
Climate Stressors on Growth, Yield, and Functional Biochemistry of two Brassica Species, Kale and Mustard
 , ,
, ,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Conditions and Plant Material
2.2. Treatments
2.3. Measurements
2.3.1. Morphological Measurements
2.3.2. Physiological Measurements
2.3.3. Epicuticular Wax Content Determination
2.3.4. Carotenoid Analysis
2.4. Data Analysis
2.4.1. Combined Stress Response Index (CSRI)
2.4.2. Statistical Analysis
3. Results
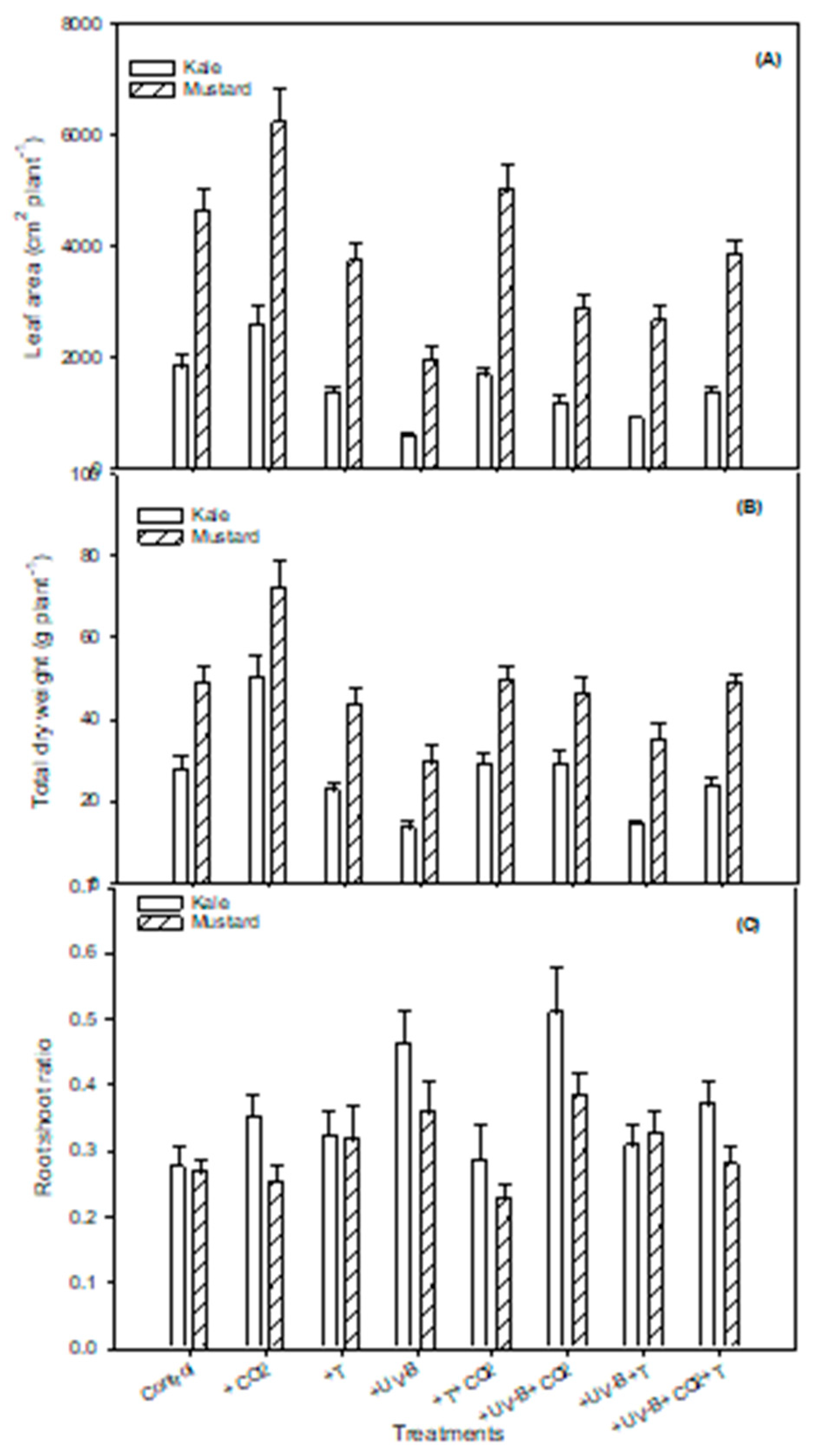
3.1. Aboveground Morphology Parameters
3.2. Dry Weight Components
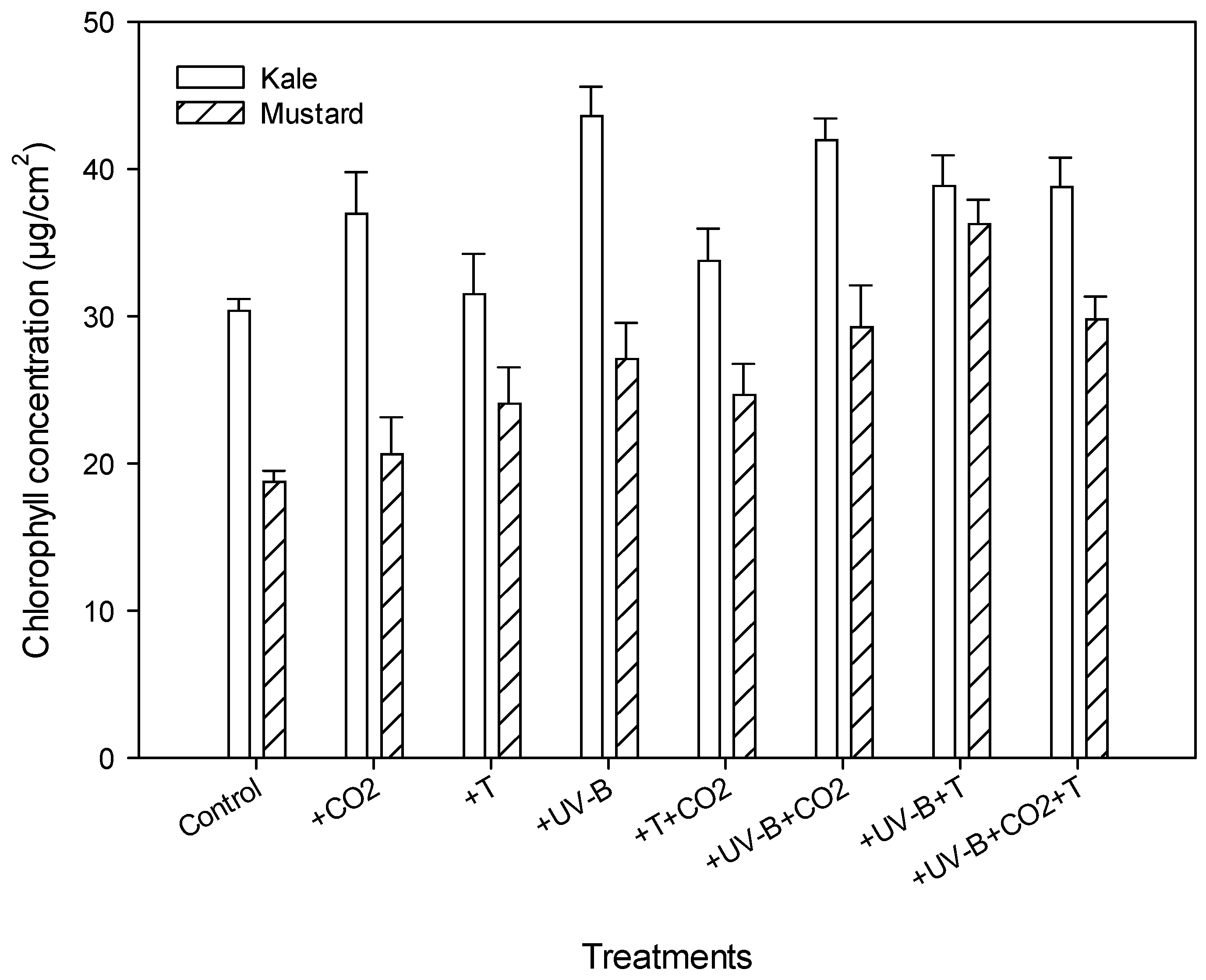
3.3. Physiological Parameters
3.4. Epicuticular Wax Content
3.5. Total Carotenoid Concentration
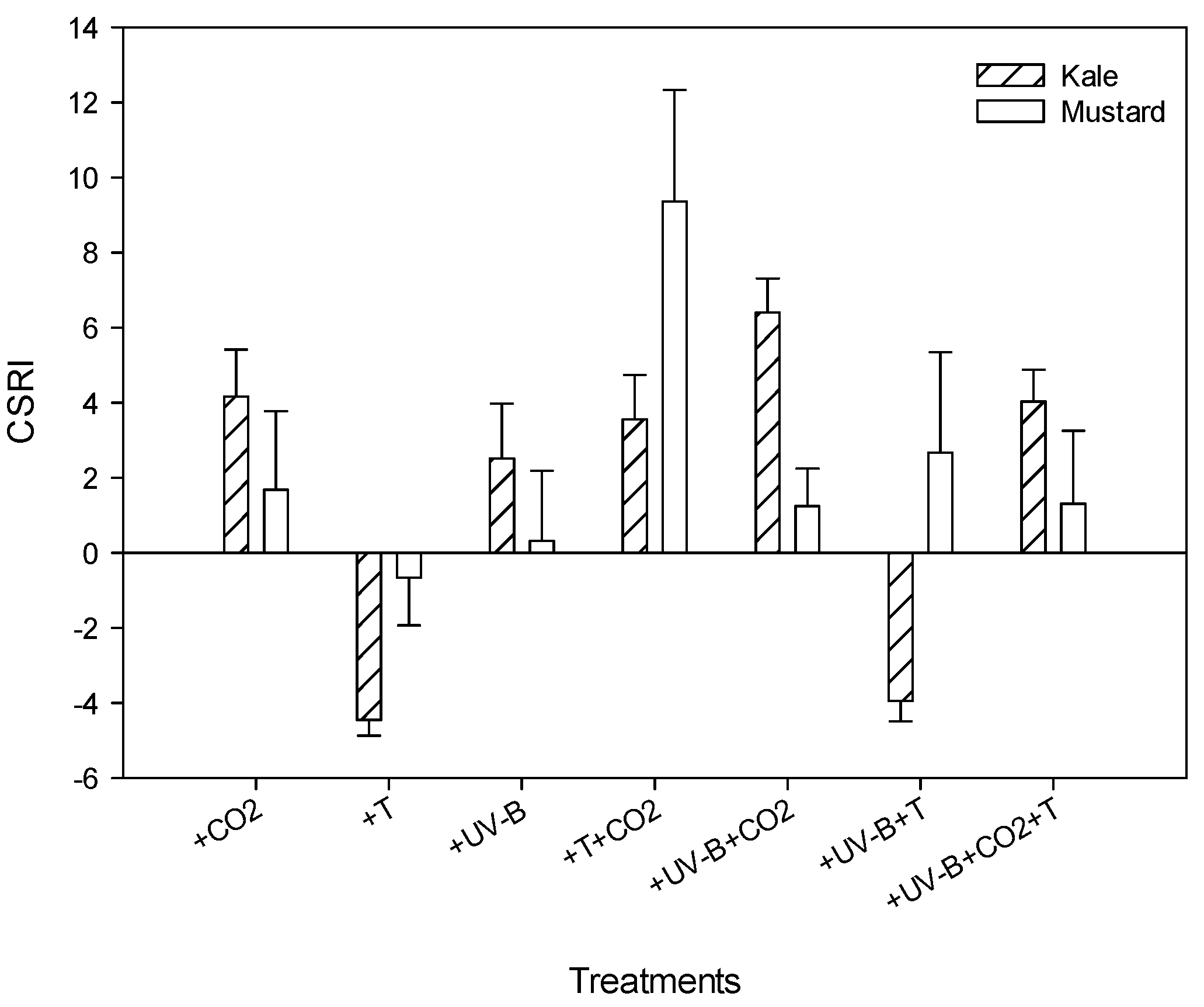
3.6. Combined Stress Response Index (CSRI)
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Meena, K.K.; Sorty, A.M.; Bitla, U.M.; Choudhary, K.; Gupta, P.; Pareek, A.; Minhas, P.S. Abiotic stress responses and microbe-mediated mitigation in plants: The omics strategies. Front. Plant Sci. 2017, 8, 172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Myers, S.S.; Smith, M.R.; Guth, S.; Golden, C.D.; Vaitla, B.; Mueller, N.D.; Huybers, P. Climate change and global food systems: Potential impacts on food security and undernutrition. Annu. Rev. Public Health 2017, 38, 259–277. [Google Scholar] [CrossRef]
- Sinha, P.; Singh, V.K.; Bohra, A.; Kumar, A.; Reif, J.C.; Varshney, R.K. Genomics and breeding innovations for enhancing genetic gain for climate resilience and nutrition traits. Theor. Appl. Genet. 2021, 134, 1829–1843. [Google Scholar] [CrossRef]
- Bilal, S.; Shahzad, R.; Imran, M.; Jan, R.; Kim, K.M.; Lee, I.J. Synergistic association of endophytic fungi enhances Glycine max L. resilience to combined abiotic stresses: Heavy metals, high temperature and drought stress. Ind. Crops Prod. 2020, 143, 111931. [Google Scholar] [CrossRef]
- Fawzy, S.; Osman, A.I.; Doran, J.; Rooney, D.W. Strategies for mitigation of climate change: A review. Environ. Chem. Lett. 2020, 18, 2069–2094. [Google Scholar] [CrossRef]
- Zandalinas, S.I.; Fritschi, F.B.; Mittler, R. Global warming, climate change, and environmental pollution: Recipe for a multifactorial stress combination disaster. Trends Plant Sci. 2021, 6, 588–599. [Google Scholar] [CrossRef]
- Intergovernmental Panel on Climate Change (IPCC). Global warming of 1.5 °C. An IPCC special report on the impacts of global warming of 1.5 °C above pre-industrial levels and related global greenhouse gas emission pathways. In The Context Of Strengthening the Global Response to the Threat of Climate Change, Sustainable Development, and Efforts to Eradicate Poverty; IPCC: Geneva, Switzerland, 2018. [Google Scholar]
- Stocker, T.F.; Qin, D.; Plattner, G.K.; Tignor, M.; Allen, S.K.; Boschung, J.; Nauels, A.; Xia, Y.; Bex, V.; Midgley, P.M. Climate Change 2013: The Physical Science Basis; Intergovernmental Panel on Climate Change, Working Group I Contribution to the IPCC Fifth Assessment Report (AR5); Cambridge University Press: New York, NY, USA, 2013. [Google Scholar]
- Intergovernmental Panel on Climate Change. Impacts, Adaptation, and Vulnerability. Part A: Global and Sectoral Aspects; Contribution of Working Group II to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; IPCC: Geneva, Switzerland, 2014. [Google Scholar]
- Obata, T.; Witt, S.; Lisec, J.; Palacios-Rojas, N.; Florez-Sarasa, I.; Yousfi, S.; Fernie, A.R. Metabolite profiles of maize leaves in drought, heat, and combined stress field trials reveal the relationship between metabolism and grain yield. Plant Physiol. 2015, 169, 2665–2683. [Google Scholar] [CrossRef] [Green Version]
- Anwar, K.; Joshi, R.; Dhankher, O.P.; Singla-Pareek, S.L.; Pareek, A. Elucidating the Response of Crop Plants towards Individual, Combined and Sequentially Occurring Abiotic Stresses. Int. J. Mol. Sci. 2021, 22, 6119. [Google Scholar] [CrossRef]
- Thavarajah, P.; Abare, A.; Basnagal, S.; Lacher, C.; Smith, P.; Combs, G.F., Jr. Mineral micronutrient and prebiotic carbohydrate profiles of USA-Grown kale (Brassica oleracea L. var. acephala). J. Food Compos. Anal. 2016, 52, 9–15. [Google Scholar] [CrossRef] [Green Version]
- Migliozzi, M.; Thavarajah, D.; Thavarajah, P.; Smith, P. Lentil and kale: Complementary nutrient-rich whole food sources to combat micronutrient and calorie malnutrition. Nutrients 2015, 7, 9285–9298. [Google Scholar] [CrossRef] [PubMed]
- Pathirana, I.; Thavarajah, P.; Siva, N.; Wickramasinghe, A.N.; Smith, P. Moisture deficit effects on kale (Brassica oleracea L. var. acephala) biomass, mineral, and low molecular weight carbohydrate concentrations. Sci. Hortic. 2017, 226, 216–222. [Google Scholar] [CrossRef]
- US Department of Health and Human Services; U.S. Department of Agriculture. 2015–2020 Dietary Guidelines for Americans, 8th ed.; 2015. Available online: http://health.gov/dietaryguidelines/2015/guidelines/ (accessed on 10 September 2022).
- Rakariyatham, N.; Sakorn, P. Biodegradation of glucosinolates in brown mustard seed meal (Brassica juncea) by Aspergillus sp. NR-4201 in liquid and solid-state cultures. Biodegradation 2002, 13, 395–399. [Google Scholar] [CrossRef] [PubMed]
- Cools, K.; Terry, L.A. The effect of processing on the glucosinolate profile in mustard seed. Food Chem. 2018, 252, 343–348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Banadyga, A.A. Greens or “Potherbs”—Chard, Collards, Kale, Mustard, Spinach; New Zealand Spinach: Washington, DC, USA, 1977; pp. 163–170. [Google Scholar]
- Deryng, D.; Conway, D.; Ramankutty, N.; Price, J.; Warren, R. Global crop yield response to extreme heat stress under multiple climate change futures. Environ. Res. Lett. 2014, 9, 034011. [Google Scholar] [CrossRef] [Green Version]
- Ugarte, C.; Calderini, D.F.; Slafer, G.A. Grain weight and grain number responsiveness to pre-anthesis temperature in wheat, barley and triticale. Field Crops Res. 2007, 100, 240–248. [Google Scholar] [CrossRef]
- Asseng, S.; Foster, I.A.N.; Turner, N.C. The impact of temperature variability on wheat yields. Glob. Change Biol. 2011, 17, 997–1012. [Google Scholar] [CrossRef]
- Allen, L.H.; Boote, K.J. Crop ecosystem responses to climatic change: Soybean. In Climate Change and Global Crop Productivity; Reddy, K.R., Hodges, H.F., Eds.; CABI Publishing: Oxon, UK, 2000; pp. 133–160. [Google Scholar] [CrossRef]
- Reddy, K.R.; Hodges, H.F. Climate Change and Global Crop Productivity; CABI Publishing: Oxon, UK, 2000. [Google Scholar]
- Ainsworth, E.A.; Davey, P.A.; Bernacchi, C.J.; Dermody, O.C.; Heaton, E.A.; Moore, D.J.; Long, S.P. A meta-analysis of elevated [CO2] effects on soybean (Glycine max) physiology, growth and yield. Glob. Change Biol. 2002, 8, 695–709. [Google Scholar] [CrossRef]
- Kakani, V.G.; Reddy, K.R.; Zhao, D.; Mohammed, A.R. Effects of ultraviolet-B radiation on cotton (Gossypium hirsutum L.) morphology and anatomy. Ann. Bot. 2003, 91, 817–826. [Google Scholar] [CrossRef]
- Kakani, V.G.; Reddy, K.R.; Zhao, D.; Sailaja, K. Field crop responses to ultraviolet-B radiation: A review. Agric. For. Meteorol. 2003, 120, 191–218. [Google Scholar] [CrossRef]
- Li, Y.; He, L.; Zu, Y. Intraspecific variation in sensitivity to ultraviolet-B radiation in endogenous hormones and photosynthetic characteristics of 10 wheat cultivars grown under field conditions. S. Afr. J. Bot. 2010, 76, 493–498. [Google Scholar] [CrossRef] [Green Version]
- Reddy, K.R.; Singh, S.K.; Koti, S.; Kakani, V.G.; Zhao, D.; Gao, W.; Reddy, V.R. Quantifying corn growth and physiological responses to ultraviolet-B radiation for modeling. Agron. J. 2013, 105, 1367–1377. [Google Scholar] [CrossRef]
- Singh, S.K.; Reddy, K.R.; Reddy, V.R.; Gao, W. Maize growth and developmental responses to temperature and ultraviolet-B radiation interaction. Photosynthetica 2014, 52, 262–271. [Google Scholar] [CrossRef]
- Kataria, S.; Jajoo, A.; Guruprasad, K.N. Impact of increasing Ultraviolet-B (UV-B) radiation on photosynthetic processes. J. Photochem. Photobiol. B Biol. 2014, 137, 55–66. [Google Scholar] [CrossRef] [PubMed]
- Allen, L.H.; Boote, K.J.; Jones, J.W.; Jones, P.H.; Pickering, N.B.; Baker, J.T.; Prasad, P.V.V. Sunlit, controlled-environment chambers are essential for comparing plant responses to various climates. Agron. J. 2020, 112, 4531–4549. [Google Scholar] [CrossRef]
- Reddy, K.R.; Read, J.J.; McKinion, J.M. Soil-Plant-Atmosphere-Research (SPAR) facility: A tool for plant research and modeling. Biotronics 2001, 30, 27–50. [Google Scholar]
- Hewitt, E.J. Sand and Water Culture Methods Used in the Study of Plant Nutrition, 2nd ed.; Commonwealth Agricultural Bureaux; Cambridge University Press: East Malling, UK, 1952. [Google Scholar]
- Ebercon, A.; Blum, A.; Jordan, W.R. A Rapid Colorimetric Method for Epicuticular Wax Contest of Sorghum Leaves. Crop Sci. 1977, 17, 179–180. [Google Scholar] [CrossRef] [Green Version]
- Kopsell, D.A.; Kopsell, D.E.; Curran-Celentano, J. Carotenoid pigments in kale are influenced by nitrogen concentration and form. J. Sci. Food Agric. 2007, 87, 900–907. [Google Scholar] [CrossRef]
- Kopsell, D.A.; Kopsell, D.E.; Lefsrud, M.G.; Curran-Celentano, J.; Dukach, L.E. Variation in Lutein, β-carotene, and Chlorophyll Concentrations among Brassica oleracea Cultigens and Seasons. HortScience 2004, 39, 361–364. [Google Scholar] [CrossRef]
- Barickman, T.C.; Kopsell, D.A.; Sams, C.E. Abscisic Acid Impacts Tomato Carotenoids, Soluble Sugars, and Organic Acids. HortScience 2016, 51, 370–376. [Google Scholar] [CrossRef] [Green Version]
- Koti, S.; Reddy, K.R.; Reddy, V.R.; Kakani, V.G.; Zhao, D. Interactive effects of carbon dioxide, temperature, and ultraviolet-B radiation on soybean (Glycine max L.) flower and pollen morphology, pollen production, germination, and tube lengths. J. Exp. Bot. 2005, 56, 725–736. [Google Scholar] [CrossRef]
- Qaderi, M.M.; Reid, D.M. Growth and physiological responses of canola (Brassica napus) to UV-B and CO2 under controlled environment conditions. Physiol. Plant. 2005, 125, 247–259. [Google Scholar] [CrossRef]
- Rodríguez-Calzada, T.; Qian, M.; Strid, Å.; Neugart, S.; Schreiner, M.; Torres-Pacheco, I.; Guevara-González, R.G. Effect of UV-B radiation on morphology, phenolic compound production, gene expression, and subsequent drought stress responses in chili pepper (Capsicum annuum L.). Plant Physiol. Biochem. 2019, 134, 94–102. [Google Scholar] [CrossRef] [PubMed]
- Son, K.H.; Ide, M.; Goto, E. Growth characteristics and phytochemicals of canola (Brassica napus) grown under U.V. radiation and low root zone temperature in a controlled environment. Hortic. Environ. Biotechnol. 2020, 61, 267–277. [Google Scholar] [CrossRef]
- Lercari, B.; Sodi, F.; Di Paola, M.L. Photomorphogenic responses to U.V. radiation: Involvement of phytochrome and U.V. photoreceptors in the control of hypocotyl elongation in Lycopersicon esculentum. Physiol. Plant. 1990, 79, 668–672. [Google Scholar] [CrossRef]
- Corlett, J.E.; Stephen, J.; Jones, H.G.; Woodfin, R.; Mepsted, R.; Paul, N.D. Assessing the impact of UV-B radiation on the growth and yield of field crops. In Plants and UV-B: Responses to Environmental Change; Lumsden, P.J., Ed.; Cambridge University Press: Cambridge, MA, USA, 1997; pp. 195–212. [Google Scholar]
- Conner, J.K.; Zangori, L.A. A garden study of the effects of ultraviolet-B radiation on pollination success and lifetime female fitness in Brassica. Oecologia 1997, 111, 388–395. [Google Scholar] [CrossRef]
- Prasad, P.V.V.; Staggenborg, S.A.; Ristic, Z. Impacts of drought and/or heat stress on physiological, developmental, growth, and yield processes of crop plants. In Response of Crops to Limited Water: Understanding and Modeling Water Stress Effects on Plant Growth Processes; Ahuja, L.H., Saseendran, S.A., Eds.; ASA-CSSA: Madison, WI, USA, 2008; pp. 301–355. [Google Scholar]
- Rodríguez, M.; Canales, E.; Borrás-Hidalgo, O. Molecular aspects of abiotic stress in plants. Biotecnol. Apl. 2005, 22, 1–10. [Google Scholar]
- Hasanuzzaman, M.; Nahar, K.; Alam, M.; Roychowdhury, R.; Fujita, M. Physiological, biochemical, and molecular mechanisms of heat stress tolerance in plants. Int. J. Mol. Sci. 2013, 14, 9643–9684. [Google Scholar] [CrossRef]
- Chauhan, J.S.; Meena, M.L.; Saini, M.K.; Meena, D.R.; Singh, M.; Meena, S.S.; Singh, K.H. Heat stress effects on morph-physiological characters of Indian Mustard (Brassica juncea L.). In Proceedings of the 16th Australian Research Assembly on Brassicas, Ballarat, Victoria, Australia, 14–16 September 2009; pp. 91–97. [Google Scholar]
- Schaarschmidt, S.; Lawas, L.M.F.; Glaubitz, U.; Li, X.; Erban, A.; Kopka, J.; Zuther, E. Season affects yield and metabolic profiles of rice (Oryza sativa) under high night temperature stress in the field. Int. J. Mol. Sci. 2020, 21, 3187. [Google Scholar] [CrossRef]
- Kosova, K.; Vitamvas, P.; Prasil, I.T.; Renaut, J. Plant proteome changes under abiotic stress—Contribution of proteomics studies to understanding plant stress response. J. Proteom. 2011, 74, 1301–1322. [Google Scholar] [CrossRef]
- Akter, N.; Islam, M.R. Heat stress effects and management in wheat. A review. Agron. Sustain. Dev. 2017, 37, 37. [Google Scholar] [CrossRef]
- Qaderi, M.M.; Kurepin, L.V.; Reid, D.M. Growth and physiological responses of canola (Brassica napus) to three components of global climate change: Temperature, carbon dioxide and drought. Physiol. Plant. 2006, 128, 710–721. [Google Scholar] [CrossRef]
- Qaderi, M.M.; Basraon, N.K.; Chinnappa, C.C.; Reid, D.M. Combined effects of temperature, ultraviolet-B radiation, and watering regime on growth and physiological processes in canola (Brassica napus) seedlings. Int. J. Plant Sci. 2010, 171, 466–481. [Google Scholar] [CrossRef]
- Pérez, M.L.; Prado, F.E.; González, J.A. Effects of ultraviolet B (UV-B) on different varieties of quinoa. I. Effects on morphology under controlled conditions. Bol. Soc. Argent. Bot. 2015, 50, 337–347. [Google Scholar] [CrossRef]
- Hinojosa, L.; González, J.A.; Barrios-Masias, F.H.; Fuentes, F.; Murphy, K.M. Quinoa abiotic stress responses: A review. Plants 2018, 7, 106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raghuvanshi, R.; Sharma, R.K. Response of two cultivars of Phaseolus vulgaris L. (French beans) plants exposed to enhanced UV-B radiation under mountain ecosystem. Environ. Sci. Pollut. Res. 2016, 23, 831–842. [Google Scholar] [CrossRef] [PubMed]
- Wargent, J.J.; Gegas, V.C.; Jenkins, G.I.; Doonan, J.H.; Paul, N.D. UVR8 in Arabidopsis thaliana regulates multiple aspects of cellular differentiation during leaf development in response to ultraviolet B radiation. New Phytol. 2009, 183, 315–326. [Google Scholar] [CrossRef] [PubMed]
- Nogués, S.; Allen, D.J.; Morison, J.I.; Baker, N.R. Characterization of stomatal closure caused by ultraviolet-B radiation. Plant Physiol. 1999, 121, 489–496. [Google Scholar] [CrossRef] [Green Version]
- Nedunchezhian, N.; Kulandaivelu, G. Changes induced by ultraviolet-B (280–320 nm) to vegetative growth and photosynthetic characteristics in filed grown Vigna unguiculate L. Plant Sci. 1997, 123, 85–92. [Google Scholar] [CrossRef]
- Cechin, I.; Fumis, T.D.F.; Dokkedal, A.L. Growth and physiological responses of sunflower plants exposed to ultraviolet-B radiation. Ciênc. Rural 2007, 37, 85–90. [Google Scholar] [CrossRef]
- Gao, W.; Zheng, Y.F.; Slusser, J.R.; Heisler, G.M.; Grant, R.H.; Xu, J.Q.; He, D.L. Effects of supplementary ultraviolet-B irradiance on maize yield and qualities: A field experiment. J. Photochem. Photobiol. 2004, 80, 127–131. [Google Scholar] [CrossRef]
- Rajabbeigi, E.; Eichholz, I.; Beesk, N.; Ulrichs, C.; Kroh, L.W.; Rohn, S.; Huyskens-Keil, S. Interaction of drought stress and UV-B radiation-impact on biomass production and flavonoid metabolism in lettuce (Lactuca sativa L.). J. Appl. Bot. Food Qual. 2013, 86, 190–197. [Google Scholar] [CrossRef]
- Balasooriya, H.N.; Dassanayake, K.B.; Seneweera, S.; Ajlouni, S. Interaction of elevated carbon dioxide and temperature on strawberry (Fragaria× ananassa) growth and fruit yield. Int. J. Agric. Biosyst. Eng. 2018, 12, 279–287. [Google Scholar]
- Poulson, M.E.; Boeger, M.R.T.; Donahue, R.A. Response of photosynthesis to high light and drought for Arabidopsis thaliana grown under a UV-B enhanced light regime. Photosynth. Res. 2006, 90, 79–90. [Google Scholar] [CrossRef] [PubMed]
- Koti, S.; Reddy, K.R.; Kakani, V.G.; Zhao, D.; Gao, W. Effects of carbon dioxide, temperature and ultraviolet-B radiation and their interactions on soybean (Glycine max L.) growth and development. Environ. Exp. Bot. 2007, 60, 1–10. [Google Scholar] [CrossRef]
- Wijewardana, C.; Henry, W.B.; Gao, W.; Reddy, K.R. Interactive effects on CO2, drought, and ultraviolet-B radiation on maize growth and development. J. Photochem. Photobiol. 2016, 160, 198–209. [Google Scholar] [CrossRef] [PubMed]
- Wanzeler, R.B.; Zanetti, L.V.; Fantinato, D.E.; Gama, V.N.; Arrivabene, H.P.; de Almeida Leite, I.T.; Milanez, C.R.D. How does UV-B radiation affect the initial growth of common bean (Phaseolus vulgaris L.)? Physiological and structural aspects. Braz. J. Dev. 2019, 5, 26947–26958. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.; Gao, W.; Reddy, K.R.; Chen, M.; Taduri, S.; Meyers, S.L.; Shankle, M.W. Ultraviolet (U.V.) B effects on growth and yield of three contrasting sweet potato cultivars. Photosynthetica 2020, 58, 37–44. [Google Scholar] [CrossRef] [Green Version]
- Barickman, T.C.; Olorunwa, O.J.; Sehgal, A.; Walne, C.H.; Reddy, K.R.; Gao, W. Interactive impacts of temperature and elevated CO2 on Basil (Ocimum basilicum L.) root and shoot morphology and growth. Horticulturae 2021, 7, 112. [Google Scholar] [CrossRef]
- Sakalauskaite, J.; Viškelis, P.; Duchovskis, P.; Dambrauskiene, E.; Sakalauskiene, S.; Samuoliene, G.; Brazaityte, A. Supplementary UV-B irradiation effects on basil (Ocimum basilicum L.) growth and phytochemical properties. J. Food Agric. Environ. 2012, 10, 342–346. [Google Scholar]
- Wang, W.; Cai, C.; He, J.; Gu, J.; Zhu, G.; Zhang, W.; Liu, G. Yield, dry matter distribution and photosynthetic characteristics of rice under elevated CO2 and increased temperature conditions. Field Crops Res. 2020, 248, 107605. [Google Scholar] [CrossRef]
- Yuan, L.; Tang, L.; Zhu, S.; Hou, J.; Chen, G.; Liu, F.; Wang, C. Influence of heat stress on leaf morphology and nitro gen–carbohydrate metabolisms in two wucai (Brassica campestris L.) genotypes. Acta Soc. Bot. Pol. 2017, 86, 1–16. [Google Scholar] [CrossRef]
- Boeger, M.R.T.; Poulson, M. Effects of ultraviolet-B radiation on leaf morphology of Arabidopsis thaliana (L.) Heynh. (Brassicaceae). Acta Bot. Bras. 2006, 20, 329–338. [Google Scholar] [CrossRef]
- Karvansara, P.R.; Razavi, S.M. Physiological and biochemical responses of sugar beet (Beta vulgaris L.) to ultraviolet-B radiation. PeerJ 2019, 7, e6790. [Google Scholar] [CrossRef]
- Al-Oudat, M.; Baydoun, S.A.; Mohammad, A. Effects of enhanced UV-B on growth and yield of two Syrian crops wheat (Triticum durum var. Horani) and broad beans (Vicia faba) under field conditions. Environ. Exp. Bot. 1998, 40, 11–16. [Google Scholar] [CrossRef]
- Zhang, X.R.; Chen, Y.H.; Guo, Q.S.; Wang, W.M.; Liu, L.; Fan, J.; Li, C. Short-term UV-B radiation effects on morphology, physiological traits and accumulation of bioactive compounds in Prunella vulgaris L. J. Plant Interact. 2017, 12, 348–354. [Google Scholar] [CrossRef]
- Hideg, É.; Jansen, M.A.; Strid, Å. UV-B exposure, ROS, and stress: Inseparable companions or loosely linked associates? Trends Plant Sci. 2013, 18, 107–115. [Google Scholar] [CrossRef] [Green Version]
- Ziska, L.H.; Teramura, A.H.; Sullivan, J.H.; McCoy, A. Influence of ultraviolet-B (UV-B) radiation on photosynthetic and growth characteristics in field-grown cassava (Manihot esculentum Crantz). Plant Cell Environ. 1993, 16, 73–79. [Google Scholar] [CrossRef]
- Idso, S.B.; Kimball, B.A.; Mauney, J.R. Effects of atmospheric CO2 enrichment on root: Shoot ratios of carrot, radish, cotton and soybean. Agric. Ecosyst. Environ. 1988, 21, 293–299. [Google Scholar] [CrossRef]
- Yang, X. Plants and Microclimate: A Quantitative Approach to Environmental Plant Physiology. Agric. Forest Meteorol. 1993, 66, 267–268. [Google Scholar] [CrossRef]
- Shah, N.H.; Paulsen, G.M. Interaction of drought and high temperature on photosynthesis and grain-filling of wheat. Plant Soil 2003, 257, 219–226. [Google Scholar] [CrossRef]
- Angadi, S.V.; Cutforth, H.W.; Miller, P.R.; McConkey, B.G.; Entz, M.H.; Brandt, S.A.; Volkmar, K.M. Response of three Brassica species to high temperature stress during reproductive growth. Can. J. Plant Sci. 2000, 80, 693–701. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez, V.M.; Soengas, P.; Alonso-Villaverde, V.; Sotelo, T.; Cartea, M.E.; Velasco, P. Effect of temperature stress on the early vegetative development of Brassica oleracea L. BMC Plant Biol. 2015, 15, 145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, E.Y.; Seo, T.C.; Lee, S.G.; Cho, I.H.; Stangoulis, J. Growth and physiological responses of Chinese cabbage and radish to long-term exposure to elevated carbon dioxide and temperature. Hortic. Environ. Biotechnol. 2011, 52, 376. [Google Scholar] [CrossRef]
- Hinojosa, L.; Matanguihan, J.B.; Murphy, K.M. Effect of high temperature on pollen morphology, plant growth and seed yield in quinoa (Chenopodium quinoa Willd.). J. Agron. Crop Sci. 2019, 205, 33–45. [Google Scholar] [CrossRef] [Green Version]
- Sun, P.; Mantri, N.; Lou, H.; Hu, Y.; Sun, D.; Zhu, Y.; Lu, H. Effects of elevated CO2 and temperature on yield and fruit quality of strawberry (Fragaria× ananassa Duch.) at two levels of nitrogen application. PLoS ONE 2012, 7, e41000. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumari, M.; Verma, S.C.; Bhardwaj, S.K. Effect of elevated CO2 and temperature on crop growth and yield attributes of bell pepper (Capsicum annuum L.). J. Agrometeorol. 2019, 21, 1–6. [Google Scholar] [CrossRef]
- Rangaswamy, T.C.; Sridhara, S.; Ramesh, N.; Gopakkali, P.; El-Ansary, D.O.; Mahmoud, E.A.; Abdel-Hamid, A.M. Assessing the impact of higher levels of CO2 and temperature and their interactions on tomato (Solanum lycopersicum L.). Plants 2021, 10, 256. [Google Scholar] [CrossRef] [PubMed]
- Zhou, R.; Yu, X.; Li, X.; Mendanha dos Santos, T.; Rosenqvist, E.; Ottosen, C.O. Combined high light and heat stress induced complex response in tomato with better leaf cooling after heat priming. Plant Physiol. Biochem. 2020, 151, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Barickman, T.C.; Olorunwa, O.J.; Sehgal, A.; Walne, C.H.; Reddy, K.R.; Gao, W. Yield, Physiological Performance, and Phytochemistry of Basil (Ocimum basilicum L.) under Temperature Stress and Elevated CO2 Concentrations. Plants 2021, 10, 1072. [Google Scholar] [CrossRef]
- Yoon, H.I.; Kim, D.; Son, J.E. Spatial and temporal bioactive compound contents and chlorophyll fluorescence of kale (Brassica oleracea L.) under UV-B exposure near harvest time in controlled environments. Photochem. Photobiol. 2020, 96, 845–852. [Google Scholar] [CrossRef]
- Pourcel, L.; Routaboul, J.M.; Cheynier, V.; Lepiniec, L.; Debeaujon, I. Flavonoid oxidation in plants: From biochemical properties to physiological functions. Trends Plant Sci. 2007, 12, 29–36. [Google Scholar] [CrossRef] [PubMed]
- Olsson, L.C.; Veit, M.; Bornman, J.F. Epidermal transmittance and phenolic composition in leaves of atrazine-tolerant and atrazine-sensitive cultivars of Brassica napus grown under enhanced UV-B radiation. Physiol. Plant. 1999, 107, 259–266. [Google Scholar] [CrossRef]
- Cartelat, A.; Cerovic, Z.G.; Goulas, Y.; Meyer, S.; Lelarge, C.; Prioul, J.L.; Barbottin, A.; Jeuffroy, M.H.; Gate, P.; Agati, G.; et al. Optically assessed contents of leaf polyphenolics and chlorophyll as indicators of nitrogen deficiency in wheat (Triticum aestivum L.). Field Crops Res. 2005, 91, 35–49. [Google Scholar] [CrossRef]
- Oliveira, A.F.; Rais, F.; Dettori, I.; Azzena, M.; Nieddu, G. UV light acclimation capacity of leaf photosynthetic and photochemical behaviour in near-isohydric and anisohydric grapevines in hot and dry environments. S. Afr. J. Enol. Vitic. 2019, 40, 188–205. [Google Scholar] [CrossRef] [Green Version]
- Pieristè, M.; Chauvat, M.; Kotilainen, T.K.; Jones, A.G.; Aubert, M.; Robson, M.; Forey, E. Solar UV-A radiation and blue light enhance tree leaf litter decomposition in a temperate forest. Oecologia 2019, 191, 191–203. [Google Scholar] [CrossRef] [Green Version]
- Martel, A.B.; Taylor, A.E.; Qaderi, M.M. Individual and interactive effects of temperature and light intensity on canola growth, physiological characteristics and methane emissions. Plant Physiol. Biochem. 2020, 157, 160–168. [Google Scholar] [CrossRef]
- Steinmüller, D.; Tevini, M. Action of ultraviolet radiation (UV-B) upon cuticular waxes in some crop plants. Planta 1985, 164, 557–564. [Google Scholar] [CrossRef]
- Gonzalez, R.; Paul, N.D.; Percy, K.; Ambrose, M.; McLaughlin, C.K.; Barnes, J.D.; Wellburn, A.R. Responses to ultraviolet-B radiation (280–315 nm) of pea (Pisum sativum) lines differing in leaf surface wax. Physiol. Plant. 1996, 98, 852–860. [Google Scholar] [CrossRef]
- Dong, J.; Gruda, N.; Lam, S.K.; Li, X.; Duan, Z. Effects of elevated CO2 on nutritional quality of vegetables: A review. Front. Plant Sci. 2018, 9, 924. [Google Scholar] [CrossRef]
- Gruda, N.; Bisbis, M.; Tanny, J. Impacts of protected vegetable cultivation on climate change and adaptation strategies for cleaner production–a review. J. Clean. Prod. 2019, 225, 324–339. [Google Scholar] [CrossRef]
- Slavin, J.L.; Lloyd, B. Health benefits of fruits and vegetables. Adv. Nutr. 2012, 3, 506–516. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
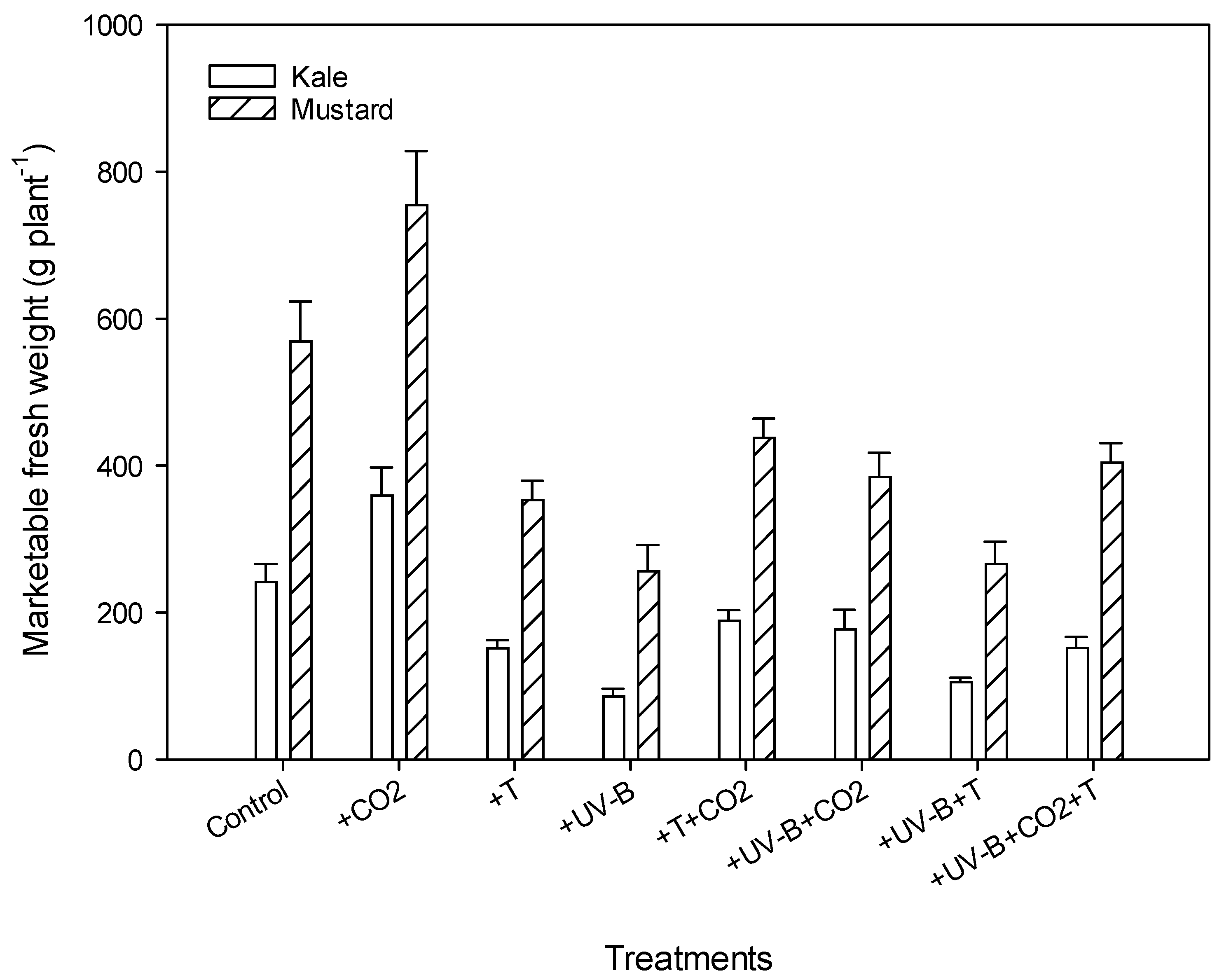{kind=link}
{kind=link}
{kind=link}
| Treatments | Measured Temperature (°C) | CO2 (µmol mol−1) | VPD (kPa) | Mean ET (L H2O d−1) | |||
|---|---|---|---|---|---|---|---|
| DAY | NIGHT | DAY/NIGHT | DAY | DAY | NIGHT | DAY/NIGHT | |
| Control | 24.90 ± 0.08 | 17.42 ± 0.03 | 21.56 ± 0.04 | 433 ± 1.92 | 1.35 ± 0.02 | 0.94 ± 0.01 | 11.55 ± 0.84 |
| +CO2 | 25.13 ± 0.06 | 17.54 ± 0.02 | 21.73 ± 0.04 | 721.22 ± 1.27 | 1.33 ± 0.02 | 0.97 ± 0.01 | 11.71 ± 1.15 |
| +T | 31.31 ± 0.66 | 23.73 ± 0.65 | 27.93 ± 0.64 | 434.31 ± 1.30 | 2.30 ± 0.11 | 1.68 ± 0.08 | 12.56 ± 1.13 |
| +UV-B | 24.84 ±0.10 | 17.31 ± 0.03 | 21.48 ± 0.05 | 439.69 ± 1.72 | 1.37 ± 0.01 | 0.97 ± 0.01 | 8.48 ± 0.57 |
| +T + CO2 | 31.69 ± 0.70 | 24.04 ± 0.69 | 28.28 ± 0.68 | 720.04 ± 2.49 | 2.75± 0.11 | 1.91 ± 0.08 | 13.79 ± 1.40 |
| +UV-B + CO2 | 24.90 ± 0.09 | 17.35 ± 0.03 | 21.53 ± 0.05 | 715.98 ± 2.17 | 1.32 ± 0.02 | 0.90 ± 0.01 | 8.71 ± 1.01 |
| +UV-B + T | 31.32 ± 0.67 | 23.75 ± 0.66 | 27.94 ± 0.65 | 435.33 ± 1.27 | 2.74 ± 0.11 | 1.90 ± 0.09 | 11.68 ± 0.96 |
| +UV-B + CO2 + T | 31.34 ± 0.67 | 23.75 ± 0.66 | 27.96 ± 0.65 | 729.37 ±1.46 | 2.89 ± 0.14 | 1.99 ± 0.10 | 11.34 ± 1.21 |
| Source of Variance | PH | LN | LA | MFW | ADW | RDW | TDW | RS | Neo | Viol | Zea | Lut | Bcar | TXan | ZA/ZAV | Chl | Flav | Anth | NBI | Wax |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Treatment | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | N.S. | ** | ** | *** | ** | *** | *** | *** | *** | *** |
| Crop | *** | *** | *** | *** | *** | *** | *** | ** | * | ** | ** | *** | N.S. | N.S. | ** | *** | N.S. | *** | *** | *** |
| Trt *Crop | *** | *** | *** | ** | N.S. | N.S. | N.S. | N.S. | N.S. | N.S. | N.S. | N.S. | N.S. | N.S. | N.S. | ** | ** | *** | * | *** |
| Traits | Crop | Treatments | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Shoot Traits | Control | +CO2 | +T | +UV-B | +T + CO2 | +UV-B + CO2 | +UV-B + T | +UV-B + CO2 + T | ||
| PH (cm) | Kale | 53.8 | 57.7 (+7%) | 37.7 (−29.8%) | 28.3 (−47.3%) | 35.7 (−33.6%) | 37.1 (−31%) | 31.1 (−42%) | 35.1 (−34.7%) | |
| Mustard | 52.4 | 56.1 (+7%) | 38.7 (−26%) | 40.3 (−23%) | 36.3 (−30.7%) | 44 (−16%) | 39.6 (−24.5%) | 42.4 (−19%) | ||
| LN (plant−1) | Kale | 14.2 | 14.7 (+3%) | 13 (−8.5%) | 14.3 (0%) | 13.8 (−3%) | 15.3 (+7.8%) | 13.4 (−5.4%) | 14 (−1.5%) | |
| Mustard | 32 | 43 (+34%) | 36.2 (+13%) | 20.6 (−35.6%) | 50.1 (+50.6%) | 28.6 (−10.7%) | 30.7 (−4%) | 36.7 (+14.8%) | ||
| LA (cm2 plant−1) | Kale | 1805.7 | 2558.7 (+39%) | 1357.3 (−24.8%) | 570.1 (−68.4%) | 1843.6 (+2%) | 1136.9 (−37%) | 882.8 (−51%) | 1153.1 (−36%) | |
| Mustard | 4613.7 | 6220.2 (+34.8%) | 3724.6 (−19%) | 1934.7 (−58%) | 4993.5 (+8%) | 2857.1 (−38%) | 2637.4 (−42.8%) | 3830.9 (−17%) | ||
| MFW (g plant1) | Kale | 241.3 | 359.2 (+48.8%) | 151 (−37.4%) | 85.9 (−64.3%) | 188.8 (−21.7%) | 176.8 (−26.7%) | 105.3 (−56.3%) | 151.7 (37%) | |
| Mustard | 569.2 | 755 (+32.6) | 353 (−40%) | 256 (−55%) | 437.8 (−23%) | 384.6 (−32.4%) | 266.2 (−53%) | 404.2 (−30%) | ||
| Dry weight traits | RS | Kale | 0.27 | 0.35 (+29.6%) | 0.32 (+18.5%) | 0.46 (+70%) | 0.28 (+3.7%) | 0.51 (+88.8%) | 0.31 (+14.8%) | 0.37 (+37%) |
| Mustard | 0.27 | 0.35 (+29.6%) | 0.32 (+18.5%) | 0.36 (+33.3%) | 0.23 (−14.8%) | 0.38 (+40.7%) | 0.33 (+22.2%) | 0.28 (+3.7%) | ||
| ADW (g plant−1) | Kale | 21.7 | 37.4 (+72.1%) | 17 (−21.7%) | 8.9 (−59%) | 22.4 (+3.2%) | 19.4 (−10.8%) | 10.9 (−49.4%) | 17.3 (−20%) | |
| Mustard | 38.6 | 57.9 (+50%) | 33.2 (−14%) | 21.7 (−43.6%) | 40.4 (+4.7%) | 33.4 (−13.3%) | 26.3 (−31.8%) | 38 (−1.5%) | ||
| RDW (g plant−1) | Kale | 6 | 11.7 (+95%) | 5.5 (−8.5%) | 4.3 (−28%) | 6.4 (+6.5%) | 9.6 (+60.5%) | 3.3 (−45.3%) | 6.3 (+5.6%) | |
| Mustard | 10 | 13.9 (+39%) | 10.4 (+3.9%) | 7.6 (−24%) | 8.9 (−10.3%) | 12.6 (+25.6%) | 8.7 (−12.8%) | 10.4 (+4.4%) | ||
| TDW (g plant−1) | Kale | 27.7 | 49 (+76.8%) | 22.5 (−19%) | 13.2 (−52.3%) | 28.8 (+4%) | 29 (+4.6%) | 14.2 (−48.5%) | 23.7 (−14.5%) | |
| Mustard | 48.6 | 71.8 (+47.8%) | 43.6 (−10%) | 29.3 (−39.6%) | 49.4 (+1.6%) | 45.9 (−5.3%) | 35 (−28%) | 48.5 (0%) | ||
| Traits | Crop | Treatments | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Control | +CO2 | +T | +UV-B | +T + CO2 | +UV-B + CO2 | +UV-B + T | +UV-B + CO2 + T | ||
| Chlorophyll conc. (µg/cm2) | Kale | 30.4 | 37 (+21.7%) | 31.5 (+3.6%) | 43.6 (+43.4%) | 33.8 (+11%) | 42 (+38%) | 38.8 (+27.6%) | 38.8 (+27.6%) |
| Mustard | 18.77 | 20.6 (+9.7%) | 24 (+27.8%) | 27.1 (+44.3%) | 24.7 (+24.2%) | 29.3 (+56%) | 36.3 (+93.3%) | 29.8 (+58.7%) | |
| Flavonoid index | Kale | 0.72 | 0.81 (+12.5%) | 0.74 (+2.7%) | 1.21 (+68%) | 0.81 (+12.5%) | 1.26 (+75%) | 1.25 (+73.6%) | 1.32 (+83.3%) |
| Mustard | 0.82 | 0.85 (+3.6%) | 0.83 (+1.2%) | 1 (+30%) | 0.84 (+2.4%) | 1.12 (+36.5%) | 1.19 (+45%) | 1.40 (+70.7%) | |
| Anthocyanin index | Kale | 0.09 | 0.09 (0%) | 0.08 (−11%) | 0.06 (−33.3%) | 0.09 (0%) | 0.06 (−33.3%) | 0.07 (−22.2%) | 0.06 (−33.3%) |
| Mustard | 0.14 | 0.13 (−7%) | 0.12 (−14.2%) | 0.12 (−14.2%) | 0.12 (−14.2%) | 0.11 (−21.4%) | 0.09 (−35.7%) | 0.10 (−28.5%) | |
| NBI | Kale | 42.8 | 46.4 (+8.4%) | 46.2 (+8%) | 37.7 (−11.9%) | 43 (+0.5%) | 34.4 (−19.6%) | 31.5 (−26.4%) | 30 (−30%) |
| Mustard | 23.4 | 24.6 (+5%) | 31.4 (+34%) | 28.5 (+21.7%) | 36.5 (+60%) | 27.4 (+17%) | 31.8 (+35.8%) | 21.9 (−6.4%) | |
| Waxes (µg/cm2) | Kale | 122.5 | 115.6 (−5.6%) | 133 (+8.5%) | 92.5 (−24.4%) | 140.82 (+15%) | 87.9 (−28.2%) | 66.3 (−45.8%) | 70.1 (−42.7%) |
| Mustard | 17.4 | 13 (−25.2%) | 14.7 (−15.5%) | 13.4 (−30%) | 18.18 (+4.5%) | 11 (−36.7%) | 17.5 (0%) | 21 (+20.6%) | |
| Traits | Crop | Treatments | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Control | +CO2 | +T | +UV-B | +T + CO2 | +UV-B + CO2 | +UV-B + T | +UV-B + CO2 + T | ||
| Neo (µg/g dry mass) | Kale | 290 | 355 (+22.4%) | 276.8 (−4.5%) | 407.7 (+40.5%) | 298.5 (+3%) | 372.1 (+28.3%) | 371.5 (+28.1%) | 397 (+36.8%) |
| Mustard | 273.3 | 256.3 (−6.2%) | 246.3 (−9.8%) | 370.4 (+35.5%) | 269 (−1.6%) | 361.2 (+32%) | 378.5 (+38.4%) | 319.3 (+16.8%) | |
| Viol (µg/g dry mass) | Kale | 163.7 | 195 (+19%) | 167.5 (−2.3%) | 314.4 (+92%) | 150 (−8.3%) | 337.8 (+106%) | 276.2 (+68.8%) | 242 (+47.8%) |
| Mustard | 323.9 | 217.3 (−40%) | 248.8 (−23%) | 327.7 (+1%) | 208 (−35.7%) | 274.8 (−15%) | 391.2 (+20.7%) | 264 (−18.4%) | |
| Anth (µg/g dry mass | Kale | 37.6 | 39.3 (+4.5%) | 23.8 (−36.7%) | 46.6 (+23.9%) | 34.6 (−8%) | 41.6 (+10.6%) | 43.6 (+16%) | 31.2 (−17%) |
| Mustard | 64.1 | 48.2 (−24.8%) | 44.6 (−30.4%) | 67.7 (+5.6%) | 38.3 (−40%) | 51.8 (−19%) | 45.4 (−29%) | 37.7 (−41%) | |
| Zea (µg/g dry mass) | Kale | 209.5 | 219.2 (+4.6%) | 132.9 (−36.5%) | 149.4 (−28.6%) | 139.2 (−33.5%) | 157.4 (−24.8%) | 140.8 (−32.7%) | 147.5 (−29.5%) |
| Mustard | 129.6 | 126.2 (−2.6%) | 99.1 (−23.5%) | 148.7 (+14.7%) | 146.8 (+13.2%) | 138.5 (+6.8%) | 123.9 (−4.3%) | 139.2 (+7.4%) | |
| Lut (µg/g dry mass) | Kale | 898.3 | 967.3 (+7.6%) | 825 (−8%) | 1093.6 (+21.7%) | 679.2 (24.3%) | 1105.5 (+23%) | 1066.9 (+18.7%) | 948.1 (−5.5%) |
| Mustard | 746.7 | 635.7 (−14.8%) | 676.4 (−9.4%) | 766.7 (+2.6%) | 654.8 (−12.3%) | 742.3 (−0.5%) | 899.6 (+20.4%) | 695 (−6.9%) | |
| Bcar (µg/g dry mass) | Kale | 548.4 | 656.7 (+19.7%) | 580.5 (+5.8%) | 477.6 (−13%) | 439.8 (−19.8%) | 710.7 (+29.5%) | 766.6 (+39.7%) | 636.8 (+16%) |
| Mustard | 649 | 521.1 (−19.7%) | 588.2 (−9.3%) | 620.9 (−4.3%) | 476.6 (−26.5%) | 517.7 (−20%) | 745.2 (+14.8%) | 568.3 (−12.4%) | |
| Total Xanth (µg/g dry mass) | Kale | 410.8 | 453.5 (+10.3%) | 324.1 (−21%) | 510.3 (+24.2%) | 323.7 (−21.2%) | 536.7 (+30.6%) | 460.6 (+12%) | 420.7 (+2.4%) |
| Mustard | 517.6 | 391.8 (−24.3%) | 392.5 (−24.1%) | 544 (+5.2%) | 393.3 (−24%) | 465 (−10%) | 560.5 (+8.2%) | 440.9 (−14.8%) | |
| ZA/ZAV | Kale | 0.60 | 0.57 (−5%) | 0.48 (−20%) | 0.38 (−36.6%) | 0.53 (−11.6%) | 0.37 (−38.3%) | 0.40 (−33.3%) | 0.44 (−26.6%) |
| Mustard | 0.37 | 0.44 (+18.9%) | 0.36 (−2.7%) | 0.41 (+10.8%) | 0.47 (+27%) | 0.41 (+10.8%) | 0.30 (−18.9%) | 0.40 (+8.1%) | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sehgal, A.; Reddy, K.R.; Walne, C.H.; Barickman, T.C.; Brazel, S.; Chastain, D.; Gao, W. Climate Stressors on Growth, Yield, and Functional Biochemistry of two Brassica Species, Kale and Mustard. Life 2022, 12, 1546. https://doi.org/10.3390/life12101546
Sehgal A, Reddy KR, Walne CH, Barickman TC, Brazel S, Chastain D, Gao W. Climate Stressors on Growth, Yield, and Functional Biochemistry of two Brassica Species, Kale and Mustard. Life. 2022; 12(10):1546. https://doi.org/10.3390/life12101546
Chicago/Turabian StyleSehgal, Akanksha, Kambham Raja Reddy, Charles Hunt Walne, T. Casey Barickman, Skyler Brazel, Daryl Chastain, and Wei Gao. 2022. "Climate Stressors on Growth, Yield, and Functional Biochemistry of two Brassica Species, Kale and Mustard" Life 12, no. 10: 1546. https://doi.org/10.3390/life12101546