
Similar Responses of Relatively Salt-Tolerant Plants to Na and K during Chloride Salinity: Comparison of Growth, Water Content and Ion Accumulation

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Plant Establishment, Cultivation and Treatments
2.3. Plant Harvest and Measurements
2.4. Data Analysis
3. Results
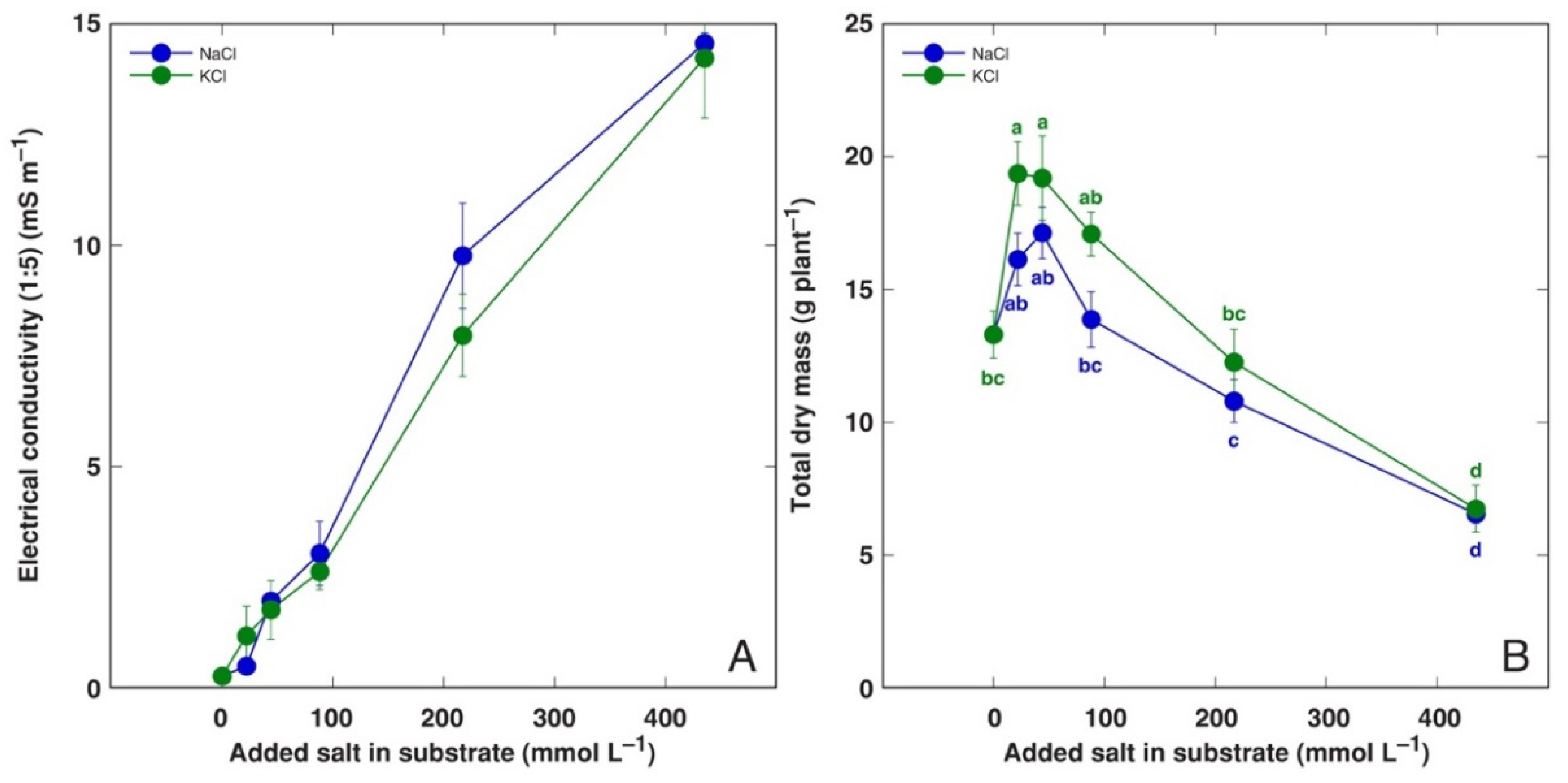
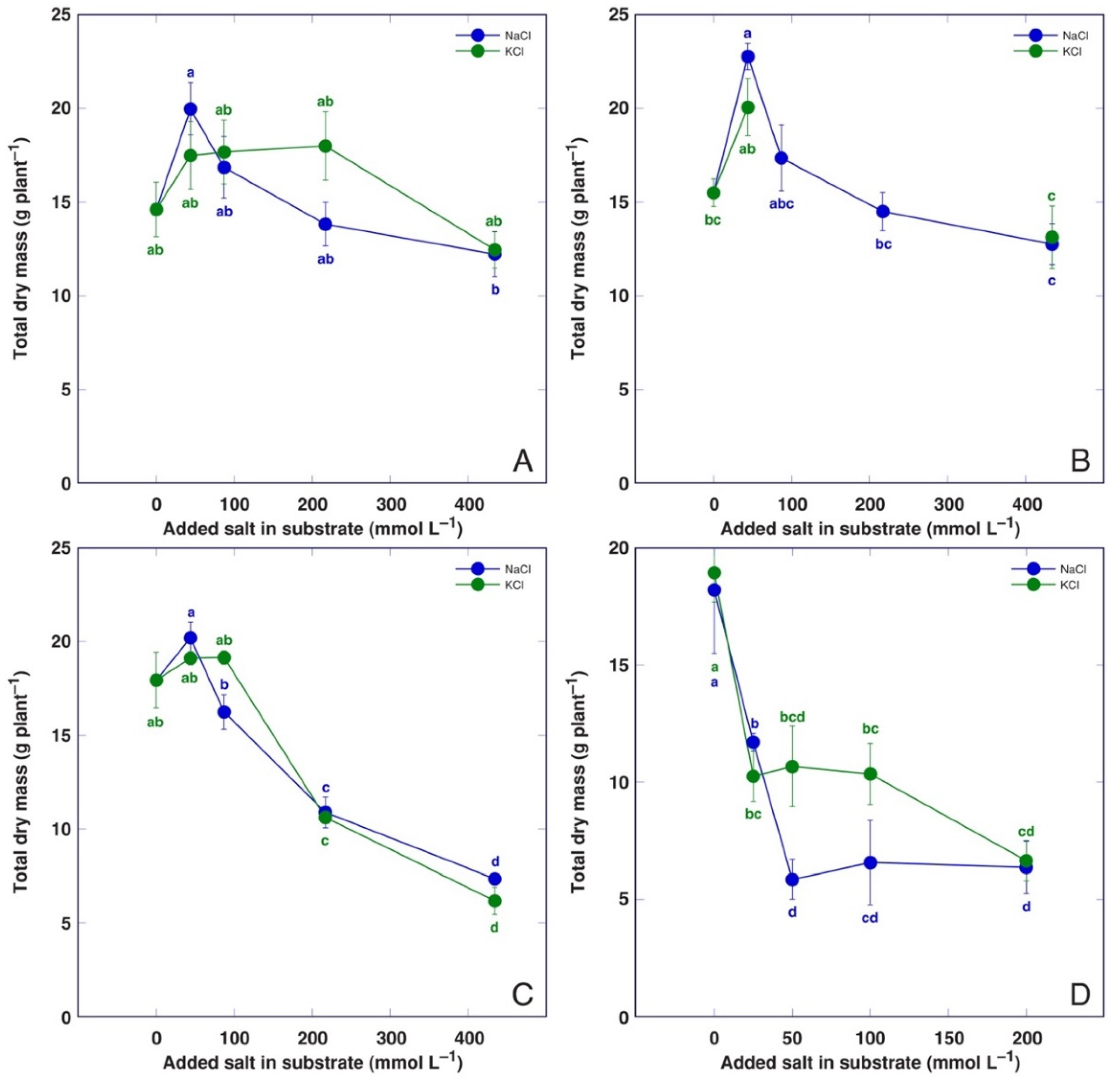
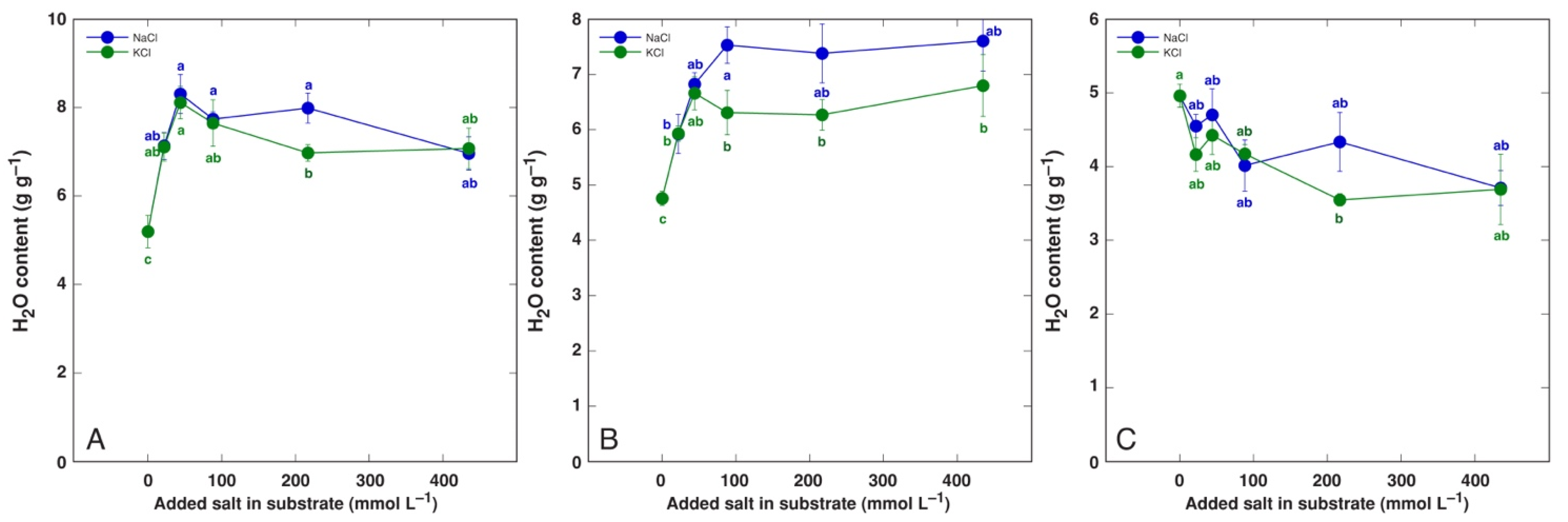
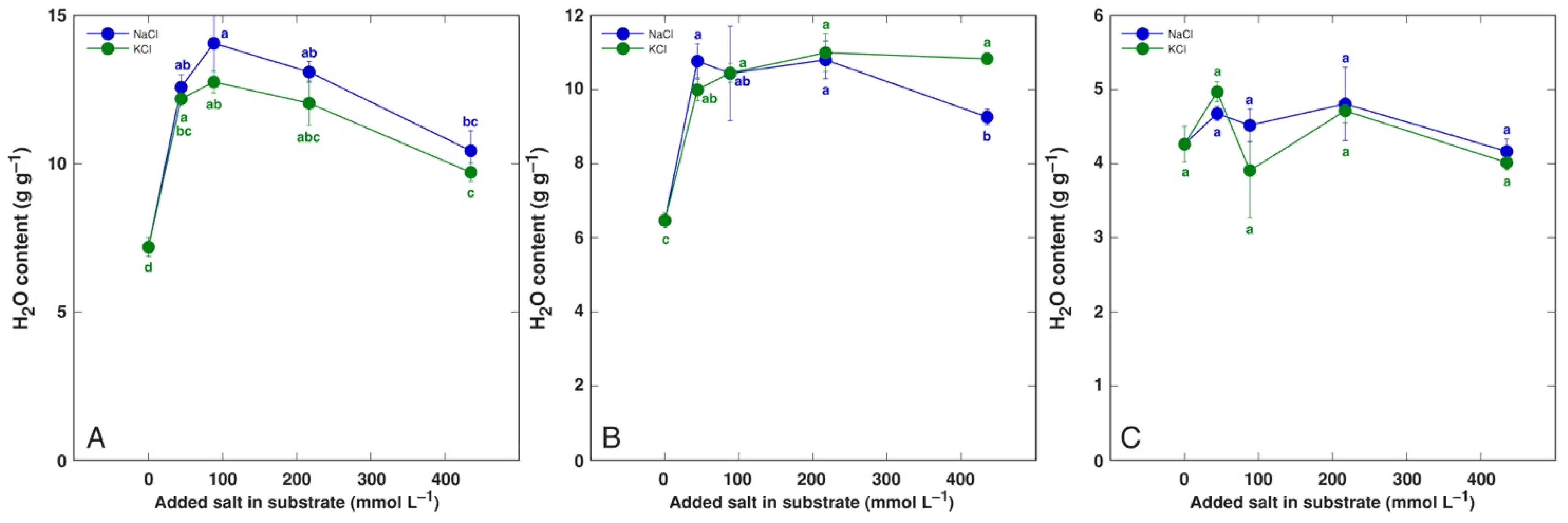
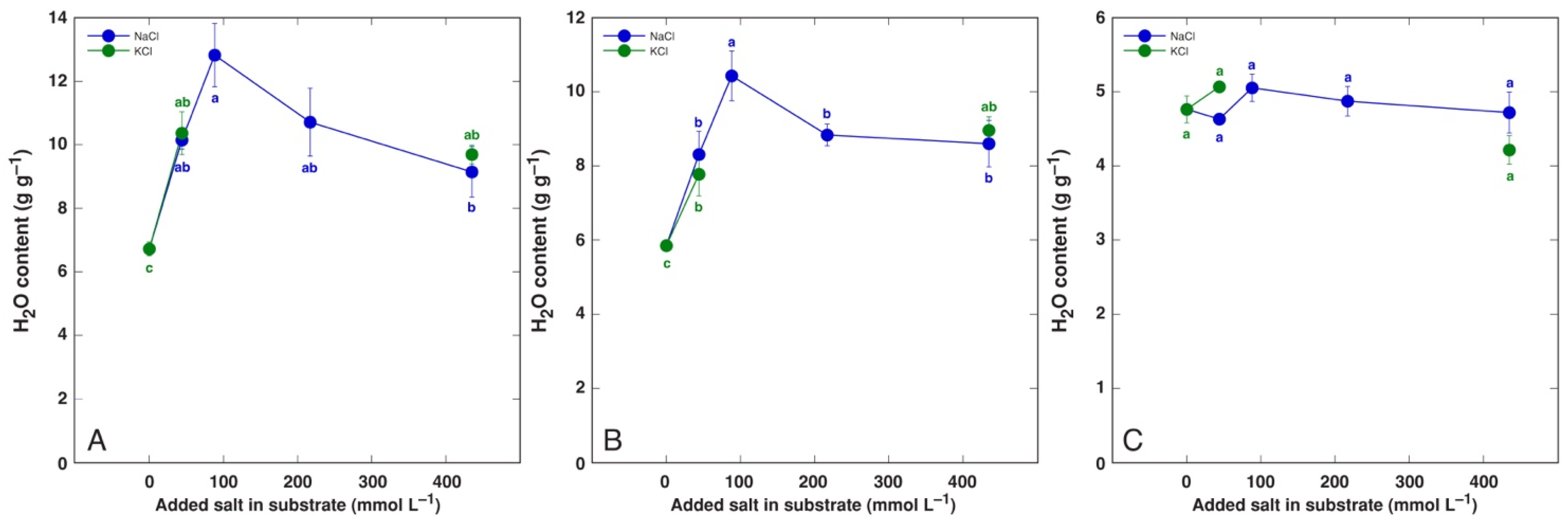
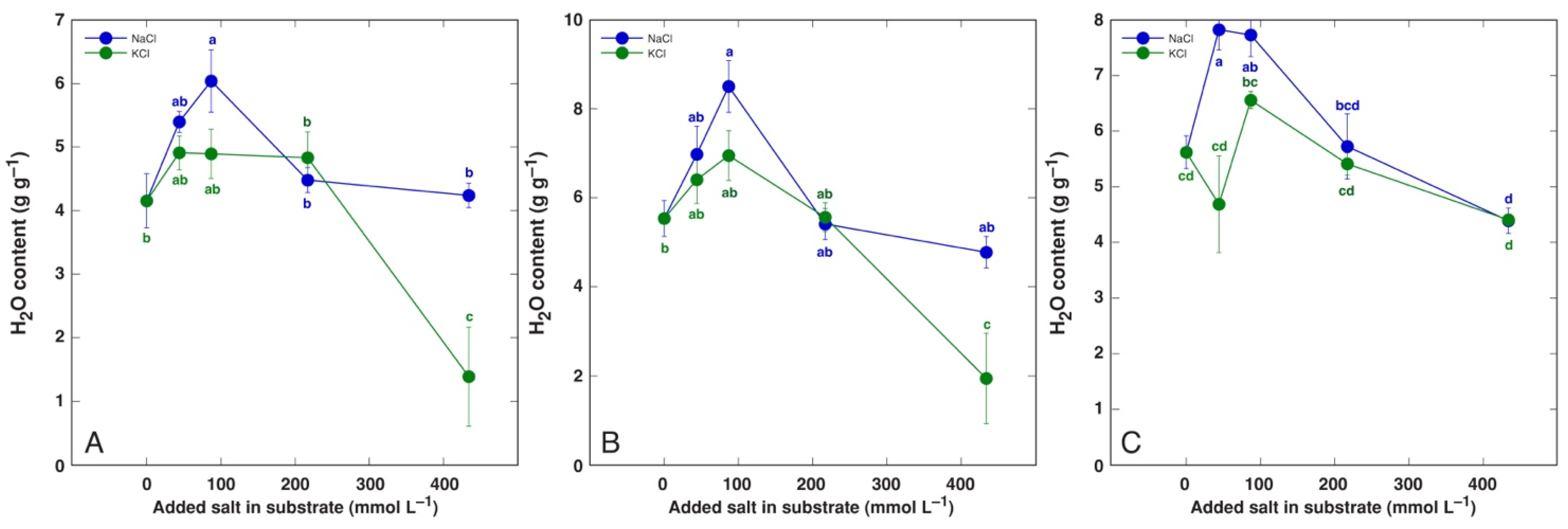
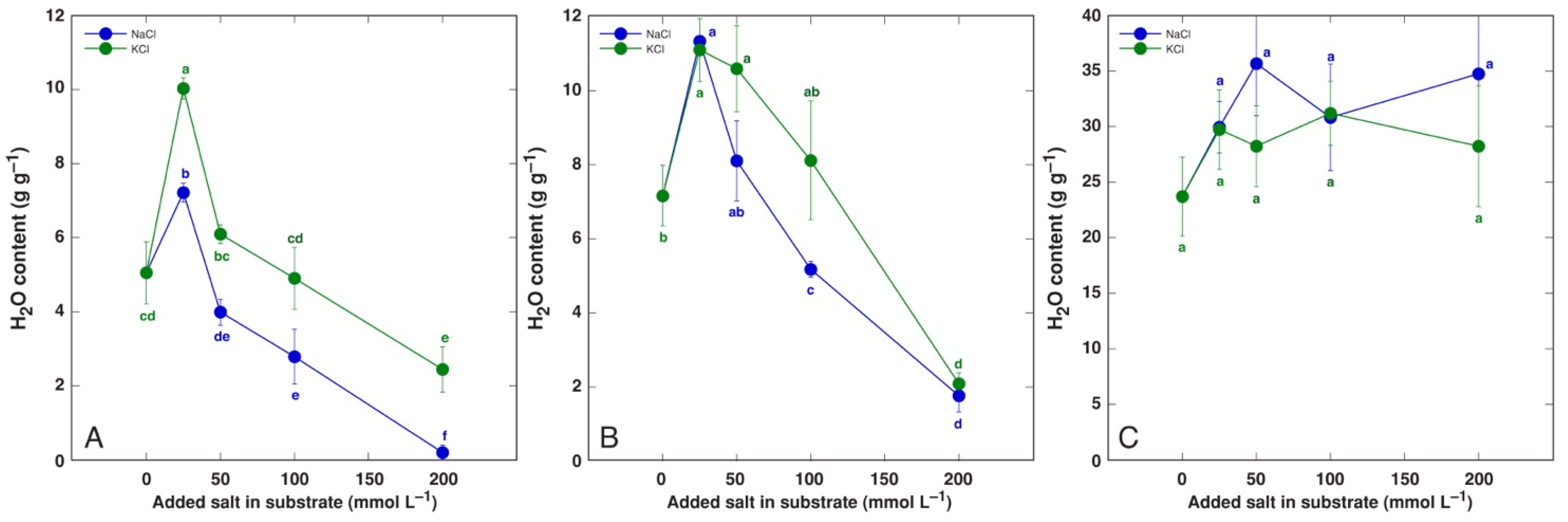
3.1. Effect on Plant Growth and Water Content
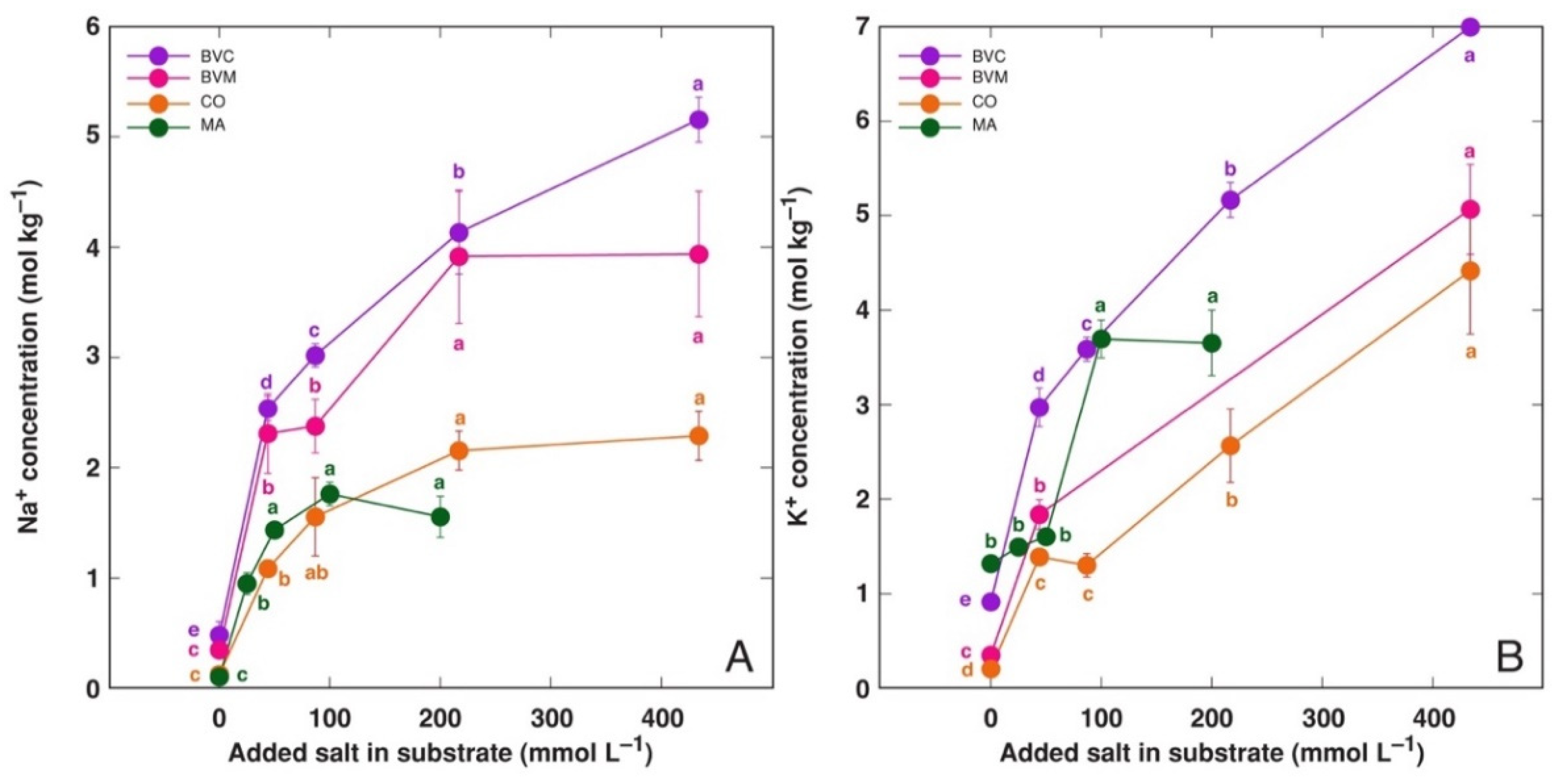
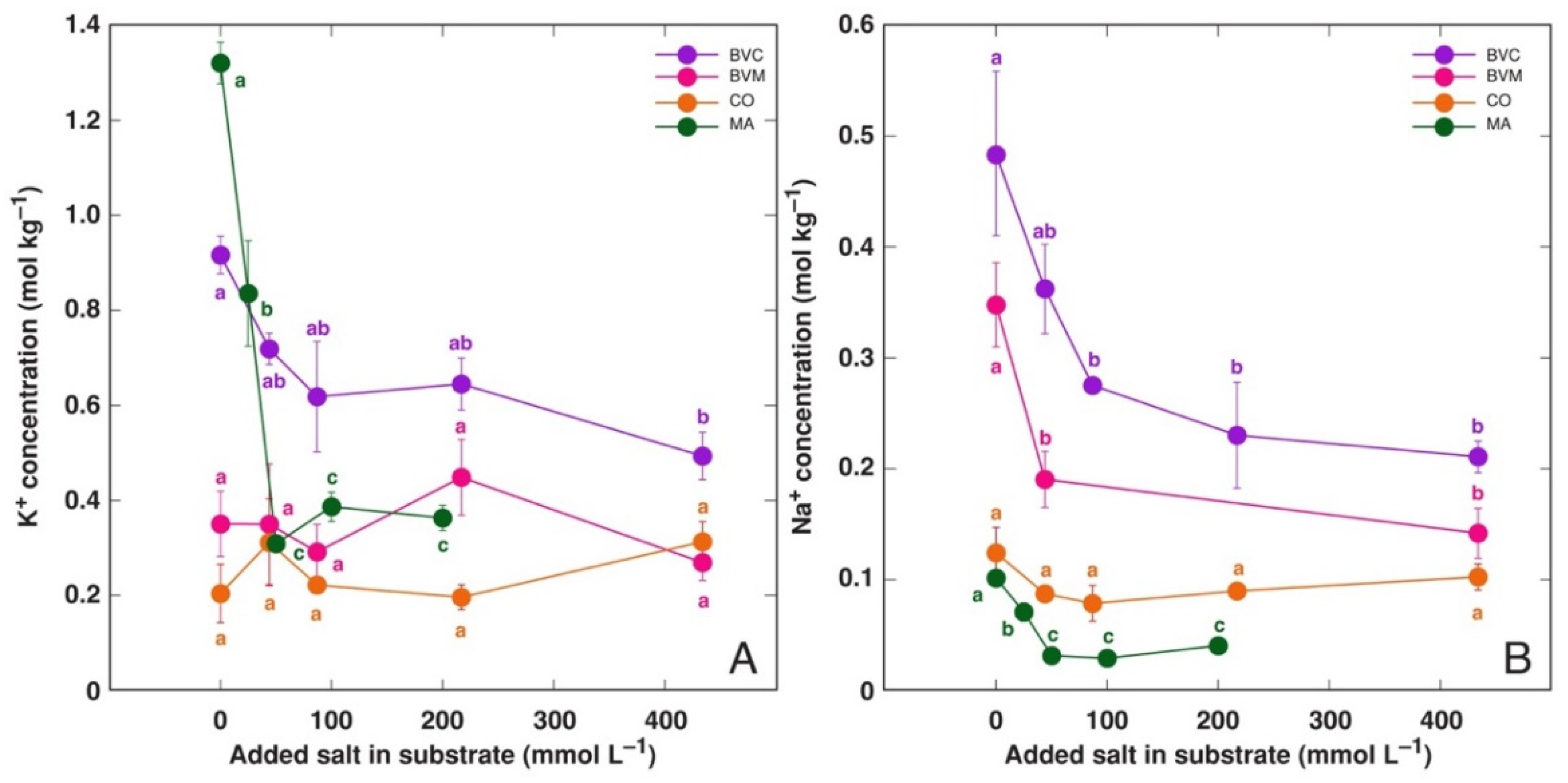
3.2. Effect on Ion Accumulation
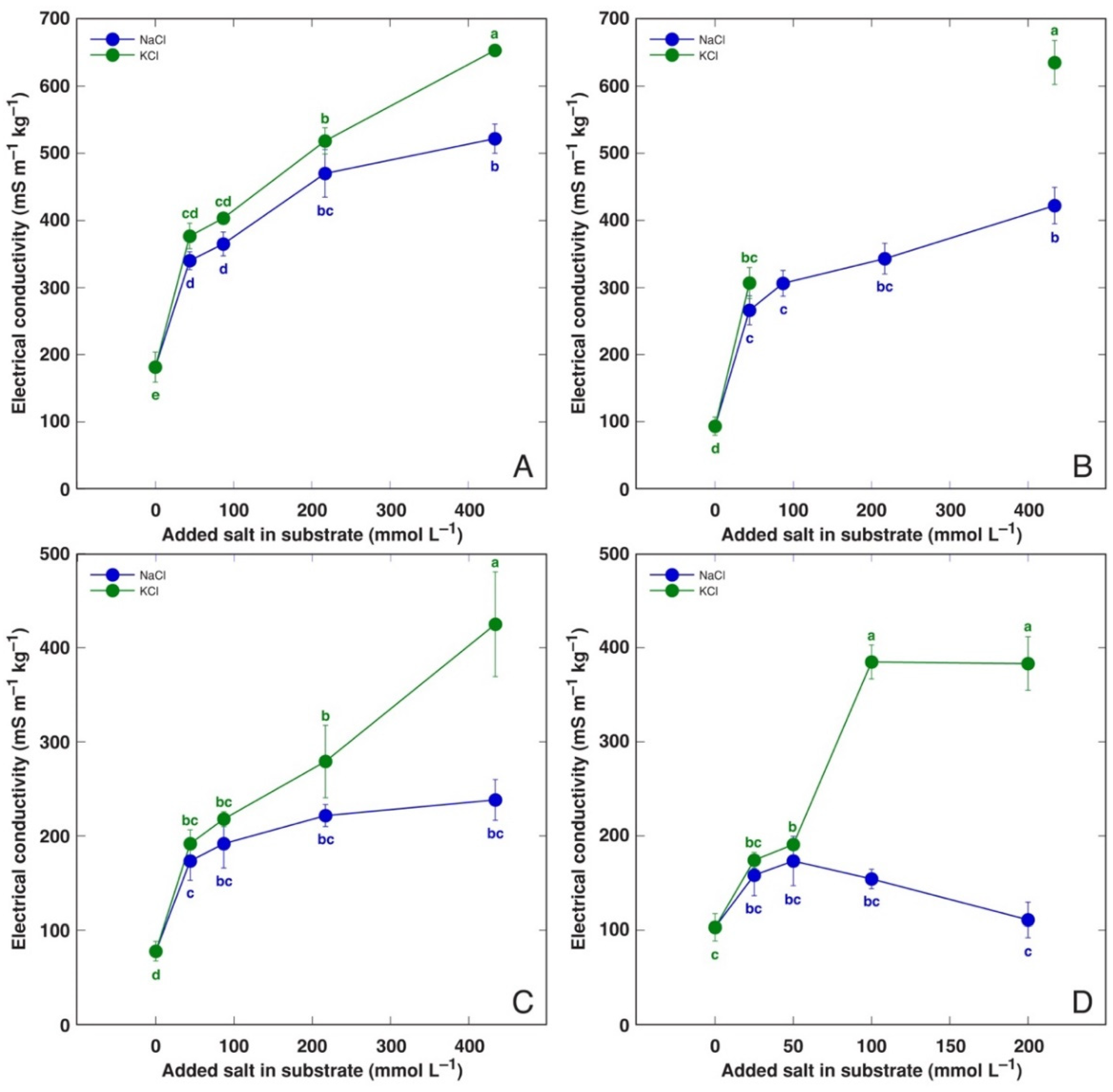
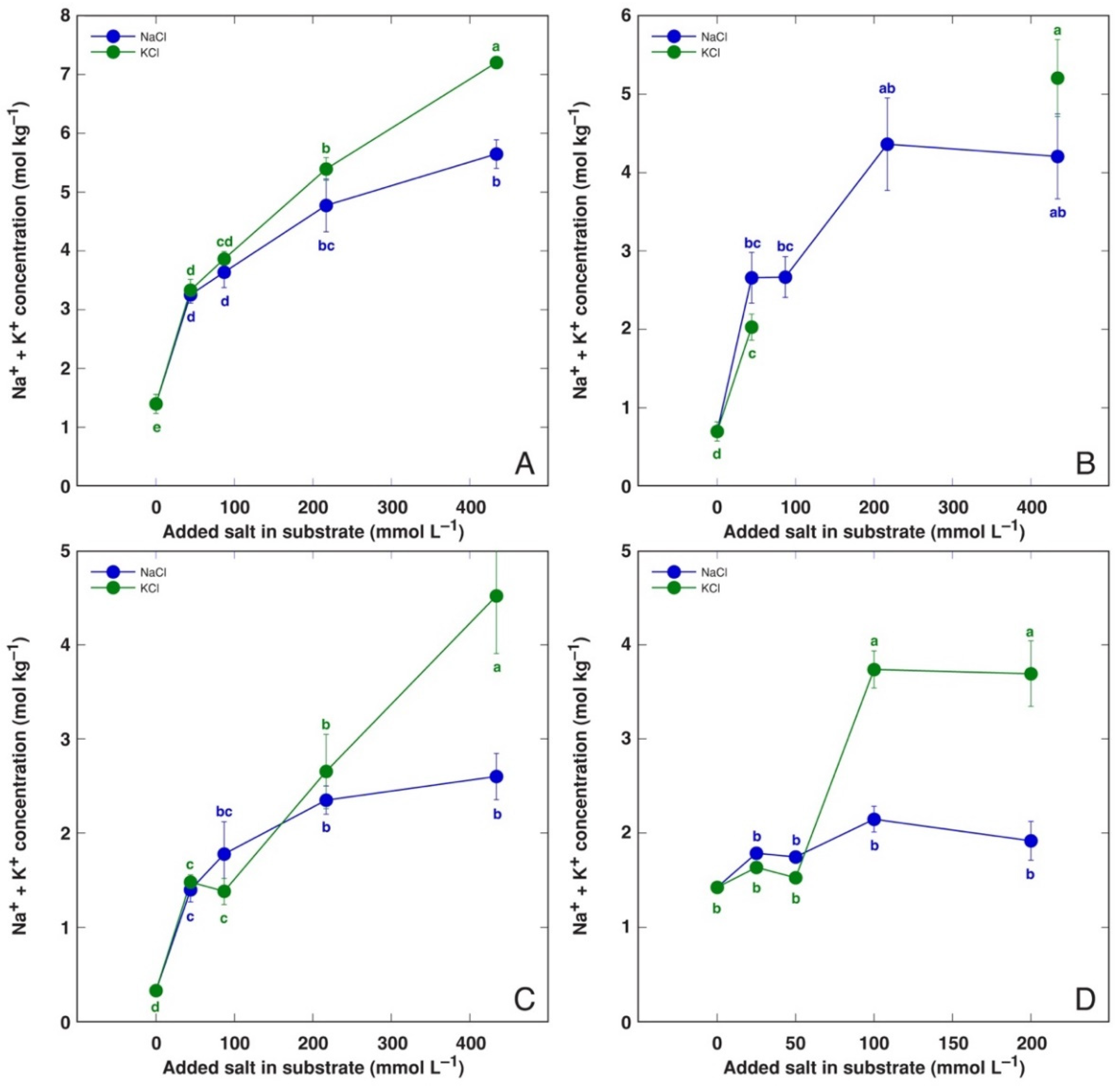
3.3. Comparison of NaCl and KCl Effects
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- van Zelm, E.; Zhang, Y.; Testerink, C. Salt tolerance mechanisms of plants. Annu. Rev. Plant Biol. 2020, 71, 403–433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fang, S.; Hou, X.; Liang, X. Response mechanisms of plants under saline-alkali stress. Front. Plant Sci. 2021, 12, 667458. [Google Scholar] [CrossRef]
- Hameed, A.; Ahmed, M.Z.; Hussain, T.; Aziz, I.; Ahmad, N.; Gul, B.; Nielsen, B.L. Effects of salinity stress on chloroplast structure and function. Cells 2021, 10, 2023. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.M.; Mostofa, M.G.; Keya, S.S.; Siddiqui, M.N.; Ansary, M.M.; Das, A.K.; Rahman, M.A.; Tran, L.S.-P. Adaptive mechanisms of halophytes and their potential in improving salinity tolerance in plants. Int. J. Mol. Sci. 2022, 22, 10733. [Google Scholar] [CrossRef] [PubMed]
- Soltabayeva, A.; Ongaltay, A.; Omondi, J.O.; Srivastava, S. Morphological, physiological and molecular markers for salt-stressed plants. Plants 2021, 10, 243. [Google Scholar] [CrossRef]
- Valenzuela, F.J.; Reineke, D.; Leventini, D.; Chen, C.C.L.; Barrett-Lennard, E.G.; Colmer, T.D.; Dodd, I.C.; Shabala, S.; Brown, P.; Bazihizina, N. Plant responses to heterogeneous salinity: Agronomic relevance and research priorities. Ann. Bot. 2022, 129, 499–517. [Google Scholar] [CrossRef] [PubMed]
- Ondrasek, G.; Rengel, Z. Environmental salinization processes: Detection, implications & solutions. Sci. Total Environ. 2021, 754, 142432. [Google Scholar]
- Maathuis, F.J.M.; Amtmann, A. K+ nutrition and Na+ toxicity: The basis of cellular K+/Na+ ratios. Ann. Bot. 1999, 84, 123–133. [Google Scholar] [CrossRef] [Green Version]
- Kronzucker, H.J.; Coskun, D.; Schulze, L.M.; Wong, J.R.; Britto, D.T. Sodium as nutrient and toxicant. Plant Soil 2013, 369, 1–23. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Ballesta, M.C.; Martínez, V.; Carvajal, M. Osmotic adjustment, water relations and gas exchange in pepper plants grown under NaCl and KCl. Environ. Exp. Bot. 2004, 52, 161–174. [Google Scholar] [CrossRef]
- Ramos, J.; López, M.J.; Benlloch, M. Effect of NaCl and KCl salts on the growth and solute accumulation of the halophyte Atriplex nummularia. Plant Soil 2004, 259, 163–168. [Google Scholar] [CrossRef]
- Wang, D.; Wang, H.; Han, B.; Wang, B.; Guo, A.; Zheng, D.; Liu, C.; Chang, L.; Peng, M.; Wang, X. Sodium instead of potassium and chloride is an important macronutrient to improve leaf succulence and shoot development for halophyte Sesuvium portulacastrum. Plant Physiol. Biochem. 2012, 51, 53–62. [Google Scholar] [CrossRef]
- Belkheiri, O.; Mulas, M. The effects of salt stress on growth, water relations and ion accumulation in two halophyte Atriplex species. Environ. Exp. Bot. 2013, 86, 17–28. [Google Scholar] [CrossRef]
- Zhao, C.; Zhang, H.; Song, C.; Zhu, J.-K.; Shabala, S. Mechanisms of plant responses and adaptation to soil salinity. Innovation 2020, 1, 100017. [Google Scholar] [CrossRef]
- Surówka, E.; Hura, T. Osmoprotectants and nonenzymatic antioxidants in halophytes. In Handbook of Halophytes; Grigore, M.-N., Ed.; Springer: Cham, Switzerland, 2021; pp. 1901–1930. [Google Scholar]
- Ievinsh, G.; Ieviņa, S.; Andersone-Ozola, U.; Samsone, I. Leaf sodium, potassium and electrolyte accumulation capacity of plant species from salt-affected coastal habitats of the Baltic Sea: Towards a definition of Na hyperaccumulation. Flora 2021, 274, 151748. [Google Scholar] [CrossRef]
- Leys, M.; Petit, E.J.; El-Bahloul, J.; Liso, C.; Fournet, S.; Arnaud, J.-F. Spatial genetic structure in Beta vulgaris subsp. maritima and Beta macrocarpa reveals the effect of contrasting mating system, influence of marine currents, and footprints of postglacial recolonization routes. Ecol. Evol. 2014, 4, 1828–1852. [Google Scholar] [CrossRef]
- Andrello, M.; Henry, K.; Devaux, P.; Desprez, B.; Manel, S. Taxonomic, spatial and adaptive genetic variation of Beta section Beta. Theor. Appl. Genet. 2015, 129, 257–271. [Google Scholar] [CrossRef]
- Frese, L.; Ford-Lloyd, B. Range of distribution. In Beta maritima, the Origin of Beets, 2nd ed.; Biancardi, E., Panella, L.W., McGrath, J.M., Eds.; Springer: Cham, Switzerland, 2020; pp. 49–60. [Google Scholar]
- Frese, L.; Ford-Lloyd, B. Taxonomy, phylogeny, and the genepool. In Beta maritima, the Origin of Beets, 2nd ed.; Biancardi, E., Panella, L.W., McGrath, J.M., Eds.; Springer: Cham, Switzerland, 2020; pp. 121–151. [Google Scholar]
- Bor, M.; Özdemir, F.; Türkan, I. The effect of salt stress on lipid peroxidation and antioxidants in leaves of sugar beet Beta vulgaris L. and wild beet Beta maritima L. Plant Sci. 2003, 164, 77–84. [Google Scholar] [CrossRef]
- Koyro, H.-W.; Daoud, S.; Harrouni, C.; Huchzermeyer, B. Strategies of a potential cash crop halophyte (Beta vulgaris ssp. maritima) to avoid salt injury. Trop. Ecol. 2006, 47, 191–200. [Google Scholar]
- Ribeiro, I.C.; Pinheiro, C.; Ribeiro, C.M.; Veloso, M.; Simões-Costa, M.C.; Evaristo, I.; Paulo, O.S.; Ricardo, C.P. Genetic diversity and physiological performance of Portuguese wild beet (Beta vulgaris spp. maritima) from three contrasting habitats. Front. Plant Sci. 2016, 7, 1293. [Google Scholar] [CrossRef] [Green Version]
- Skorupa, M.; Gołȩbiewski, M.; Kurnik, K.; Niedojadło, J.; Kȩsy, J.; Klamowski, K.; Wójcik, K.; Treder, W.; Tretyn, A.; Tyburski, J. Salt stress vs. salt shock—The case of sugar beet and its halophytic ancestor. BMC Plant Biol. 2019, 19, 57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, L.; Ueda, A.; Saneoka, H. Physiological responses of white Swiss chard (Beta vulgaris L. subsp. cicla) to saline and alkaline stress. Austr. J. Crop Sci. 2013, 7, 1046–1052. [Google Scholar]
- Kaburagi, E.; Morikawa, Y.; Yamada, M.; Fujiyama, H. Sodium enhances nitrate uptake in Swiss chard (Beta vulgaris var. cicla L.). Soil Sci. Plant Nutr. 2014, 60, 651–658. [Google Scholar] [CrossRef]
- Puccinelli, M.; Carmassi, G.; Botrini, L.; Bindi, A.; Rossi, L.; Fierro-Sañudo, J.F.; Pardossi, A.; Incrocci, L. Growth and mineral relations of Beta vulgaris var. cicla and Beta vulgaris ssp. maritima cultivated hydroponically with diluyed seawater and low nitrogen level in the nutrient solution. Horticulturae 2022, 8, 638. [Google Scholar]
- Rozema, J.; Cornelisse, D.; Zhang, Y.; Li, H.; Bruning, B.; Katschnig, D.; Broekman, R.; Ji, B.; van Bodegom, P. Comparing salt tolerance of beet cultivars and their halophytic ancestor: Consequences of domestication and breeding programmes. AoB Plants 2014, 7, plu083. [Google Scholar] [CrossRef] [Green Version]
- Nawaz, I.; Iqbal, M.; Bliek, M.; Schat, H. Salt and heavy metal tolerance and expression levels of candidate tolerance genes among four extremophile Cochlearia species with contrasting habitat preferences. Sci. Total Environ. 2017, 584–585, 731–741. [Google Scholar] [CrossRef]
- de Vos, A.C.; Broekman, R.; de Almeida Guerra, C.C.; van Rijsselberghe, M.; Rozema, J. Developing and testing new halophyte crops: A case study of salt tolerance of two species of the Brassicaceae, Diplotaxis tenuifolia and Cochlearia officinalis. Environ. Exp. Bot. 2013, 92, 154–164. [Google Scholar] [CrossRef]
- Nordal, I.; Eriksen, A.B.; Laane, M.M.; Solberg, Y. Biogeographic and biosystematic studies in the genus Cochlearia in Northern Scandinavia. Acta Univ. Upsaliensis. Symb. Bot. Ups. 1986, 27, 83–93. [Google Scholar]
- Janssen, J.A.M.; Rodwell, J.S. European Red List of Habitats: Part 2. Terrestrial and Freshwater Habitats; European Union: Brussels, Belgium, 2016. [Google Scholar]
- Puijalon, S.; Bouma, T.J.; van Groenendael, J.; Bornette, G. Clonal plasticity of aquatic plant species submitted to mechanical stress: Escape versus resistance strategy. Ann. Bot. 2008, 102, 898–996. [Google Scholar] [CrossRef] [Green Version]
- Haddadi, B.S.; Hassanpour, H.; Niknam, V. Effect of salinity and waterlogging on growth, anatomical and antioxidative responses in Mentha aquatica L. Acta Physiol. Plant. 2016, 38, 119. [Google Scholar] [CrossRef]
- Sniedze-Kretalova, R. 6430. Hydrophilous tall herb fringe communities of plains and the montane to alpine levels. In European Union Protected Habitats in Latvia. Interpretation Manual; Auniņš, A., Ed.; Latvian Fund for Nature, Ministry of Environmental Protection and Regional Development: Riga, Latvia, 2013; pp. 186–189. [Google Scholar]
- Rūsiņa, S. 6430. Water courses of plain to montane levels with the Ranunculion fluitantis and Callitricho-Batrachion vegetation. In European Union Protected Habitats in Latvia. Interpretation Manual; Auniņš, A., Ed.; Latvian Fund for Nature, Ministry of Environmental Protection and Regional Development: Riga, Latvia, 2013; pp. 123–126. [Google Scholar]
- Kalniņš, M.; Andersone-Ozola, U.; Gudrā, D.; Sieriņa, A.; Fridmanis, D.; Ievinsh, G.; Muter, O. Effect of bioaugumentation on the growth and rhizosphere microbiome assembly of hydroponic cultures of Mentha aquatica. Ecol. Genet. Genom. 2022, 22, 100107. [Google Scholar]
- Erdei, L.; Kuiper, P.J.C. The effect of salinity on growth, cation content, Na+ uptake and translocation in salt-sensitive and salt-tolerant Plantago species. Physiol. Plant. 1979, 47, 95–99. [Google Scholar] [CrossRef]
- Tánczos, O.G.; Erdei, L.; Snijder, J. Uptake and translocation of sodium in salt-sensitive and salt-tolerant Plantago species. Plant Soil 1981, 63, 27–32. [Google Scholar] [CrossRef]
- Königshofer, H. Changes in ion composition and hexitol content of different Plantago species under the influence of salt stress. Plant Soil 1983, 72, 289–296. [Google Scholar] [CrossRef]
- Blom, C.W.P.M. Phenotypic plasticity in Plantago maritima. In Plantago: A Multidisciplinary Study; Kuiper, P.C.J., Bos, M., Eds.; Springer: Berlin, Germany, 1992; pp. 240–250. [Google Scholar]
- Sleimi, N.; Guerfali, S.; Bankaji, I. Biochemical indicators of salt stress in Plantago maritima: Implications for environmental stress assessment. Ecol. Indic. 2015, 48, 570–577. [Google Scholar] [CrossRef]
- Rubinigg, M.; Posthumus, F.; Ferschke, M.; Elzenga, J.T.M.; Stulen, I. Effects of NaCl salinity on 15N-nitrate fluxes and specific root length in the halophyte Plantago maritima L. Plant Soil 2003, 250, 201–213. [Google Scholar] [CrossRef]
- Jerling, L. The impact of some environmental factors on the establishment of Plantago maritima seedlings and juveniles along a distributional gradient. Holarctic Ecol. 1984, 7, 271–279. [Google Scholar] [CrossRef]
- Jerling, L. Population dynamics of Plantago maritima along a distributional gradient on a Baltic seashore meadow. Vegetatio 1985, 61, 155–161. [Google Scholar] [CrossRef]
- Jerling, L.; Liljelund, L.-E. Dynamics of Plantago maritima along a distributional gradient: A demographic study. Holarctic Ecol. 1984, 7, 280–288. [Google Scholar] [CrossRef]
- He, H.; Zhou, H.; Lü, H.; Liang, B. Growth, morphological and physiological adaptability of leaf beat (Beta vulgaris var. cicla) to salt stress: A soil culture experiment. Agronomy 2022, 12, 1393. [Google Scholar]
- Tavakkoli, E.; Fatehi, F.; Rengasamy, P.; McDonald, G.K. A comparison of hydroponic and soil-based screening methods to identify salt tolerance in the field in barley. J. Exp. Bot. 2012, 63, 3853–3868. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walker, D.J.; Leigh, R.A.; Miller, A.J. Potassium homeostasis in vacuolated plant cells. Proc. Natl. Acad. Sci. USA 1996, 93, 10510–10514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodríguez-Navarro, A.; Rubio, F. High-affinity potassium and sodium transport systems in plants. J. Exp. Bot. 2006, 57, 1149–1160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adams, E.; Shin, R. Transport, signaling, and homeostasis of potassium and sodium in plants. J. Integr. Plant Biol. 2014, 56, 231–249. [Google Scholar] [CrossRef]
- Jennings, D.H. Halophytes, succulence and sodium in plants–a unified theory. New Phytol. 1968, 67, 899–911. [Google Scholar] [CrossRef]
- Song, J.; Wang, B. Using euhalophytes to understand salt tolerance and to develop saline agriculture: Suaeda salsa as a promising model. Ann. Bot. 2015, 115, 541–553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, F.; Xu, X.; Leng, B.; Wang, B. Beneficial effects of salinity on halophyte growth: Morphology, cells, and genes. Open Life Sci. 2019, 14, 191–200. [Google Scholar] [CrossRef] [PubMed]
- Allu, A.D.; Soja, A.M.; Wu, A.; Szymanski, J.; Balazadeh, S. Salt stress and senescence: Identification of cross-talk regulatory components. J. Exp. Bot. 2014, 65, 3993–4008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Purmale, L.; Jēkabsone, A.; Andersone-Ozola, U.; Ievinsh, G. Salinity tolerance, ion accumulation potential and osmotic adjustment in vitro and in planta of different Armeria maritima accessions from a dry coastal meadow. Plants 2022, 11, 2570. [Google Scholar] [CrossRef]
- Schulze, E.D.; Beck, E.; Buchmann, N.; Clemens, S.; Müller-Hohenstein, K.; Scherer-Lorenzen, M. Adverse soil mineral availability. In Plant Ecology; Springer: Berlin/Heidelberg, Germany, 2019; pp. 203–256. [Google Scholar]
- Bonales-Alatorre, E.; Shabala, S.; Chen, Z.-H.; Pottosin, I. Reduced tonoplast fast-activating and slow-activating channel activity is essential for conferring salinity tolerance in a facultative halophyte, quinoa. Plant Physiol. 2013, 162, 940–952. [Google Scholar] [CrossRef] [Green Version]
- Szarek-Łukaszewska, G.; Słysz, A.; Wierzbicka, M. Response of Armeria maritima (Mill.) Willd. to Cd, Zn and Pb. Acta Biol. Cracov. Ser. Bot. 2004, 46, 19–24. [Google Scholar]
- Zajc, J.; Džeroski, S.; Kocev, D.; Oren, A.; Sonjak, S.; Tkavc, R.; Gunde-Cimerman, N. Chaophilic or chaotolerant fungi: A new category of extremophiles? Front. Microbiol. 2014, 5, 708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collins, K.D. Why continuum electrostatic theories cannot explain biological structure, polyelectrolytes or ionic strength effects in ion–protein interactions. Biophys. Chem. 2012, 167, 43–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, G.; Magen, H.; Tarchitzky, J.; Kafkafi, U. Advances in chloride nutrition of plants. In Advances in Agronomy; Academic Press: Cambridge, MA, USA, 2000; Volume 68, pp. 97–150. [Google Scholar]
- Colmenero-Flores, J.M.; Franco-Navarro, J.D.; Cuberto-Font, P.; Peinado-Torrubia, P.; Rosales, M.A. Chloride as a beneficial macronutrient in higher plants: New roles and regulation. Int. J. Mol. Sci. 2019, 20, 4684. [Google Scholar] [CrossRef] [Green Version]
- Martin, P.K.; Koebner, R.M.D. Sodium and chloride ions contribute synergistically to salt toxicity in wheat. Biol. Plant. 1995, 37, 265–271. [Google Scholar] [CrossRef]
- Raven, J.A. Chloride: Essential micronutrient and multifunctional beneficial ion. J. Exp. Bot. 2017, 68, 359–367. [Google Scholar] [CrossRef]
- Geilfus, C.M. Chloride: From nutrient to toxicant. Plant Cell Physiol. 2018, 59, 877–886. [Google Scholar] [CrossRef]
- Bazihizina, N.; Colmer, T.D.; Cuin, T.A.; Mancuso, S.; Shabala, S. Friend or foe? Chloride patterning in halophytes. Trends Plant Sci. 2019, 24, 142–151. [Google Scholar] [CrossRef]
- Flowers, T.J.; Glenn, E.P.; Volkov, V. Could vesicular transport of Na+ and Cl− be a feature of salt tolerance in halophytes? Ann. Bot. 2019, 123, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Le, L.T.T.; Kotula, L.; Siddique, K.H.M.; Colmer, T.D. Na+ and/or Cl– toxicities determine salt sensitivity in soybean (Glycine max (L.) Merr.), mungbean (Vigna radiata (L.) R. Wilczek), cowpea (Vigna unguiculata (L.) Walp.) and common bean (Phaseolus vulgaris L.). Int. J. Mol. Sci. 2021, 22, 1909. [Google Scholar] [CrossRef]
- Kawakami, K.; Umena, Y.; Kamiya, N.; Shen, J.R. Location of chloride and its possible functions in oxygen-evolving photosystem II revealed by X-ray crystallography. Proc. Natl. Acad. Sci. USA 2009, 106, 8567–8572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franco-Navarro, J.D.; Brumos, J.; Rosales, M.A.; Cubero-Font, P.; Talon, M.; Colmenero-Flores, J.M. Chloride regulates leaf cell size and water relations in tobacco plants. J. Exp. Bot. 2016, 67, 873–891. [Google Scholar] [CrossRef] [Green Version]
- Yamada, M.; Kuroda, C.; Fujiyama, H. Growth promotion by sodium in amaranthaceous plants. J. Plant Nutr. 2016, 39, 1186–1193. [Google Scholar] [CrossRef]
- Yamada, M.; Kuroda, C.; Fujiyama, H. Function of sodium and potassium in growth of sodium-lowing Amaranthaceae species. Soil Sci. Plant Nutr. 2016, 62, 20–26. [Google Scholar] [CrossRef] [Green Version]
- Kaburagi, E.; Yamada, M.; Fujiyama, H. Sodium, but not potassium, enhances root to leaf nitrate translocation in Swicc chard (Beta vulgaris var. cicla L.). Environ. Exp. Bot. 2015, 112, 27–32. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Code | Model Plant | Propagation Material (Source) | Cultivation System | Treatments | Analyzed Parameters |
|---|---|---|---|---|---|
| BVC | Beta vulgaris subsp. vulgaris L. var. cicla cv. ‘Magenta Sunset’ | Seeds (Sėklos, Vilnius, Lithuania) | Garden soil/quartz sand 3:1 (v/v) | NaCl (44, 87, 217, 434 mmol L−1). KCl (44, 87, 217, 434 mmol L−1) | Dry biomass, H2O, Na, K, EC |
| BVM | Beta vulgaris subsp. maritima (L.) Arcang. | Seeds (Agroforestry Research Trust, Dartington, Devon, UK) | Garden soil/quartz sand 3:1 (v/v) | NaCl (44, 87, 217, 434 mmol L−1). KCl (44, 434 mmol L−1) | Dry biomass, H2O, Na, K, EC |
| CO | Cochlearia officinalis L. | Seeds (Jelitto Staudensamen, Schwarmstedt, Germany) | Garden soil/quartz sand 5:1 (v/v) | NaCl (44, 87, 217, 434 mmol L−1). KCl (44, 87, 217, 434 mmol L−1) | Dry biomass, H2O, Na, K, EC |
| MA | Mentha aquatica L. | Stem explants (seawater-affected sandy beach, Ainaži, Latvia) | Hydroponics, Kristalon Red + Calcinit (0.5 g L−1) | NaCl (25, 50, 100, 200 mmol L−1). KCl (25, 50, 100, 200 mmol L−1) | Dry biomass, H2O, Na, K, EC |
| PM | Plantago maritima L. | Seeds (seawater-affected gravel beach, Ohesaare, island of Saaremaa, Estonia) | Garden soil/quartz sand 3:1 (v/v) | NaCl (22, 44, 87, 217, 434 mmol L−1). KCl (22, 44, 87, 217, 434 mmol L−1) | Dry biomass, H2O |
| Model Plant | Plant Part | R2 | r |
|---|---|---|---|
| BVC | Leaf petioles | 0.978 | 0.989 |
| Leaf blades | 0.984 | 0.992 | |
| Roots | 0.970 | 0.985 | |
| BVM | Leaf petioles | 0.873 | 0.934 |
| Leaf blades | 0.860 | 0.927 | |
| Roots | 0.863 | 0.929 | |
| CO | Leaf petioles | 0.995 | 0.997 |
| Leaf blades | 0.982 | 0.991 | |
| Roots | 0.970 | 0.985 | |
| MA | Stems | 0.548 | 0.741 |
| Leaves | 0.822 | 0.907 | |
| Roots | 0.625 | 0.790 |
| Parameter | Model Plant | Effect of NaCl | Effect of KCl | Na+ vs. K+ |
|---|---|---|---|---|
| Maximum dry mass increase (%) | BVC | 37 | 20 | n.s. |
| BVM | 47 * | 29 | n.s. | |
| CO | 13 | 7 | n.s. | |
| MA | 0 | 0 | n.s. | |
| PM | 29 | 46 * | n.s. | |
| Maximum dry mass decrease (%) | BVC | 16 | 15 | n.s. |
| BVM | 18 | 15 | n.s. | |
| CO | 59 * | 66 * | n.s. | |
| MA | 65 * | 65 * | n.s. | |
| PM | 51 * | 49 * | n.s. | |
| Maximum increase in H2O in leaf blades (%) | BVC | 96 * | 78 * | n.s. |
| BVM | 91 * | (54 *) | - | |
| CO | 45 * | 18 | n.s. | |
| MA | 43 * | 97 * | s. | |
| PM | 60 * | 56 * | n.s. | |
| Maximum increase in EC in leaf blades (%) | BVC | 187 * | 260 * | s. |
| BVM | 351 * | 579 * | s. | |
| CO | 207 * | 447 * | s. | |
| MA | 68 | 272 * | s. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ievinsh, G.; Andersone-Ozola, U.; Jēkabsone, A. Similar Responses of Relatively Salt-Tolerant Plants to Na and K during Chloride Salinity: Comparison of Growth, Water Content and Ion Accumulation. Life 2022, 12, 1577. https://doi.org/10.3390/life12101577
Ievinsh G, Andersone-Ozola U, Jēkabsone A. Similar Responses of Relatively Salt-Tolerant Plants to Na and K during Chloride Salinity: Comparison of Growth, Water Content and Ion Accumulation. Life. 2022; 12(10):1577. https://doi.org/10.3390/life12101577
Chicago/Turabian StyleIevinsh, Gederts, Una Andersone-Ozola, and Astra Jēkabsone. 2022. "Similar Responses of Relatively Salt-Tolerant Plants to Na and K during Chloride Salinity: Comparison of Growth, Water Content and Ion Accumulation" Life 12, no. 10: 1577. https://doi.org/10.3390/life12101577