
Alleviate the Drought Stress on Triticum aestivum L. Using the Algal Extracts of Sargassum latifolium and Corallina elongate Versus the Commercial Algal Products
 , , , , and
, , , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material and Treatments
2.2. Preparation of Algal Extract
2.3. Treatments and Experimental Design
2.4. Growth Traits Measurement
2.4.1. Morphological Parameters Measurement
2.4.2. Photosynthetic Pigments Measurement
2.4.3. Total Soluble Carbohydrates and Proteins
2.4.4. Determination of Antioxidant Enzymes
2.4.5. Extraction and Determination of Endogenous Acidic Phytohormones
2.4.6. Yield Analysis
2.5. Statistical Analysis
3. Results and Discussion
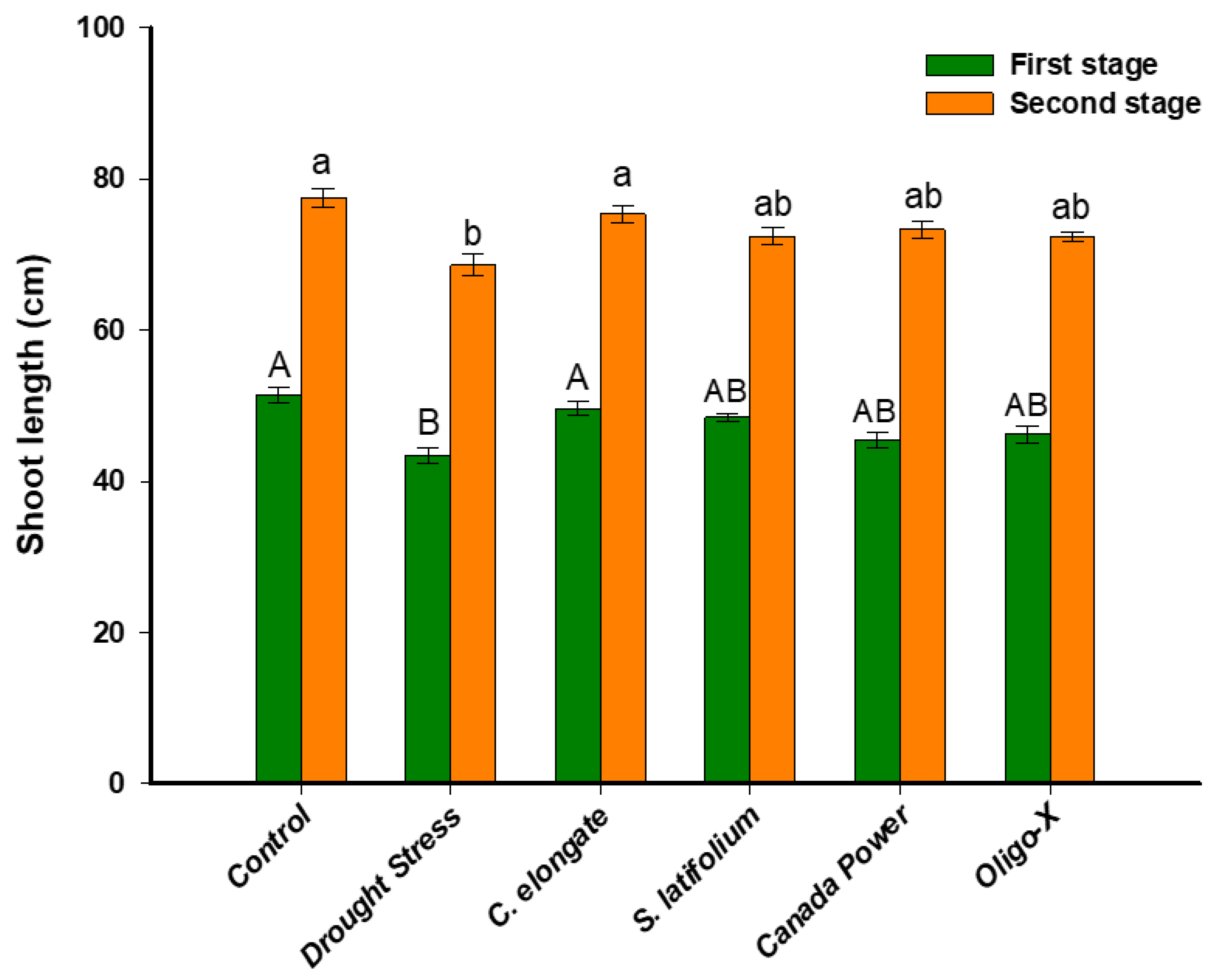
3.1. Effect of the Foliar Spray of Aqueous Algal Extracts on the Growth of Wheat Plants under Drought Stress
3.2. Effect of Algal Extracts on the Pigments Contents of Wheat Plants Grown under Drought Stress
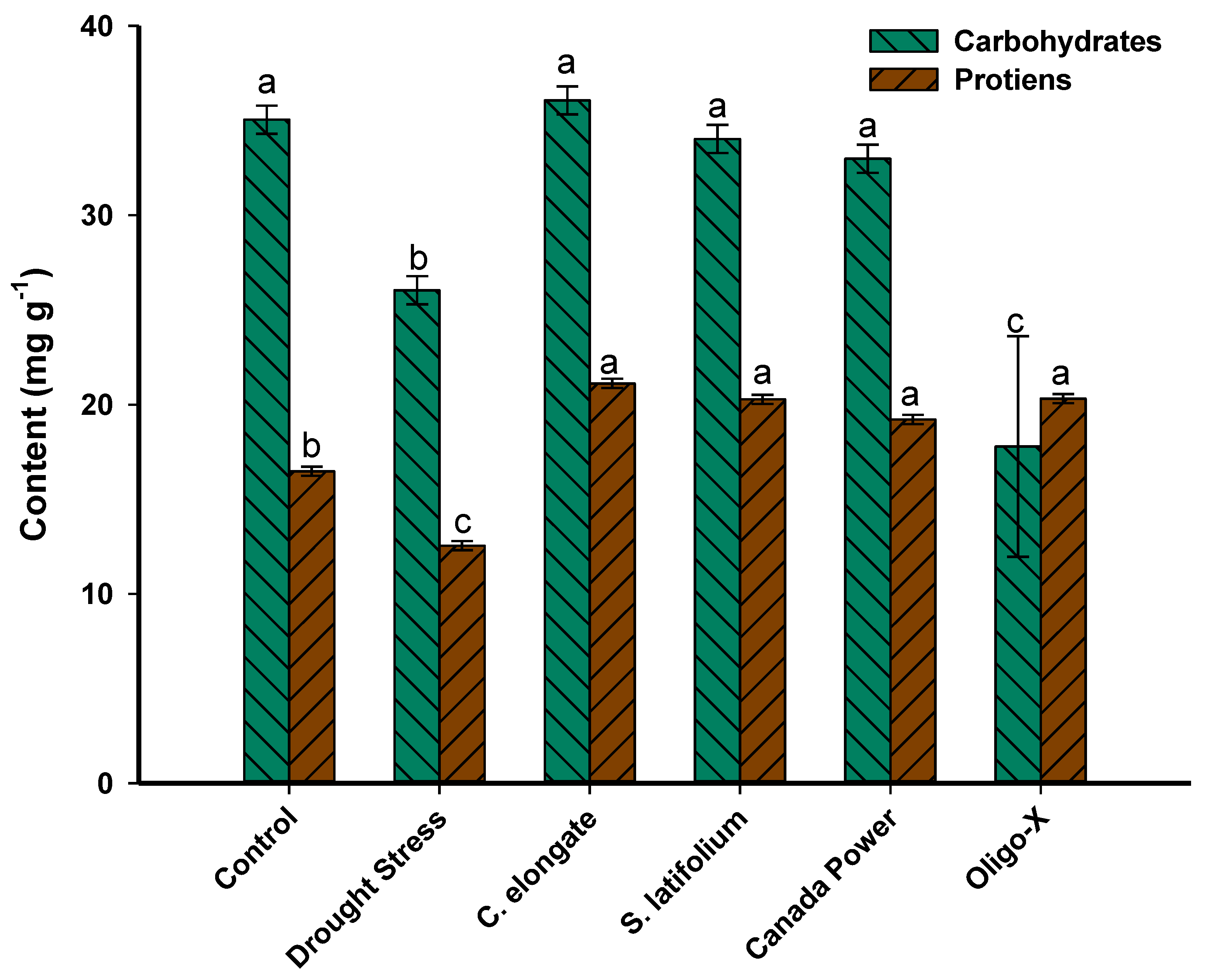
3.3. Effect of Foliar Spraying of the Algal Extract on the Carbohydrates and Protein Contents
3.4. Antioxidant Enzymatic Activity of Wheat Plant Grown under Drought Stress
3.5. Acidic Growth Hormones
3.6. Yield Parameter Measurement
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Li, W.; Wang, Y.; Zhang, Y.; Wang, R.; Guo, Z.; Xie, Z. Impacts of drought stress on the morphology, physiology, and sugar content of Lanzhou lily (Lilium davidii var. unicolor). Acta Physiol. Plant. 2020, 42, 127. [Google Scholar] [CrossRef]
- Shakeel, A.A.; Xiao-yu, X.; Long-chang, W.; Muhammad, F.S.; Chen, M.; Wang, L. Morphological, physiological and biochemical responses of plants to drought stress. Afr. J. Agric. Res. 2011, 6, 2026–2032. [Google Scholar]
- Yang, X.; Lu, M.; Wang, Y.; Wang, Y.; Liu, Z.; Chen, S. Response Mechanism of Plants to Drought Stress. Horticulturae 2021, 7, 50. [Google Scholar] [CrossRef]
- Singh, P.K.; Indoliya, Y.; Agrawal, L.; Awasthi, S.; Deeba, F.; Dwivedi, S.; Chakrabarty, D.; Shirke, P.A.; Pandey, V.; Singh, N.; et al. Genomic and proteomic responses to drought stress and biotechnological interventions for enhanced drought tolerance in plants. Curr. Plant Biol. 2022, 29, 100239. [Google Scholar] [CrossRef]
- Ullah, A.; Manghwar, H.; Shaban, M.; Khan, A.H.; Akbar, A.; Ali, U.; Ali, E.; Fahad, S. Phytohormones enhanced drought tolerance in plants: A coping strategy. Environ. Sci. Pollut. Res. Int. 2018, 25, 33103–33118. [Google Scholar] [CrossRef] [PubMed]
- Lau, S.-E.; Hamdan, M.F.; Pua, T.-L.; Saidi, N.B.; Tan, B.C. Plant Nitric Oxide Signaling under Drought Stress. Plants 2021, 10, 360. [Google Scholar] [CrossRef]
- Asseng, S.; Kheir, A.M.S.; Kassie, B.T.; Hoogenboom, G.; Abdelaal, A.I.N.; Haman, D.Z.; Ruane, A.C. Can Egypt become self-sufficient in wheat? Environ. Res. Lett. 2018, 13, 094012. [Google Scholar] [CrossRef] [Green Version]
- Shalaby, E.M.M.; Galall, E.H.; Ali, M.B.; Amro, A.; El Ramly, A. Growth and yield responses of ten wheat (Triticum aestivum L) genotypes to drought. SVU-Int. J. Agric. Sci. 2020, 2, 1–17. [Google Scholar] [CrossRef]
- Ullah, A.; Bano, A.; Khan, N. Climate Change and Salinity Effects on Crops and Chemical Communication Between Plants and Plant Growth-Promoting Microorganisms Under Stress. Front. Sustain. Food Syst. 2021, 5, 161. [Google Scholar] [CrossRef]
- Carillo, P.; Ciarmiello, L.F.; Woodrow, P.; Corrado, G.; Chiaiese, P.; Rouphael, Y. Enhancing Sustainability by Improving Plant Salt Tolerance through Macro-and Micro-Algal Biostimulants. Biology 2020, 9, 253. [Google Scholar] [CrossRef]
- Franzoni, G.; Cocetta, G.; Prinsi, B.; Ferrante, A.; Espen, L. Biostimulants on Crops: Their Impact under Abiotic Stress Conditions. Horticulturae 2022, 8, 189. [Google Scholar] [CrossRef]
- Ali, O.; Ramsubhag, A.; Jayaraman, J. Biostimulant Properties of Seaweed Extracts in Plants: Implications towards Sustainable Crop Production. Plants 2021, 10, 531. [Google Scholar] [CrossRef]
- Kumari, R.; Kaur, I.; Bhatnagar, A.K. Effect of aqueous extract of Sargassum johnstonii Setchell & Gardner on growth, yield and quality of Lycopersicon esculentum Mill. J. Appl. Phycol. 2011, 23, 623–633. [Google Scholar]
- El-Sheekh, M.M.; El-Saied, A.e.-D. Effect of crude seaweed extracts on seed germination, seedling growth and some metabolic processes of Vicia faba L. Cytobios 2000, 101, 23–35. [Google Scholar]
- Anisimov, M.; Chaikina, E.; Klykov, A.; Rasskazov, V. Effect of seaweeds extracts on the growth of seedling roots of buckwheat (Fagopyrum esculentum Moench) is depended on the season of algae collection. Agric. Sci. Dev. 2013, 2, 67–75. [Google Scholar]
- Si, Y.; Haxim, Y.; Wang, L. Optimum Sterilization Method for In Vitro Cultivation of Dimorphic Seeds of the Succulent Halophyte Suaeda aralocaspica. Horticulturae 2022, 8, 289. [Google Scholar] [CrossRef]
- Selim, M.T.; Salem, S.S.; Mohamed, A.A.; El-Gamal, M.S.; Awad, M.F.; Fouda, A. Biological Treatment of Real Textile Effluent Using Aspergillus flavus and Fusarium oxysporium and Their Consortium along with the Evaluation of Their Phytotoxicity. J. Fungi 2021, 7, 193. [Google Scholar] [CrossRef]
- Ismail, M.A.; Amin, M.A.; Eid, A.M.; Hassan, S.E.; Mahgoub, H.A.M.; Lashin, I.; Abdelwahab, A.T.; Azab, E.; Gobouri, A.A.; Elkelish, A.; et al. Comparative Study between Exogenously Applied Plant Growth Hormones versus Metabolites of Microbial Endophytes as Plant Growth-Promoting for Phaseolus vulgaris L. Cells 2021, 10, 1059. [Google Scholar] [CrossRef]
- Strain, H.H.; Svec, W.A. 2-Extraction, Separation, Estimation, and Isolation of the Chlorophylls**Based on work carried out under the auspices of the U.S. Atomic Energy Commission. In The Chlorophylls; Vernon, L.P., Seely, G.R., Eds.; Academic Press: London, UK, 1966; pp. 21–66. [Google Scholar]
- Lightenthaler, H. Chlorophylls and carotenoids: Pigments of photosynthetic biomembranes. Methods Enzymol. 1987, 148, 350–382. [Google Scholar]
- Umbreit, W.W.; Burris, R.H.; Stauffer, J.F. Manometric Techniques. A Manual Describing Methods Applicable to the Study of Tissue Metabolism; Burgess Publishing: Clayton, NC, USA, 1957. [Google Scholar]
- Lowry, O.H. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Mahgoub, H.A.M.; Fouda, A.; Eid, A.M.; Ewais, E.E.-D.; Hassan, S.E.-D. Biotechnological application of plant growth-promoting endophytic bacteria isolated from halophytic plants to ameliorate salinity tolerance of Vicia faba L. Plant Biotechnol. Rep. 2021, 15, 819–843. [Google Scholar] [CrossRef]
- Kar, M.; Mishra, D. Catalase, Peroxidase, and Polyphenoloxidase Activities during Rice Leaf Senescence. Plant Physiol. 1976, 57, 315–319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beyer, W.F.; Fridovich, I. Assaying for superoxide dismutase activity: Some large consequences of minor changes in conditions. Anal. Biochem. 1987, 161, 559–566. [Google Scholar] [CrossRef]
- Yang, R.; Yang, T.; Zhang, H.; Qi, Y.; Xing, Y.; Zhang, N.; Li, R.; Weeda, S.; Ren, S.; Ouyang, B.; et al. Hormone profiling and transcription analysis reveal a major role of ABA in tomato salt tolerance. Plant Physiol. Biochem. 2014, 77, 23–34. [Google Scholar] [CrossRef] [PubMed]
- Badawy, A.A.; Abdelfattah, N.A.H.; Salem, S.S.; Awad, M.F.; Fouda, A. Efficacy Assessment of Biosynthesized Copper Oxide Nanoparticles (CuO-NPs) on Stored Grain Insects and Their Impacts on Morphological and Physiological Traits of Wheat (Triticum aestivum L.) Plant. Biology 2021, 10, 233. [Google Scholar] [CrossRef]
- Sarkar, D.; Mandal, B.; Kundu, M.C. Increasing use efficiency of boron fertilisers by rescheduling the time and methods of application for crops in India. Plant Soil 2007, 301, 77–85. [Google Scholar] [CrossRef]
- Zhu, J.K. Salt and drought stress signal transduction in plants. Annu. Rev. Plant Biol. 2002, 53, 247–273. [Google Scholar] [CrossRef] [Green Version]
- Agnew, C.; Warren, A. A framework for tackling drought and land degradation. J. Arid. Environ. 1996, 33, 309–320. [Google Scholar] [CrossRef]
- Bassal, S.; Zahran, F. Effect of farmyard manure, bio and mineral nitrogen fertilizer and hill spaces on rice crop productivity. J. Plant Prod. 2002, 27, 1975–1988. [Google Scholar] [CrossRef]
- Huang, J.; Hirji, R.; Adam, L.; Rozwadowski, K.L.; Hammerlindl, J.K.; Keller, W.A.; Selvaraj, G. Genetic engineering of glycinebetaine production toward enhancing stress tolerance in plants: Metabolic limitations. Plant Physiol. 2000, 122, 747–756. [Google Scholar] [CrossRef] [Green Version]
- Ashraf, M.; Harris, P.J.C. Potential biochemical indicators of salinity tolerance in plants. Plant Sci. 2004, 166, 3–16. [Google Scholar] [CrossRef]
- van Staden, J.; Beckett, R.P.; Rijkenberg, M.J. Effect of seaweed concentrate on the growth of the seedlings of three species of Eucalyptus. S. Afr. J. Bot. 1995, 61, 169–172. [Google Scholar] [CrossRef] [Green Version]
- Crouch, I.J.; Smith, M.T.; van Staden, J.; Lewis, M.J.; Hoad, G.V. Identification of Auxins in a Commercial Seaweed Concentrate. J. Plant Physiol. 1992, 139, 590–594. [Google Scholar] [CrossRef]
- Ahmed, F.; Baloch, D.; Sadiq, S.; Ahmed, S.; Hanan, A.; Taran, S.; Ahmed, N.; Hassan, M. Plant growth regulators induced drought tolerance in sunflower (Helianthus annuus L.) hybrids. JAPS J. Anim. Plant Sci. 2014, 24, 886–890. [Google Scholar]
- Fouda, A.; Eid, A.M.; Elsaied, A.; El-Belely, E.F.; Barghoth, M.G.; Azab, E.; Gobouri, A.A.; Hassan, S.E. Plant Growth-Promoting Endophytic Bacterial Community Inhabiting the Leaves of Pulicaria incisa (Lam.) DC Inherent to Arid Regions. Plants 2021, 10, 76. [Google Scholar] [CrossRef]
- Mansori, M.; Chernane, H.; Latique, S.; Benaliat, A.; Hsissou, D.; El Kaoua, M. Seaweed extract effect on water deficit and antioxidative mechanisms in bean plants (Phaseolus vulgaris L.). J. Appl. Phycol. 2015, 27, 1689–1698. [Google Scholar] [CrossRef]
- Briceño-Domínguez, D.; Hernández-Carmona, G.; Moyo, M.; Stirk, W.; van Staden, J. Plant growth promoting activity of seaweed liquid extracts produced from Macrocystis pyrifera under different pH and temperature conditions. J. Appl. Phycol. 2014, 26, 2203–2210. [Google Scholar] [CrossRef]
- dos Santos, T.B.; Ribas, A.F.; de Souza, S.G.H.; Budzinski, I.G.F.; Domingues, D.S. Physiological Responses to Drought, Salinity, and Heat Stress in Plants: A Review. Stresses 2022, 2, 113–135. [Google Scholar] [CrossRef]
- Eissa, M.A.; Roshdy, N.M.K. Nitrogen fertilization: Effect on Cd-phytoextraction by the halophytic plant quail bush [Atriplex lentiformis (Torr.) S. Wats]. S. Afr. J. Bot. 2018, 115, 126–131. [Google Scholar] [CrossRef]
- Ramya, S.S.; Vijayanand, N.; Rathinavel, S. Foliar application of liquid biofertilizer of brown alga Stoechospermum marginatum on growth, biochemical and yield of Solanum melongena. Int. J. Recycl. Org. Waste Agric. 2015, 4, 167–173. [Google Scholar] [CrossRef] [Green Version]
- Khan, W.; Rayirath, U.P.; Subramanian, S.; Jithesh, M.N.; Rayorath, P.; Hodges, D.M.; Critchley, A.T.; Craigie, J.S.; Norrie, J.; Prithiviraj, B. Seaweed Extracts as Biostimulants of Plant Growth and Development. J. Plant Growth Regul. 2009, 28, 386–399. [Google Scholar] [CrossRef]
- Döring, J.; Frisch, M.; Tittmann, S.; Stoll, M.; Kauer, R. Growth, yield and fruit quality of grapevines under organic and biodynamic management. PLoS ONE 2015, 10, e0138445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ali, Q.; Ashraf, M.; Anwar, F. Seed Composition and Seed Oil Antioxidant Activity of Maize Under Water Stress. J. Am. Oil Chem. Soc. 2010, 87, 1179–1187. [Google Scholar] [CrossRef]
- Manzoor, K.; Ilyas, N.; Batool, N.; Ahmad, B.; Arshad, M. Effect of Salicylic Acid on the Growth and Physiological Characteristics of Maize under Stress Conditions. J. Chem. Soc. Pak. 2015, 37, 588–593. [Google Scholar]
- Noreen, S.; Fatima, K.; Athar, H.; Ahmad, S.; Hussain, K. Enhancement of physio-biochemical parameters of wheat through exogenous application of salicylic acid under drought stress. JAPS: J. Anim. Plant Sci. 2017, 27, 153–163. [Google Scholar]
- El-Sayed, S.; Hellal, F.; Nofal, O.; El-Karamany, M.; Bakry, B. Influence of algal extracts on yield and chemical composition of moringa and alfalfa grown under drought condition. Int. J. Environ. 2015, 4, 151–157. [Google Scholar]
- Fouda, A.; Eid, A.M.; Abdelkareem, A.; Said, H.A.; El-Belely, E.F.; Alkhalifah, D.H.M.; Alshallash, K.S.; Hassan, S.E.-D. Phyco-Synthesized Zinc Oxide Nanoparticles Using Marine Macroalgae, Ulva fasciata Delile, Characterization, Antibacterial Activity, Photocatalysis, and Tanning Wastewater Treatment. Catalysts 2022, 12, 756. [Google Scholar] [CrossRef]
- Zewail, R. Effect of seaweed extract and amino acids on growth and productivity and some biocostituents of common bean (Phaseolus vulgaris L) plants. J. Plant Prod. 2014, 5, 1441–1453. [Google Scholar] [CrossRef] [Green Version]
- Fike, J.H.; Allen, V.G.; Schmidt, R.E.; Zhang, X.; Fontenot, J.P.; Bagley, C.P.; Ivy, R.L.; Evans, R.R.; Coelho, R.W.; Wester, D.B. Tasco-Forage: I. Influence of a seaweed extract on antioxidant activity in tall fescue and in ruminants. J. Anim. Sci. 2001, 79, 1011–1021. [Google Scholar] [CrossRef]
- Ayad, J. The Effect of Seaweed Extract (Ascophyllum nodosum) on Antioxidant Activities and Drought Tolerance of Tall Fescue (Festuca arundinacea Schreb). Ph.D. Thesis, Texas Tech University, Lubbock, TX, USA, 1998. [Google Scholar]
- O’Sullivan, A.M.; O’Callaghan, Y.C.; O’Grady, M.N.; Queguineur, B.; Hanniffy, D.; Troy, D.J.; Kerry, J.P.; O’Brien, N.M. In vitro and cellular antioxidant activities of seaweed extracts prepared from five brown seaweeds harvested in spring from the west coast of Ireland. Food Chem. 2011, 126, 1064–1070. [Google Scholar] [CrossRef]
- Eid, A.M.; Fouda, A.; Abdel-Rahman, M.A.; Salem, S.S.; Elsaied, A.; Oelmüller, R.; Hijri, M.; Bhowmik, A.; Elkelish, A.; Hassan, S.E. Harnessing Bacterial Endophytes for Promotion of Plant Growth and Biotechnological Applications: An Overview. Plants 2021, 10, 935. [Google Scholar] [CrossRef]
- Grace, S.C. Phenolics as antioxidants. In Antioxidants and Reactive Oxygen Species in Plants; John Wiley & Son: Hoboken, NJ, USA, 2005; pp. 141–168. [Google Scholar]
- Kurutas, E.B. The importance of antioxidants which play the role in cellular response against oxidative/nitrosative stress: Current state. Nutr. J. 2016, 15, 71. [Google Scholar] [CrossRef] [Green Version]
- El-Sersawy, M.M.; Hassan, S.E.; El-Ghamry, A.A.; El-Gwad, A.M.A.; Fouda, A. Implication of plant growth-promoting rhizobacteria of Bacillus spp. as biocontrol agents against wilt disease caused by Fusarium oxysporum Schlecht. in Vicia faba L. Biomol. Concepts 2021, 12, 197–214. [Google Scholar] [CrossRef]
- Kazan, K. Auxin and the integration of environmental signals into plant root development. Ann. Bot. 2013, 112, 1655–1665. [Google Scholar] [CrossRef] [Green Version]
- Abdel-Hamid, M.S.; Fouda, A.; El-Ela, H.K.A.; El-Ghamry, A.A.; Hassan, S.E. Plant growth-promoting properties of bacterial endophytes isolated from roots of Thymus vulgaris L. and investigate their role as biofertilizers to enhance the essential oil contents. Biomol. Concepts 2021, 12, 175–196. [Google Scholar] [CrossRef]
- Iqbal, S.; Wang, X.; Mubeen, I.; Kamran, M.; Kanwal, I.; Díaz, G.A.; Abbas, A.; Parveen, A.; Atiq, M.N.; Alshaya, H.; et al. Phytohormones Trigger Drought Tolerance in Crop Plants: Outlook and Future Perspectives. Front. Plant Sci. 2021, 12, 799318. [Google Scholar] [CrossRef]
- Colebrook, E.H.; Thomas, S.G.; Phillips, A.L.; Hedden, P. The role of gibberellin signalling in plant responses to abiotic stress. J. Exp. Biol. 2014, 217, 67–75. [Google Scholar] [CrossRef] [Green Version]
- Achard, P.; Genschik, P. Releasing the brakes of plant growth: How GAs shutdown DELLA proteins. J. Exp. Bot. 2009, 60, 1085–1092. [Google Scholar] [CrossRef] [Green Version]
- Kaya, C.; Levent Tuna, A.; Alfredo, A.C.A. Gibberellic acid improves water deficit tolerance in maize plants. Acta Physiol. Plant. 2006, 28, 331–337. [Google Scholar] [CrossRef]
- Salih, S.; Tuncturk, R. Low doses of gibberellic acid can enhance germination of wheat seed under drought stress. Adv. Crop. Sci. Tech. 2020, 8, 432. [Google Scholar]
- Islam, M.R.; Feng, B.; Chen, T.; Tao, L.; Fu, G. Role of Abscisic Acid in Thermal Acclimation of Plants. J. Plant Biol. 2018, 61, 255–264. [Google Scholar] [CrossRef]
- Qi, J.; Song, C.P.; Wang, B.; Zhou, J.; Kangasjärvi, J.; Zhu, J.K.; Gong, Z. Reactive oxygen species signaling and stomatal movement in plant responses to drought stress and pathogen attack. J. Integr. Plant Biol. 2018, 60, 805–826. [Google Scholar] [CrossRef] [Green Version]
- Chhaya; Yadav, B.; Jogawat, A.; Gnanasekaran, P.; Kumari, P.; Lakra, N.; Lal, S.K.; Pawar, J.; Narayan, O.P. An overview of recent advancement in phytohormones-mediated stress management and drought tolerance in crop plants. Plant Gene 2021, 25, 100264. [Google Scholar] [CrossRef]
- Hernández-Herrera, R.M.; Santacruz-Ruvalcaba, F.; Ruiz-López, M.A.; Norrie, J.; Hernández-Carmona, G. Effect of liquid seaweed extracts on growth of tomato seedlings (Solanum lycopersicum L.). J. Appl. Phycol. 2014, 26, 619–628. [Google Scholar] [CrossRef]
- Almaroai, Y.A.; Eissa, M.A. Role of Marine Algae Extracts in Water Stress Resistance of Onion Under Semiarid Conditions. J. Soil Sci. Plant Nutr. 2020, 20, 1092–1101. [Google Scholar] [CrossRef]
- Shukla, P.S.; Shotton, K.; Norman, E.; Neily, W.; Critchley, A.T.; Prithiviraj, B. Seaweed extract improve drought tolerance of soybean by regulating stress-response genes. AoB Plants 2018, 10, plx051. [Google Scholar] [CrossRef]
- Elewa, T.A.; Sadak, M.S.; Dawood, M.G. Improving drought tolerance of quinoa plant by foliar treatment of trehalose. Agric. Eng. Int. CIGR J. 2017, 245–254. [Google Scholar]
- Agami, R.A. Salicylic acid mitigates the adverse effect of water stress on lettuce (Lactuca sativa L.). J. Appl. Sci. Res. 2013, 9, 5701–5711. [Google Scholar]
- El-Saadony, F.; Nawar, D.A.; Zyada, H. Effect of foliar application with salicylic acid, garlic extract and proline on growth, yield and leaf anatomy of pea (Pisum sativum L.) grown under drought stress. Middle East J. Appl. Sci. 2017, 7, 633–650. [Google Scholar]
- El-Beltagi, H.S.; Mohamed, A.A.; Mohamed, H.I.; Ramadan, K.M.A.; Barqawi, A.A.; Mansour, A.T. Phytochemical and Potential Properties of Seaweeds and Their Recent Applications: A Review. Mar. Drugs 2022, 20, 342. [Google Scholar] [CrossRef]
- Djapic, N. Corallina officinalis chemical compounds obtained by supercritical fluid extraction. Aquac. Aquar. Conserv. Legis. 2018, 11, 422–428. [Google Scholar]



{kind=link}
{kind=link}
| Treatments | Shoot Fresh Weight (g) | Shoot Dry Weight (g) | Water Shoot Content (%) | |||
|---|---|---|---|---|---|---|
| First Stage | Second Stage | First Stage | Second Stage | First Stage | Second Stage | |
| Control | 5.5 ± 0.7 a | 9.1 ± 0.8 a | 1.6 ± 0.02 a | 2.1 ± 0.3 a | 71.1 ± 0.7 c | 76.5 ± 0.8 a |
| Drought Stress | 4.04 ± 0.4 b | 7.9 ± 0.4 b | 0.90 ± 0.001 b | 1.8 ± 0.2 a | 75.6 ± 0.2 b | 77.2 ± 0.5 a |
| C. elongate | 5.7 ± 0.2 a | 9.1 ± 0.7 a | 1.6 ± 0.1 a | 2.1 ± 0.2 a | 71.5 ± 0.4 c | 77.2 ± 0.4 a |
| S. latifolium | 4.7 ± 0.2 ab | 8.2 ± 0.4 b | 1.5 ± 0.02 a | 1.9 ± 0.2 a | 66.4 ± 1.8 d | 76.4 ± 0.5 a |
| Canada Power | 4.9 ± 0.3 ab | 7.9 ± 0.4 b | 1.1 ± 0.05 ab | 1.9 ± 0.2 a | 78.2 ± 1.4 a | 76.6 ± 0.9 a |
| Oligo-X | 4.2 ± 0.2 b | 8.04 ± 0.3 b | 1.1 ± 0.1 ab | 1.9 ± 0.1 a | 72.9 ± 0.6 c | 76.1 ± 0.1 a |
| Treatments | Chlorophyll (a) | Chlorophyll (b) | Chlorophyll (a + b) | Carotenoids | ||||
|---|---|---|---|---|---|---|---|---|
| First Stage | Second Stage | First Stage | Second Stage | First Stage | Second Stage | First Stage | Second Stage | |
| Control | 6.8 ± 1.9 a | 7.4 ± 0.6 a | 5.3 ± 1.8 a | 9.1 ± 1.6 a | 12.1 ± 3.7 a | 16.5 ± 1.1 a | 2.8 ± 0.8 a | 1.3 ± 0.33 b |
| Drought Stress | 3.1 ± 1.5 b | 5.7 ± 0.1 c | 4.2 ± 2.9 b | 7.5 ± 0.5 b | 7.3 ± 1.4 c | 13.2± 0.4 b | 2.4 ± 0.5 a | 1.1 ± 0.13 b |
| C.elongate | 5.9 ± 1.8 a | 7.5 ± 0.5 a | 5.3 ± 2.7 a | 9.5 ± 1.6 a | 11.2 ± 1.7 a | 17.1 ± 1.04 a | 2.1 ± 1.6 a | 1.01 ± 0.3 b |
| S. latifolium | 4.2 ± 0.1 ab | 6.7 ± 0.02 b | 4.9 ± 0.1 ab | 9.9 ± 0.75 a | 9.1 ± 0.04 b | 16.7 ± 0.8 a | 1.0 ± 0.6 b | 1.2 ± 0.5 a |
| Canada Power | 3.8 ± 2.6 b | 6.6 ± 0.02 b | 4.3 ± 3.3 b | 10.2 ± 0.1 a | 8.1 ± 5.9 c | 16.8 ± 0.2 a | 1.5 ± 0.3 ab | 1.5 ± 0.2 ab |
| Oligo-X | 3.4 ± 1.4 b | 6.6 ± 0.03 b | 4.4 ± 1.01 b | 11.9 ± 1.1 a | 7.8 ± 2.5 c | 18.5 ± 1.1 a | 0.8 ± 0.5 b | 1.9± 0.7 a |
| Treatments | Carbohydrate Shoot (mg g−1 DW) | Protein Shoot (mg g−1 DW) | ||
|---|---|---|---|---|
| First Stage | Second Stage | First Stage | Second Stage | |
| Control | 13.3 ± 0.4 ab | 11.7 ± 0.4 a | 4.9 ± 0.2 b | 6.4 ± 0.1 a |
| Drought Stress | 10.4 ± 0.4 b | 9.8 ± 0.1 a | 3.8 ± 0.1 c | 4.6 ± 0.1 b |
| C. elongate | 15.9 ± 0.1 a | 12.7 ± 0.1 a | 7.9 ± 0.3 a | 7.6 ± 0.3 a |
| S. latifolium | 12.9 ± 0.1 b | 12.5 ± 0.1 a | 6.7 ± 0.1 a | 6.9 ± 0.1 a |
| Canada Power | 14.0 ± 0.2 a | 12.5 ± 0.2 a | 5.6 ± 0.2 b | 6.7 ± 0.1 a |
| Oligo-X | 15.0 ± 0.3 a | 12.1 ± 0.4 a | 5.9 ± 0.1 ab | 7.0 ± 0.1 a |
| Treatments | Peroxidase (Unit/µg FW) | Superoxide Dismutase (Unit/µg FW) | Polyphenol Oxidase (Unit/µg FW) | |||
|---|---|---|---|---|---|---|
| First Stage | Second Stage | First Stage | Second Stage | First Stage | Second Stage | |
| Control | 76.5 ± 7.5 b | 144 ± 12.0 b | 36 ± 12.0 e | 180 ± 8.4 e | 1.2 ± 0.6 b | 13.2 ± 0.6 b |
| Drought Stress | 86.5 ± 8.5 a | 165.5 ± 8.5 a | 60 ± 12.0 a | 300 ± 4.2 a | 2.4 ± 1.2 a | 16.2 ± 1.8 a |
| C. elongate | 45 ± 5.2 e | 140.5 ± 3.5 c | 50 ± 9.3 b | 174 ± 13.8 e | 2.1 ± 1.5 a | 6.6 ± 1.21 d |
| S. latifolium | 56.5 ± 2.3 d | 106.5 ± 13.5 e | 46 ± 9.9 c | 240 ± 4.8 b | 1.5 ± 3.3 ab | 10.6 ± 2.12 c |
| Canada Power | 55.5 ± 3.3 d | 138 ± 18.0 c | 41 ± 6.0 d | 219 ± 6.9 d | 1.25 ± 0.3 b | 14.3 ± 2.25 ab |
| Oligo-X | 65.5 ± 2.3 c | 124.5 ± 7.5 d | 39 ± 3.6 de | 228 ± 2.4 c | 2.1 ± 1.2 a | 8.4 ± 1.62 d |
| Treatment | IAA (mg/100 g) | GA3 (mg/100 g) | ABA (µg/100 g) |
|---|---|---|---|
| Control | 0.4 ± 0.02 b | 9.6 ± 0.03 a | 0.5 ± 0.01 b |
| Drought Stress | 0.3 ± 0.05 b | 7.5 ± 0.5 b | 0.9 ± 0.01 a |
| C. elongate | 1.1 ± 0.1 a | 9.4 ± 0.4 a | 0.4 ± 0.03 b |
| S. latifolium | 0.6 ± 0.05 b | 8.9 ± 0.3 ab | 0.6 ± 0.05 ab |
| Canada Power | 0.5 ± 0.05 b | 8.7 ± 0.4 ab | 0.3 ± 0.08 b |
| Oligo-X | 0.4 ± 0.01 b | 8.8 ± 0.1 ab | 0.3 ± 0.09 b |
| Treatment | Wt. of the Spike (g) | Spike Length (cm) | No. of Grains/Spike | Wt. Grains (g)/Spike |
|---|---|---|---|---|
| Control | 3.4 ± 0.4 a | 11.7 ± 0.4 a | 49.9 ± 4.8 b | 1.6 ± 0.3 a |
| Drought Stress | 1.6 ± 0.2 b | 8.9 ± 0.4 b | 38.7 ± 3.3 c | 0.9 ± 0.1 b |
| C. elongate | 3.2 ± 0.3 a | 11.6 ± 0.4 a | 54.6 ± 4.2 a | 1.8 ± 0.1 a |
| S. latifolium | 3.1 ± 0.2 a | 11.3 ± 0.5 a | 47.7 ± 3.03 b | 1.6 ± 0.2 a |
| Canada Power | 2.9 ± 0.2 a | 11.1 ± 0.4 a | 46.4 ± 2.6 b | 1.6 ± 0.1 a |
| Oligo-X | 2.8 ± 0.2 a | 11.1 ± 0.6 a | 45.7 ± 2.9 b | 1.5 ± 0.2 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alharbi, K.; Amin, M.A.; Ismail, M.A.; Ibrahim, M.T.S.; Hassan, S.E.-D.; Fouda, A.; Eid, A.M.; Said, H.A. Alleviate the Drought Stress on Triticum aestivum L. Using the Algal Extracts of Sargassum latifolium and Corallina elongate Versus the Commercial Algal Products. Life 2022, 12, 1757. https://doi.org/10.3390/life12111757
Alharbi K, Amin MA, Ismail MA, Ibrahim MTS, Hassan SE-D, Fouda A, Eid AM, Said HA. Alleviate the Drought Stress on Triticum aestivum L. Using the Algal Extracts of Sargassum latifolium and Corallina elongate Versus the Commercial Algal Products. Life. 2022; 12(11):1757. https://doi.org/10.3390/life12111757
Chicago/Turabian StyleAlharbi, Khadiga, Mohamed A. Amin, Mohamed A. Ismail, Mariam T. S. Ibrahim, Saad El-Din Hassan, Amr Fouda, Ahmed M. Eid, and Hanan A. Said. 2022. "Alleviate the Drought Stress on Triticum aestivum L. Using the Algal Extracts of Sargassum latifolium and Corallina elongate Versus the Commercial Algal Products" Life 12, no. 11: 1757. https://doi.org/10.3390/life12111757