
Salinity and Heavy Metal Tolerance, and Phytoextraction Potential of Ranunculus sceleratus Plants from a Sandy Coastal Beach
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Experiments
2.2. Plant Establishment and Cultivation
2.3. Treatments
2.4. Termination of Experiments and Measurements
2.5. Data Analysis
3. Results
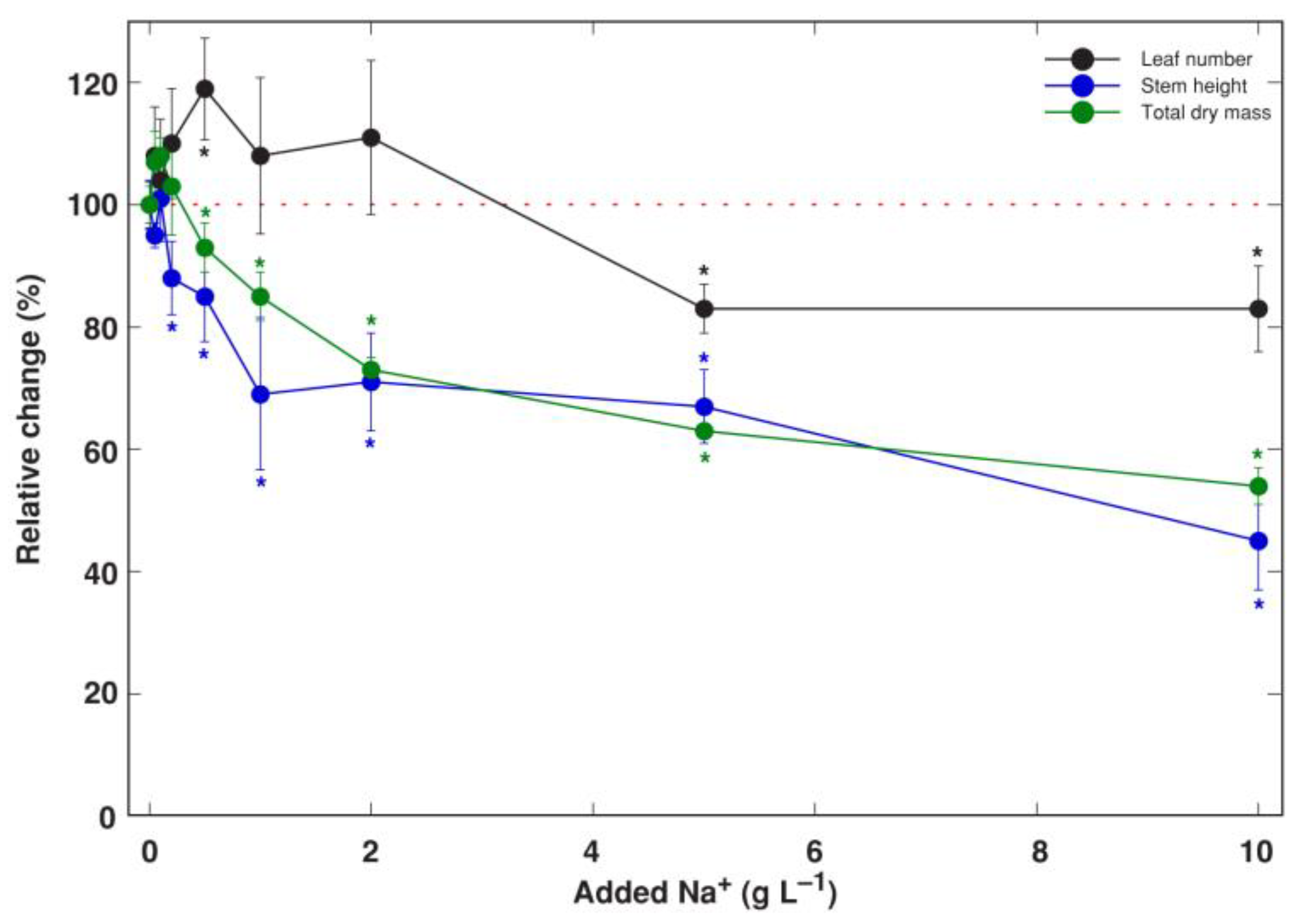
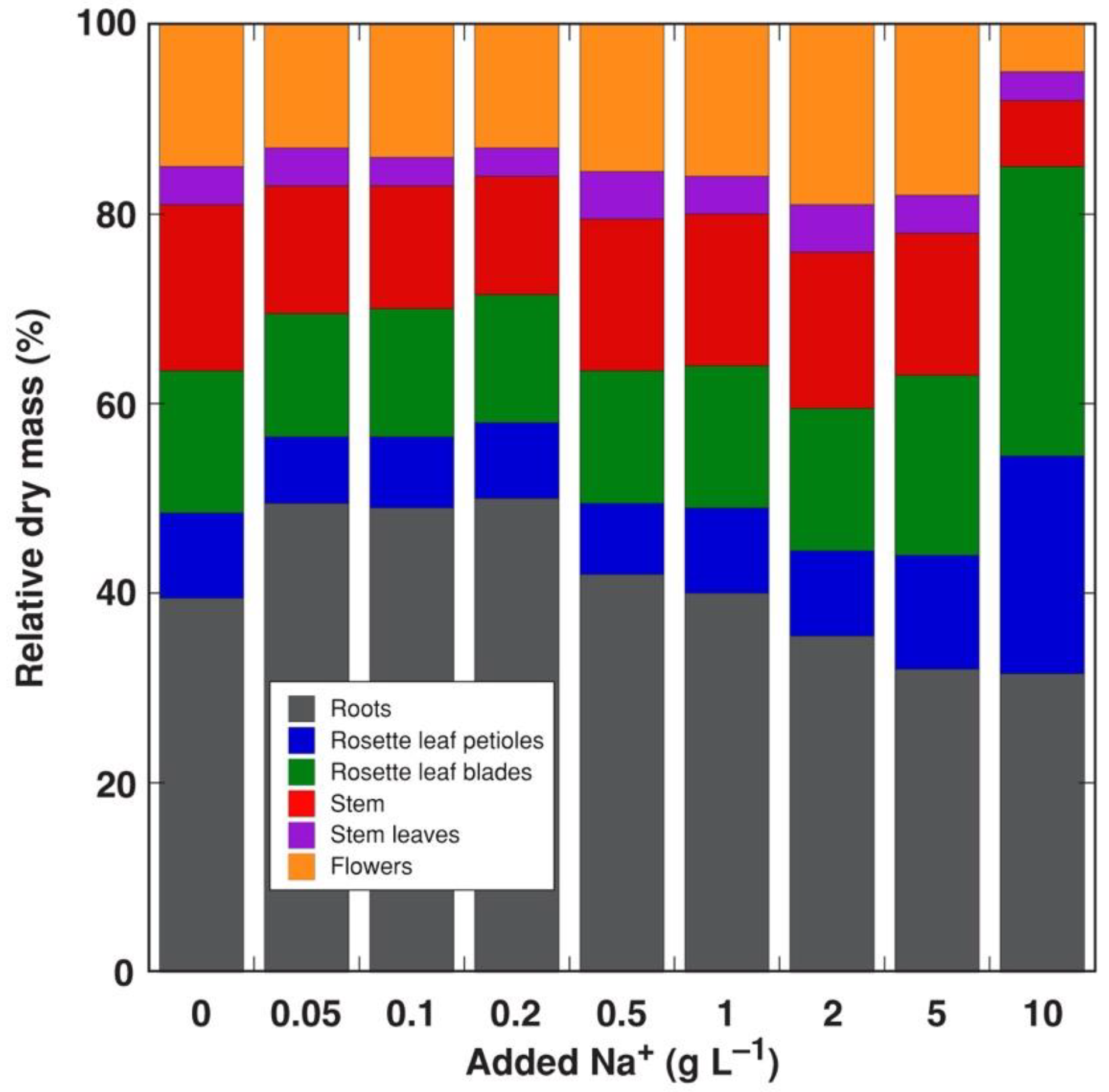
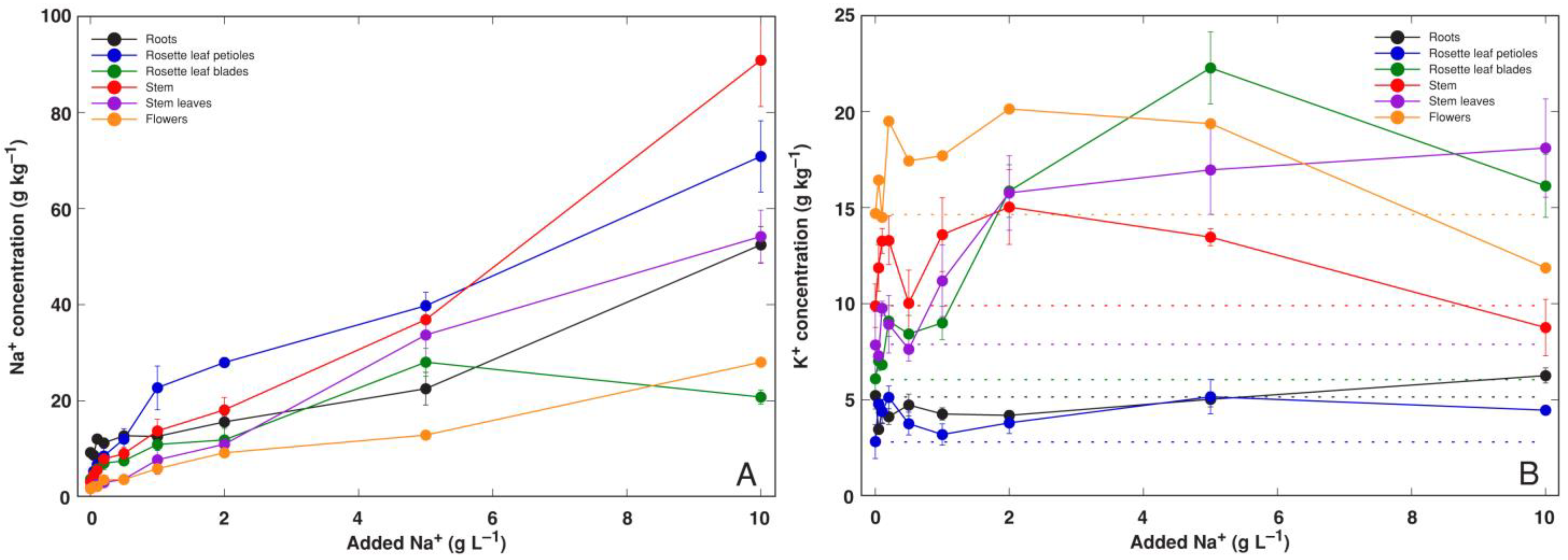
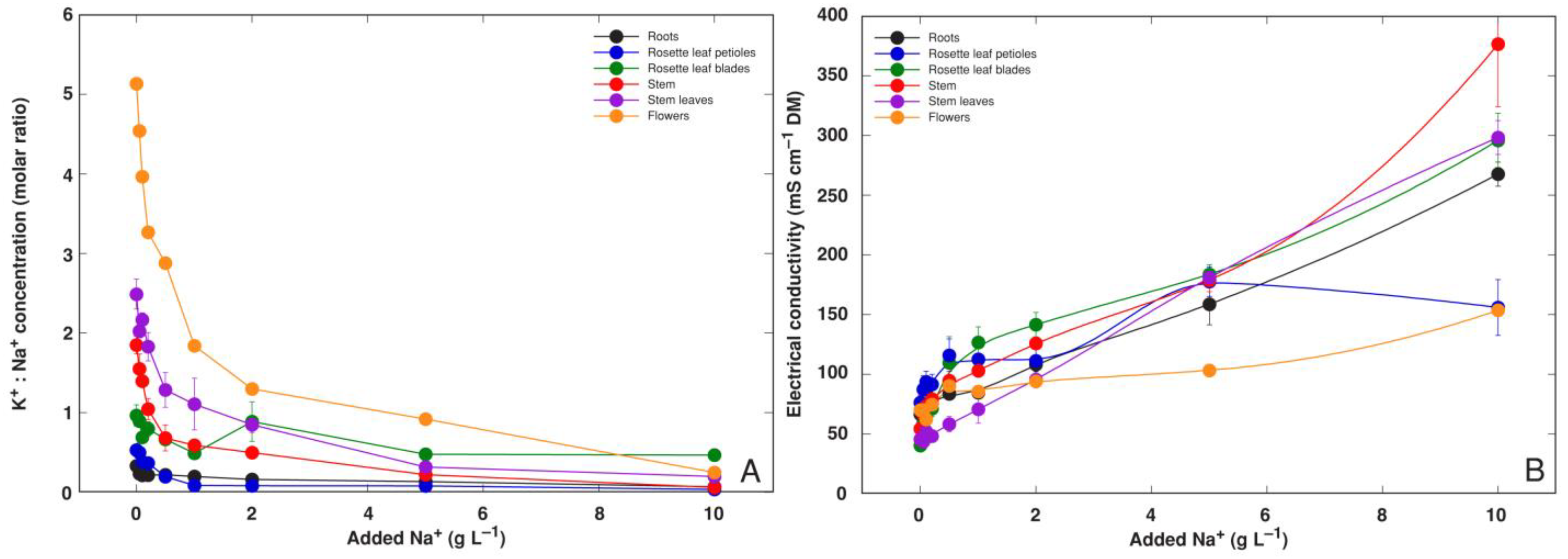
3.1. Experiment 1: NaCl Gradient
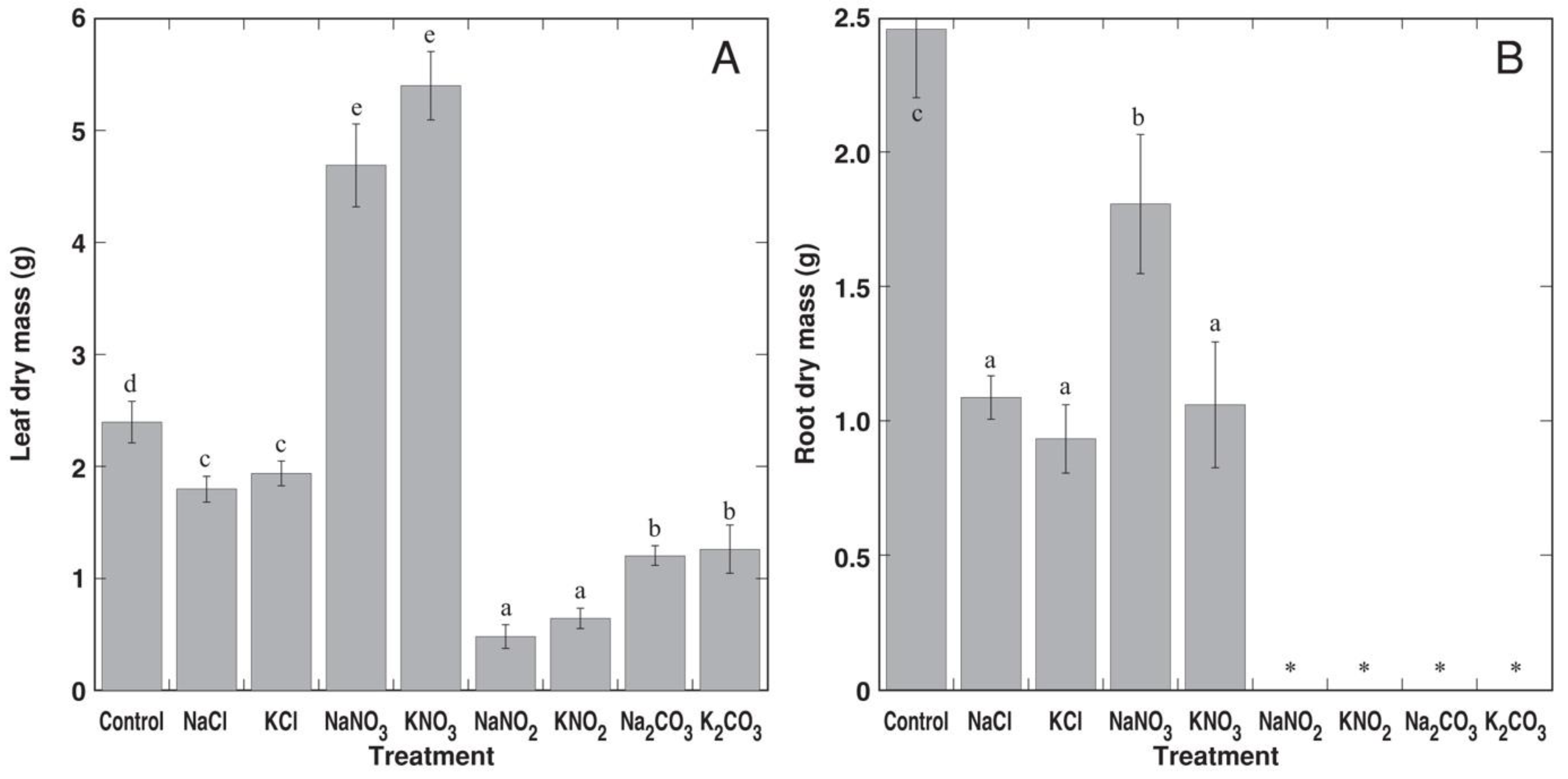
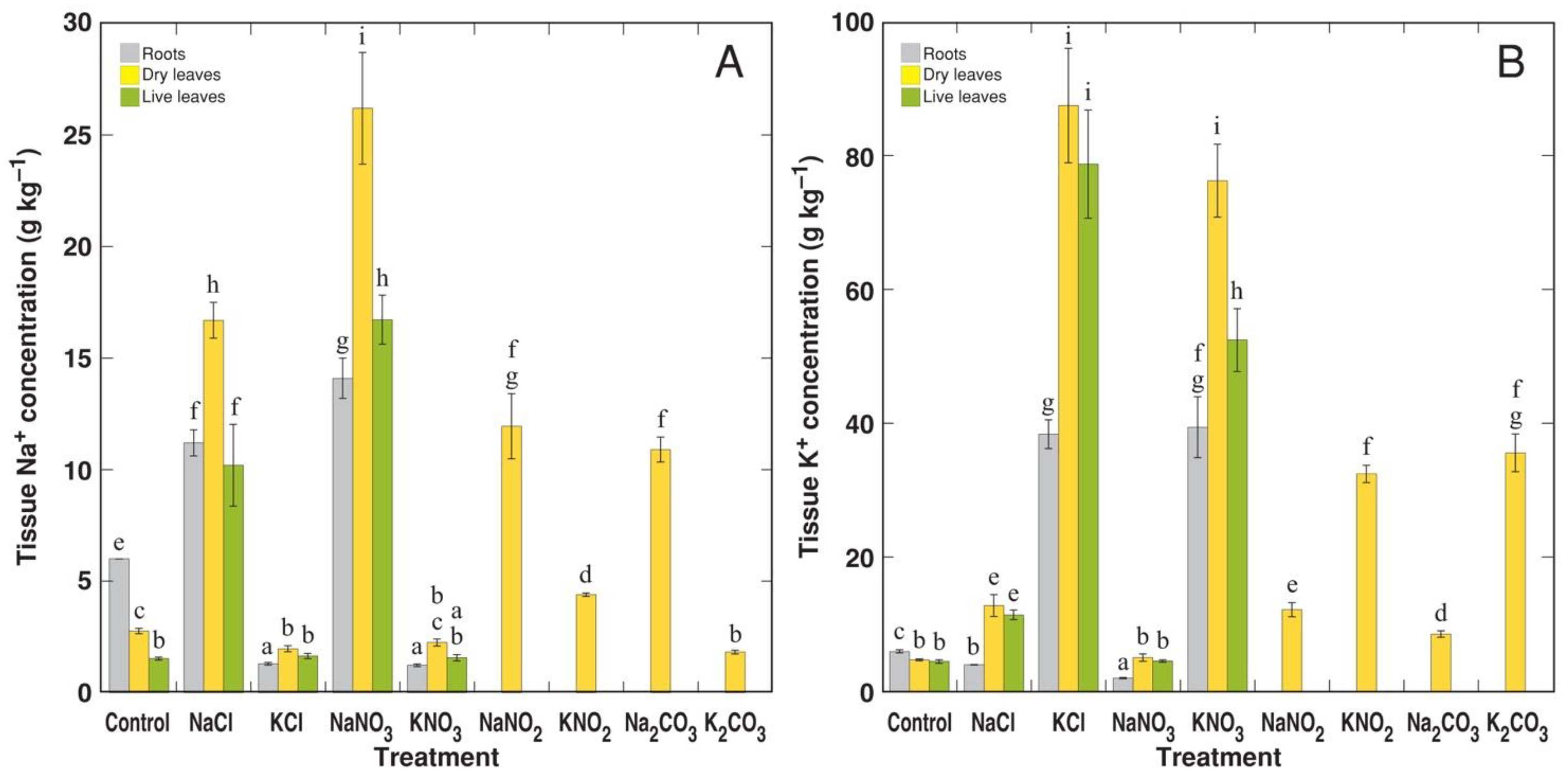
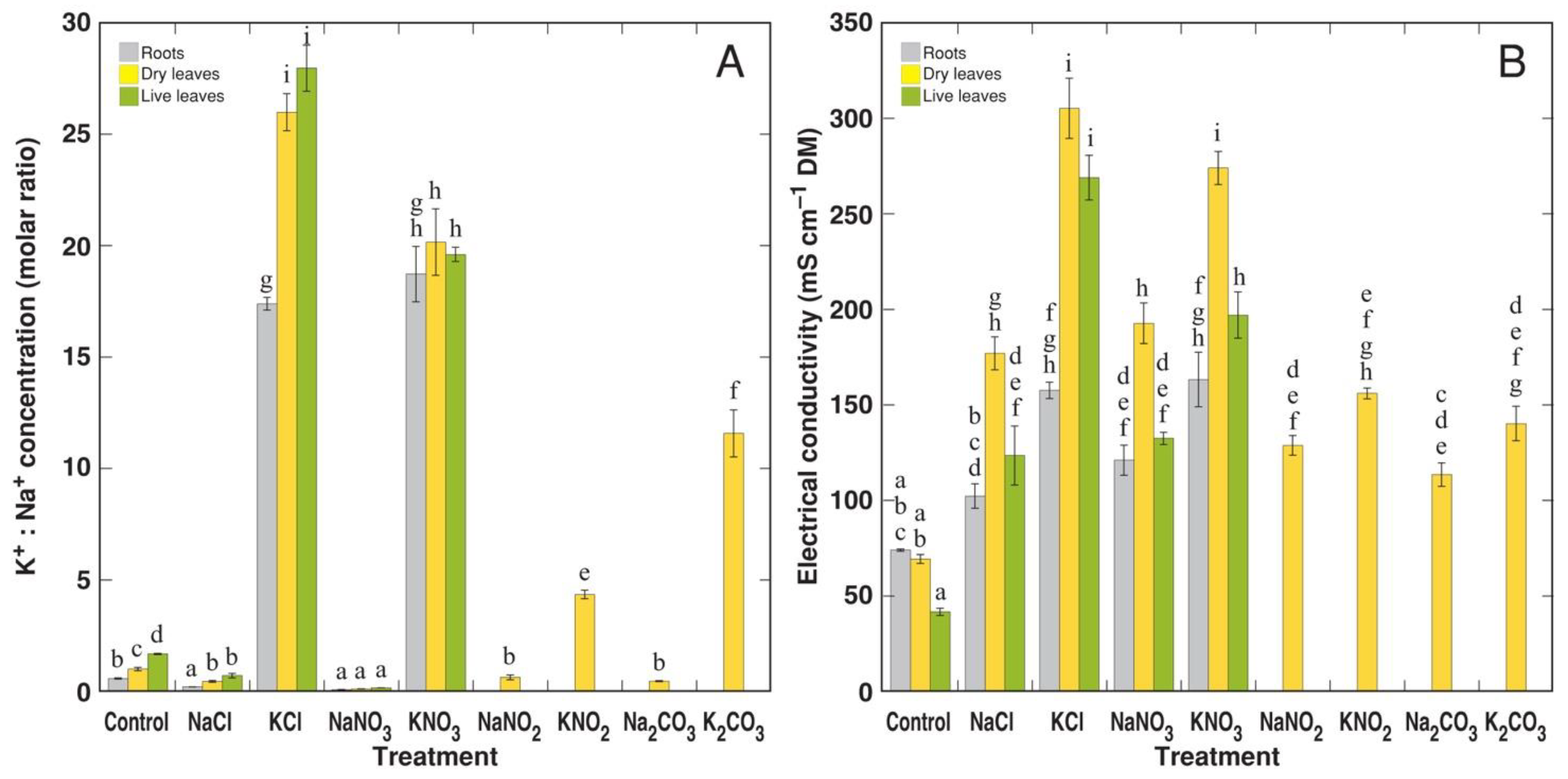
3.2. Experiment 2: Na+ and K+ Salts
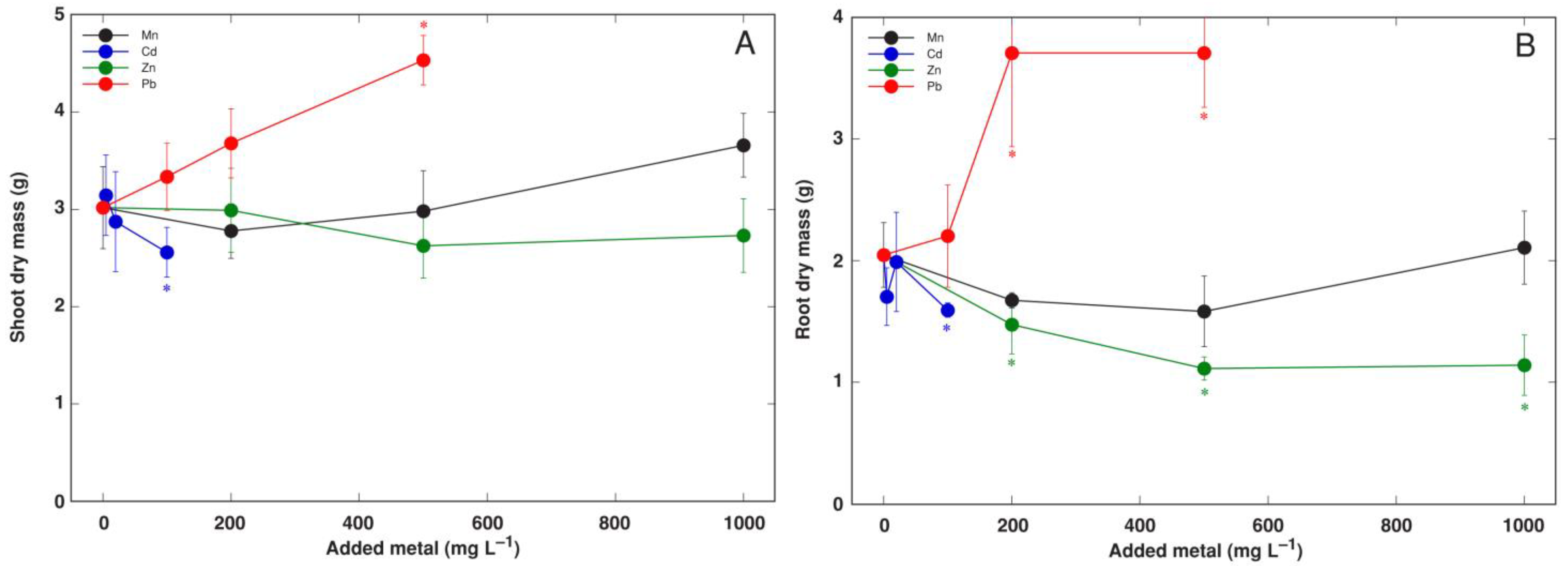
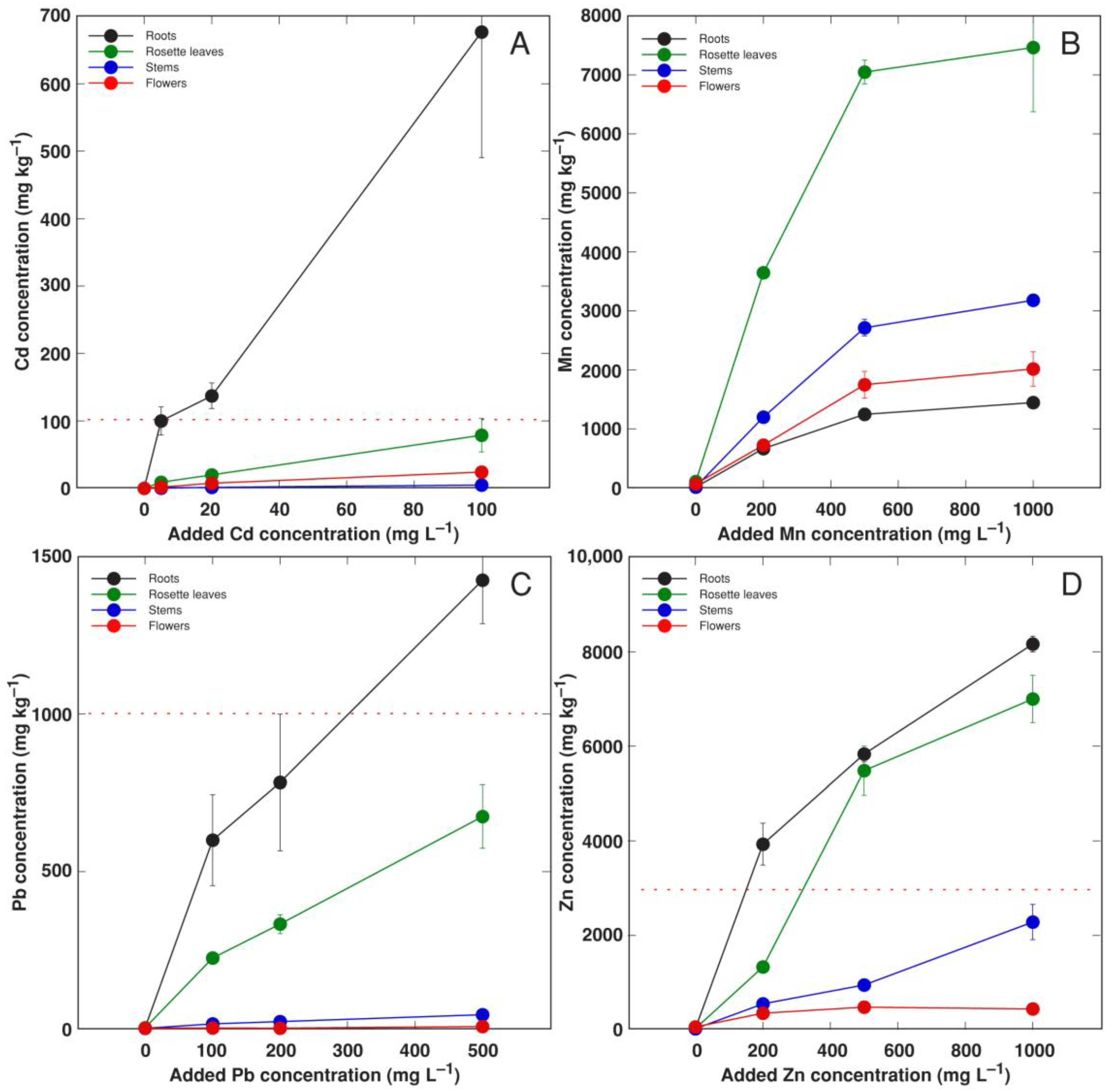
3.3. Experiment 3: Heavy Metals
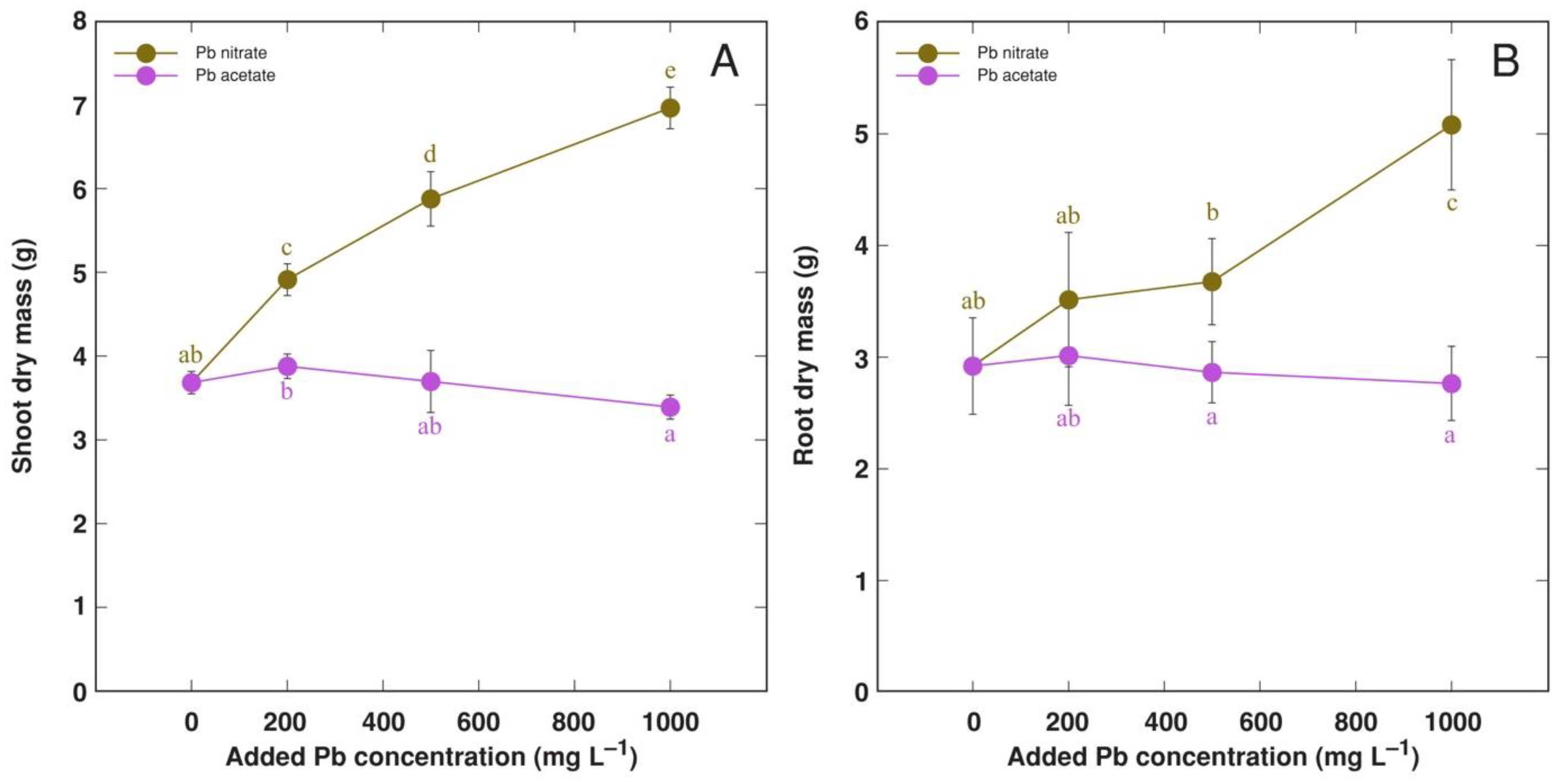
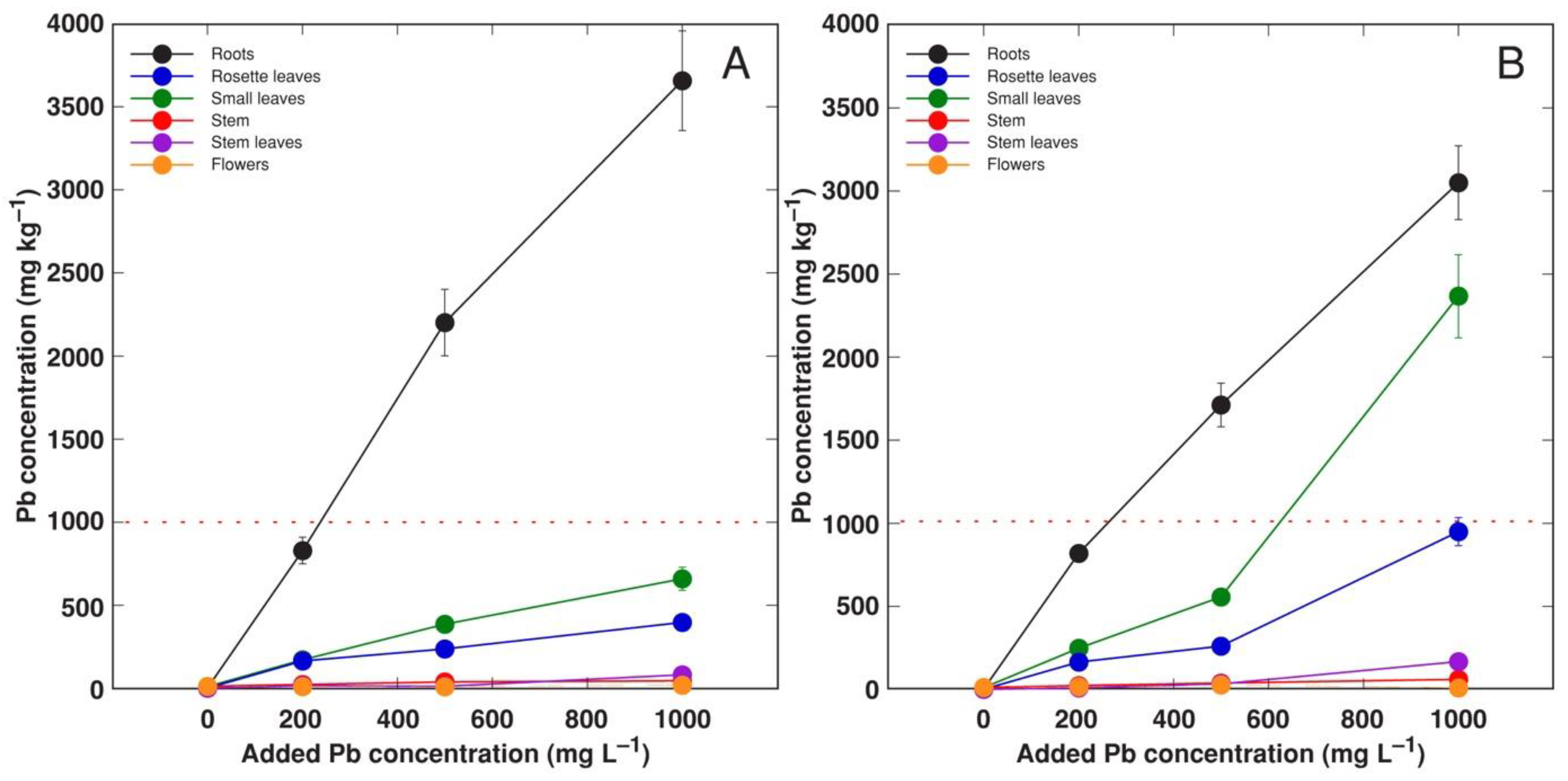
3.4. Experiment 4: Forms of Pb Salts
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Padmavathiamma, P.K.; Li, L.Y. Phytoremediation technology, hyper-accumulation metals in plants. Water Air Soil Pollut. 2007, 184, 105–126. [Google Scholar] [CrossRef]
- Mayerová, M.; Petrová, Š.; Madaras, M.; Lipavský, J.; Šimon, T.; Vaněk, T. Non-enhanced phytoextraction of cadmium, zinc, and lead by high-yielding crops. Environ. Sci. Pollut. Res. 2017, 24, 14706–14716. [Google Scholar] [CrossRef]
- Mikołajczak, P.; Borowiak, K.; Niedzielski, P. Phytoextraction of rare earth elements in herbaceous plant species growing close to roads. Environ. Sci. Pollut. Res. 2017, 24, 14091–14103. [Google Scholar] [CrossRef] [Green Version]
- Goolsby, E.W.; Mason, C.M. Toward a more physiologically and evolutionarily relevant definition of metal hyperaccumulation in plants. Front. Plant Sci. 2015, 6, 33. [Google Scholar] [CrossRef] [Green Version]
- Favas, P.C.J.; Pratas, J.; Varun, M.; D’Souza, R.; Paul, M.S. Phytoremediation of soils contaminated with metals and metalloids at mining areas, potential of native flora. In Environmental Risk Assessment of Soil Contamination; Hernandez-Soriano, M.C., Ed.; Intech Open: London, UK, 2014; pp. 485–517. [Google Scholar]
- Yang, W.; Li, H.; Zhang, T.; Sen, L.; Ni, W. Classification and identification of metal-accumulating plant species by cluster analysis. Environ. Sci. Pollut. Res. 2014, 21, 10626–10637. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Ye, Z. Metal accumulation and tolerance in wetland plants. Front. Biol. China 2009, 4, 282–288. [Google Scholar] [CrossRef]
- Kissoon, L.T.T.; Jacob, D.L.; Otte, M.L. Multi-element accumulation near Rumex crispus roots under wetland and dryland conditions. Environ. Pollut. 2010, 158, 1834–1841. [Google Scholar] [CrossRef] [Green Version]
- Teuchies, J.; Jacobs, S.; Oosterlee, L.; Bervoets, L.; Meire, P. Role of plants in metal cycling in a tidal wetland: Implications for phytoremediation. Sci. Total Environ. 2013, 445–446, 146–154. [Google Scholar] [CrossRef]
- Bonanno, G.; Vymazal, J.; Cirelli, G.L. Translocation; accumulation and bioindication of trace elements in wetland plants. Sci. Total Environ. 2018, 631–632, 252–261. [Google Scholar] [CrossRef]
- Nikalje, G.C.; Suprasanna, P. Coping with metal toxicity—Cues from halophytes. Front. Plant Sci. 2018, 9, 777. [Google Scholar] [CrossRef]
- Aziz, I.; Mujeeb, A. Halophytes for phytoremediation of hazardous metal(oid)s: A terse review on metal tolerance, bio-indication and hyperaccumulation. J. Hazard. Mater. 2022, 424, 127309. [Google Scholar] [CrossRef]
- Sruthi, P.; Shackira, A.M.; Puthur, J.T. Heavy metal detoxification mechanisms in halophytes, an overview. Wetl. Ecol. Manag. 2017, 25, 129–148. [Google Scholar] [CrossRef]
- Anjum, N.A.; Duarte, B.; Caçador, I.; Sleimi, N.; Duarte, A.C.; Pereira, E. Biophysical and biochemical markers of metal/metalloid impacts in salt marsh halophytes and their implications. Front. Environ. Sci. 2016, 4, 24. [Google Scholar] [CrossRef] [Green Version]
- Smulders, M.J.M.; Horton, R.F. Ethylene promotes elongation growth and auxin promotes radial growth in Ranunculus sceleratus petioles. Plant Physiol. 1991, 96, 806–811. [Google Scholar] [CrossRef] [Green Version]
- He, J.B.; Bögemann, G.M.; van de Steeg, H.M.; Rijnders, J.G.H.M.; Voesenek, L.A.C.J.; Blom, C.W.P.M. Survival tactics of Ranunculus species in river floodplains. Oecologia 1999, 118, 1–8. [Google Scholar] [CrossRef]
- Laime, B. 1310 Salicornia and other annuals colonising mud and sand. In European Union Protected Habitats in Latvia. Interpretation Manual; Auniņš, A., Ed.; Latvian Fund for Nature, Ministry of Environmental Protection and Regional Development: Riga, Latvia, 2013; pp. 52–54. [Google Scholar]
- Rove, I. 1640 Boreal Baltic sandy beaches with perennial vegetation. In European Union Protected Habitats in Latvia. Interpretation Manual; Auniņš, A., Ed.; Latvian Fund for Nature, Ministry of Environmental Protection and Regional Development: Riga, Latvia, 2013; pp. 58–61. [Google Scholar]
- Hill, M.O.; Ellenberg, H.H. Ellenberg’s Indicator Values for British Plants; ECOFACT Research Report. Technical Annex; Institute of Terrestrial Ecology: Wallingford, UK, 1999; Volume 2. [Google Scholar]
- Tyler, T.; Herbertsson, L.; Olofsson, J.; Olsson, P.A. Ecological indicator and traits values for Swedish vascular plants. Ecol. Indic. 2021, 120, 106923. [Google Scholar] [CrossRef]
- Ievinsh, G.; Ieviņa, S.; Andersone-Ozola, U.; Samsone, I. Leaf sodium; potassium and electrolyte accumulation capacity of plant species from salt-affected coastal habitats of the Baltic Sea: Towards a definition of Na hyperaccumulation. Flora 2021, 274, 151748. [Google Scholar] [CrossRef]
- Zuo, S.; Wan, K.; Ma, S. Environmental restoration effects of Ranunculus sceleratus L. in a eutrophic sewage system. Biochem. Syst. Ecol. 2014, 55, 34–40. [Google Scholar] [CrossRef]
- Farahat, E.A.; Galal, T.M. Trace metal accumulation by Ranunculus sceleratus, implications for phytostabilization. Environ. Sci. Pollut. Res. 2018, 25, 4214–4222. [Google Scholar] [CrossRef] [PubMed]
- Sharma, P.; Tripathi, S.; Sirohi, R.; Kim, S.H.; Ngo, H.H.; Pandey, A. Uptake and mobilization of heavy metals through phytoremediation process from native plant species growing on complex pollutants: Antioxidant enzymes and photosynthetic pigments response. Environ. Technol. Innov. 2021, 23, 101629. [Google Scholar] [CrossRef]
- Khalifa, S.M.; Badr, B.M. Habitats, phytochemical metabolites and phytoremediation potential of Ranunculus sceleratus L. Sebha Univ. J. Pure Appl. Sci. 2021, 20, 20–24. [Google Scholar]
- Losfeld, G.; L’Huillier, L.; Fogliani, B.; Mc Coy, S.; Grison, C.; Jaffré, T. Leaf-age and soil-plant relationships: Key factors for reporting trace-elements hyperaccumulation by plants and design applications. Environ. Sci. Pollut. Res. 2015, 22, 5620–5632. [Google Scholar] [CrossRef] [PubMed]
- Muszyńska, E.; Labudda, M. Dual role of metallic trace elements in stress biology—From negative to beneficial impact on plants. Int. J. Mol. Sci. 2019, 20, 3117. [Google Scholar] [CrossRef] [Green Version]
- Corso, M.; de la Torre, V.S.G. Biomolecular approaches to understanding metal tolerance and hyperaccumulation in plants. Metallomics 2020, 12, 840–859. [Google Scholar] [CrossRef]
- Manara, A.; Fasani, E.; Furini, A.; DalCorso, G. Evolution of the metal hyperaccumulation and hypertolerance traits. Plant Cell Environ. 2020, 43, 2969–2986. [Google Scholar] [CrossRef] [PubMed]
- Angulo-Bejarrano, P.I.; Puente-Rivera, J.; Cruz-Ortega, R. Metal and metalloid toxicity in plants: And overview on molecular aspects. Plants 2021, 10, 635. [Google Scholar] [CrossRef] [PubMed]
- Peng, J.-S.; Guan, Y.-H.; Lin, X.-J.; Xu, X.-J.; Xiao, L.; Wang, H.-H.; Meng, S. Comparative understanding of metal hyperaccumulation in plants: A mini review. Environ. Geochem. Health 2021, 43, 1599–1607. [Google Scholar] [CrossRef] [PubMed]
- Sytar, O.; Ghosh, S.; Malinska, H.; Zivcak, M.; Brestic, M. Physiological and molecular mechanisms of metal accumulation and hyperaccumulation. Physiol. Plant 2021, 173, 148–166. [Google Scholar]
- Seregin, I.V.; Kozhevnikova, A.D. Low-molecular-weight ligands in plants: Role in metal homeostasis and hyperaccumulation. Photosynth. Res. 2021, 150, 51–96. [Google Scholar]
- Ernst, W.H.O. Evolution of heavy metal tolerance in higher plants. For. Snow Landsc. Res. 2006, 80, 251–274. [Google Scholar]
- Baumbach, H.; Hellwig, F.H. Genetic differentiation of metallicolous and non-metallicolous Armeria maritima (Mill.) Willd. taxa (Plumbaginaceae) in Central Europe. Plant Syst. Evol. 2007, 269, 245–258. [Google Scholar] [CrossRef]
- Purmale, L.; Jēkabsone, A.; Andersone-Ozola, U.; Karlsons, A.; Osvalde, A.; Ievinsh, G. Comparison of in vitro and in planta heavy metal tolerance and accumulation potential of different Armeria maritima accessions from a dry coastal meadow. Plants 2022, 11, 2104. [Google Scholar] [CrossRef] [PubMed]
- Pollard, A.J.; Reeves, R.D.; Baker, A.J.M. Facultative hyperaccumulation of heavy metals and metaloids. Plant Sci. 2014, 217–218, 8–17. [Google Scholar] [CrossRef] [PubMed]
- Bothe, H.; Słomka, A. Divergent biology of facultative heavy metal plants. J. Plant Physiol. 2017, 219, 45–61. [Google Scholar] [CrossRef]
- Dou, X.; Dai, H.; Skuza, L.; Wei, S. Bidens pilosa L. hyperaccumulating Cd with different species in soil and the role of EDTA on the hyperaccumulation. Environ. Sci. Pollut. Res. 2019, 26, 25668–25675. [Google Scholar] [CrossRef]
- Liu, J.; Mo, L.; Zhang, X.; Yao, S.; Wang, Y. Simultaneous accumulation of cadmium and manganese in Celosia argentea Linn. Int. J. Phytoremediat. 2018, 20, 1106–1112. [Google Scholar] [CrossRef] [PubMed]
- Abubakari, F.; Nkrumah, P.N.; Fernando, D.R.; Brown, G.K.; Erdkine, P.D.; Echevarria, G.; van der Ent, A. Incidence of hyperaccumulation and tissue-level distribution of manganese, cobalt, and zinc in the genus Gossia (Myrtaceae). Metallomics 2021, 13, mfam008. [Google Scholar] [CrossRef]
- Coakley, S.; Gahill, G.; Enright, A.-M.; O’Rourke, B.; Petti, C. Cadmium hyperaccumulation and translocation in Impatiens glandulifera: From foe to friend? Sustainability 2019, 11, 5018. [Google Scholar] [CrossRef] [Green Version]
- Moray, C.; Goolsby, E.W.; Bromham, L. The phylogenetic association between salt tolerance and heavy metal hyperaccumulation in angiosperms. Evol. Biol. 2016, 43, 119–130. [Google Scholar] [CrossRef]
- Reeves, R.D.; Baker, A.J.M.; Jaffré, T.; Erskine, P.D.; Echevarria, G.; van der Ent, A. A global database for plants that hyperaccumulate metal and metalloid trace elements. New Phytol. 2018, 218, 407–411. [Google Scholar] [CrossRef] [Green Version]
- Liang, H.-M.; Lin, T.-H.; Chiou, J.-M.; Yeh, K.-C. Model evaluation of the phytoextraction potential of heavy metal hyperaccumulators and non-hyperaccumulators. Environ. Pollut. 2009, 157, 1945–1952. [Google Scholar] [CrossRef] [PubMed]
- Ievinsh, G.; Andersone-Ozola, U.; Jēkabsone, A. Similar responses of relatively salt tolerant plants to Na and K during chloride salinity: Comparison of growth, water content and ion accumulation. Life 2022, 12, 1577. [Google Scholar] [CrossRef] [PubMed]
- Purmale, L.; Jēkabsone, A.; Andersone-Ozola, U.; Ievinsh, G. Salinity tolerance and ion accumulation potential in vitro and in planta of different Armeria maritima accessions from a dry coastal meadow. Plants 2022, 11, 2570. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Hussain, T.; Feng, X.; Guo, K.; Chen, H.; Yang, C.; Liu, X. Comparative study on the resistance of Suaeda glauca and Suaeda salsa to drought, salt, and alkali stresses. Ecol. Eng. 2019, 140, 105593. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, W.; Yan, X.; Guo, J. Physiological and transcriptomic analyses of yellow horn (Xanthoceras sorbifolia) provide important insights into salt and saline-alkali stress tolerance. PLoS ONE 2020, 15, e0244365. [Google Scholar] [CrossRef]
- Geng, G.; Wang, G.; Stevanato, P.; Lv, C.; Wang, Q.; Yu, L.; Wang, Y. Physiological and proteomic analysis of different molecular mechanisms of sugar beet response to acidic and alkaline pH environment. Front. Plant Sci. 2021, 12, 682799. [Google Scholar] [CrossRef]
- Yang, C.W.; Wang, P.; Li, C.Y.; Shi, D.C.; Wang, D.L. Comparison of effects of salt and alkali stresses on the growth and photosynthesis of wheat. Photosynthetica 2008, 46, 107–114. [Google Scholar] [CrossRef]
- Guo, R.; Shi, L.; Yan, G.; Zhong, X.; Gu, F.; Liu, Q.; Xia, X.; Li, H. Ionomic and metabolic responses to salt or alkaline salt stresses in maize (Zea mays L.) seedlings. BMC Plant Biol. 2017, 17, 41. [Google Scholar] [CrossRef] [Green Version]
- Cole, J.J.; Prairie, Y.T. Dissolved CO2 in freshwater systems. In Encyclopedia of Inland Waters; Elsevier: Amsterdam, The Netherlands, 2009; pp. 30–34. [Google Scholar]
- Poschenrieder, C.; Fernández, J.A.; Rubio, L.; Pérez, L.; Terés, J.; Barcélo, J. Transport and use of bicarbonate in plants: Current knowledge and challenges ahead. Int. J. Mol. Sci. 2018, 19, 1352. [Google Scholar] [CrossRef] [Green Version]
- Csitári, B.; Bedics, A.; Felföldi, T.; Boros, E.; Nagy, H.; Máthé, I.; Szkely, A.J. Anion-type modulates the effect of salt stress on saline lake bacteria. Extremophiles 2022, 26, 12. [Google Scholar] [CrossRef]
- Crawford, N.M. Nitrate: Nutrient and signal for plant growth. Plant Cell 1995, 7, 859–868. [Google Scholar]
- Bharucha, F.R.; Dubash, P.J. The problem of nitrophily. Vegetatio 1951, 3, 183–194. [Google Scholar] [CrossRef]
- Moreau, D.; Milard, G.; Munier-Jolain, N. A plant nitrophily index based on plant leaf area response to soil nitrogen availability. Agron. Sustain. Dev. 2013, 33, 809–815. [Google Scholar] [CrossRef]
- Xu, G.; Fan, X.; Miller, A.J. Plant nitrogen assimilation and use efficiency. Annu. Rev. Plant Biol. 2012, 63, 153–182. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Hu, B.; Chu, C. Nitrogen assimilation in plants: Current status and future perspectives. J. Genet. Genom. 2022, 49, 394–404. [Google Scholar] [CrossRef] [PubMed]
- Kotur, Z.; Siddiqi, Y.M.; Glass, A.D.M. Characterization of nitrite uptake in Arabidopsis thaliana: Evidence for a nitrite-specific transporter. New Phytol. 2013, 200, 201–210. [Google Scholar] [CrossRef] [PubMed]
- Maia, L.B.; Moura, J.J.G. How biology handles nitrite. Chem. Rev. 2014, 114, 5273–5357. [Google Scholar]
- Ezzine, M.; Debouba, M.; Ghorbel, M.H.; Gouia, H. Ion uptake and structural modifications induced by nitrogen source in tomato (Solanum lycopersicum Mill. Cv. Ibiza F1). C. R. Biol. 2011, 334, 526–534. [Google Scholar] [CrossRef]
- Zsoldos, F.; Vashegyi, Á.; Pécsváradi, A.; Haunold, E.; Herger, P. Unhibition of ion uptake and growth of wheat and rice exposed to nitrite at low pH. Cereal Res. Commun. 1997, 25, 35–42. [Google Scholar] [CrossRef]
- Samater, A.H.; Van Cleemput, O.; Ertebo, T. Influence of the presence of nitrite and nitrate in soil on maize biomass production, nitrogen immobilization and nitrogen recover. Biol. Fertil. Soils 1998, 27, 211–218. [Google Scholar] [CrossRef]
- Sharma, P.; Ahmad, M.; Rathee, S.; Singh, H.P.; Batish, D.R.; Kohli, R.K. Bridging the gap: Linking morpho-functional traits’ plasticity with hyperaccumulation. Environ. Monit. Assess. 2021, 193, 762. [Google Scholar] [CrossRef] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experiment No. | Description | Treatments (Final Concentrations in Substrate) | Duration of Experiment |
|---|---|---|---|
| 1 | Effect of NaCl gradient | NaCl (equivalent to 0.05, 0.1, 0.2, 0.5, 1, 2, 5, 10 g L−1 Na+) | Treatment phase 3 weeks, cultivation phase 5 weeks |
| 2 | Effect of different Na+ and K+ salts | NaCl, NaNO3, NaNO2, Na2CO3 (equivalent to 4.0 g L−1 Na+); KCl, KNO3, KNO2, K2CO3 (equivalent to 6.8 g L−1 K+) | Treatment phase 2 weeks, cultivation phase 5 weeks |
| 3 | Comparison of heavy metal tolerance and accumulation | MnSO4 (equivalent to 200, 500, 1000 mg L−1 Mn), CdSO4 (equivalent to 5, 20, 100 mg L−1 Cd), ZnSO4 (equivalent to 200, 500, 1000 mg L−1 Zn), Pb(NO3)2 (equivalent to 100, 200, 500 mg L−1 Pb) | Treatment phase 2 weeks, cultivation phase 5 weeks |
| 4 | Comparison of Pb nitrate and acetate | Pb(NO3)2, Pb(CH3COO)2 (equivalent to 200, 500, 1000 mg L−1 Pb) | Treatment phase 2 weeks, cultivation phase 7 weeks |
| Na+ (g L−1) | Roots | Rosette Leaf Petioles | Rosette Leaf Blades | Stem | Stem Leaves | Flowers |
|---|---|---|---|---|---|---|
| 0.05 | 174 | 107 | 93 | 91 | 43 | 43 |
| 0.1 | 121 | 68 | 59 | 56 | 27 | 22 |
| 0.2 | 96 | 43 | 35 | 39 | 15 | 18 |
| 0.5 | 25 | 24 | 15 | 18 | 7 | 7 |
| 1 | 13 | 23 | 11 | 14 | 8 | 6 |
| 2 | 8 | 14 | 6 | 9 | 6 | 5 |
| 5 | 5 | 8 | 6 | 7 | 7 | 3 |
| 10 | 5 | 7 | 2 | 9 | 5 | 3 |
| Metal | Concentration (mg L−1) | Roots | Leaves | Stem | Flowers |
|---|---|---|---|---|---|
| Mn | 200 | 3.3 | 18.3 | 6.0 | 3.6 |
| 500 | 2.5 | 14.1 | 5.4 | 3.6 | |
| 1000 | 1.5 | 7.5 | 3.2 | 2.0 | |
| Cd | 5 | 20.0 | 1.8 | 0.10 | 0.33 |
| 20 | 6.9 | 1.0 | 0.08 | 0.38 | |
| 100 | 6.8 | 0.8 | 0.05 | 0.24 | |
| Zn | 200 | 19.7 | 6.7 | 2.8 | 1.76 |
| 500 | 11.7 | 11.0 | 1.9 | 0.96 | |
| 1000 | 8.2 | 7.0 | 2.3 | 0.44 | |
| Pb | 100 | 6.0 | 2.3 | 0.16 | 0.03 |
| 200 | 3.9 | 1.7 | 0.12 | 0.01 | |
| 500 | 2.9 | 1.4 | 0.09 | 0.02 |
| Salt | Concentration (mg L−1) | Roots | Small Leaves | Rosette Leaves | Stem | Stem Leaves | Flowers |
|---|---|---|---|---|---|---|---|
| Pb nitrate | 200 | 4.2 | 0.87 | 0.83 | 0.12 | 0.09 | 0.06 |
| 500 | 4.0 | 0.77 | 0.48 | 0.08 | 0.03 | 0.02 | |
| 1000 | 3.7 | 0.66 | 0.40 | 0.05 | 0.08 | 0.02 | |
| Pb acetate | 200 | 4.1 | 1.25 | 1.83 | 0.12 | 0.05 | 0.09 |
| 500 | 3.4 | 1.11 | 0.52 | 0.08 | 0.07 | 0.06 | |
| 1000 | 3.1 | 2.37 | 0.95 | 0.06 | 0.17 | 0.01 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ievinsh, G.; Landorfa-Svalbe, Z.; Andersone-Ozola, U.; Karlsons, A.; Osvalde, A. Salinity and Heavy Metal Tolerance, and Phytoextraction Potential of Ranunculus sceleratus Plants from a Sandy Coastal Beach. Life 2022, 12, 1959. https://doi.org/10.3390/life12121959
Ievinsh G, Landorfa-Svalbe Z, Andersone-Ozola U, Karlsons A, Osvalde A. Salinity and Heavy Metal Tolerance, and Phytoextraction Potential of Ranunculus sceleratus Plants from a Sandy Coastal Beach. Life. 2022; 12(12):1959. https://doi.org/10.3390/life12121959
Chicago/Turabian StyleIevinsh, Gederts, Zaiga Landorfa-Svalbe, Una Andersone-Ozola, Andis Karlsons, and Anita Osvalde. 2022. "Salinity and Heavy Metal Tolerance, and Phytoextraction Potential of Ranunculus sceleratus Plants from a Sandy Coastal Beach" Life 12, no. 12: 1959. https://doi.org/10.3390/life12121959