
Alleviation of Cadmium Stress by Silicon Supplementation in Peas by the Modulation of Morpho-Physio-Biochemical Variables and Health Risk Assessment
 and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site, Experimental Design and Treatments
2.2. Crop Management
2.3. Morphological, Phenological and Biomass Variables
2.4. Physio-Biochemical and Water-Related Variables
2.5. Enzymatic Antioxidant Variables
2.6. Cadmium Contents in Plant Tissues
2.7. Health Risk Assessment
2.8. Statistical Analysis
3. Results
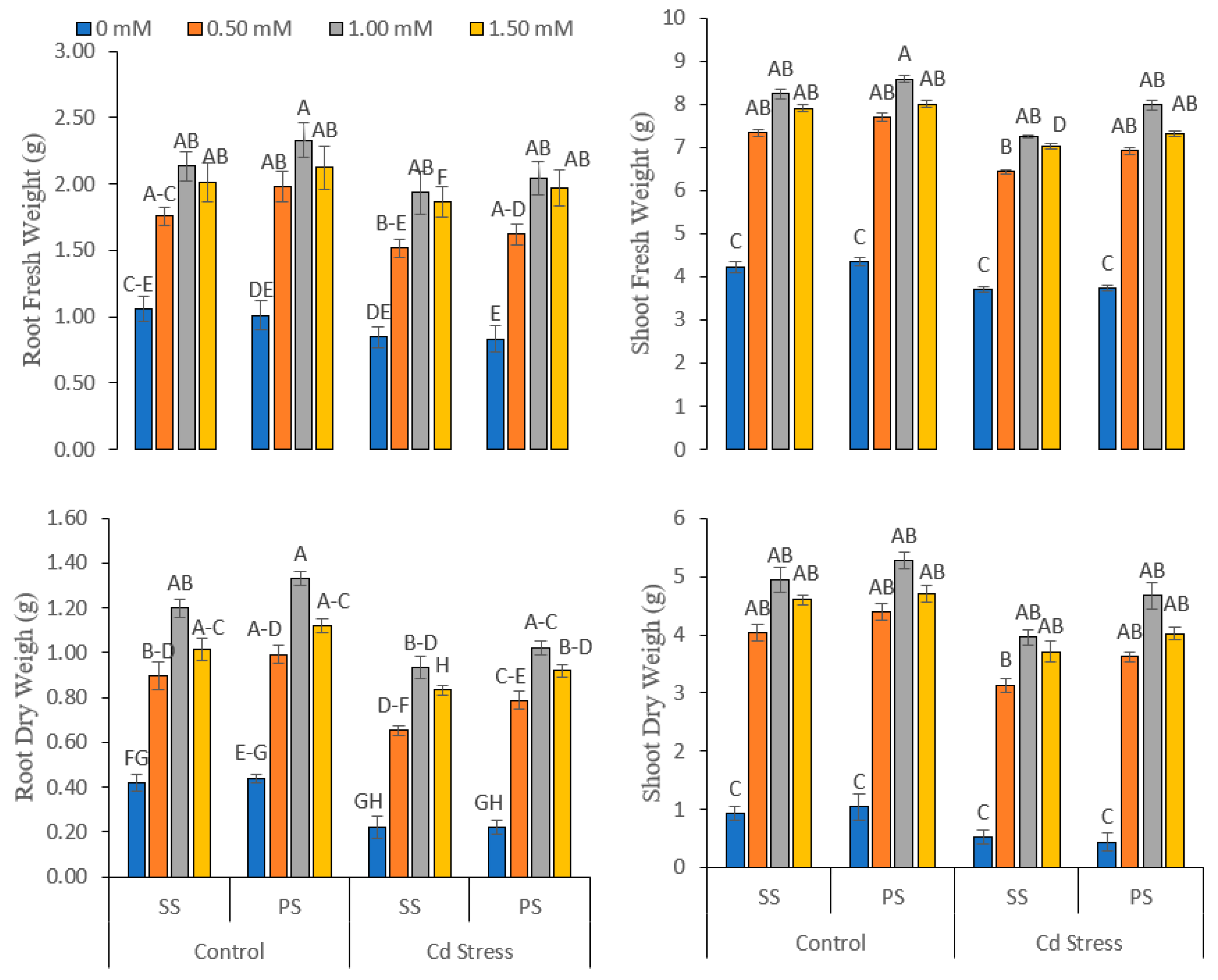
3.1. Morphological, Phenological and Biomass Variables
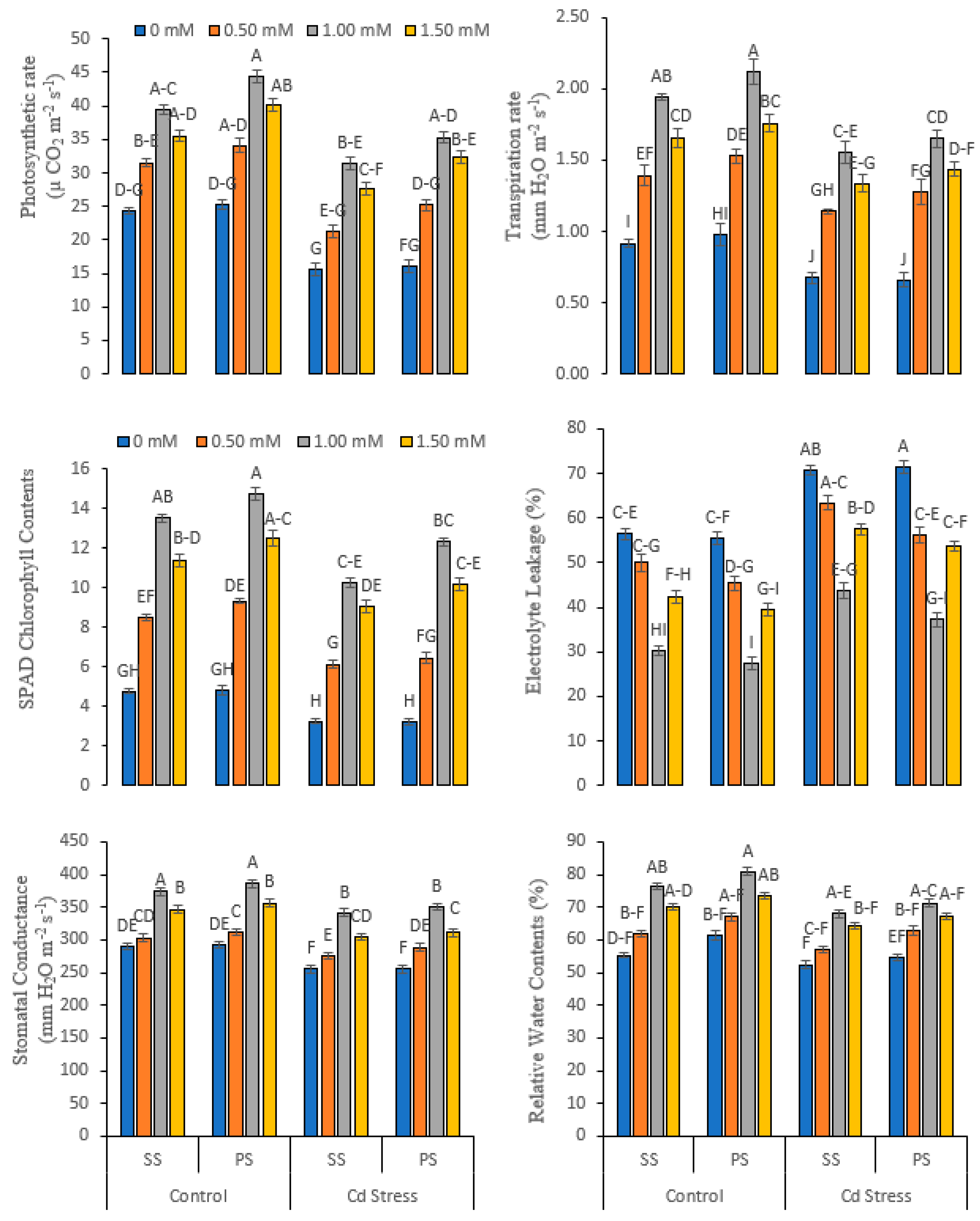
3.2. Physio-Biochemical and Water-Related Variables
3.3. Enzymatic Antioxidants Variables
3.4. Cadmium Contents in Plant Tissues
3.5. Health Risk Index
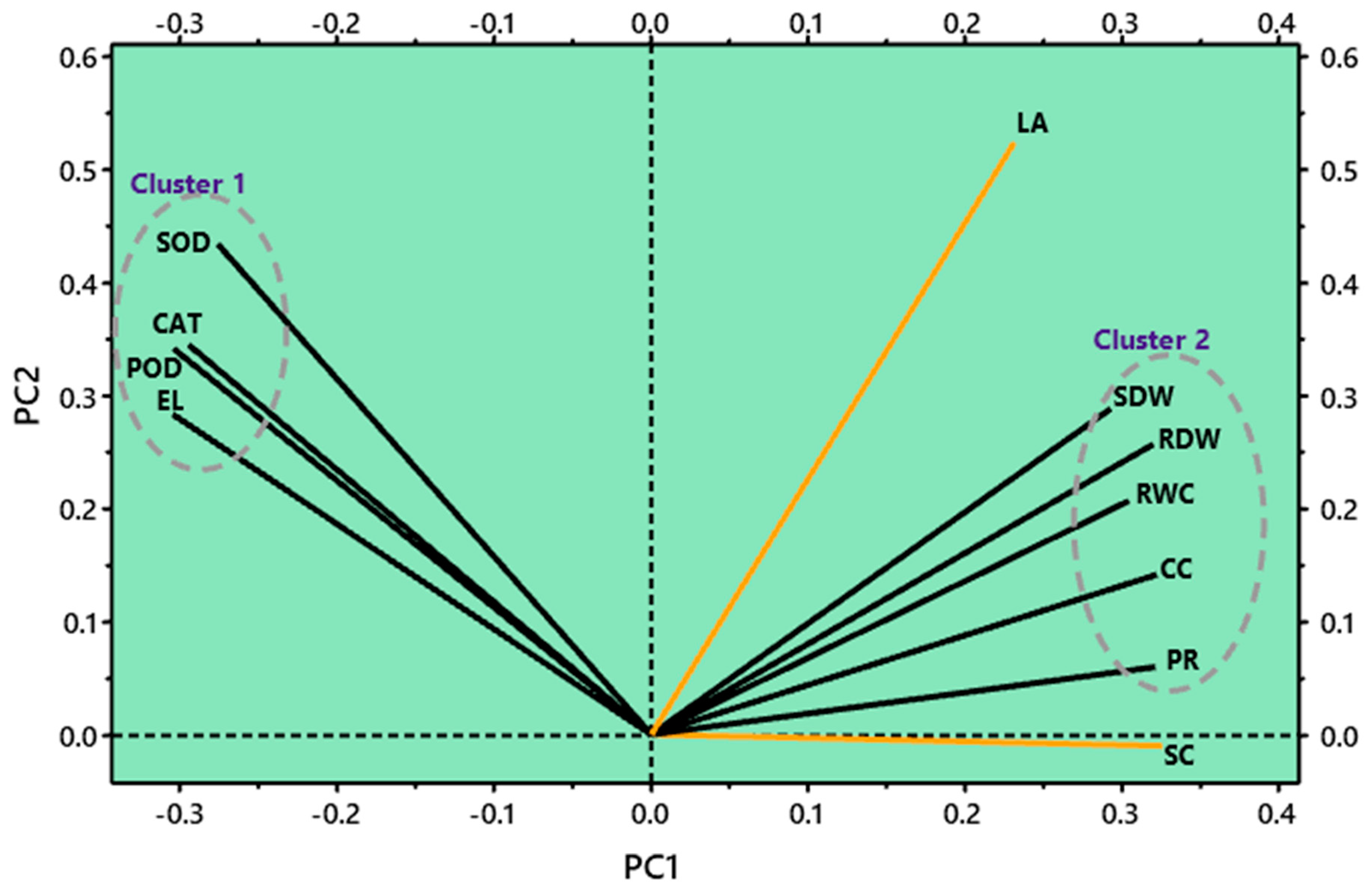
3.6. Correlation and Principal Component Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zaman, Q.; Rashid, M.; Nawaz, R.; Hussain, A.; Ashraf, K.; Latif, M.; Heile, A.O.; Chen, Y. Silicon Fertilization: A Step towards Cadmium-Free Fragrant Rice. Plants 2021, 10, 2440. [Google Scholar] [CrossRef] [PubMed]
- Heile, A.O.; Zaman, Q.; Aslam, Z.; Hussain, A.; Aslam, M.; Saleem, M.H.; Abualreesh, M.H.; Ali, S. Alleviation of Cadmium Phytotoxicity Using Silicon Fertilization in Wheat by Altering Antioxidant Metabolism and Osmotic Adjustment. Sustainability 2021, 13, 11317. [Google Scholar] [CrossRef]
- Ma, Q.; Haider, F.U.; Farooq, M.; Adeel, M.; Shakoor, N.; Wu, J.; Xu, J.; Xu, W.W.; Luo, P.; Cai, L. Selenium treated Foliage and biochar treated soil for improved lettuce (Lactuca sativa L.) growth in Cd-polluted soil. J. Clean. Prod. 2022, 335, 130267. [Google Scholar]
- Ahmad, P.; Ahanger, M.A.; Egamberdieva, D.; Alam, P.; Alyemeni, M.N.; Ashraf, M. Modification of osmolytes and antioxidant enzymes by 24-epibrassinolide in chickpea Seedlings under mercury (Hg) toxicity. J. Plant Growth Regul. 2018, 37, 309–322. [Google Scholar] [CrossRef]
- Yousaf, B.; Amina, L.G.; Wang, R.; Imtiaz, M.; Rizwan, M.S.; Zia-ur-Rehman, M.; Qadir, A.; Si, Y. The importance of evaluating metal exposure and predicting human health risks in urban–periurban environments influenced by emerging industry. Chemosphere 2016, 150, 79–89. [Google Scholar] [CrossRef] [PubMed]
- Zulfiqar, U.; Jiang, W.; Xiukang, W.; Hussain, S.; Ahmad, M.; Maqsood, M.F.; Ali, N.; Ishfaq, M.; Kaleem, M.; Haider, F.U.; et al. Cadmium Phytotoxicity, Tolerance, and Advanced Remediation Approaches in Agricultural Soils; A Comprehensive Review. Front. Plant Sci. 2022, 13, 773815. [Google Scholar] [CrossRef] [PubMed]
- Haider, F.U.; Virk, A.L.; Rehmani, M.I.A.; Skalicky, M.; Ata-ul-Karim, S.T.; Ahmad, N.; Soufan, W.; Brestic, M.; Sabagh, A.E.L.; Liqun, C. Integrated application of thiourea and biochar improves maize growth, antioxidant activity and reduces cadmium bioavailability in cadmium-contaminated soil. Front. Plant Sci. 2021, 12, 809322. [Google Scholar] [CrossRef]
- Haider, F.U.; Farooq, M.; Naveed, M.; Cheema, S.A.; Ain, U.-N.; Salim, M.A.; Liqun, C.; Mustafa, A. Influence of biochar and microorganism co-application on the remediation and maize growth in cadmium-contaminated soil. Front. Plant Sci. 2022, 13, 983830. [Google Scholar] [CrossRef]
- O’Mara, K.; Cresswell, T. Trophic variations in uptake and assimilation of cadmium, manganese and zinc: An estuarine food-chain radiotracer experiment. World Academy of Science, Engineering and Technology. Int. J. Environ. Ecol. Eng. 2018, 5, 1. [Google Scholar]
- Rahman, M.F.; Ghosal, A.; Alam, M.F.; Kabir, A.H. Remediation of cadmium toxicity in field peas (Pisum sativum L.) through exogenous silicon. Ecotoxicol. Environ. Saf. 2017, 135, 165–172. [Google Scholar] [CrossRef]
- Haider, F.U.; Wang, X.; Farooq, M.; Hussain, S.; Cheema, S.A.; Ain, U.-N.; Virk, A.L.; Ejaz, M.; Janyshova, U.; Liqun, C. Biochar application for the remediation of trace metals in contaminated soils: Implications for stress tolerance and crop production. Ecotoxicol. Environ. Saf. 2022, 230, 113165. [Google Scholar] [CrossRef]
- Akhter, N.; Rahman, M.M.; Hasanuzzaman, M.; Nahar, K. Physiological response of garden pea (Pisum sativum L.) grown under different light environment. Bot. Res. Int. 2009, 2, 304–309. [Google Scholar]
- Jan, S.; Alyemeni, M.N.; Wijaya, L.; Alam, P.; Siddique, K.H.; Ahmad, P. Interactive effect of 24-epibrassinolide and silicon alleviates cadmium stress via the modulation of antioxidant defense and glyoxalase systems and macronutrient content in Pisum sativum L. seedlings. BMC Plant Biol. 2018, 18, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Aziz, R.; Rafiq, M.T.; Li, T.; Liu, D.; He, Z.; Stoffella, P.J.; Sun, K.; Xiaoe, Y. Uptake of Cadmium by Rice Grown on Contaminated Soils and Its Bioavailability/Toxicity in Human Cell Lines (Caco-2/HL-7702). J. Agric. Food Chem. 2015, 63, 3599–3608. [Google Scholar] [CrossRef] [PubMed]
- Lux, A.; Martinka, M.; Vaculík, M.; White, P.J. Root responses to cadmium in the rhizosphere: A review. J. Exp. Bot. 2010, 62, 21–37. [Google Scholar] [CrossRef]
- Naveed, M.; Saifullah; Riaz, U.; Murtaza, G.; Bibi, S.; Arooj, A.; Zaman, Q.U. Strategic use of water: A step toward cadmium-free basmati rice (Oryza sativa L.). Paddy Water Environ. 2018, 16, 867–873. [Google Scholar] [CrossRef]
- Sandalio, L.M.; Dalurzo, H.C.; Gomez, M.; Romero-Puertas, M.C.; Del Rio, L.A. Cadmium-induced changes in the growth and oxidative metabolism of pea plants. J. Exp. Bot. 2001, 52, 2115–2126. [Google Scholar] [CrossRef]
- Hamid, A.; Wasim, A.; Azfar, A.; Amjad, R.; Nazir, R. Monitoring and health risk assessment of selected trace metals in wheat rice and soil samples. Food Sci. Technol. 2020, 40, 917–923. [Google Scholar] [CrossRef]
- Ashraf, S.; Ali, Q.; Zahir, Z.A.; Ashraf, S.; Asghar, H.N. Phytoremediation: Environmentally sustainable way for reclamation of heavy metal polluted soils. Ecotoxicol. Environ. Saf. 2019, 174, 714–727. [Google Scholar] [CrossRef]
- Azeem, M.; Ali, A.; Jeyasundar, P.G.S.A.; Li, Y.; Abdelrahman, H.; Latif, A.; Li, R.; Basta, N.; Li, G.; Shaheen, S.M. Bone-derived biochar improved soil quality and reduced Cd and Zn phytoavailability in a multi-metal contaminated mining soil. Environ. Pollut. 2021, 277, 116800. [Google Scholar] [CrossRef]
- Luo, W.; Ma, J.; Aman Khan, M.; Liao, S.; Ruan, Z.; Liu, H.; Zhong, B.; Zhu, Y.; Duan, L.; Fu, L. Cadmium accumulation in rice and its bioavailability in paddy soil with application of silicon fertilizer under different water management regimes. Soil Use Manag. 2021, 37, 299–306. [Google Scholar] [CrossRef]
- Radzali, N.; Kadir, W.; Shariff, S.M.; Nawahwi, M.Z.; Wakid, S.A.; Jaafar, Z.; Rahim, M.I. Phytoremediation: Environmental friendly clean up method. World 2015, 5, 16–22. [Google Scholar]
- Tahir, N.; Ullah, A.; Tahir, A.; Rashid, H.U.; Rehman, T.U.; Danish, S.; Hussain, B.; Akca, H. Strategies for reducing Cd concentration in paddy soil for rice safety. J. Clean. Prod. 2021, 316, 128116. [Google Scholar]
- Ali, S.; Rizwan, M.; Hussain, A.; Zia Rehman, M.; Ali, B.; Yousaf, B.; Wijaya, L.; Alyemeni, M.N.; Ahmad, P. Silicon nano-particles enhanced the growth and reduced the cadmium accumulation in grains of wheat (Triticum aestivum L.). Plant Physiol. Biochem. 2019, 140, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Putko, P.; Kwaśny, M. Bioavailable silicon forms in dietary supplements. Bull. Mil. Univ. Technol. 2020, 69, 35–41. [Google Scholar] [CrossRef]
- Farooq, M.A.; Ali, S.; Hameed, A.; Ishaque, W.; Mahmood, K.; Iqbal, Z. Alleviation of cadmium toxicity by silicon is related to elevated photosynthesis, antioxidant enzymes; suppressed cadmium uptake and oxidative stress in cotton. Ecotoxicol. Environ. Saf. 2013, 96, 242–249. [Google Scholar] [CrossRef] [PubMed]
- Song, A.; Li, Z.; Zhang, J.; Xue, G.; Fan, F.; Liang, Y. Silicon-enhanced resistance to cadmium toxicity in Brassica chinensis L. is attributed to Si-suppressed cadmium uptake and transport and Si-enhanced antioxidant defense capacity. J. Hazard. Mater. 2009, 172, 74–83. [Google Scholar] [CrossRef]
- Rizwan, M.; Meunier, J.-D.; Davidian, J.; Pokrovsky, O.S.; Bovet, N.; Keller, C. Silicon alleviates Cd stress of wheat seedlings (Triticum turgidum L. cv. Claudio) grown in hydroponics. Environ. Sci. Pollut. Res. 2016, 23, 1414–1427. [Google Scholar] [CrossRef]
- Shi, G.; Cai, Q.; Liu, C.; Wu, L. Silicon alleviates cadmium toxicity in peanut plants in relation to cadmium distribution and stimulation of antioxidative enzymes. Plant Growth Regul. 2010, 61, 45–52. [Google Scholar] [CrossRef]
- Li, P.; Wang, X.; Zhang, T.; Zhou, D.; He, Y. Effects of several amendments on rice growth and uptake of copper and cadmium from a contaminated soil. J. Environ. Sci. 2008, 20, 449–455. [Google Scholar] [CrossRef]
- Wang, Y.; Hu, Y.; Duan, Y.; Feng, R.; Gong, H. Silicon reduces long-term cadmium toxicities in potted garlic plants. Acta Physiol. Plant. 2016, 38, 211. [Google Scholar] [CrossRef]
- Gill, R.A.; Zang, L.; Ali, B.; Farooq, M.A.; Cui, P.; Yang, S.; Ali, S.; Zhou, W. Chromium-induced physio-chemical and ultrastructural changes in four cultivars of Brassica napus L. Chemosphere 2015, 120, 154–164. [Google Scholar] [CrossRef] [PubMed]
- Hussain, A.; Ali, S.; Rizwan, M.; ur Rehman, M.Z.; Hameed, A.; Hafeez, F.; Alamri, S.A.; Alyemeni, M.N.; Wijaya, L. Role of zinc–lysine on growth and chromium uptake in rice plants under Cr stress. J. Plant Growth Regul. 2018, 37, 1413–1422. [Google Scholar] [CrossRef]
- Ahmad, R.; Ali, S.; Rizwan, M.; Dawood, M.; Farid, M.; Hussain, A.; Wijaya, L.; Alyemeni, M.N.; Ahmad, P. Hydrogen sulfide alleviates chromium stress on cauliflower by restricting its uptake and enhancing antioxidative system. Physiol. Plant. 2020, 168, 289–300. [Google Scholar] [CrossRef]
- Saleem, M.H.; Fahad, S.; Adnan, M.; Ali, M.; Rana, M.S.; Kamran, M.; Ali, Q.; Hashem, I.A.; Bhantana, P.; Ali, M. Foliar application of gibberellic acid endorsed phytoextraction of copper and alleviates oxidative stress in jute (Corchorus capsularis L.) plant grown in highly copper-contaminated soil of China. Environ. Sci. Pollut. Res. 2020, 27, 37121–37133. [Google Scholar] [CrossRef]
- Rico-Garcia, E.; Hernandez-Hernandez, F.; Soto-Zarazua, G.M.; Herrera-Ruiz, G. Two new methods for the estimation of leaf area using digital photography. Int. J. Agric. Biol. 2009, 11, 97–400. [Google Scholar]
- Lutts, S.; Guerrier, G. Peroxidase activities of two rice cultivars differing in salinity tolerance as affected by proline and NaCl. Biol. Plant. 1995, 37, 577–586. [Google Scholar] [CrossRef]
- Velikova, V.; Yordanov, I.; Edreva, A. Oxidative stress and some antioxidant systems in acid rain-treated bean plants: Protective role of exogenous polyamines. Plant Sci. 2000, 151, 59–66. [Google Scholar] [CrossRef]
- Aebi, H. Catalase. In Methods of Enzymatic Analysis; Academic Press: Cambridge, MA, USA, 1974; pp. 673–684. [Google Scholar]
- Beauchamp, C.; Fridovich, I. Superoxide dismutase: Improved assays and an assay applicable to acrylamide gels. Anal. Biochem. 1971, 44, 276–287. [Google Scholar] [CrossRef]
- Ryan, P.R.; Delhaize, E.; Jones, D.J. Function and mechanism of organic anion exudation from plant roots. Annu. Rev. Plant Physiol. Plant Mol. Biol. 2001, 52, 527–560. [Google Scholar] [CrossRef]
- Wachirawongsakorn, P. Health risk assessment via consumption of Pb and Cd contaminated vegetables collected from fresh markets in the lower north of Thailand. Hum. Ecol. Risk Assess. Int. J. 2016, 22, 611–622. [Google Scholar] [CrossRef]
- Zhuang, P.; Lu, H.; Li, Z. Multiple exposure and effects assessment of heavy metals in the population near mining area in South China. PLoS Med. 2014, 9, e94484. [Google Scholar] [CrossRef] [PubMed]
- United States Environmental Protection Agency. Risk-Based Concentration Table; United States Environmental Protection Agency: Philadelphia, PA, USA, 2000.
- Imtiaz, M.; Tu, S.; Xie, Z.; Han, D.; Ashraf, M.; Rizwan, M.S. Growth, V uptake, and antioxidant enzymes responses of chickpea (Cicer arietinum L.) genotypes under vanadium stress. Plant Soil 2015, 390, 17–27. [Google Scholar] [CrossRef]
- Guo, L.; Chen, A.; He, N.; Yang, D.; Liu, M. Exogenous silicon alleviates cadmium toxicity in rice seedlings in relation to Cd distribution and ultrastructure changes. J. Soils Sediments 2017, 18, 1691–1700. [Google Scholar] [CrossRef]
- Keller, C.; Rizwan, M.; Davidian, J.C.; Pokrovsky, O.S.; Bovet, N.; Chaurand, P.; Meunier, J.D. Effect of silicon on wheat seedlings (Triticum turgidum L.) grown in hydroponics and exposed to 0 to 30 mM Cu. Planta 2015, 241, 847–860. [Google Scholar] [CrossRef]
- Fan, X.Y.; Wen, X.H.; Huang, F.; Cai, Y.X.; Cai, K.Z. Effects of silicon on morphology, ultrastructure and exudates of rice root under heavy metal stress. Acta Physiol. Plant. 2016, 38, 197. [Google Scholar] [CrossRef]
- Hussain, A.; Rizwan, M.; Ali, Q.; Ali, S. Seed priming with silicon nanoparticles improved the biomass and yield while reduced the oxidative stress and cadmium concentration in wheat grains. Environ. Sci. Pollut. Res. 2019, 26, 7579–7588. [Google Scholar] [CrossRef]
- Gheshlaghpour, J.; Asghari, B.; Khademian, R.; Sedaghati, B. Silicon alleviates cadmium stress in basil (Ocimum basilicum L.) through alteration of phytochemical and physiological characteristics. Indus. Crop. Prod. 2021, 163, 113338. [Google Scholar] [CrossRef]
- Khaliq, A.; Ali, S.; Hameed, A.; Farooq, M.A.; Farid, M.; Shakoor, M.B.; Mahmood, K.; Ishaque, W.; Rizwan, M. Silicon alleviates nickel toxicity in cotton seedlings through enhancing growth, photosynthesis and suppressing Ni uptake and oxidative stress. Arch. Agron. Soil Sci. 2016, 62, 633–647. [Google Scholar] [CrossRef]
- Ghasemi Mianaei, A.; Mobasser, H.R.; Madani, H.; Dastan, S. Silicon and potassium application facts on lodging related characteristics and quantity yield in rice (Oryza sativa L.) Tarom Hashemi variety. J. New Find. Agric. 2011, 5, 423–435. [Google Scholar]
- Chen, R.; Zhang, C.; Zhao, Y.; Huang, Y.; Liu, Z. Foliar application with nano-silicon reduced cadmium accumulation in grains by inhibiting cadmium translocation in rice plants. Environ. Sci. Pollut. Res. 2018, 25, 2361–2368. [Google Scholar] [CrossRef] [PubMed]
- Rios, J.J.; Martínez-Ballesta, M.C.; Ruiz, J.M.; Blasco, B.; Carvajal, M. Silicon- mediated improvement in plant salinity tolerance: The role of aquaporins. Front. Plant Sci. 2017, 8, 948. [Google Scholar] [CrossRef] [PubMed]
- Ming, D.F.; Pei, Z.F.; Naeem, M.S.; Gong, H.J.; Zhou, W.J. Silicon alleviates PEG-induced water-deficit stress in upland rice seedlings by enhancing osmotic adjustment. J. Agron. Crop Sci. 2012, 198, 14–26. [Google Scholar] [CrossRef]
- Kabir, A.H.; Hossain, M.M.; Khatun, M.A.; Mandal, A.; Haider, S.A. Role of silicon counteracting cadmium toxicity in Alfalfa (Medicago sativa L.). Front. Plant Sci. 2016, 7, 1117. [Google Scholar] [CrossRef]
- Kim, Y.H.; Khan, A.L.; Lee, I.J. Silicon: A duo synergy for regulating crop growth and hormonal signaling under abiotic stress conditions. Crit. Rev. Biotechnol. 2016, 36, 1099–1109. [Google Scholar] [CrossRef]
- Rizwan, M.; Meunier, J.D.; Helene, M.; Keller, C. Effect of silicon on reducing cadmium toxicity in durum wheat (Triticum turgidum L. cv. Claudio, W.) grown in a soil with aged contamination. J. Hazard. Mater. 2012, 209, 326–334. [Google Scholar] [CrossRef]
- Sen, A. Oxidative stress studies in plant tissue culture. In Biochemistry, Genetics and Molecular Biology “Antioxidant Enzyme”; El-Missiry, M.A., Ed.; World’s largest Science, Technology & Medicine Open Access book publisher (INTECH): London, UK, 2012; pp. 59–88. [Google Scholar]
- Gong, H.; Zhu, X.; Chen, K.; Wang, S.; Zhang, C. Silicon alleviates oxidative damage of wheat plants in pots under drought. Plant Sci. 2003, 169, 313–321. [Google Scholar] [CrossRef]
- An, T.; Gao, Y.; Kuang, Q.; Wu, Y.; Zaman, Q.; Zhang, Y.; Xu, B.; Chen, Y. Effect of silicon on morpho-physiological attributes, yield and cadmium accumulation in two maize genotypes with contrasting root system size and health risk assessment. Plant Soil 2022, 477, 117–134. [Google Scholar] [CrossRef]
- Rehman, M.Z.U.; Rizwan, M.; Rauf, A.; Ayub, M.; Ali, S.; Qayyum, M.F.; Waris, A.A.; Naeem, A.; Sanaullah, M. Split application of silicon in cadmium (Cd) spiked alkaline soil plays a vital role in decreasing Cd accumulation in rice (Oryza sativa L.) grains. Chemosphere 2019, 226, 454–462. [Google Scholar] [CrossRef]
- Silva, A.J.; Nascimento, C.W.A.; Gouveia-Neto, A.S. Assessment of cadmium phytotoxicity alleviation by silicon using chlorophyll a fluorescence. Photosynthetica 2017, 55, 648–654. [Google Scholar] [CrossRef]
- Khan, Z.S.; Rizwan, M.; Hafeez, M.; Ali, S.; Adrees, M.; Qayyum, M.F.; Khalid, S.; Rehman, M.Z.U.; Sarwar, M.A. Effects of silicon nanoparticles on growth and physiology of wheat in cadmium contaminated soil under different soil moisture levels. Environ. Sci. Pollut. Res. 2020, 27, 4958–4968. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Platts-Mills, J.A.; Juma, J.; Kabir, F.; Nkeze, J.; Okoi, C.; Operario, D.J.; Uddin, J.; Ahmed, S.; Alonso, P.L. Use of quantitative molecular diagnostic methods to identify causes of diarrhoea in children: A reanalysis of the GEMS case-control study. Lancet 2016, 388, 1291–1301. [Google Scholar] [CrossRef]
- Mahmood, A.; Malik, R.N. Human health risk assessment of heavy metals via consumption of contaminated vegetables collected from different irrigation sources in Lahore, Pakistan. Arab. J. Chem. 2014, 7, 91–99. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experimental Treatments | HRI Adults | HRI Children | ||
|---|---|---|---|---|
| Control (0 mg kg−1) | Sodium Silicate | 0.00 mM | 0.11 | 0.20 |
| 0.50 mM | 0.08 | 0.15 | ||
| 1.00 mM | 0.03 | 0.05 | ||
| 1.50 mM | 0.05 | 0.10 | ||
| Potassium Silicate | 0.00 mM | 0.11 | 0.21 | |
| 0.50 mM | 0.05 | 0.10 | ||
| 1.00 mM | 0.02 | 0.03 | ||
| 1.50 mM | 0.04 | 0.07 | ||
| Cd Stress (20 mg kg−1) | Sodium Silicate | 0.00 mM | 0.51 | 0.95 |
| 0.50 mM | 0.42 | 0.79 | ||
| 1.00 mM | 0.35 | 0.65 | ||
| 1.50 mM | 0.45 | 0.83 | ||
| Potassium Silicate | 0.00 mM | 0.53 | 0.98 | |
| 0.50 mM | 0.39 | 0.73 | ||
| 1.00 mM | 0.30 | 0.57 | ||
| 1.50 mM | 0.41 | 0.75 | ||
| Variables | CAT | CC | EL | LA | POD | PR | RDW | RWC | SC | SDW |
|---|---|---|---|---|---|---|---|---|---|---|
| CC | −0.72 ** | |||||||||
| EL | 0.85 ** | −0.86 ** | ||||||||
| LA | −0.46 ** | 0.65 ** | −0.35 * | |||||||
| POD | 0.92 ** | −0.77 ** | 0.87 ** | −0.47 ** | ||||||
| PR | −0.78 ** | 0.90 ** | −0.82 ** | 0.67 ** | −0.83 ** | |||||
| RDW | −0.73 ** | 0.94 ** | −0.76 ** | 0.77 ** | −0.74 ** | 0.89 ** | ||||
| RWC | −0.68 ** | 0.87 ** | −0.75 ** | 0.69 ** | −0.72 ** | 0.87 ** | 0.89 ** | |||
| SC | −0.78 ** | 0.95 ** | −0.91 ** | 0.61 ** | −0.84 ** | 0.90 ** | 0.88 ** | 0.88 ** | ||
| SDW | −0.65 ** | 0.88 ** | −0.71 ** | 0.66 ** | −0.64 ** | 0.79 ** | 0.93 ** | 0.78 ** | 0.79 ** | |
| SOD | 0.89 ** | −0.66 ** | 0.79 ** | −0.41 ** | 0.91 ** | −0.73 ** | −0.65 ** | −0.60 ** | −0.76 ** | −0.54 ** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Batool, T.; Javied, S.; Ashraf, K.; Sultan, K.; Zaman, Q.u.; Haider, F.U. Alleviation of Cadmium Stress by Silicon Supplementation in Peas by the Modulation of Morpho-Physio-Biochemical Variables and Health Risk Assessment. Life 2022, 12, 1479. https://doi.org/10.3390/life12101479
Batool T, Javied S, Ashraf K, Sultan K, Zaman Qu, Haider FU. Alleviation of Cadmium Stress by Silicon Supplementation in Peas by the Modulation of Morpho-Physio-Biochemical Variables and Health Risk Assessment. Life. 2022; 12(10):1479. https://doi.org/10.3390/life12101479
Chicago/Turabian StyleBatool, Tahira, Sabiha Javied, Kamran Ashraf, Khawar Sultan, Qamar uz Zaman, and Fasih Ullah Haider. 2022. "Alleviation of Cadmium Stress by Silicon Supplementation in Peas by the Modulation of Morpho-Physio-Biochemical Variables and Health Risk Assessment" Life 12, no. 10: 1479. https://doi.org/10.3390/life12101479