

Effects of Precipitation Change and Nitrogen and Phosphorus Additions on Traits and Abundance of Potentilla anserina in an Alpine Meadow
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Description
2.2. Experimental Design
2.3. Measurement of Plant Traits and Abundance
2.4. Statistical Analyses
3. Results
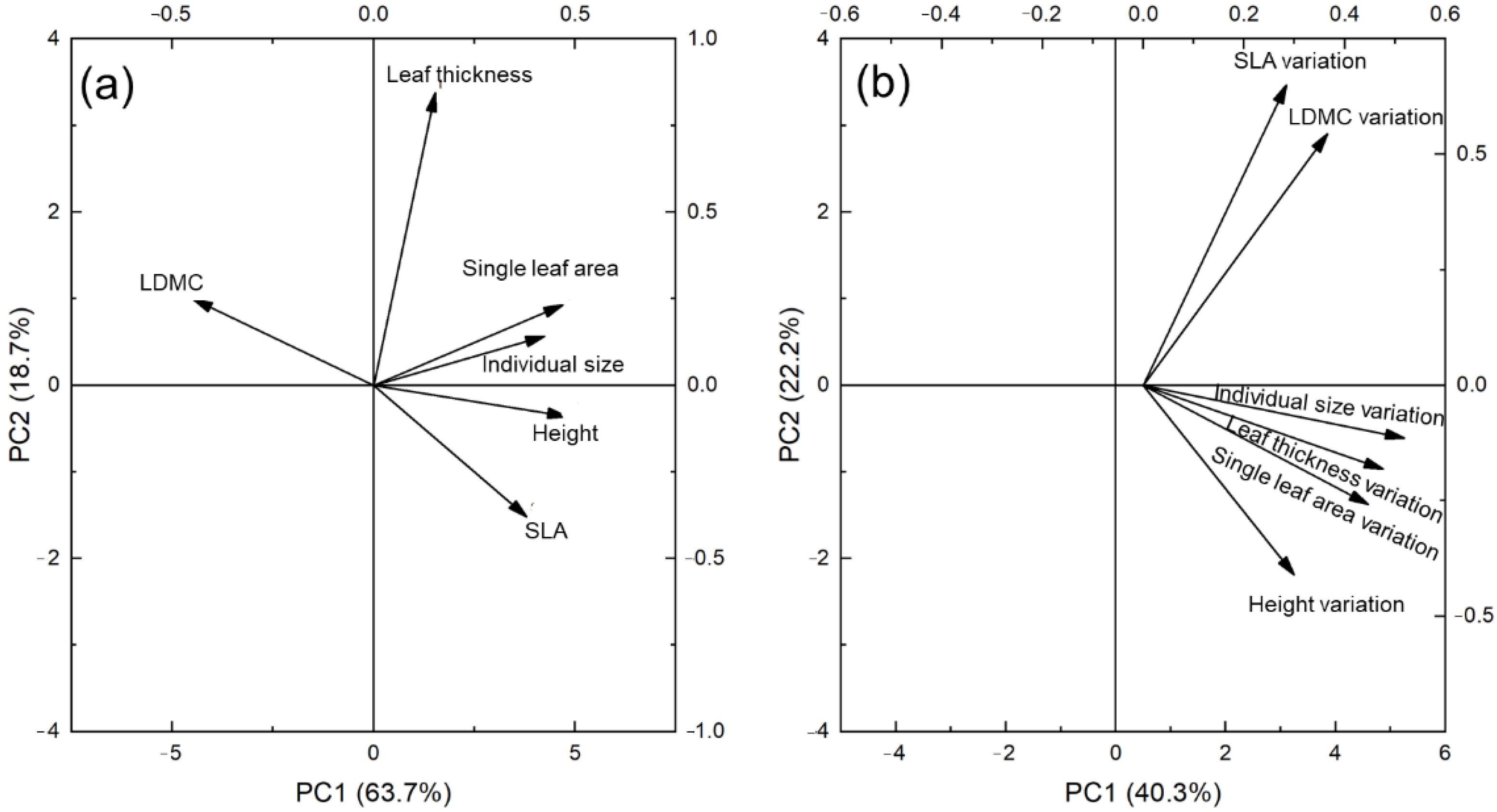
3.1. PCA Analysis of Traits and Intraspecific Trait Variation
3.2. Response of Traits to Precipitation Change, N Addition, P Addition and Their Interactions
3.3. Response of Intraspecific Variation of Traits and Plant Abundance to Precipitation Change, N Addition, P Addition and Their Interactions
3.4. Relationships of Plant Abundance with Traits and Their Intraspecific Variation
4. Discussion
4.1. Effects of Precipitation Change and N and P Addition on Traits
4.2. Effects of Precipitation Change and N and P Additions on Plant Abundance
4.3. Effects of Mean Value of Traits on Plant Abundance
4.4. Effects of Intraspecific Trait Variation on Plant Abundance
4.5. Limitation of P on Traits of Potentilla Anserina
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chen, H.; Zhu, Q.; Peng, C.; Wu, N.; Wang, Y.; Fang, X.; Gao, Y.; Zhu, D.; Yang, G.; Tian, J. The impacts of climate change and human activities on biogeochemical cycles on the Qinghai-Tibetan Plateau. Glob. Chang. Biol. 2013, 19, 2940–2955. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Willis, C.G.; Klein, J.A.; Ma, Z.; Li, J.; Zhou, H.; Zhao, X. Recovery of plant species diversity during long-term experimental warming of a species-rich alpine meadow community on the Qinghai-Tibet plateau. Biol. Conserv. 2017, 213, 218–224. [Google Scholar] [CrossRef]
- Yu, C.; Wang, J.; Shen, Z.; Fu, G. Effects of experimental warming and increased precipitation on soil respiration in an alpine meadow in the Northern Tibetan Plateau. Sci. Total Environ. 2019, 647, 1490–1497. [Google Scholar] [CrossRef] [PubMed]
- Fu, G.; Wang, J.; Li, W. Response of forage nutritional quality to climate change and human activities in alpine grasslands. Sci. Total Environ. 2022, 845, 157552. [Google Scholar] [CrossRef]
- Wang, Y.; Lv, W.; Xue, K.; Wang, S.; Zhang, L.; Hu, R.; Zeng, H.; Xu, X.; Li, Y.; Hao, Y.; et al. Grassland changes and adaptive management on the Qinghai–Tibetan Plateau. Nat. Rev. Earth Environ. 2022, 3, 668–683. [Google Scholar] [CrossRef]
- Yang, Y.H.; Park, S.-L. Vegetation cover changes in grasslands on the Qinghai-Tibet Plateau and their relationship with climate factors. J. Plant Ecol. 2006, 30, 1–8. [Google Scholar] [CrossRef]
- Wang, J.; Yu, C.; Fu, G. Warming reconstructs the elevation distributions of aboveground net primary production, plant species and phylogenetic diversity in alpine grasslands. Ecol. Indic. 2021, 133, 108355. [Google Scholar] [CrossRef]
- Wang, J.; Li, M.; Yu, C.; Fu, G. The change in environmental variables linked to climate change has a stronger effect on aboveground net primary productivity than does phenological change in alpine grasslands. Front. Plant Sci. 2022, 12, 798633. [Google Scholar] [CrossRef]
- Editorial Committee of the Flora of China, Chinese Academy of Sciences. Flora of China; Science Press: Beijing, China, 1993; Volume 37, p. 275. [Google Scholar]
- Tan, L.; Li, J.; Li, Y.; Wang, H.; Bao, X.; Zhao, J.; Ma, J.; Ji, T.; Wang, H. Nutrient composition analysis and quality evaluation of Potentilla anserine from different habitats in Qinghai. J. Food Sci. Biotechnol. 2022, 41, 95–111. [Google Scholar] [CrossRef]
- Xiong, Y.; Diao, Z.; Wu, B.; Li, Q. Resources of Potentilla anserina in Qinghai Grasslands and its development and application value. Qinghai Pratacult. 2004, 4, 22–26. [Google Scholar] [CrossRef]
- Zhou, H.K.; Zhao, X.Q.; Zhou, L.; Liu, W.; Han, F.; Gu, S. Effects of alpine meadow degradation on the growth characteristics of Potentilla anserina clones. Acta Ecol. Sin. 2006, 2, 508–520. [Google Scholar] [CrossRef]
- Díaz, S.; Kattge, J.; Cornelissen, J.H.; Wright, I.J.; Lavorel, S.; Dray, S.; Reu, B.; Kleyer, M.; Wirth, C.; Prentice, I.C.; et al. The global spectrum of plant form and function. Nature 2016, 529, 167–171. [Google Scholar] [CrossRef]
- Wright, I.J.; Reich, P.B.; Westoby, M.; Ackerly, D.D.; Baruch, Z.; Bongers, F.; Cavender-Bares, J.; Chapin, T.; Cornelissen, J.H.C.C.; Villar, R.; et al. The worldwide leaf economics spectrum. Nature 2004, 428, 821–827. [Google Scholar] [CrossRef] [PubMed]
- Migliavacca, M.; Musavi, T.; Mahecha, M.D.; Nelson, J.A.; Knauer, J.; Baldocchi, D.D.; Perez-Priego, O.; Christiansen, R.; Peters, J.; Anderson, K.; et al. The three major axes of terrestrial ecosystem function. Nature 2021, 598, 468–472. [Google Scholar] [CrossRef] [PubMed]
- Van der Plas, F.; Schröder-Georgi, T.; Weigelt, A.; Barry, K.; Meyer, S.; Alzate, A.; Wirth, C. Plant traits alone are poor predictors of ecosystem properties and long-term ecosystem functioning. Nat. Ecol. Evol. 2020, 4, 1602–1611. [Google Scholar] [CrossRef]
- Kovenock, M.; Swann, A.L. Leaf trait acclimation amplifies simulated climate warming in response to elevated carbon dioxide. Glob. Biogeochem. Cycles 2018, 32, 1437–1448. [Google Scholar] [CrossRef]
- Xing, X.; Zhang, L.; Bao, Y.; Yang, Z. Effect of grazing on the aboveground functional traits of Cleistogenes squarrosa. Chin. J. Grassl. 2019, 41, 116–122. [Google Scholar] [CrossRef]
- Zhang, C.; Cadotte, M.W.; Chiarucci, A.; Loreau, M.; Willis, C.G.; Si, X.; Li, L.; Cianciaruso, M.V. Scale-dependent shifts in functional and phylogenetic structure of Mediterranean island plant communities over two centuries. J. Ecol. 2021, 109, 3513–3523. [Google Scholar] [CrossRef]
- Fu, G.; Shen, Z.X. Response of alpine plants to nitrogen addition on the Tibetan Plateau: A meta-analysis. J. Plant Growth Regul. 2016, 35, 974–979. [Google Scholar] [CrossRef]
- Liang, X.S.; Ma, W.; Wang, H.Y. Responses of functional traits of three plants to addition in Hulunbuir grassland. Chin. J. Grassl. 2019, 41, 61–67. [Google Scholar] [CrossRef]
- Jung, V.; Albert, C.H.; Violle, C.; Kunstler, G.; Loucougaray, G.; Spiegelberger, T. Intraspecific trait variability mediates the response of subalpine grassland communities to extreme drought events. J. Ecol. 2014, 102, 45–53. [Google Scholar] [CrossRef] [Green Version]
- Hu, Z.; Yu, G.; Fan, J.; Zhong, H.; Wang, S.; Li, S. Precipitation-use efficiency along a 4500-km grassland transect. Glob. Ecol. Biogeogr. 2010, 19, 842–851. [Google Scholar] [CrossRef]
- Zhang, C.; Ma, Z.; Zhou, H.; Zhao, X. Long-term warming results in species-specific shifts in seed mass in alpine communities. PeerJ 2019, 7, e7416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pérez, H.N.; Díaz, S.; Garnier, E.; Lavore, S.; Poorter, H.; Jaureguiberry, P.; Bret-Harte, M.S.; Cornwell, W.K.; Craine, J.M.; Gurvich, D.E.; et al. New handbook for standardised measurement of plant functional traits worldwide. Aust. J. Bot. 2013, 61, 167–234. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022; Available online: https://www.R-project.org/ (accessed on 1 March 2022).
- Suding, K.N.; Collins, S.L.; Gough, L.; Clark, C.; Cleland, E.E.; Gross, K.L.; Milchunas, D.G.; Pennings, S. Functional- and abundance-based mechanisms explain diversity loss due to N fertilization. Proc. Natl. Acad. Sci. USA 2005, 102, 4387–4392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sherry, R.A.; Weng, E.; Arnone III, J.A.; Johnson, D.W.; Schimel, D.S.; Verburc, P.S.; Wallace, L.L.; Luo, Y. Lagged effects of experimental warming and doubled precipitation on annual and seasonal aboveground biomass production in a tallgrass prairie. Glob. Chang. Biol. 2008, 14, 2923–2936. [Google Scholar] [CrossRef]
- Fu, G.; Shen, Z.; Zhang, X. Increased precipitation has stronger effects on plant production of an alpine meadow than does experimental warming in the Northern Tibetan Plateau. Agric. For. Meteorol. 2018, 249, 11–21. [Google Scholar] [CrossRef]
- Fang, Y.; Xun, F.; Bai, W.; Zhang, W.; Li, L. Long-term nitrogen addition leads to loss of species richness due to litter accumulation and soil acidification in a temperate steppe. PLoS ONE 2012, 7, e47369. [Google Scholar] [CrossRef]
- Niu, S.; Wu, M.; Han, Y.I.; Xia, J.; Zhang, Z.H.E.; Yang, H.; Wan, S. Nitrogen effects on net ecosystem carbon exchange in a temperate steppe. Glob. Chang. Biol. 2010, 16, 144–155. [Google Scholar] [CrossRef]
- Mo, J.M.; Peng, S.L.; Fang, Y.T.; Yu, M.D.; Kong, G.H.; Zhang, Y.C. A preliminary study on the dynamics of bio-available nitrogen in soils of pine-broadleaf mixed forest in Dinghushan Biosphere Reserve. Acta Ecol. Sin. 2001, 21, 492–497. [Google Scholar] [CrossRef]
- Chinese Journal of Plant Ecology. Available online: https://www.plant-ecology.com/EN/Y2001/V25/I2/210#goTop (accessed on 30 June 2022).
- Su, B.; Han, X.; Qu, C.; Li, G. Factors affecting soil N availability in forest ecosystems: A literature review. Chin. J. Ecol. 2002, 21, 40–46. [Google Scholar] [CrossRef]
- Wang, X.; Chen, G.; Wang, Y.; Zhao, T.; Zhao, M. Response of stocking rates on the functional traits of Stipa breviflora desert steppe. Chin. J. Grassl. 2017, 39, 40–46. [Google Scholar] [CrossRef]
- Yang, H.; Li, Y.; Wu, M.; Zhang, Z.H.E.; Li, L.; Wan, S. Plant community responses to nitrogen addition and increased precipitation: The importance of water availability and species traits. Glob. Chang. Biol. 2011, 17, 2936–2944. [Google Scholar] [CrossRef]
- Dickson, T.L.; Mittelbach, G.G.; Reynolds, H.L.; Gross, K.L. Height and clonality traits determine plant community responses to fertilization. Ecology 2014, 95, 2443–2452. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Willis, C.G.; Burghardt, L.T.; Qi, W.; Liu, K.; de Moura Souza-Filho, P.R.; Ma, Z.; Du, G. The community-level effect of light on germination timing in relation to seed mass: A source of regeneration niche differentiation. New Phytol. 2014, 204, 496–506. [Google Scholar] [CrossRef]
- Albert, C.H.; Thuiller, W.; Yoccoz, N.G.; Soudant, A.; Boucher, F.; Saccone, P.; Lavorel, S. Intraspecific functional variability: Extent, structure and sources of variation. J. Ecol. 2010, 98, 604–613. [Google Scholar] [CrossRef]
- Lecerf, A.; Chauvet, E. Intraspecific variability in leaf traits strongly affects alder leaf decomposition in a stream. Basic Appl. Ecol. 2008, 9, 598–605. [Google Scholar] [CrossRef] [Green Version]
- Yu, H.; Pan, Z.; Chen, Z.; Chen, Z.; Feng, L. Effects of nitrogen addition on soil chemical properties and leaf functional traits of Dicranopteris dichotoma in the red soil erosion area of southern China. Chin. J. Appl. Environ. Biol. 2020, 26, 46–53. [Google Scholar] [CrossRef]
- Lü, X.T.; Reed, S.; Yu, Q.; He, N.; Wang, Z.; Han, X. Convergent responses of nitrogen and phosphorus resorption to nitrogen inputs in a semiarid grassland. Glob. Chang. Biol. 2013, 19, 2775–2784. [Google Scholar] [CrossRef]
- Mooney, H.A.; Vitousek, P.M.; Matson, P.A. Exchange of materials between terrestrial ecosystems and the atmosphere. Science 1987, 238, 926–932. [Google Scholar] [CrossRef]
- Du, E.; Terrer, C.; Pellegrini, A.F.; Ahlström, A.; van Lissa, C.J.; Zhao, X.; Xin, N.; Wu, X.; Jackson, R.B. Global patterns of terrestrial nitrogen and phosphorus limitation. Nat. Geosci. 2020, 13, 221–226. [Google Scholar] [CrossRef]
- Reed, S.C.; Yang, X.; Thornton, P.E. Incorporating phosphorus cycling into global modeling efforts: A worthwhile, tractable endeavor. New Phytol. 2015, 208, 324–329. [Google Scholar] [CrossRef] [PubMed]
- Cui, E.; Lu, R.; Xu, X.; Sun, H.; Qiao, Y.; Ping, J.; Qiu, S.; Lin, Y.; Bao, J.; Yong, Y.; et al. Soil phosphorus drives plant trait variations in a mature subtropical forest. Glob. Chang. Biol. 2022, 28, 3310–3320. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 50% Precipitation Reduction (W−) | No Change (W0) | 50% Precipitation Addition (W+) | |
|---|---|---|---|
| No addition (N0P0) | W−N0P0 | W0N0P0 | W+N0P0 |
| 10 g N m−2 year−1 (N+P0) | W−N+P0 | W0N+P0 | W+N+P0 |
| 10 g P m−2 year−1 (N0P+) | W−N0P+ | W0N0P+ | W+N0P+ |
| 10 g N m−2 year−1+ 10 g P m−2 year−1 (N+P+) | W−N+P+ | W0N+P+ | W+N+P+ |
| Source of Variation | W | N | P | W × N | W × P | N × P | W × N × P |
|---|---|---|---|---|---|---|---|
| SLA | 9.608 *** | 1.782 | 40.203 *** | 0.225 | 0.944 | 1.204 | 0.909 |
| LDMC | 12.234 *** | 10.093 ** | 54.556 *** | 1.423 | 2.733 | 0.130 | 0.558 |
| Single leaf area | 10.501 *** | 26.468 *** | 34.617 *** | 0.575 | 2.054 | 0.833 | 1.146 |
| Leaf thickness | 0.909 | 0.793 | 0.112 | 0.867 | 0.002 | 3.360 | 1.561 |
| Height | 24.289 *** | 41.334 *** | 126.907 *** | 0.101 | 2.400 | 3.540 | 0.807 |
| Individual size | 7.542 ** | 43.766 *** | 26.571 *** | 0.837 | 0.197 | 0.354 | 0.177 |
| Abundance | 35.918 *** | 11.643 ** | 7.700 * | 1.485 | 3.530 * | 0.217 | 0.361 |
| Source of Variation | W | N | P | W × N | W × P | N × P | W × N × P |
|---|---|---|---|---|---|---|---|
| SLA variation | 0.693 | 0.109 | 4.916 * | 1.177 | 0.627 | 0.159 | 0.043 |
| LDMC variation | 0.407 | 1.302 | 0.576 | 1.126 | 0.394 | 1.918 | 0.263 |
| Single leaf area variation | 1.496 | 0.002 | 0.440 | 0.412 | 0.265 | 0.007 | 0.601 |
| Leaf thickness variation | 0.296 | 0.126 | 1.840 | 0.473 | 0.435 | 0.474 | 1.794 |
| Height variation | 2.773 | 2.427 | 0.639 | 0.041 | 0.117 | 2.359 | 0.513 |
| Individual size variation | 0.072 | 1.667 | 5.561 * | 1.661 | 1.332 | 1.720 | 2.781 |
| Source of Variation | Slope | Intercept | R2 | p |
|---|---|---|---|---|
| SLA variation | −0.231 | 0.853 | 0.007 | 0.5247 |
| LDMC variation | −0.231 | 0.836 | 0.005 | 0.5788 |
| Single leaf area variation | 0.181 | 1.224 | 0.003 | 0.6584 |
| Leaf thickness variation | 0.329 | 1.475 | 0.011 | 0.4339 |
| Height variation | −0.575 | 0.614 | 0.051 | 0.0841 |
| Individual size variation | 0.181 | 1.224 | 0.003 | 0.6584 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, L.; Ren, Y.; Wan, J.-Z.; Wang, M.; Wang, Z.; Fu, F.; Sun, J.; Fu, Y.; Ma, Z.; Zhang, C. Effects of Precipitation Change and Nitrogen and Phosphorus Additions on Traits and Abundance of Potentilla anserina in an Alpine Meadow. Atmosphere 2022, 13, 1820. https://doi.org/10.3390/atmos13111820
Wu L, Ren Y, Wan J-Z, Wang M, Wang Z, Fu F, Sun J, Fu Y, Ma Z, Zhang C. Effects of Precipitation Change and Nitrogen and Phosphorus Additions on Traits and Abundance of Potentilla anserina in an Alpine Meadow. Atmosphere. 2022; 13(11):1820. https://doi.org/10.3390/atmos13111820
Chicago/Turabian StyleWu, Lin, Yanmei Ren, Ji-Zhong Wan, Mengyan Wang, Zuoyi Wang, Feiyan Fu, Jianping Sun, Yanjin Fu, Zhen Ma, and Chunhui Zhang. 2022. "Effects of Precipitation Change and Nitrogen and Phosphorus Additions on Traits and Abundance of Potentilla anserina in an Alpine Meadow" Atmosphere 13, no. 11: 1820. https://doi.org/10.3390/atmos13111820