
Biotransformation of High Concentrations of Ginsenoside Substrate into Compound K by β-glycosidase from Sulfolobus solfataricus
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Construction of the Expression Strain
2.3. Preparation of Crude Enzyme
2.4. Assay of β-glycosidase Activity
2.5. Enzyme Reactions and HPLC Analysis of CK
2.6. Effects of pH, Temperature, and Metal Ions on CK Production
2.7. Effects of Substrate and Enzyme Concentrations on CK Production
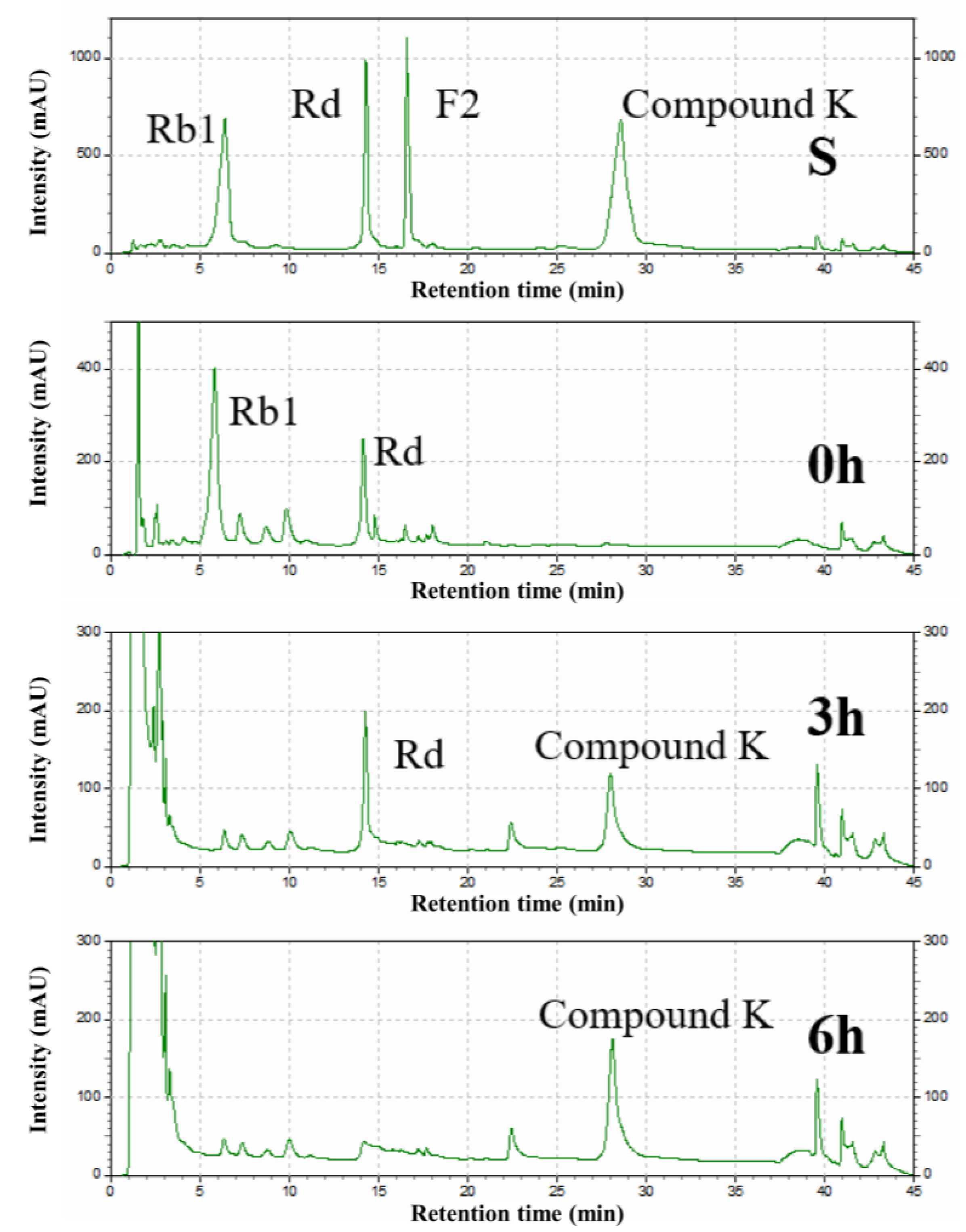
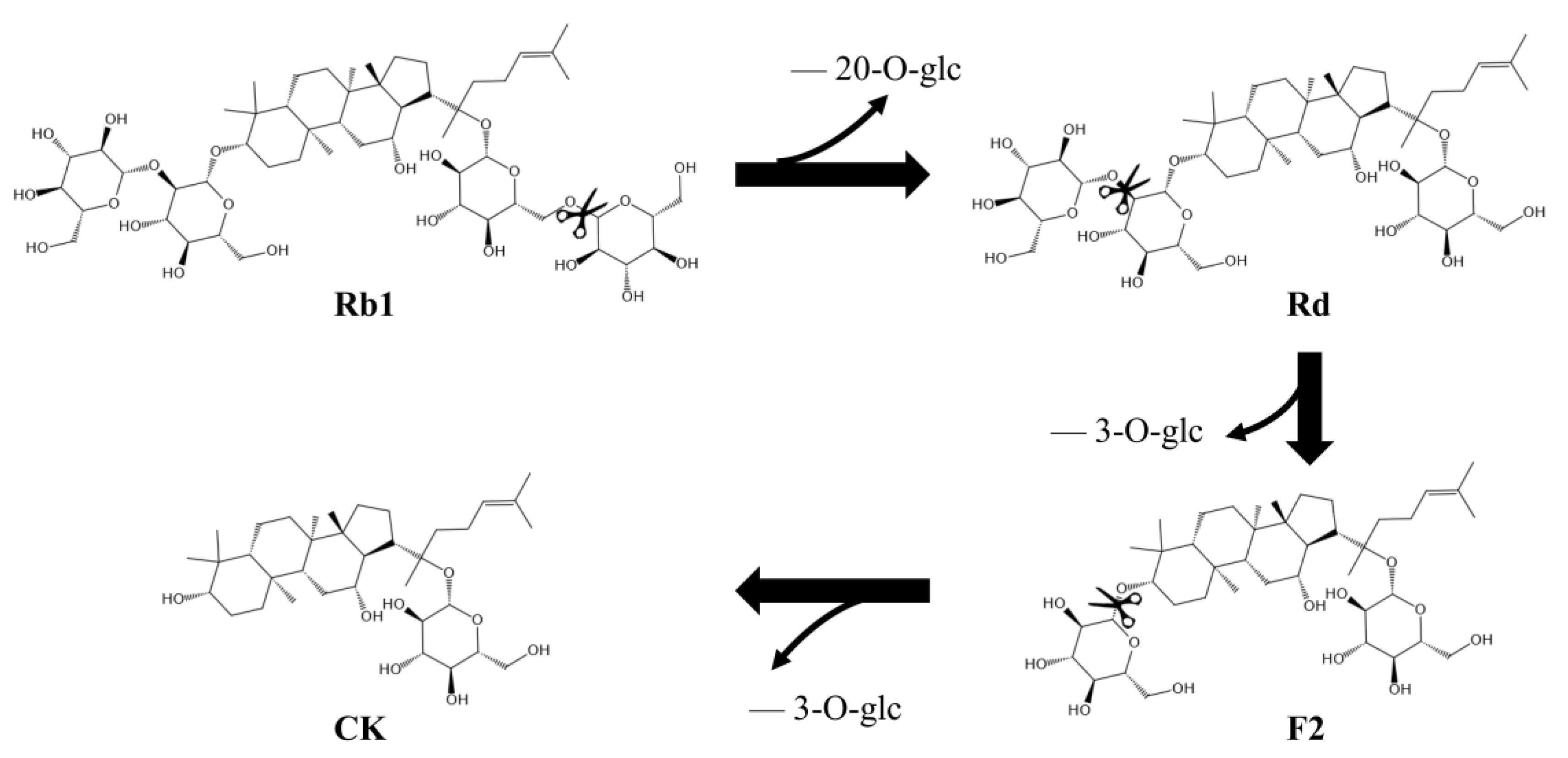
2.8. Analysis of Biotransformation Pathways
3. Results
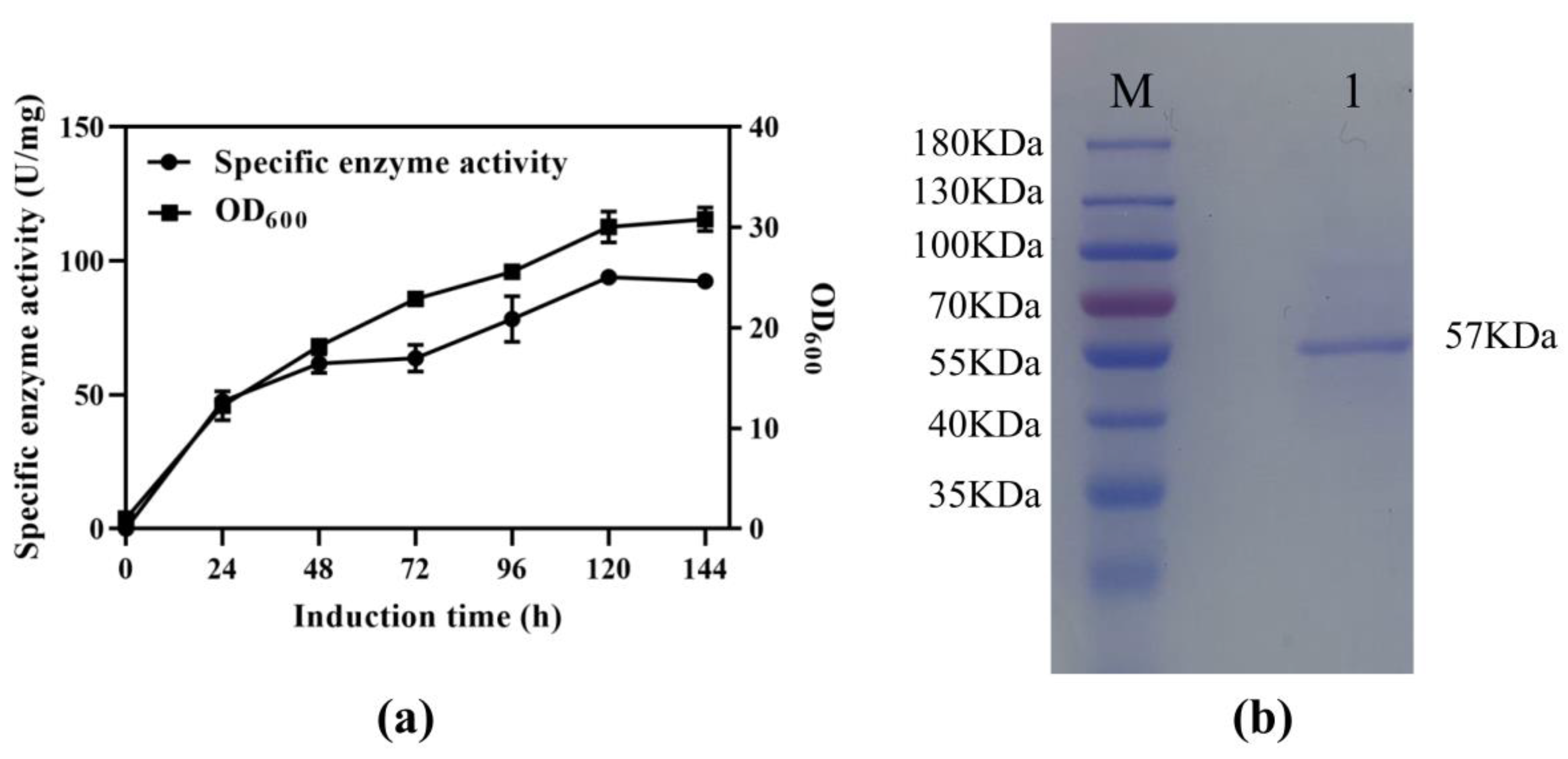
3.1. Expression and Analysis of Recombinant SS-bgly in P. pastoris
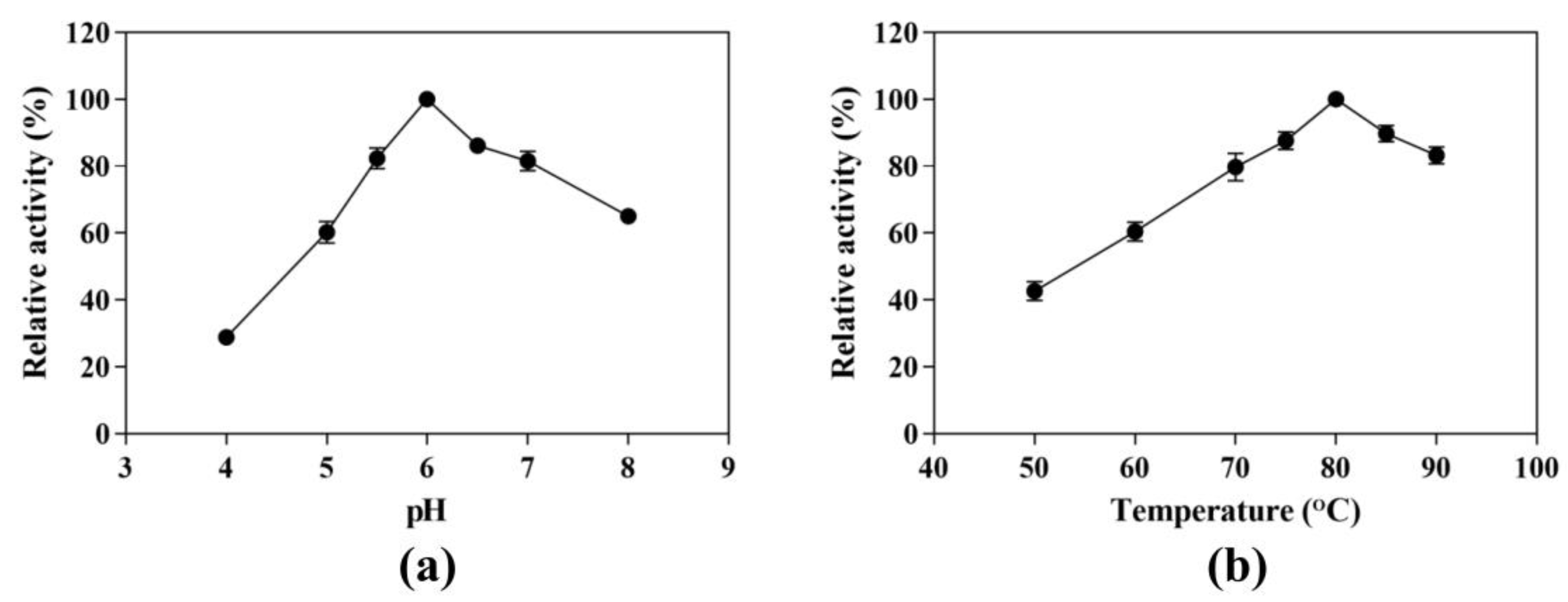
3.2. Effects of pH and Temperature on CK-Producing Activity
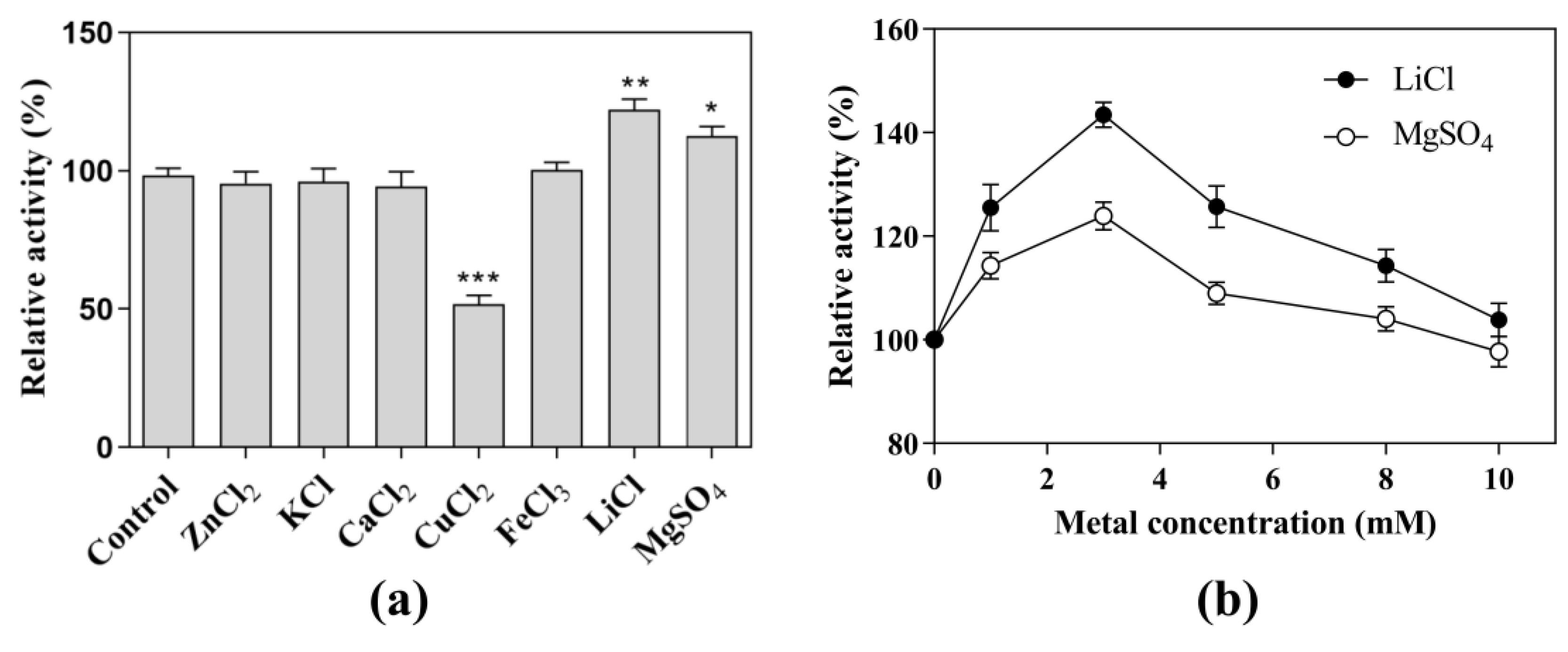
3.3. Effects of Metal Ions on CK-Producing Activity
3.4. Effects of Substrate Concentrations on CK Production
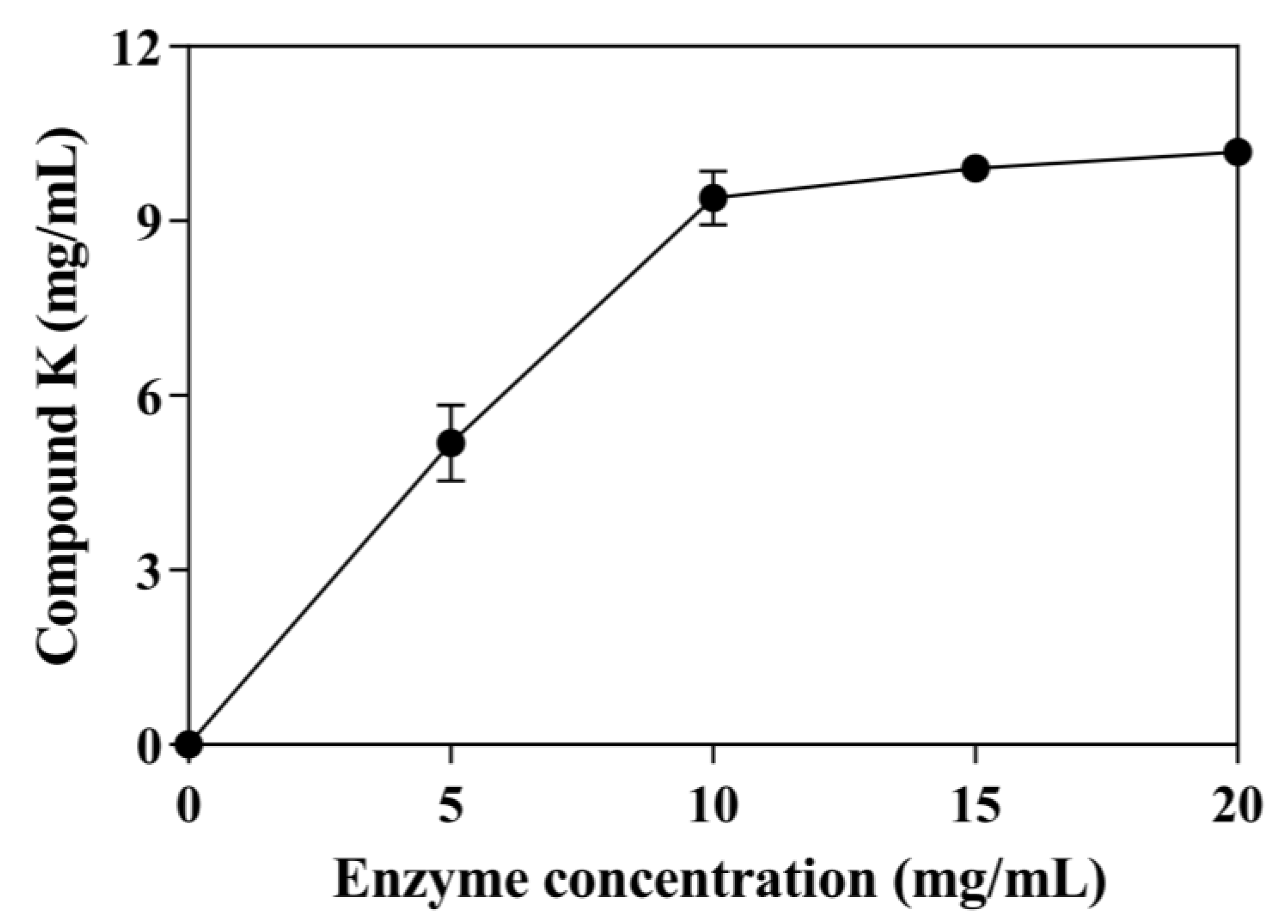
3.5. Effects of Enzyme Concentrations on CK Production
3.6. Determination of the Biotransformation Pathway of Recombinant SS-bgly
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fan, W.X.; Huang, Y.L.; Zheng, H.; Li, S.Q.; Li, Z.H.; Li, Y.; Cheng, X.; He, C.S.; Sun, J.F. Ginsenosides for the treatment of metabolic syndrome and cardiovascular diseases: Pharmacology and mechanisms. Biomed. Pharm. 2020, 132, 110915. [Google Scholar] [CrossRef] [PubMed]
- Duan, Z.G.; Wei, B.; Deng, J.J.; Mi, Y.; Dong, Y.F. The anti-tumor effect of ginsenoside Rh4 in MCF-7 breast cancer cells in vitro and in vivo. Biochem. Biophys. Res. Commun. 2018, 499, 482–487. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.; Seo, Y.J.; Song, J.H.; Chei, S.W.; Lee, B.Y. Ginsenoside Rg1 promotes browning by inducing UCP1 expression and mitochondrial activity in 3T3-L1 and subcutaneous white adipocytes. J. Ginseng Res. 2019, 43, 589–599. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Yi, Y.S.; Kim, M.Y.; Cho, J.Y. Role of ginsenosides, the main active components of Panax ginseng, in inflammatory responses and diseases. J. Ginseng Res. 2017, 41, 435–443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mo, J.; Zhou, Y.L.; Yang, R.H.; Zhang, P.L.; He, B.; Yang, J.Y.; Li, S.D.; Shen, Z.Q.; Chen, P. Ginsenoside Rg1 ameliorates palmitic acid-induced insulin resistance in HepG2 cells in association with modulating Akt and JNK activity. Pharm. Rep. 2019, 71, 1160–1167. [Google Scholar] [CrossRef]
- Hossen, M.J.; Hong, Y.D.; Baek, K.S.; Yoo, S.; Hong, Y.H.; Kim, J.H.; Lee, J.O.; Kim, D.; Park, J.; Cho, J.Y. In vitro antioxidative and anti-inflammatory effects of the compound K-rich fraction BIOGF1K, prepared from Panax ginseng. J. Ginseng Res. 2017, 41, 43–51. [Google Scholar] [CrossRef] [Green Version]
- Li, C.W.; Deng, M.Z.; Gao, Z.J.; Dang, Y.Y.; Zheng, G.D.; Yang, X.J.; Chao, Y.X.; Cai, Y.F.; Wu, X.L. Effects of compound K, a metabolite of ginsenosides, on memory and cognitive dysfunction in db/db mice involve the inhibition of ER stress and the NLRP3 inflammasome pathway. Food Funct. 2020, 11, 4416–4427. [Google Scholar] [CrossRef]
- Hong, Y.H.; Kim, D.; Nam, G.; Yoo, S.; Han, S.Y.; Jeong, S.G.; Kim, E.; Jeong, D.; Yoon, K.; Kim, S.; et al. Photoaging protective effects of BIOGF1K, a compound-K-rich fraction prepared from Panax ginseng. J. Ginseng Res. 2018, 42, 81–89. [Google Scholar] [CrossRef]
- Kim, E.; Kim, D.; Yoo, S.; Hong, Y.H.; Han, S.Y.; Jeong, S.; Jeong, D.; Kim, J.H.; Cho, J.Y.; Park, J. The skin protective effects of compound K, a metabolite of ginsenoside Rb1 from Panax ginseng. J. Ginseng Res. 2018, 42, 218–224. [Google Scholar] [CrossRef]
- Yang, X.D.; Yang, Y.Y.; Ouyang, D.S.; Yang, G.P. A review of biotransformation and pharmacology of ginsenoside compound K. Fitoterapia 2015, 100, 208–220. [Google Scholar] [CrossRef]
- Hwang, C.R.; Lee, S.H.; Jang, G.Y.; Hwang, I.G.; Kim, H.Y.; Woo, K.S.; Lee, J.; Jeong, H.S. Changes in ginsenoside compositions and antioxidant activities of hydroponic-cultured ginseng roots and leaves with heating temperature. J. Ginseng Res. 2014, 38, 180–186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, C.P.; Gao, W.P.; Zhao, B.Z.; Cheng, L.Q. Optimization of the selective preparation of 20(R)-ginsenoside Rg3 catalyzed by D, L-tartaric acid using response surface methodology. Fitoterapia 2013, 84, 213–221. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.T.; Yang, M.; Song, Y.; Lu, Z.Q.; Zhang, J.Q.; Huang, H.L.; Guo, W. Microbial transformation of ginsenoside Rb1 by Acremonium strictum. Appl. Microbiol. Biot. 2008, 77, 1345–1350. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Huang, H.; Zhu, H.Y.; Zhou, P.; Shi, X.L. New metabolites from the biotransformation of ginsenoside Rb1 by Paecilomyces bainier sp.229 and activities in inducing osteogenic differentiation by Wnt/beta-catenin signaling activation. J. Ginseng Res. 2018, 42, 199–207. [Google Scholar] [CrossRef]
- Kim, T.H.; Yang, E.J.; Shin, K.C.; Hwang, K.H.; Park, J.S.; Oh, D.K. Enhanced Production of β-D-glycosidase and α-L-arabinofuranosidase in recombinant Escherichia coli in Fed-batch culture for the biotransformation of ginseng leaf extract to ginsenoside Compound K. Biotechnol. Bioproc. E 2018, 23, 183–193. [Google Scholar] [CrossRef]
- Liu, C.Y.; Zhou, R.X.; Sun, C.K.; Jin, Y.H.; Yu, H.S.; Zhang, T.Y.; Xu, L.Q.; Jin, F.X. Preparation of minor ginsenosides C-Mc, C-Y, F2, and C-K from American ginseng PPD-ginsenoside using special ginsenosidase type-I from Aspergillus niger g.848. J. Ginseng Res. 2015, 39, 221–229. [Google Scholar] [CrossRef] [Green Version]
- Wang, P.P.; Wang, J.L.; Zhao, G.P.; Yan, X.; Zhou, Z.H. Systematic optimization of the yeast cell factory for sustainable and high efficiency production of bioactive ginsenoside compound K. Synth. Syst. Biotechnol. 2021, 6, 69–76. [Google Scholar] [CrossRef]
- Li, D.; Wu, Y.; Zhang, C.; Sun, J.; Zhou, Z.; Lu, W. Production of triterpene ginsenoside Compound K in the non-conventional yeast Yarrowia lipolytica. J. Agric. Food Chem. 2019, 67, 2581–2588. [Google Scholar] [CrossRef]
- Xu, W.; Klumbys, E.; Ang, E.L.; Zhao, H.M. Emerging molecular biology tools and strategies for engineering natural product biosynthesis. Metab. Eng. Commun. 2020, 10, e00108. [Google Scholar] [CrossRef]
- Duan, Z.G.; Zhu, C.H.; Shi, J.J.; Fan, D.D.; Deng, J.J.; Fu, R.Z.; Huang, R.; Fan, C.Y. High efficiency production of ginsenoside compound K by catalyzing ginsenoside Rb1 using snailase. Chin. J. Chem. Eng. 2018, 26, 1591–1597. [Google Scholar] [CrossRef]
- Kim, S.A.; Jeong, E.B.; Oh, D.K. Complete bioconversion of protopanaxadiol-type ginsenosides to Compound K by extracellular enzymes from the isolated strain Aspergillus tubingensis. J. Agric. Food. Chem. 2021, 69, 315–324. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.Y.; Li, W.N.; Fan, D.D. Biotransformation of ginsenoside Rb1 to ginsenoside CK by strain XD101: A safe bioconversion strategy. Appl. Biochem. Biotech. 2021, 193, 2110–2127. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.; Mi, Y.; Han, X.; Li, H.H.; Tian, M.; Duan, Z.G.; Fan, D.D.; Ma, P. Transformation of ginsenoside via deep eutectic solvents based on choline chloride as an enzymatic reaction medium. Bioproc. Biosyst. Eng. 2020, 43, 1195–1208. [Google Scholar] [CrossRef] [PubMed]
- Noh, K.H.; Son, J.W.; Kim, H.J.; Oh, D.K. Ginsenoside compound K production from ginseng root extract by a thermostable β-glycosidase from Sulfolobus solfataricus. Biosci. Biotechnol. Biochem. 2009, 73, 316–321. [Google Scholar] [CrossRef] [PubMed]
- Park, S.E.; Na, C.S.; Yoo, S.A.; Seo, S.H.; Son, H.S. Biotransformation of major ginsenosides in ginsenoside model culture by lactic acid bacteria. J. Ginseng Res. 2017, 41, 36–42. [Google Scholar] [CrossRef] [Green Version]
- Wang, R.; Yang, J.; Jin, M.J.; Liu, J.; Yuan, H.J.B.T. Efficient ferulic acid and xylo-oligosaccharides production by a novel multi-modular bifunctional xylanase/feruloyl esterase using agricultural residues as substrates. Bioresour. Technol. 2019, 297, 122487. [Google Scholar] [CrossRef]
- Kim, B.N.; Yeom, S.J.; Kim, Y.S.; Oh, D.K. Characterization of a beta-glucosidase from Sulfolobus solfataricus for isoflavone glycosides. Biotechnol. Lett. 2012, 34, 125–129. [Google Scholar] [CrossRef]
- Shin, K.C.; Choi, H.Y.; Seo, M.J.; Oh, D.K. Compound K production from red ginseng extract by beta-glycosidase from Sulfolobus solfataricus supplemented with alpha-L-arabinofuranosidase from Caldicellulosiruptor saccharolyticus. PLoS One. 2015, 10, e0145876. [Google Scholar] [CrossRef]
- Ye, L.; Zhang, C.; Li, J.; Shi, X.; Feng, M. Effects of external calcium on the biotransformation of ginsenoside Rb1 to ginsenoside Rd by Paecilomyces bainier 229-7. World J. Microbiol. Biotechnol. 2012, 28, 857–863. [Google Scholar] [CrossRef]
- Xie, J.C.; Zhao, D.X.; Zhao, L.G.; Pei, J.J.; Xiao, W.; Ding, G.; Wang, Z.Z. Overexpression and characterization of a Ca2+ activated thermostable β-glucosidase with high ginsenoside Rb1 to ginsenoside 20(S)-Rg3 bioconversion productivity. J. Ind. Microbiol. Biot. 2015, 42, 839–850. [Google Scholar] [CrossRef]
- Quan, L.H.; Min, J.W.; Jin, Y.; Wang, C.; Kim, Y.J.; Yang, D.C. Enzymatic biotransformation of ginsenoside Rb1 to compound K by recombinant beta-glucosidase from Microbacterium esteraromaticum. J. Agric. Food Chem. 2012, 60, 3776–3781. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.Z.; Yu, H.S.; Bao, Y.M.; An, L.J.; Jin, F.X. Purification and characterization of ginsenoside-β-glucosidase from ginseng. Chem. Pharm. Bull. 2001, 49, 795–798. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, I.S.; Wi, S.G.; Jung, S.R.; Patel, D.H.; Bae, H.J. Characterization and application of recombinant β-glucosidase (BglH) from Bacillus licheniformis KCTC 1918. J. Wood Sci. 2009, 55, 329–334. [Google Scholar] [CrossRef]
- Shin, K.C.; Choi, H.Y.; Seo, M.J.; Oh, D.K. Improved conversion of ginsenoside Rb1 to compound K by semi-rational design of Sulfolobus solfataricus β-glycosidase. Amb Expr 2017, 7, 186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, X.; Li, W.N.; Duan, Z.G.; Ma, X.X.; Fan, D.D. Biocatalytic production of compound K in a deep eutectic solvent based on choline chloride using a substrate fed-batch strategy. Bioresour. Technol. 2020, 305, 123039. [Google Scholar] [CrossRef]
- Jin, Z.; Han, S.Y.; Zhang, L.; Zheng, S.P.; Wang, Y.; Lin, Y. Combined utilization of lipase-displaying Pichia pastoris whole-cell biocatalysts to improve biodiesel production in co-solvent media. Bioresour. Technol. 2013, 130, 102–109. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Substrate Concentration (mg/mL) | CK Production (mg/mL) | Molar Conversion Rate (%) | Productivity (μM/h) | Reaction Time (h) |
|---|---|---|---|---|
| 10 | 3.79 ± 0.19 | 100.31 ± 5.17 | 507.06 ± 26.11 | 12 |
| 20 | 7.58 ± 0.46 | 100.41 ± 6.03 | 405.65 ± 24.37 | 30 |
| 30 | 9.39 ± 0.19 | 82.50 ± 6.03 | 314.07 ± 6.46 | 48 |
| 40 | 9.08 ± 0.36 | 59.83 ± 2.38 | 303.70 ± 12.07 | 48 |
| 50 | 8.59 ± 0.34 | 45.28 ± 1.81 | 287.31 ± 11.42 | 48 |
| Substrate | Rb1 | Rd | F2 |
|---|---|---|---|
| Km (mM) | 3.64 ± 0.28 | 5.08 ± 0.32 | 1.86 ± 0.05 |
| Kcat (s−1) | 20.03 ± 1.65 | 16.35 ± 2.26 | 82.42 ± 5.86 |
| Kcat/Km (s·mM)−1 | 6.25 ± 0.84 | 3.24 ± 0.25 | 42.58 ± 4.46 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, P.; Tang, C.; Liu, Y.; Yang, J.; Fan, D. Biotransformation of High Concentrations of Ginsenoside Substrate into Compound K by β-glycosidase from Sulfolobus solfataricus. Genes 2023, 14, 897. https://doi.org/10.3390/genes14040897
Wang P, Tang C, Liu Y, Yang J, Fan D. Biotransformation of High Concentrations of Ginsenoside Substrate into Compound K by β-glycosidase from Sulfolobus solfataricus. Genes. 2023; 14(4):897. https://doi.org/10.3390/genes14040897
Chicago/Turabian StyleWang, Pan, Congcong Tang, Yannan Liu, Jing Yang, and Daidi Fan. 2023. "Biotransformation of High Concentrations of Ginsenoside Substrate into Compound K by β-glycosidase from Sulfolobus solfataricus" Genes 14, no. 4: 897. https://doi.org/10.3390/genes14040897