
Reconstruction of a Comprehensive Interactome and Experimental Data Analysis of FRA10AC1 May Provide Insights into Its Biological Role in Health and Disease
 and
and 
Abstract
:1. Introduction
2. Material and Methods
2.1. Protein–Protein Interaction (PPI) Data Retrieval and Protein Network Reconstruction
2.2. GTEx Data Collection
2.3. HeLa Cell Line Culture and Transfection
2.4. Home-Made Antibodies
2.5. GST Pull-Down Assays
2.6. Co-Immunoprecipitation and Immunofluorescence
2.7. RNA In Situ Hybridization
3. Results
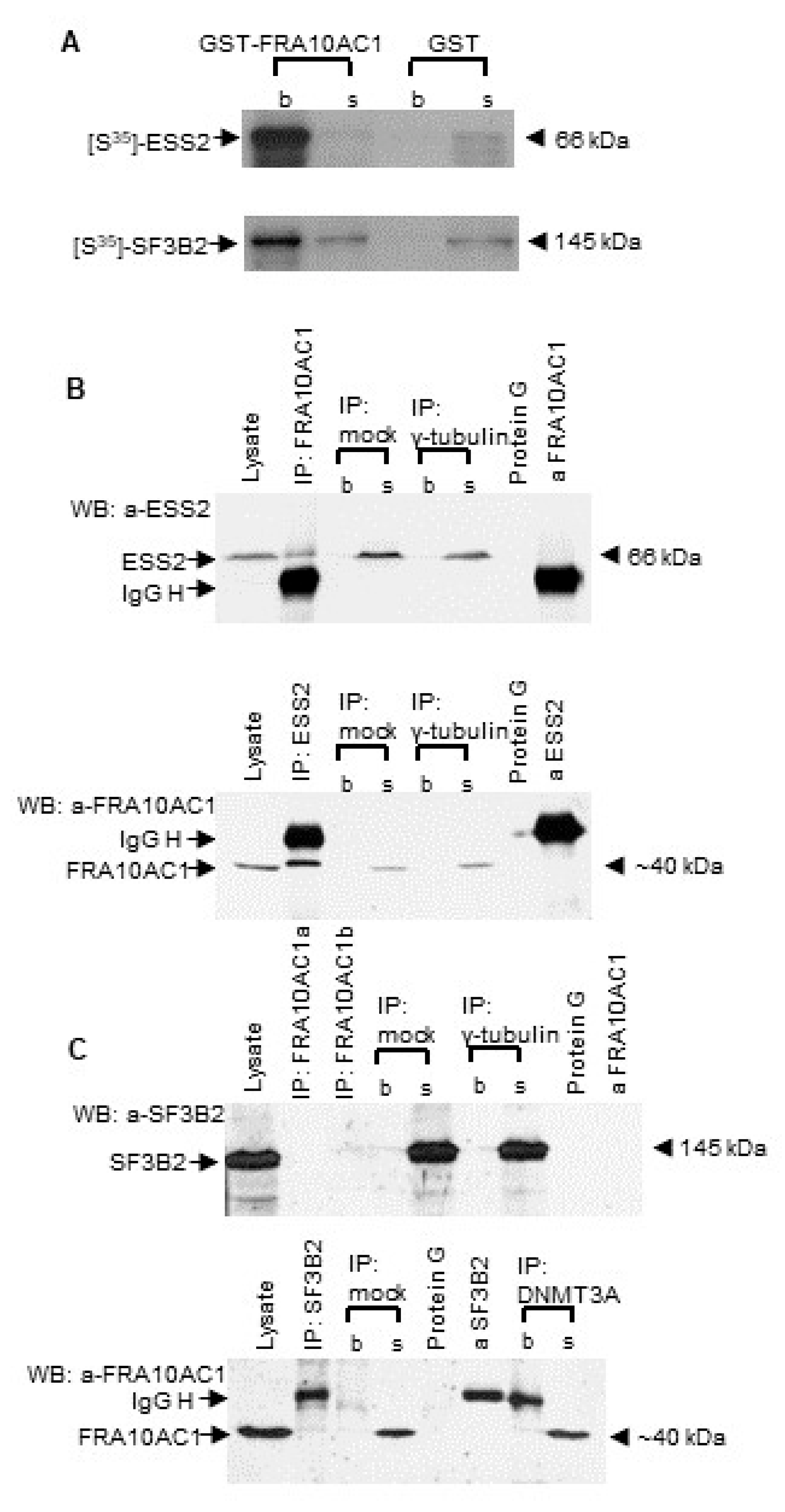
3.1. Validating FRA10AC1 Protein Interactions with SF3B2 and ESS2 Spliceosomal Proteins
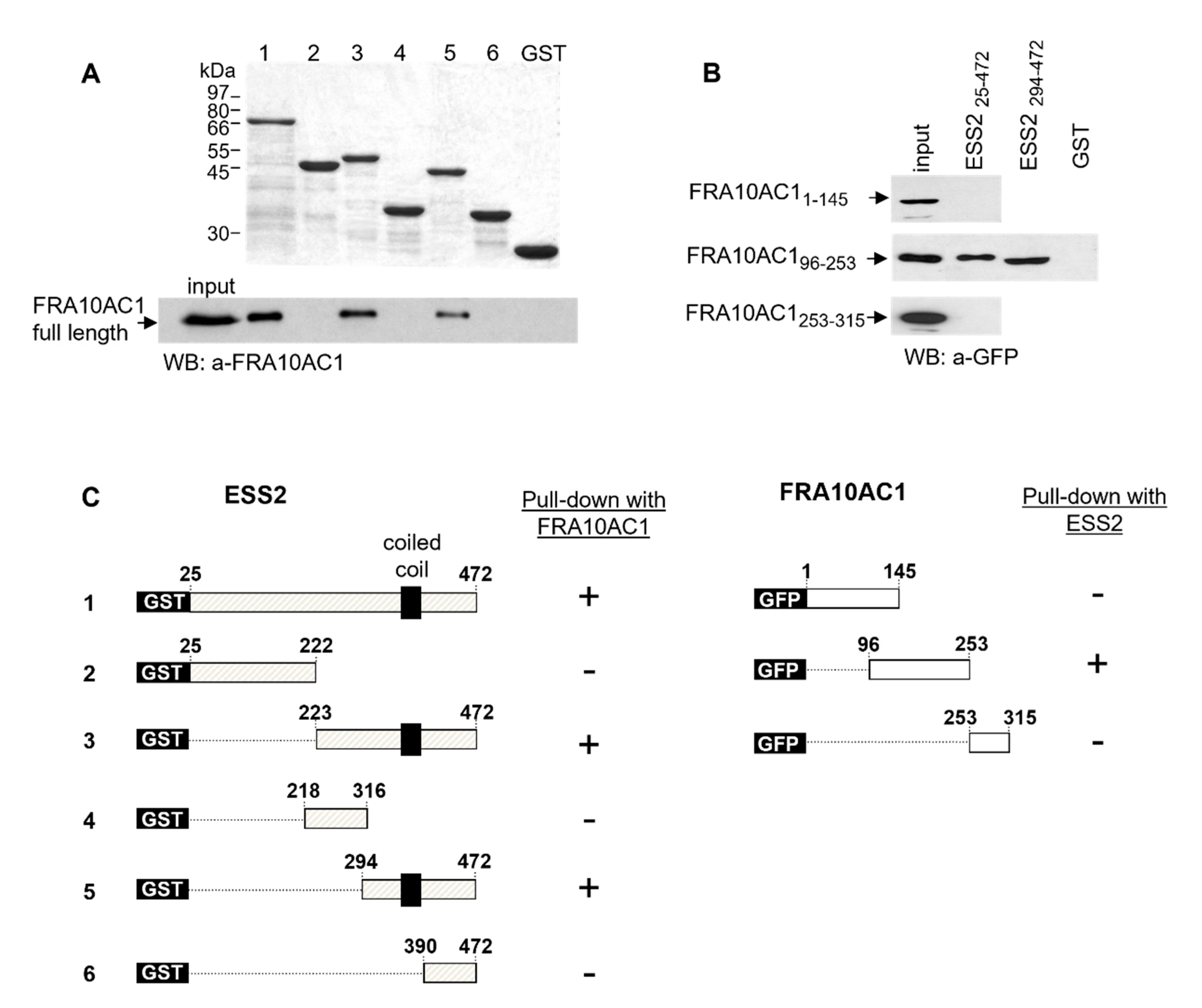
3.2. Mapping the FRA10AC1 Protein Interacting Region
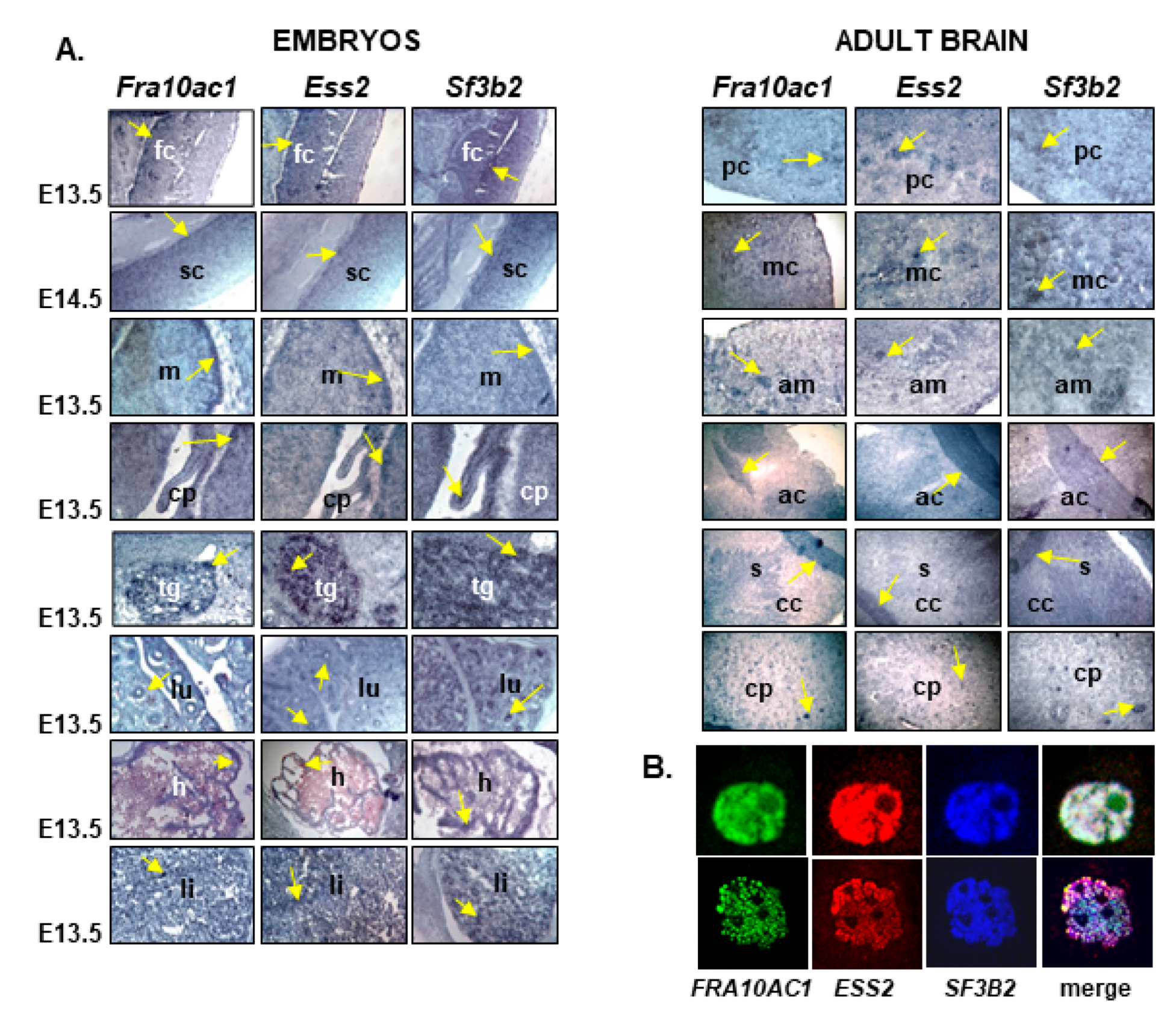
3.3. Fra10ac1, Ess2, and Sf3b2 Are Co-Expressed in the Mouse Embryo and Adult Mouse Tissue
3.4. FRA10AC1, ESS2, and SF3B2 Co-Localize in the HeLa Cell Nucleus
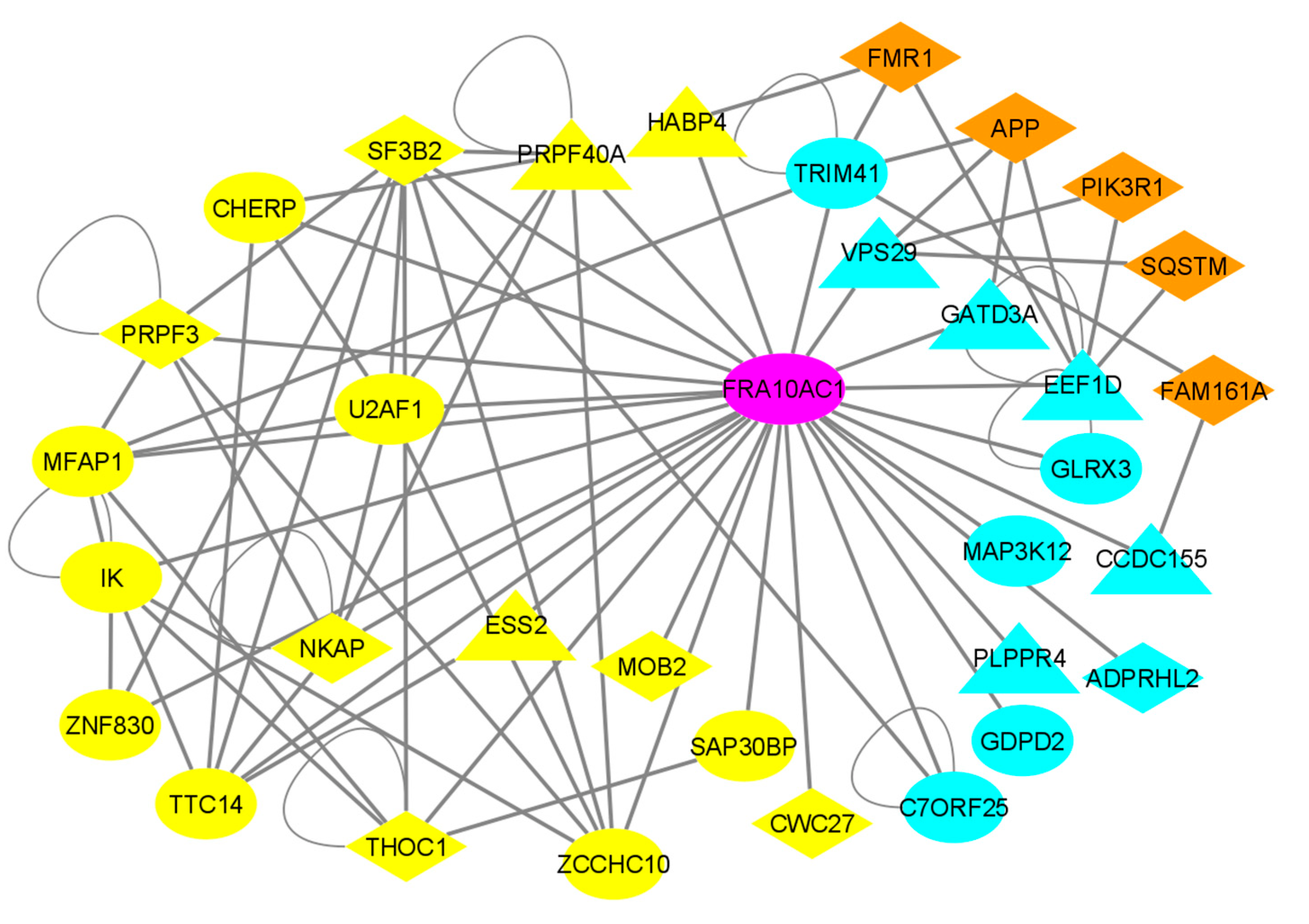
3.5. Reconstruction of the FRA10AC1 Protein Interactome
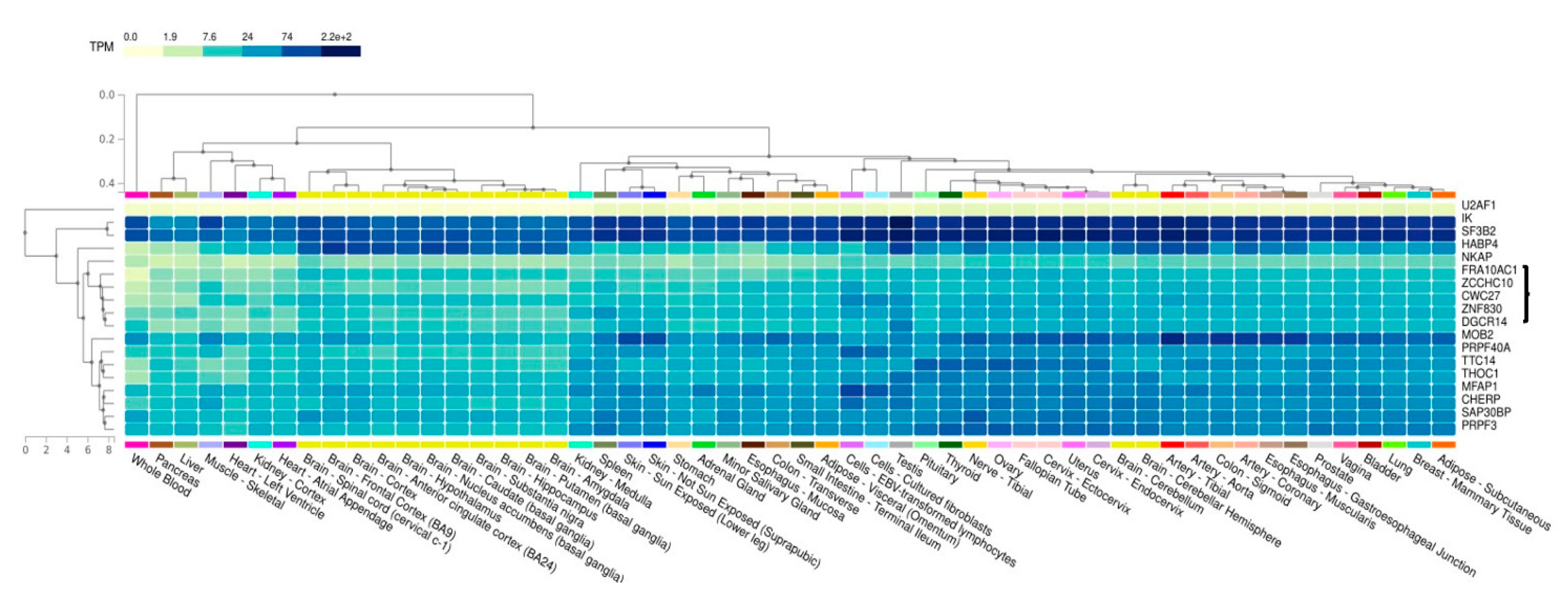
3.6. Co-Expression Analysis of FRA10AC1 Spliceosomal Interactors
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ruiz-Herrera, A.; Robinson, T.J. Chromosomal instability in Afrotheria: Fragile sites, evolutionary breakpoints and phylogenetic inference from genome sequence assemblies. BMC Evol. Biol. 2007, 7, 199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, X.; Supiwong, W.; Weise, A.; Mrasek, K.; Kosyakova, N.; Tanomtong, A.; Pinthong, K.; Trifonov, V.A.; Cioffi, M.D.B.; Grothmann, P.; et al. Comprehensive characterization of evolutionary conserved breakpoints in four New World Monkey karyotypes compared to Chlorocebus aethiops and Homo sapiens. Heliyon 2015, 1, e00042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glover, T.W.; Wilson, T.E.; Arlt, M.F. Fragile sites in cancer: More than meets the eye. Nat. Rev. Cancer 2017, 17, 489–501. [Google Scholar] [CrossRef]
- Kumar, R.; Nagpal, G.; Kumar, V.; Usmani, S.S.; Agrawal, P.; Raghava, G.P.S. HumCFS: A database of fragile sites in human chromosomes. BMC Genom. 2019, 19, 985. [Google Scholar] [CrossRef] [Green Version]
- Lokanga, R.A.; Kumari, D.; Usdin, K. Common Threads: Aphidicolin-Inducible and Folate-Sensitive Fragile Sites in the Human Genome. Front. Genet. 2021, 12, 708860. [Google Scholar] [CrossRef]
- Feng, W.; Chakraborty, A. Fragility Extraordinaire: Unsolved Mysteries of Chromosome Fragile Sites. DNA Replication Old Princ. New Discov. 2017, 1042, 489–526. [Google Scholar] [CrossRef]
- Sutherland, G.R. Heritable fragile sites on human chromosomes. VIII. Preliminary population cytogenetic data on the folic-acid-sensitive fragile sites. Am. J. Hum. Genet. 1982, 34, 452–458. [Google Scholar]
- Hecht, F.; Hecht, B.K. Autosomal fragile sites not a current indication for prenatal diagnosis. Hum. Genet. 1984, 67, 352–353. [Google Scholar] [CrossRef]
- Sutherland, G.R.; Hecht, F. Fragile Sites on Human Chromosomes; Oxford University Press: New York, NY, USA, 1985. [Google Scholar]
- Mavrou, A.; Syrrou, M.; Tsenghi, C.; Metaxotou, C. Autosomal folate sensitive fragile sites in normal and mentally retarded individuals in Greece. Am. J. Med. Genet. 1991, 38, 437–439. [Google Scholar] [CrossRef]
- Sitzmann, A.F.; Hagelstrom, R.T.; Tassone, F.; Hagerman, R.J.; Butler, M.G. Rare FMR1 gene mutations causing fragile X syndrome: A review. Am. J. Med. Genet. Part A 2017, 176, 11–18. [Google Scholar] [CrossRef]
- Banerjee, A.; Ifrim, M.F.; Valdez-Sinon, A.; Raj, N.; Bassell, G.J. Aberrant RNA translation in fragile X syndrome: From FMRP mechanisms to emerging therapeutic strategies. Brain Res. 2018, 1693, 24–36. [Google Scholar] [CrossRef] [PubMed]
- Richter, J.D.; Zhao, X. The molecular biology of FMRP: New insights into fragile X syndrome. Nat. Rev. Neurosci. 2021, 22, 209–222. [Google Scholar] [CrossRef] [PubMed]
- Sarafidou, T.; Moschonas, N.K. “Chromosome 10” Encyclopedia of Life Sciences; J. Wiley & Sons, Ltd.: Chichester, UK, 2017; Available online: https://onlinelibrary.wiley.com/action/doSearch?AllField=chromosome+10&ContentGroupKey=10.1002%2F047001590X (accessed on 6 March 2017).
- Sutherland, G.R. Heritable fragile sites on human chromosomes XII. Population cytogenetics. Ann. Hum. Genet. 1985, 49, 153–161. [Google Scholar] [CrossRef] [PubMed]
- Petit, P.; Fryns, J.P.; Berghe, H.V.D.; Hecht, F. Population cytogenetics of autosomal fragile sites. Clin. Genet. 2008, 29, 96–100. [Google Scholar] [CrossRef]
- Alitalo, T.; Matilainen, R.; Kaski, M.; Airaksinen, E. Population cytogenetics of folate-sensitive fragile sites. Hum. Genet. 1989, 82, 3–8. [Google Scholar] [CrossRef]
- Sarafidou, T.; Kahl, C.; Garay, I.M.; Mangelsdorf, M.; Gesk, S.; Baker, E.; Kokkinaki, M.; Talley, P.; Maltby, E.L.; French, L.; et al. Folate-sensitive fragile site FRA10A is due to an expansion of a CGG repeat in a novel gene, FRA10AC1, encoding a nuclear protein. Genomics 2004, 84, 69–81. [Google Scholar] [CrossRef]
- Lin, H.; Zhang, Z.; Iomini, C.; Dutcher, S.K. Identifying RNA splicing factors using IFT genes in Chlamydomonas reinhardtii. Open Biol. 2018, 8, 170211. [Google Scholar] [CrossRef] [Green Version]
- Bessonov, S.; Anokhina, M.; Krasauskas, A.; Golas, M.M.; Sander, B.; Will, C.L.; Urlaub, H.; Stark, H.; Lührmann, R. Characterization of purified human Bact spliceosomal complexes reveals compositional and morphological changes during spliceosome activation and first step catalysis. RNA 2010, 16, 2384–2403. [Google Scholar] [CrossRef] [Green Version]
- Bessonov, S.; Anokhina, M.; Will, C.L.; Urlaub, H.; Lührmann, R. Isolation of an active step I spliceosome and composition of its RNP core. Nature 2008, 452, 846–850. [Google Scholar] [CrossRef] [Green Version]
- Ilagan, J.O.; Chalkley, R.J.; Burlingame, A.; Jurica, M.S. Rearrangements within human spliceosomes captured after exon ligation. RNA 2013, 19, 400–412. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, C.; Grønborg, M.; Deckert, J.; Bessonov, S.; Conrad, T.; Lührmann, R.; Urlaub, H. Mass spectrometry–based relative quantification of proteins in precatalytic and catalytically active spliceosomes by metabolic labeling (SILAC), chemical labeling (iTRAQ), and label-free spectral count. RNA 2014, 20, 406–420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hegele, A.; Kamburov, A.; Grossmann, A.; Sourlis, C.; Wowro, S.; Weimann, M.; Will, C.L.; Pena, V.; Lührmann, R.; Stelzl, U. Dynamic Protein-Protein Interaction Wiring of the Human Spliceosome. Mol. Cell 2012, 45, 567–580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- von Elsner, L.; Chai, G.; Schneeberger, P.E.; Harms, F.L.; Casar, C.; Qi, M.; Alawi, M.; Abdel-Salam, G.M.H.; Zaki, M.S.; Arndt, F.; et al. Biallelic FRA10AC1 variants cause a neurodevelopmental disorder with growth retardation. Brain 2021, 145, 1551–1563. [Google Scholar] [CrossRef] [PubMed]
- Banka, S.; Shalev, S.; Park, S.-M.; Wood, K.A.; Thomas, H.B.; Wright, H.L.; Alyahya, M.; Bankier, S.; Alimi, O.; Chervinsky, E.; et al. Bi-allelic FRA10AC1 variants in a multisystem human syndrome. Brain 2022, 145, e86–e89. [Google Scholar] [CrossRef]
- Alsaleh, N.; Alhashem, A.; Tabarki, B.; Mohamed, S.; Alharby, E.; Alkuraya, F.S.; Almontashiri, N.A. A Biallelic Variant in FRA10AC1 Is Associated With Neurodevelopmental Disorder and Growth Retardation. Neurol. Genet. 2022, 8, e200010. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.S.; Parrado, A.R.; Samtani, M.N.; Narayan, V.A.; Alzheimer’s Disease Neuroimaging Initiative. Variations in the FRA10AC1 Fragile Site and 15q21 Are Associated with Cerebrospinal Fluid Aβ1-42 Level. PLoS ONE 2015, 10, e0134000. [Google Scholar] [CrossRef]
- Dimitrakopoulos, G.N.; Klapa, M.I.; Moschonas, N.K. PICKLE 3.0: Enriching the human meta-database with the mouse protein interactome extended via mouse–human orthology. Bioinformatics 2020, 37, 145–146. [Google Scholar] [CrossRef]
- Dimitrakopoulos, G.N.; Klapa, M.I.; Moschonas, N.K. How Far Are We from the Completion of the Human Protein Interactome Reconstruction? Biomolecules 2022, 12, 140. [Google Scholar] [CrossRef]
- Klapa, M.I.; Tsafou, K.; Theodoridis, E.; Tsakalidis, A.; Moschonas, N.K. Reconstruction of the experimentally supported human protein interactome: What can we learn? BMC Syst. Biol. 2013, 7, 96. [Google Scholar] [CrossRef] [Green Version]
- Gioutlakis, A.; Klapa, M.I.; Moschonas, N.K. PICKLE 2.0: A human protein-protein interaction meta-database employing data integration via genetic information ontology. PLoS ONE 2017, 12, e0186039. [Google Scholar] [CrossRef] [Green Version]
- The UniProt Consortium; Bateman, A.; Martin, M.-J.; Orchard, S.; Magrane, M.; Ahmad, S.; Alpi, E.; Bowler-Barnett, E.H.; Britto, R.; Bye-A-Jee, H.; et al. UniProt: The Universal Protein Knowledgebase in 2023. Nucleic Acids Res. 2022, 51, D523–D531. [Google Scholar] [CrossRef]
- Aguet, F.; Barbeira, A.N.; Bonazzola, R.; Jo, B.; Kasela, S.; Liang, Y.; Parsana, P.; Aguet, F.; Battle, A.; Brown, A.; et al. The GTEx Consortium The GTEx Consortium atlas of genetic regulatory effects across human tissues. Science 2020, 369, 1318–1330. [Google Scholar] [CrossRef]
- Graham, F.L.; Van Der Eb, A.J. A new technique for the assay of infectivity of human adenovirus 5 DNA. Virology 1973, 52, 456–467. [Google Scholar] [CrossRef]
- Chiotaki, R.; Petrou, P.; Giakoumaki, E.; Pavlakis, E.; Sitaru, C.; Chalepakis, G. Spatiotemporal distribution of Fras1/Frem proteins during mouse embryonic development. Gene Expr. Patterns 2007, 7, 381–388. [Google Scholar] [CrossRef]
- Koutelou, E.; Sato, S.; Tomomori-Sato, C.; Florens, L.; Swanson, S.K.; Washburn, M.P.; Kokkinaki, M.; Conaway, R.C.; Conaway, J.W.; Moschonas, N.K. Neuralized-like 1 (Neurl1) targeted to the plasma membrane by n-myristoylation regulates the Notch ligand Jagged1. J. Biol. Chem. 2008, 283, 3846–3853. [Google Scholar] [CrossRef] [Green Version]
- Agafonov, D.E.; Deckert, J.; Wolf, E.; Odenwälder, P.; Bessonov, S.; Will, C.L.; Urlaub, H.; Lührmann, R. Semiquantitative Proteomic analysis of the human spliceosome via a novel two-dimensional gel electrophoresis method. Mol. Cell. Biol. 2011, 31, 2667–2682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayes, S.; Malacrida, B.; Kiely, M.; Kiely, P.A. Studying protein–protein interactions: Progress, pitfalls and solutions. Biochem. Soc. Trans. 2016, 44, 994–1004. [Google Scholar] [CrossRef] [PubMed]
- Chan, J.S.K.; Teo, Z.Q.; Sng, M.K.; Tan, N.S. Probing for protein-protein interactions during cell migration: Limitations and challenges. Histol. Histopathol. 2014, 29, 965–976. [Google Scholar] [CrossRef] [PubMed]
- Oliver, S. Guilt-by-association goes global. Nature 2000, 403, 601–602. [Google Scholar] [CrossRef]
- Ideker, T.; Sharan, R. Protein networks in disease. Genome Res. 2008, 18, 644–652. [Google Scholar] [CrossRef] [Green Version]
- Barabási, A.-L.; Gulbahce, N.; Loscalzo, J. Network medicine: A network-based approach to human disease. Nat. Rev. Genet. 2010, 12, 56–68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Griffin, C.; Saint-Jeannet, J. Spliceosomopathies: Diseases and mechanisms. Dev. Dyn. 2020, 249, 1038–1046. [Google Scholar] [CrossRef]
- Noma, K.; Goncharov, A.; Jin, Y. Systematic Analyses of rpm-1 Suppressors Reveal Roles for ESS-2 in mRNA Splicing in Caenorhabditis elegans. Genetics 2014, 198, 1101–1115. [Google Scholar] [CrossRef] [Green Version]
- Kanno, T.; Venhuizen, P.; Wu, M.-T.; Chiou, P.; Chang, C.-L.; Kalyna, M.; Matzke, A.J.M.; Matzke, M. A Collection of Pre-mRNA Splicing Mutants in Arabidopsis thaliana. G3 Genes Genomes Genet. 2020, 10, 1983–1996. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, D.; McIntosh, C.S.; Mastaglia, F.L.; Wilton, S.D.; Aung-Htut, M.T. Neurodegenerative diseases: A hotbed for splicing defects and the potential therapies. Transl. Neurodegener. 2021, 10, 16. [Google Scholar] [CrossRef]
- Carter, H.; Hofree, M.; Ideker, T. Genotype to phenotype via network analysis. Curr. Opin. Genet. Dev. 2013, 23, 611–621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bertrand, R.E.; Wang, J.; Li, Y.; Cheng, X.; Wang, K.; Stoilov, P.; Chen, R. Cwc27, associated with retinal degeneration, functions as a splicing factor in vivo. Hum. Mol. Genet. 2021, 31, 1278–1292. [Google Scholar] [CrossRef]
- Wu, S.; Romfo, C.M.; Nilsen, T.W.; Green, M.R. Functional recognition of the 3′ splice site AG by the splicing factor U2AF35. Nature 1999, 402, 832–835. [Google Scholar] [CrossRef]
- Lardelli, R.M.; Thompson, J.X.; Yates, J.R.; Stevens, S.W. Release of SF3 from the intron branchpoint activates the first step of pre-mRNA splicing. RNA 2010, 16, 516–528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maita, H.; Kitaura, H.; Ariga, H.; Iguchi-Ariga, S.M. Association of PAP-1 and Prp3p, the products of causative genes of dominant retinitis pigmentosa, in the tri-snRNP complex. Exp. Cell Res. 2005, 302, 61–68. [Google Scholar] [CrossRef]
- Andersen, D.S.; Tapon, N. Drosophila MFAP1 is required for pre-mRNA processing and G2/M progression. J. Biol. Chem. 2008, 283, 31256–31267. [Google Scholar] [CrossRef] [Green Version]
- Ma, L.; Gao, X.; Luo, J.; Huang, L.; Teng, Y.; Horvitz, H.R. The caenorhabditis elegans gene mfap-1 encodes a nuclear protein that affects alternative splicing. PLoS Genet. 2012, 8, e1002827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abovich, N.; Rosbash, M. Cross-intron bridging interactions in the yeast commitment complex are conserved in mammals. Cell 1997, 89, 403–412. [Google Scholar] [CrossRef] [Green Version]
- Spartz, A.K.; Herman, R.K.; Shaw, J.E. SMU-2 and SMU-1, Caenorhabditis elegans homologs of mammalian spliceosome-associated proteins RED and fSAP57, work together to affect splice site choice. Mol. Cell. Biol. 2004, 24, 6811–6823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bressan, G.C.; Quaresma, A.J.C.; Moraes, E.C.; Manfiolli, A.O.; Passos, D.O.; Gomes, M.D.; Kobarg, J. Functional association of human Ki-1/57 with pre-mRNA splicing events. FEBS J. 2009, 276, 3770–3783. [Google Scholar] [CrossRef]
- Sasaki-Osugi, K.; Imoto, C.; Takahara, T.; Shibata, H.; Maki, M. Nuclear ALG-2 protein interacts with Ca2+ homeostasis endoplasmic reticulum protein (CHERP) Ca2+-dependently and participates in regulation of alternative splicing of inositol trisphosphate receptor type 1 (IP3R1) pre-mRNA. J. Biol. Chem. 2013, 288, 33361–33375. [Google Scholar] [CrossRef] [Green Version]
- Chen, D.; Li, Z.; Yang, Q.; Zhang, J.; Zhai, Z.; Shu, H.-B. Identification of a nuclear protein that promotes NF-κB activation. Biochem. Biophys. Res. Commun. 2003, 310, 720–724. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Li, Y.-M.; Li, J.-F.; Liu, L.-D.; Liao, Y.; Na, R.-X.; Wang, J.-J.; Wang, L.-C.; Li, Q.-H. Transcriptional regulation by HSV-1 induced HTRP via acetylation system. Virol. Sin. 2010, 25, 417–424. [Google Scholar] [CrossRef]
- Kaitsuka, T.; Tomizawa, K.; Matsushita, M. Transformation of eEF1Bδ into heat-shock response transcription factor by alternative splicing. EMBO Rep. 2011, 12, 673–681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohira, T.; Kojima, H.; Kuroda, Y.; Aoki, S.; Inaoka, D.; Osaki, M.; Wanibuchi, H.; Okada, F.; Oshimura, M.; Kugoh, H. PITX1 protein interacts with ZCCHC10 to regulate hTERT mRNA transcription. PLoS ONE 2019, 14, e0217605. [Google Scholar] [CrossRef] [Green Version]
- Cheng, H.; Dufu, K.; Lee, C.-S.; Hsu, J.L.; Dias, A.; Reed, R. Human mRNA export machinery recruited to the 5′ end of mRNA. Cell 2006, 127, 1389–1400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Busetto, V.; Barbosa, I.; Basquin, J.; Marquenet, E.; Hocq, R.; Hennion, M.; Paternina, J.A.; Namane, A.; Conti, E.; Bensaude, O.; et al. Structural and functional insights into CWC27/CWC22 heterodimer linking the exon junction complex to spliceosomes. Nucleic Acids Res. 2020, 48, 5670–5683. [Google Scholar] [CrossRef] [PubMed]
- Singh, G.; Kucukural, A.; Cenik, C.; Leszyk, J.D.; Shaffer, S.A.; Weng, Z.; Moore, M.J. The Cellular EJC Interactome reveals higher-order mRNP structure and an EJC-SR protein nexus. Cell 2012, 151, 750–764. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Disease | Reference | Inheritance | Association | ||
|---|---|---|---|---|---|
| Spliceosomal FRA10AC1 interactors | PRPF40A | Memory dysfunction in frontotemporal lobe dementia, Male puberty timing (late vs. average onset voice breaking) | GWAS catalog | # | |
| SF3B2 | Craniofacial microsomia | MIM No. 164210 | AD | ||
| PRPF3 | Retinitis pigmentosa 18 | MIM No. 601414 | AD | ||
| THOC1 | Late-onset, progressive, non-syndromic hearing loss | PMID: 32776944 | AD | ||
| CWC27 | Retinitis pigmentosa & neurological defects | MIM No. 250410 | AR | ||
| ESS2 | Schizophrenia | PMID: 16432632 | # | ||
| NKAP | Hackmann-Di Donato-type X-linked syndromic intellectual developmental disorder | MIM No. 301039 | XLR | ||
| HABP4 | Cognitive ability | GWAS catalog | # | ||
| MOB2 | Neuronal mispositioning and periventricular nodular heterotopia (probable cause of) | PMID: 29593499 | AR? | ||
| Non-spliceosomal FRA10AC1 interactors | EEF1D | Neurodevelopmental disorder (specific to the long protein isoform) | PMID: 36344539 | # | |
| ADPRHL2 | Neurodegeneration, childhood-onset, stress-induced, with variable ataxia and seizures | MIM No. 618170 | AR | ||
| CCDC155 | Idiopathic non-obstructive azoospermia | PMID: 29790874 | # | ||
| VPS29 | Schizophrenia, neurodegenerative diseases (Alzheimer’s, Parkinson’s, frontotemporal lobar degeneration) | PMID: 29755290, 32398722 | # | ||
| PLPPR4 | Intellectual disability | PMID: 32388443 | # | ||
| GATD3A | Crohn’s disease; Parkinson’s disease | PMID: 18587394; 35307029 | # | ||
| FRA10AC1 second neighbors (selected) | FMR1 | Fragile X syndrome, Fragile X tremor/ataxia syndrome, Premature ovarian failure 1 | MIM No. 300624; 300623; 311360 | XLD, XL | |
| FAM161A | Retinitis pigmentosa 28 | MIM No. 613596 | AR | ||
| APP | Alzheimer disease, familial; cerebral amyloid angiopathy | MIM No. 104300; 605714 | AD | ||
| SQSTM | Amyotrophic Lateral Sclerosis 3, Paget Disease of Bone 3; Neurodegeneration with ataxia, dystonia, and gaze palsy, childhood-onset | MIM No. 616437; 167250; 617145 | AD, AR | ||
| PIK3R1 | SHORT Syndrome, Immunodeficiency 36 | MIM No. 269880; 616005 | AD |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sarafidou, T.; Galliopoulou, E.; Apostolopoulou, D.; Fragkiadakis, G.A.; Moschonas, N.K. Reconstruction of a Comprehensive Interactome and Experimental Data Analysis of FRA10AC1 May Provide Insights into Its Biological Role in Health and Disease. Genes 2023, 14, 568. https://doi.org/10.3390/genes14030568
Sarafidou T, Galliopoulou E, Apostolopoulou D, Fragkiadakis GA, Moschonas NK. Reconstruction of a Comprehensive Interactome and Experimental Data Analysis of FRA10AC1 May Provide Insights into Its Biological Role in Health and Disease. Genes. 2023; 14(3):568. https://doi.org/10.3390/genes14030568
Chicago/Turabian StyleSarafidou, Theologia, Eleni Galliopoulou, Despina Apostolopoulou, Georgios A. Fragkiadakis, and Nicholas K. Moschonas. 2023. "Reconstruction of a Comprehensive Interactome and Experimental Data Analysis of FRA10AC1 May Provide Insights into Its Biological Role in Health and Disease" Genes 14, no. 3: 568. https://doi.org/10.3390/genes14030568