
Genetic Analysis of HIBM Myopathy-Specific GNE V727M Hotspot Mutation Identifies a Novel COL6A3 Allied Gene Signature That Is Also Deregulated in Multiple Neuromuscular Diseases and Myopathies


 , and
, and 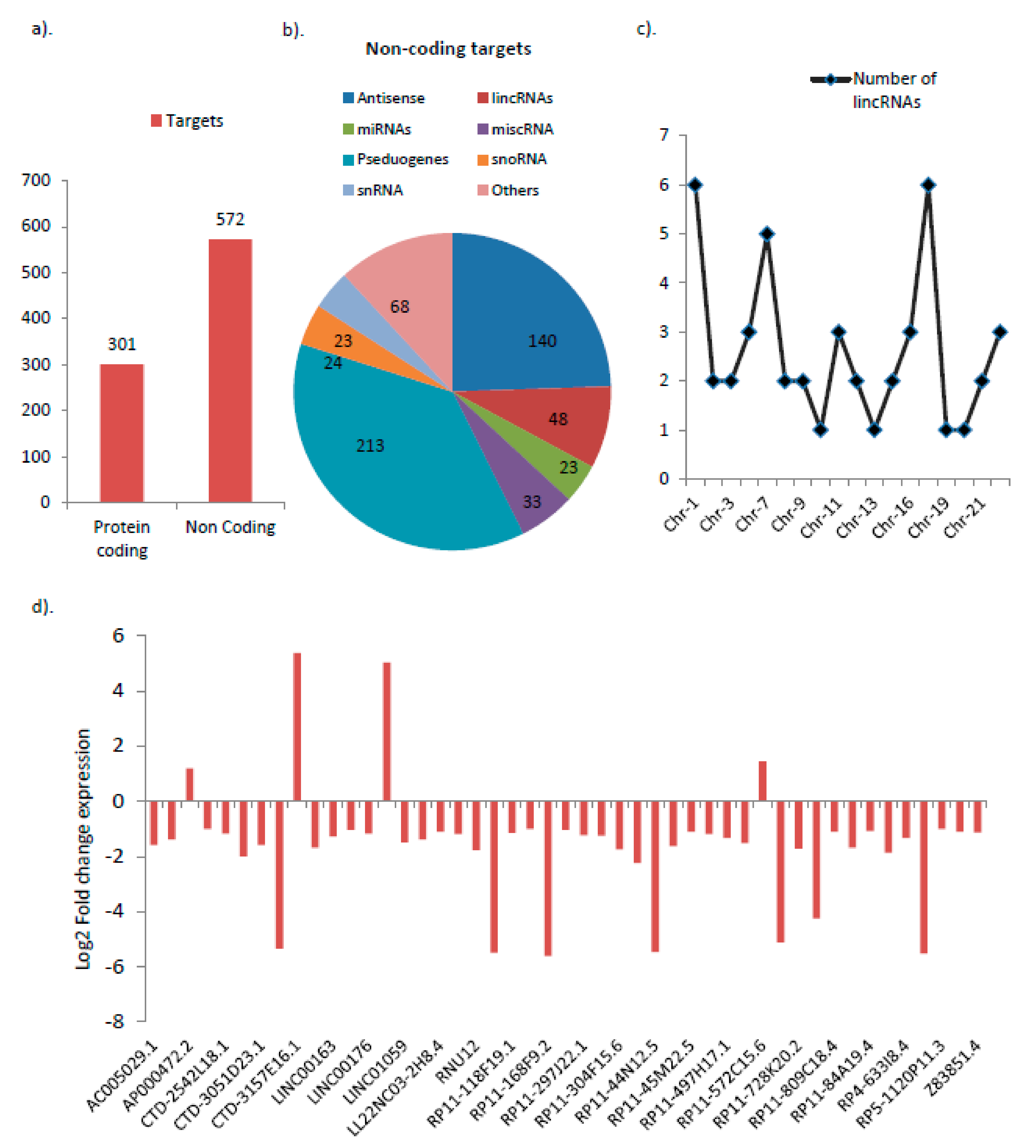{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Background
2. Materials and Methods
2.1. Cell Culture
2.2. GNE Gene Cloning
2.3. Site-Directed Mutagenesis (SDM)
2.4. Plasmid Extraction and Purification
2.5. Transfection
2.6. Total RNA Isolation
2.7. Reverse Transcription and Real-Time PCR Studies
2.8. RNAseq Study
2.9. Data Processing and Quality Control
2.10. Differential Expression Analysis
2.11. LincRNA Target Prediction
2.12. Gene Set Enrichment Analysis (GSEA)
2.13. String Analysis
2.14. GEO Data Analysis
2.15. Statistical Analysis
3. Results
3.1. Differential Expression of Various Noncoding RNAs Are Found to Be Associated with GNE V727M Mutation
3.2. lincRNAs Are the Most Highly Deregulated Group by the GNE V727M Mutation
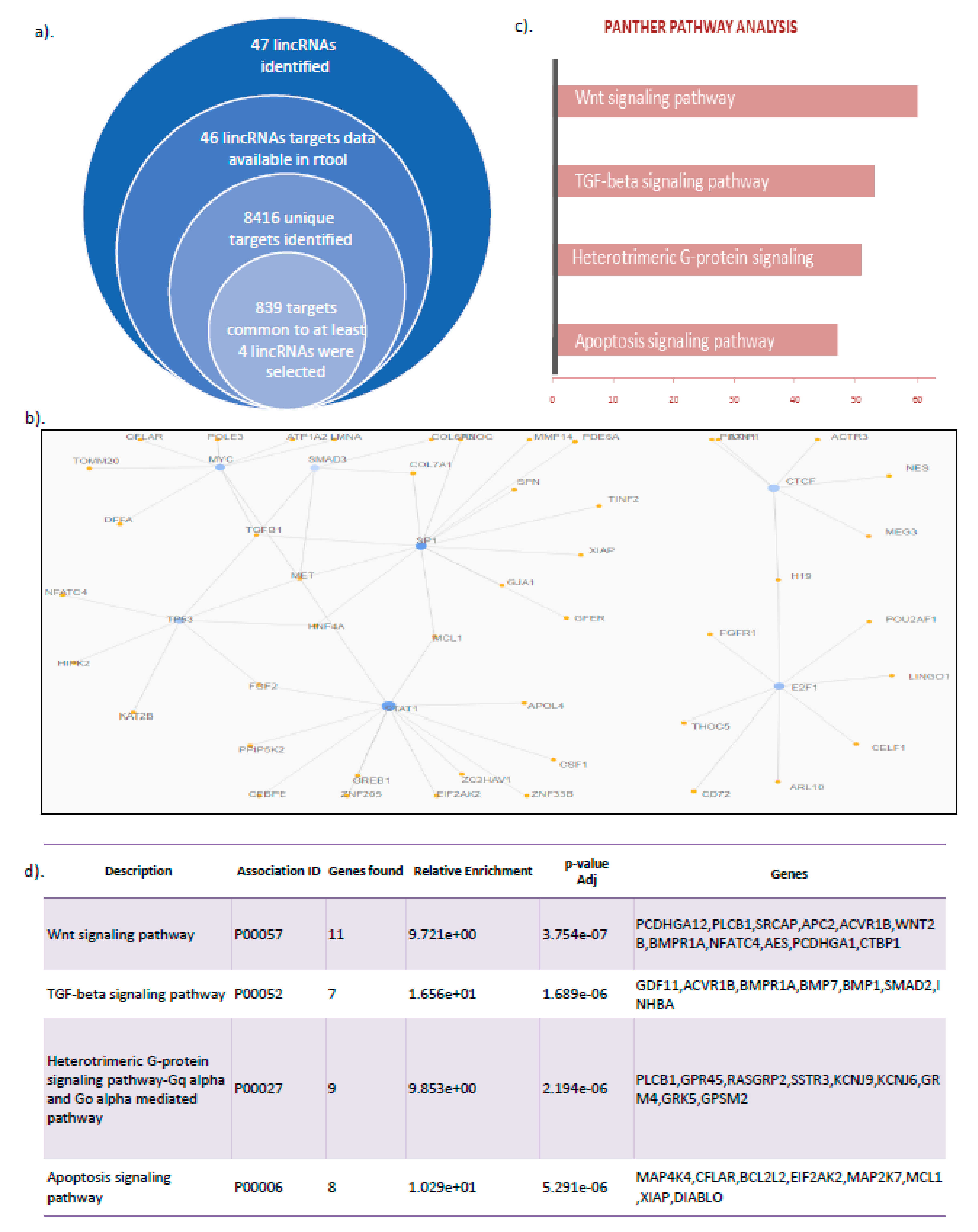
3.3. LincRNA Target Analysis Showed Their Role in the Regulation of Wnt Signaling, TGF β Pathway, and Apoptotic Signaling
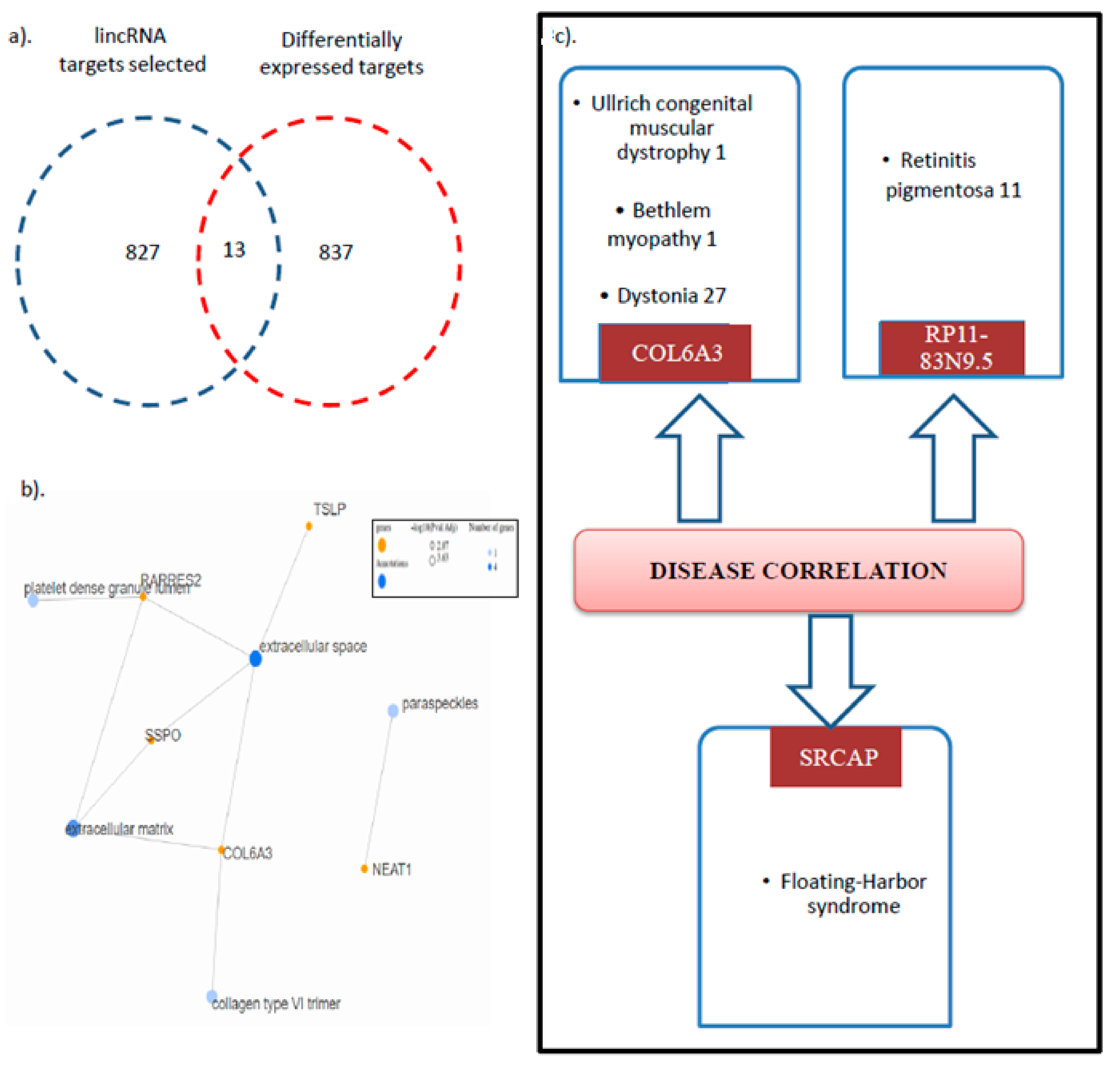
3.4. Collagen VI Family Member COL6a3 Is a Key Target Deregulated via GNE V727M Mutation
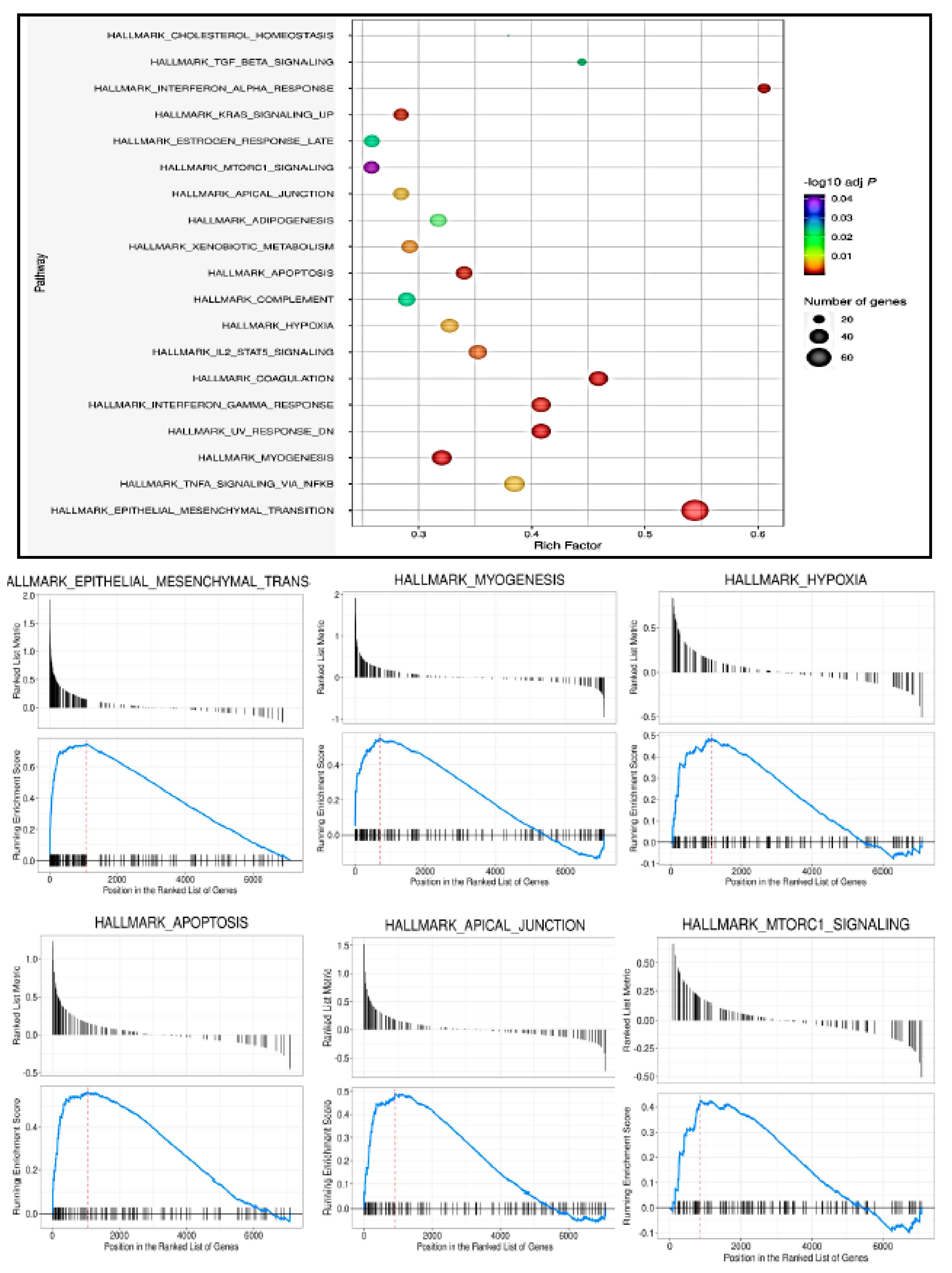
3.5. HIBM Patients with Varying COL6A3 Expression Show Alteration of Myogenic-Associated Pathways
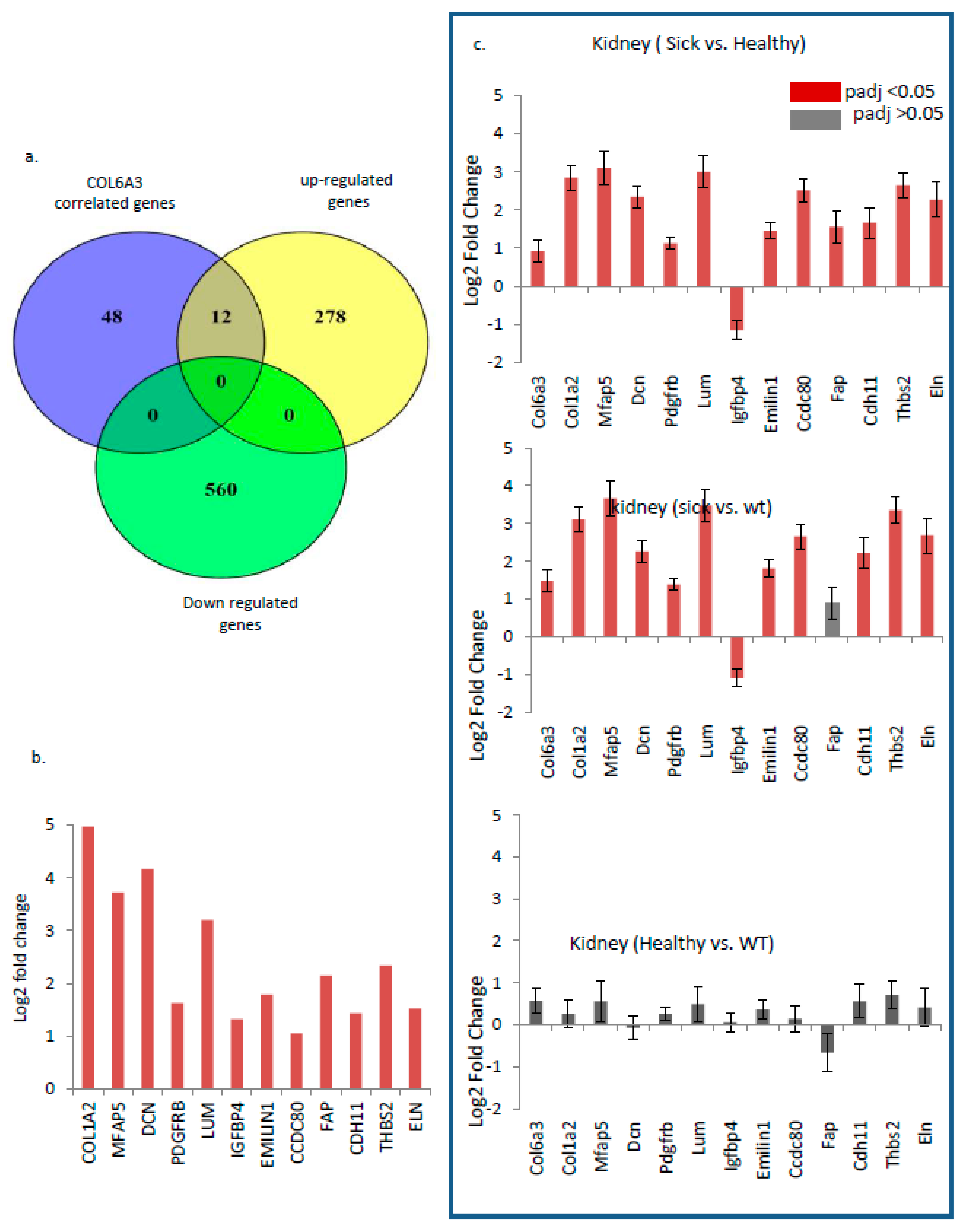
3.6. COL6a3 Associated 13 Gene Signature Is Highly Deregulated by GNE V727M Mutation
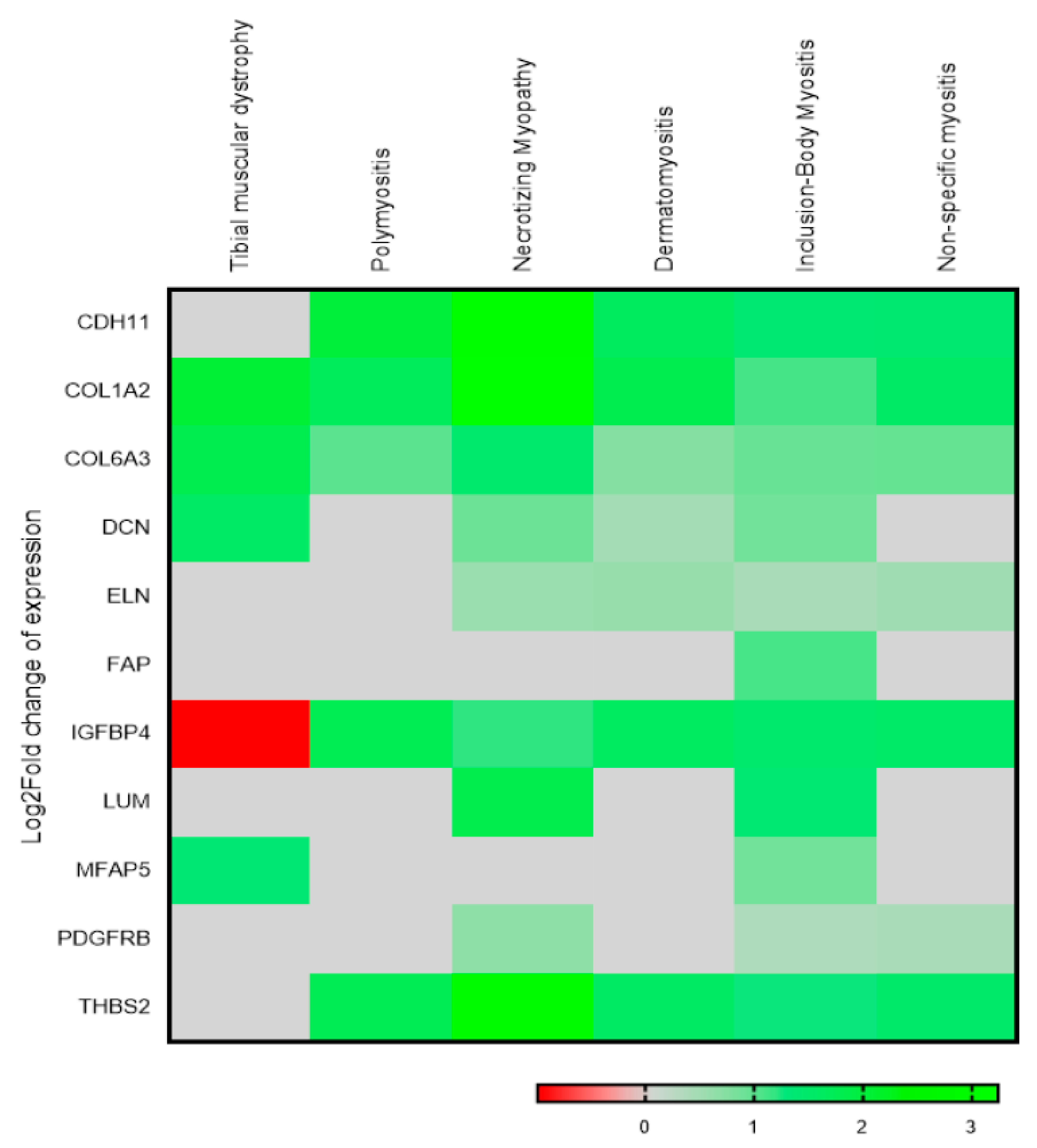
3.7. Multiple Musculoskeletal Disorders also Show a Deregulated Trend for the Identified 13 Gene Signature
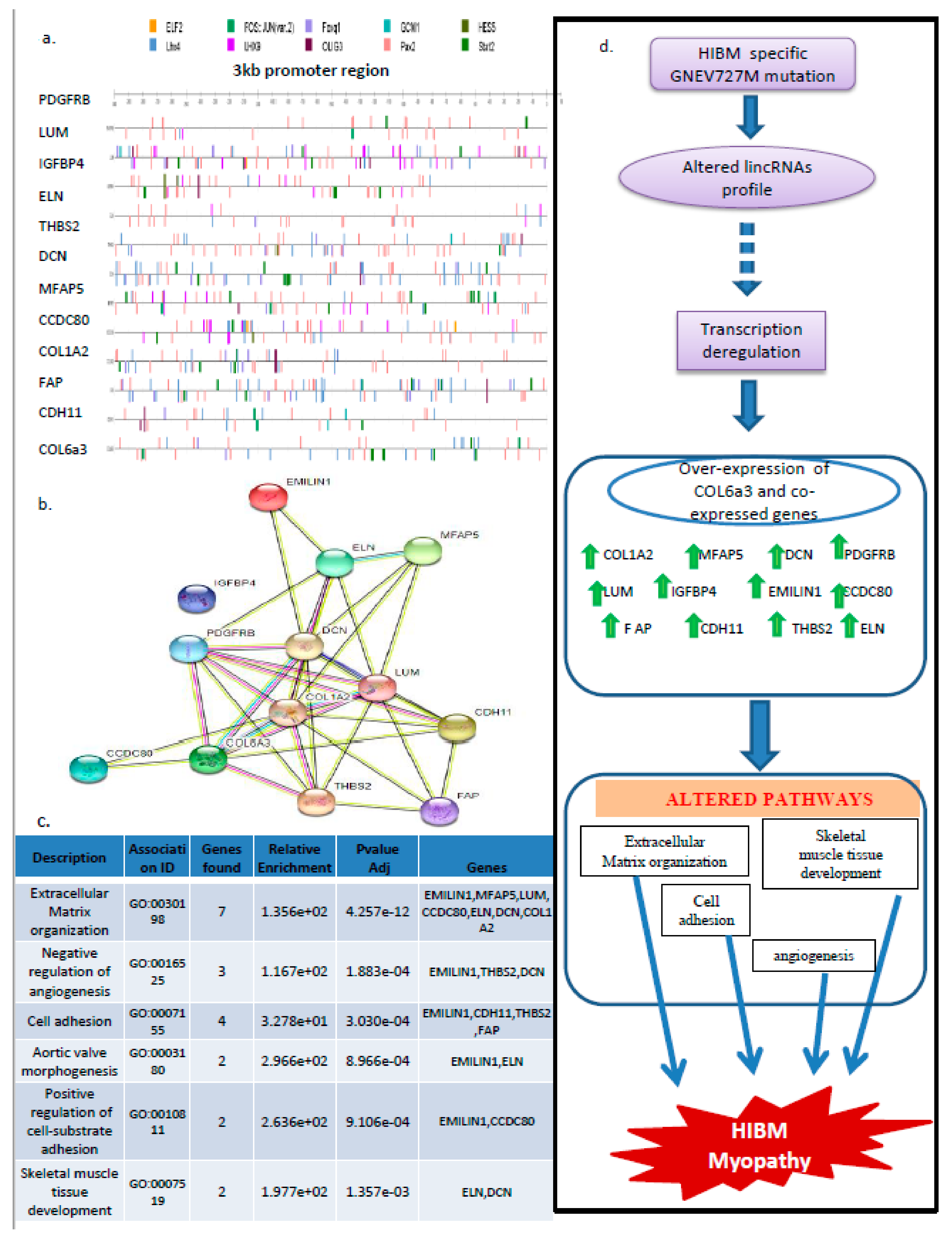
3.8. PAX2, STAT2, and LHX4 Seem to Be the Key Players in the Regulation of the 13 Gene Signatures
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Argov, Z.; Yarom, R. “Rimmed Vacuole Myopathy” Sparing the Quadriceps. A Unique Disorder in Iranian Jews. J. Neurol. Sci. 1984, 64, 33–43. [Google Scholar] [CrossRef] [PubMed]
- Nishino, I.; Carrillo-Carrasco, N.; Argov, Z. GNE Myopathy: Current Update and Future Therapy. J. Neurol. Neurosurg. Psychiatry 2015, 86, 385–392. [Google Scholar] [CrossRef] [PubMed]
- Argov, Z.; Mitrani Rosenbaum, S. GNE Myopathy: Two Clusters with History and Several Founder Mutations. J. Neuromuscul. Dis. 2015, 2, S73–S76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Celeste, F.V.; Vilboux, T.; Ciccone, C.; de Dios, J.K.; Malicdan, M.C.V.; Leoyklang, P.; Mckew, J.C.; Gahl, W.A.; Carrillo-Carrasco, N.; Huizing, M. Mutation Update for GNE Gene Variants Associated with GNE Myopathy. Hum. Mutat. 2014, 35, 915–926. [Google Scholar] [CrossRef] [Green Version]
- Attri, S.; Gahlawat, S. Challenges and Advances in Molecular Diagnosis of Myopathies and Dystrophies in Perspective of Their Use in Developing Countries: Past, Present, and Future. Neurol. India 2021, 69, 797. [Google Scholar] [CrossRef]
- Huizing, M.; Krasnewich, D.M. Hereditary Inclusion Body Myopathy: A Decade of Progress. Biochim. Biophys. Acta-Mol. Basis Dis. 2009, 1792, 881–887. [Google Scholar] [CrossRef] [Green Version]
- Eisenberg, I.; Avidan, N.; Potikha, T.; Hochner, H.; Chen, M.; Olender, T.; Barash, M.; Shemesh, M.; Sadeh, M.; Grabov-Nardini, G.; et al. The UDP-N-Acetylglucosamine 2-Epimerase/N-Acetylmannosamine Kinase Gene Is Mutated in Recessive Hereditary Inclusion Body Myopathy. Nat. Genet. 2001, 29, 83–87. [Google Scholar] [CrossRef]
- Varki, A.; Cummings, R.; Esko, J. Chapter 40, Natural and Synthetic Inhibitors of Glycosylation. In Essentials of Glycobiology; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 1999. [Google Scholar]
- Krause, S.; Hinderlich, S.; Amsili, S.; Horstkorte, R.; Wiendl, H.; Argov, Z.; Mitrani-Rosenbaum, S.; Lochmüller, H. Localization of UDP-GlcNAc 2-Epimerase/ManAc Kinase (GNE) in the Golgi Complex and the Nucleus of Mammalian Cells. Exp. Cell Res. 2005, 304, 365–379. [Google Scholar] [CrossRef]
- Effertz, K.; Hinderlich, S.; Reutter, W. Selective Loss of Either the Epimerase or Kinase Activity of UDP-N- Acetylglucosamine 2-Epimerase/N-Acetylmannosamine Kinase Due to Site-Directed Mutagenesis Based on Sequence Alignments. J. Biol. Chem. 1999, 274, 28771–28778. [Google Scholar] [CrossRef] [Green Version]
- Attri, S.; Sharma, V.; Kumar, A.; Verma, C.; Gahlawat, S.K. Dissecting Role of Founder Mutation p.V727M in GNE in Indian HIBM Cohort. Open Med. 2021, 16, 1733–1744. [Google Scholar] [CrossRef]
- Smith, M.A.; Mattick, J.S. Structural and Functional Annotation of Long Noncoding RNAs. In Methods in Molecular Biology; Humana Press: Totowa, NJ, USA, 2017. [Google Scholar]
- Fitzgerald, K.A.; Caffrey, D.R. Long Noncoding RNAs in Innate and Adaptive Immunity. Curr. Opin. Immunol. 2014, 26, 140–146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.L. Linking Long Noncoding RNA Localization and Function. Trends Biochem. Sci. 2016, 41, 761–772. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Liang, H.; Yang, H.; Zhou, K.; Xu, L.; Liu, J.; Lai, B.; Song, L.; Luo, H.; Peng, J.; et al. LincRNa-P21: Function and Mechanism in Cancer. Med. Oncol. 2017, 34, 98. [Google Scholar] [CrossRef]
- Spurlock, C.F.; Tossberg, J.T.; Matlock, B.K.; Olsen, N.J.; Aune, T.M. Methotrexate Inhibits NF-ΚB Activity via Long Intergenic (Noncoding) RNA-P21 Induction. Arthritis Rheumatol. 2014, 66, 2947–2957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Y.; Zhang, F.; Ma, J.; Zhang, X.; Wu, L.; Qu, B.; Xia, S.; Chen, S.; Tang, Y.; Shen, N. Association of Large Intergenic Noncoding RNA Expression with Disease Activity and Organ Damage in Systemic Lupus Erythematosus. Arthritis Res. Ther. 2015, 17, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Gupta, R.A.; Shah, N.; Wang, K.C.; Kim, J.; Horlings, H.M.; Wong, D.J.; Tsai, M.C.; Hung, T.; Argani, P.; Rinn, J.L.; et al. Long Noncoding RNA HOTAIR Reprograms Chromatin State to Promote Cancer Metastasis. Nature 2010, 464, 1071–1076. [Google Scholar] [CrossRef] [Green Version]
- Penny, G.D.; Kay, G.F.; Sheardown, S.A.; Rastan, S.; Brockdorff, N. Requirement for Xist in X Chromosome Inactivation. Nature 1996, 379, 131–137. [Google Scholar] [CrossRef]
- Yoon, J.H.; Abdelmohsen, K.; Srikantan, S.; Yang, X.; Martindale, J.L.; De, S.; Huarte, M.; Zhan, M.; Becker, K.G.; Gorospe, M. LincRNA-P21 Suppresses Target MRNA Translation. Mol. Cell 2012, 47, 648–655. [Google Scholar] [CrossRef] [Green Version]
- Chaleshi, V.; Irani, S.; Alebouyeh, M.; Mirfakhraie, R.; Aghdaei, H.A. Association of LncRNA-P53 Regulatory Network (LincRNA-P21, LincRNA-ROR and MALAT1) and P53 with the Clinicopathological Features of Colorectal Primary Lesions and Tumors. Oncol. Lett. 2020, 19, 3937–3949. [Google Scholar] [CrossRef] [Green Version]
- Ding, J.; Zhao, J.; Huan, L.; Liu, Y.; Qiao, Y.; Wang, Z.; Chen, Z.; Huang, S.; Zhao, Y.; He, X. Inflammation-Induced Long Intergenic Noncoding RNA (LINC00665) Increases Malignancy Through Activating the Double-Stranded RNA–Activated Protein Kinase/Nuclear Factor Kappa B Pathway in Hepatocellular Carcinoma. Hepatology 2020, 72, 1666–1681. [Google Scholar] [CrossRef]
- Shaker, O.G.; Golam, R.M.; Ayoub, S.; Daker, L.I.; Elguaad, M.K.A.; Said, E.S.; Khalil, M.A.F. Correlation between LincR-Gng2-5′and LincR-Epas1-3′as with the Severity of Multiple Sclerosis in Egyptian Patients. Int. J. Neurosci. 2020, 130, 515–521. [Google Scholar] [CrossRef]
- Wu, X.; Cao, Z.; Chen, H.; Ou, Q.; Huang, X.; Wang, Y. Downregulation of Linc-RNA Activator of Myogenesis LncRNA Participates in FGF2-Mediated Proliferation of Human Periodontal Ligament Stem Cells. J. Periodontol. 2020, 91, 422–427. [Google Scholar] [CrossRef]
- Khademian, H.; Mehravar, E.; Urtizberea, J.A.; Sagoo, S.; Sandoval, L.; Carbajo, R.; Darvish, B.; Valles-Ayoub, Y.; Darvish, D. Prevalence of GNE p.M712T and Hereditary Inclusion Body Myopathy (HIBM) in Sangesar Population of Northern Iran. Clin. Genet. 2013, 84, 589–592. [Google Scholar] [CrossRef]
- Awasthi, K.; Arya, R.; Bhattacharya, A.; Bhattacharya, S. The Inherited Neuromuscular Disorder GNE Myopathy: Research to Patient Care. Neurol. India 2019, 67, 1213–1219. [Google Scholar]
- Askanas, V.; Engel, W.K. Inclusion-Body Myositis and Myopathies: Different Etiologies, Possibly Similar Pathogenic Mechanisms. Curr. Opin. Neurol. 2002, 15, 525–531. [Google Scholar] [CrossRef]
- Benyamini, H.; Kling, Y.; Yakovlev, L.; Becker Cohen, M.; Nevo, Y.; Elgavish, S.; Harazi, A.; Argov, Z.; Sela, I.; Mitrani-Rosenbaum, S. Upregulation of Hallmark Muscle Genes Protects GneM743T/M743T Mutated Knock-In Mice from Kidney and Muscle Phenotype. J. Neuromuscul. Dis. 2020, 7, 119–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, F.; Liang, Z.; Xu, J.; Li, W.; Zhao, D.; Zhao, Y.; Yan, C. Activation of the Wnt/β-Catenin Signaling Pathway in Polymyositis, Dermatomyositis and Duchenne Muscular Dystrophy. J. Clin. Neurol. 2016, 12, 351–360. [Google Scholar] [CrossRef] [PubMed]
- Okada, K.; Naito, A.T.; Higo, T.; Nakagawa, A.; Shibamoto, M.; Sakai, T.; Hashimoto, A.; Kuramoto, Y.; Sumida, T.; Nomura, S.; et al. Wnt/β-Catenin Signaling Contributes to Skeletal Myopathy in Heart Failure via Direct Interaction with Forkhead Box O. Circ. Heart Fail. 2015, 8, 799–808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burks, T.N.; Cohn, R.D. Role of TGF-β Signaling in Inherited and Acquired Myopathies. Skelet. Muscle 2011, 1, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- MacDonald, E.M.; Cohn, R.D. TGFβ Signaling: Its Role in Fibrosis Formation and Myopathies. Curr. Opin. Rheumatol. 2012, 24, 628–634. [Google Scholar] [CrossRef] [PubMed]
- Harazi, A.; Chaouat, M.; Shlomai, Z.; Levitzki, R.; Becker-Cohen, M.; Sadeh, M.; Dabby, R.; Ben-Bassat, H.; Mitrani-Rosenbaum, S. Survival-Apoptosis Associated Signaling in GNE Myopathy-Cultured Myoblasts. J. Recept. Signal Transduct. 2015, 35, 249–257. [Google Scholar] [CrossRef] [PubMed]
- Barnes, B.T.; Confides, A.L.; Rich, M.M.; Dupont-Versteegden, E.E. Distinct Muscle Apoptotic Pathways Are Activated in Muscles with Different Fiber Types in a Rat Model of Critical Illness Myopathy. J. Muscle Res. Cell Motil. 2015, 36, 243–253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, M.J.; Lee, J.S.; Lee, M.C. Apoptosis of Skeletal Muscle on Steroid-Induced Myopathy in Rats. J. Korean Med. Sci. 2001, 16, 467–474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.Y.; Kim, W.J.; Kim, H.; Choi, S.A.; Lee, J.S.; Cho, A.; Jang, S.S.; Lim, B.C.; Kim, K.J.; Kim, J.I.; et al. Collagen VI-Related Myopathy: Expanding the Clinical and Genetic Spectrum. Muscle Nerve 2018, 58, 381–388. [Google Scholar] [CrossRef]
- Bönnemann, C.G. The Collagen VI-Related Myopathies: Muscle Meets Its Matrix. Nat. Rev. Neurol. 2011, 7, 379–390. [Google Scholar] [CrossRef] [Green Version]
- Foley, A.R.; Mohassel, P.; Donkervoort, S.; Bolduc, V.; Bönnemann, C.G. Collagen VI-Related Dystrophies. In GeneReviews ®; University of Washington: Seattle, WA, USA, 1993. [Google Scholar]
- Merlini, L.; Bernardi, P. Therapy of Collagen VI-Related Myopathies (Bethlem and Ullrich). Neurotherapeutics 2008, 5, 613–618. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.H.; Shin, H.Y.; Park, H.J.; Kim, S.H.; Kim, S.M.; Choi, Y.C. Clinical, Pathologic, and Genetic Features of Collagen VI-Related Myopathy in Korea. J. Clin. Neurol. 2017, 13, 331–339. [Google Scholar] [CrossRef] [Green Version]
- Chakravorty, S.; Berger, K.; Arafat, D.; Nallamilli, B.R.R.; Subramanian, H.P.; Joseph, S.; Anderson, M.E.; Campbell, K.P.; Glass, J.; Gibson, G.; et al. Clinical Utility of RNA Sequencing to Resolve Unusual GNE Myopathy with a Novel Promoter Deletion. Muscle Nerve 2019, 60, 98–103. [Google Scholar] [CrossRef]
- Zealy, R.W.; Fomin, M.; Davila, S.; Makowsky, D.; Thigpen, H.; McDowell, C.H.; Cummings, J.C.; Lee, E.S.; Kwon, S.H.; Min, K.W.; et al. Long Noncoding RNA Complementarity and Target Transcripts Abundance. Biochim. Biophys. Acta-Gene Regul. Mech. 2018, 1861, 224–234. [Google Scholar] [CrossRef]
- Gong, C.; Maquat, L.E. LncRNAs Transactivate STAU1-Mediated MRNA Decay by Duplexing with 39 UTRs via Alu Eleme. Nature 2011, 470, 284–288. [Google Scholar] [CrossRef] [Green Version]
- Carrieri, C.; Cimatti, L.; Biagioli, M.; Beugnet, A.; Zucchelli, S.; Fedele, S.; Pesce, E.; Ferrer, I.; Collavin, L.; Santoro, C.; et al. Long Non-Coding Antisense RNA Controls Uchl1 Translation through an Embedded SINEB2 Repeat. Nature 2012, 491, 454–457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krämer, O.H.; Heinzel, T. Phosphorylation-Acetylation Switch in the Regulation of STAT1 Signaling. Mol. Cell. Endocrinol. 2010, 315, 40–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beishline, K.; Azizkhan-Clifford, J. Sp1 and the “Hallmarks of Cancer. FEBS J. 2015, 282, 224–258. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Pan, L.; Feng, Y.; Wang, Y.; Han, Q.; Han, L.; Han, S.; Guo, J.; Huang, B.; Lu, J. P300 Plays a Role in P16INK4a Expression and Cell Cycle Arrest. Oncogene 2008, 27, 1894–1904. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Feng, Y.; Pan, L.; Wang, Y.; Xu, X.; Lu, J.; Huang, B. The Proximal GC-Rich Region of P16INK4a Gene Promoter Plays a Role in Its Transcriptional Regulation. Mol. Cell. Biochem. 2007, 301, 259–266. [Google Scholar] [CrossRef]
- Wu, J.; Xue, L.; Weng, M.; Sun, Y.; Zhang, Z.; Wang, W.; Tong, T. Sp1 Is Essential for P16INK4a Expression in Human Diploid Fibroblasts during Senescence. PLoS ONE 2007, 2, e164. [Google Scholar] [CrossRef] [Green Version]
- Xu, W.; Zhu, Q.; Wu, Z.; Guo, H.; Wu, F.; Mashausi, D.S.; Zheng, C.; Li, D. A Novel Evolutionarily Conserved Element Is a General Transcriptional Repressor of P21WAF1/CIP1. Cancer Res. 2012, 72, 6236–6246. [Google Scholar] [CrossRef] [Green Version]
- Chae, J.I.; Cho, J.H.; Lee, K.A.; Choi, N.J.; Seo, K.S.; Kim, S.B.; Lee, S.H.; Shim, J.H. Role of Transcription Factor Sp1 in the Quercetin-Mediated Inhibitory Effect on Human Malignant Pleural Mesothelioma. Int. J. Mol. Med. 2012, 30, 835–841. [Google Scholar] [CrossRef] [Green Version]
- Waby, J.S.; Chirakkal, H.; Yu, C.W.; Griffiths, G.J.; Benson, R.S.P.; Bingle, C.D.; Corfe, B.M. Sp1 Acetylation Is Associated with Loss of DNA Binding at Promoters Associated with Cell Cycle Arrest and Cell Death in a Colon Cell Line. Mol. Cancer 2010, 9, 275. [Google Scholar] [CrossRef] [Green Version]
- Milanini-Mongiat, J.; Pouysségur, J.; Pagès, G. Identification of Two Sp1 Phosphorylation Sites for P42/P44 Mitogen-Activated Protein Kinases: Their Implication in Vascular Endothelial Growth Factor Gene Transcription. J. Biol. Chem. 2002, 277, 20631–20639. [Google Scholar] [CrossRef] [Green Version]
- Dang, C.V.; O’Donnell, K.A.; Zeller, K.I.; Nguyen, T.; Osthus, R.C.; Li, F. The C-Myc Target Gene Network. Semin. Cancer Biol. 2006, 16, 253–264. [Google Scholar] [CrossRef] [PubMed]
- Cisternas, P.; Henriquez, J.P.; Brandan, E.; Inestrosa, N.C. Wnt Signaling in Skeletal Muscle Dynamics: Myogenesis, Neuromuscular Synapse and Fibrosis. Mol. Neurobiol. 2014, 49, 574–589. [Google Scholar] [CrossRef] [PubMed]
- Faenza, I.; Blalock, W.; Bavelloni, A.; Schoser, B.; Fiume, R.; Pacella, S.; Piazzi, M.; D’Angelo, A.; Cocco, L. A Role for PLCβ1 in Myotonic Dystrophies Type 1 and 2. FASEB J. 2012, 26, 3042–3048. [Google Scholar] [CrossRef] [PubMed]
- Elbaz, M.; Yanay, N.; Aga-Mizrachi, S.; Brunschwig, Z.; Kassis, I.; Ettinger, K.; Barak, V.; Nevo, Y. Losartan, a Therapeutic Candidate in Congenital Muscular Dystrophy: Studies in the Dy 2J/Dy 2J Mouse. Ann. Neurol. 2012, 71, 699–708. [Google Scholar] [CrossRef] [PubMed]
- Cohn, R.D.; Van Erp, C.; Habashi, J.P.; Soleimani, A.A.; Klein, E.C.; Lisi, M.T.; Gamradt, M.; Ap Rhys, C.M.; Holm, T.M.; Loeys, B.L.; et al. Angiotensin II Type 1 Receptor Blockade Attenuates TGF-β-Induced Failure of Muscle Regeneration in Multiple Myopathic States. Nat. Med. 2007, 13, 204–210. [Google Scholar] [CrossRef] [Green Version]
- Bernasconi, P.; Di Blasi, C.; Mora, M.; Morandi, L.; Galbiati, S.; Confalonieri, P.; Cornelio, F.; Mantegazza, R. Transforming Growth Factor-Β1 and Fibrosis in Congenital Muscular Dystrophies. Neuromuscul. Disord. 1999, 9, 28–33. [Google Scholar] [CrossRef]
- Choi, E.; Shin, S.; Lee, S.; Lee, S.J.; Park, J. Coexistence of Digenic Mutations in the Collagen VI Genes (COL6A1 and COL6A3) Leads to Bethlem Myopathy. Clin. Chim. Acta 2020, 508, 28–32. [Google Scholar] [CrossRef]
- Marakhonov, A.V.; Tabakov, V.Y.; Zernov, N.V.; Dadali, E.L.; Sharkova, I.V.; Skoblov, M.Y. Two Novel COL6A3 Mutations Disrupt Extracellular Matrix Formation and Lead to Myopathy from Ullrich Congenital Muscular Dystrophy and Bethlem Myopathy Spectrum. Gene 2018, 672, 165–171. [Google Scholar] [CrossRef]
- Lampe, A.K.; Dunn, D.M.; Von Niederhausern, A.C.; Hamil, C.; Aoyagi, A.; Laval, S.H.; Marie, S.K.; Chu, M.L.; Swoboda, K.; Muntoni, F.; et al. Automated Genomic Sequence Analysis of the Three Collagen VI Genes: Applications to Ullrich Congenital Muscular Dystrophy and Bethlem Myopathy. J. Med. Genet. 2005, 42, 108–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marrosu, E.; Ala, P.; Muntoni, F.; Zhou, H. Gapmer Antisense Oligonucleotides Suppress the Mutant Allele of COL6A3 and Restore Functional Protein in Ullrich Muscular Dystrophy. Mol. Ther.-Nucleic Acids 2017, 8, 416–427. [Google Scholar] [CrossRef] [Green Version]
- Mihaylova, V.; Chablais, F.; Bremer, J.; Guggenberger, R.; Rushing, E.J.; Bethge, T.; Spiegel, R.; Jung, H.H. Collagen VI-Related Myopathy Caused by Compound Heterozygous Mutations of COL6A3 in a Consanguineous Kurdish Family. J. Clin. Neuromuscul. Dis. 2021, 22, 173–179. [Google Scholar] [CrossRef] [PubMed]
- Lamandé, S.R.; Bateman, J.F. Collagen VI Disorders: Insights on Form and Function in the Extracellular Matrix and Beyond. Matrix Biol. 2018, 71–72, 348–367. [Google Scholar] [CrossRef] [PubMed]
- Kamma-Lorger, C.S.; Pinali, C.; Martinez, J.C.; Harris, J.; Young, R.D.; Bredrup, C.; Crosas, E.; Malfois, M.; RÃdahl, E.; Meek, K.M.; et al. Role of Decorin Core Protein in Collagen Organisation in Congenital Stromal Corneal Dystrophy (CSCD). PLoS ONE 2016, 11, e0147948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bornstein, P.; Kyriakides, T.R.; Yang, Z.; Armstrong, L.C.; Birk, D.E. Thrombospondin 2 Modulates Collagen Fibrillogenesis and Angiogenesis. J. Investig. Dermatol. Symp. Proc. 2000, 5, 61–66. [Google Scholar] [CrossRef] [Green Version]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Attri, S.; Lone, M.; Katiyar, A.; Sharma, V.; Kumar, V.; Verma, C.; Gahlawat, S.K. Genetic Analysis of HIBM Myopathy-Specific GNE V727M Hotspot Mutation Identifies a Novel COL6A3 Allied Gene Signature That Is Also Deregulated in Multiple Neuromuscular Diseases and Myopathies. Genes 2023, 14, 567. https://doi.org/10.3390/genes14030567
Attri S, Lone M, Katiyar A, Sharma V, Kumar V, Verma C, Gahlawat SK. Genetic Analysis of HIBM Myopathy-Specific GNE V727M Hotspot Mutation Identifies a Novel COL6A3 Allied Gene Signature That Is Also Deregulated in Multiple Neuromuscular Diseases and Myopathies. Genes. 2023; 14(3):567. https://doi.org/10.3390/genes14030567
Chicago/Turabian StyleAttri, Shivangi, Moien Lone, Amit Katiyar, Vikas Sharma, Vinay Kumar, Chaitenya Verma, and Suresh Kumar Gahlawat. 2023. "Genetic Analysis of HIBM Myopathy-Specific GNE V727M Hotspot Mutation Identifies a Novel COL6A3 Allied Gene Signature That Is Also Deregulated in Multiple Neuromuscular Diseases and Myopathies" Genes 14, no. 3: 567. https://doi.org/10.3390/genes14030567