
LncPLAAT3-AS Regulates PLAAT3-Mediated Adipocyte Differentiation and Lipogenesis in Pigs through miR-503-5p
 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Cell Culture and Differentiation
2.3. Cell Transfection
2.4. Western Blot Assay
2.5. Luciferase Reporter Assay
2.6. Assessment of Cell Proliferation via CCK-8 and EdU Assays
2.7. RNA Isolation and Reverse Transcription
2.8. Real-Time Quantitative PCR
2.9. Oil Red O Staining
2.10. RNA-Seq and Collection of Sequencing Data
2.11. Data Analysis and Statistics
3. Results
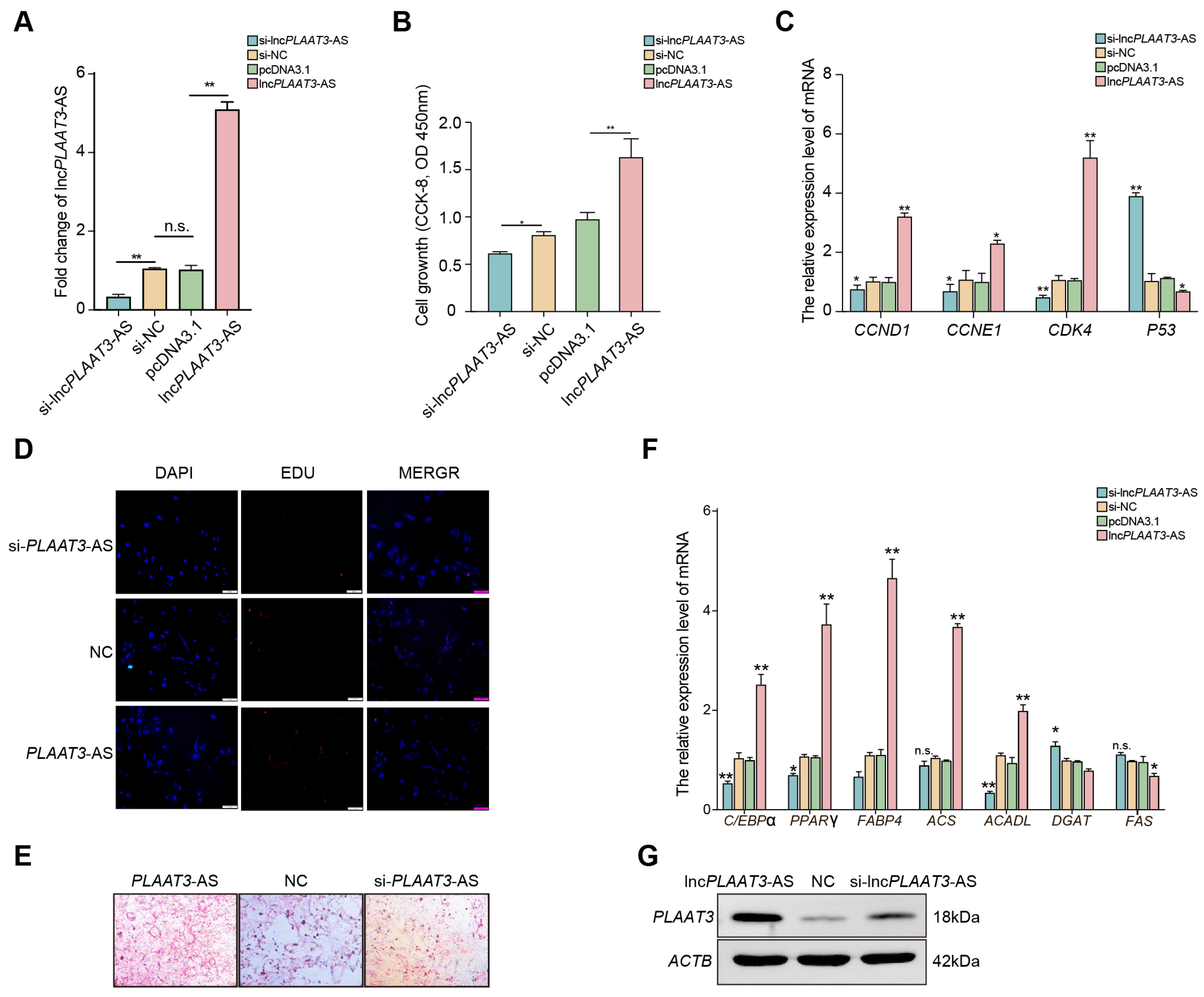
3.1. LncPLAAT3-AS Regulates Porcine Primary Preadipocyte Proliferation
3.2. LncPLAAT3 Promotes Porcine Primary Preadipocyte Differentiation
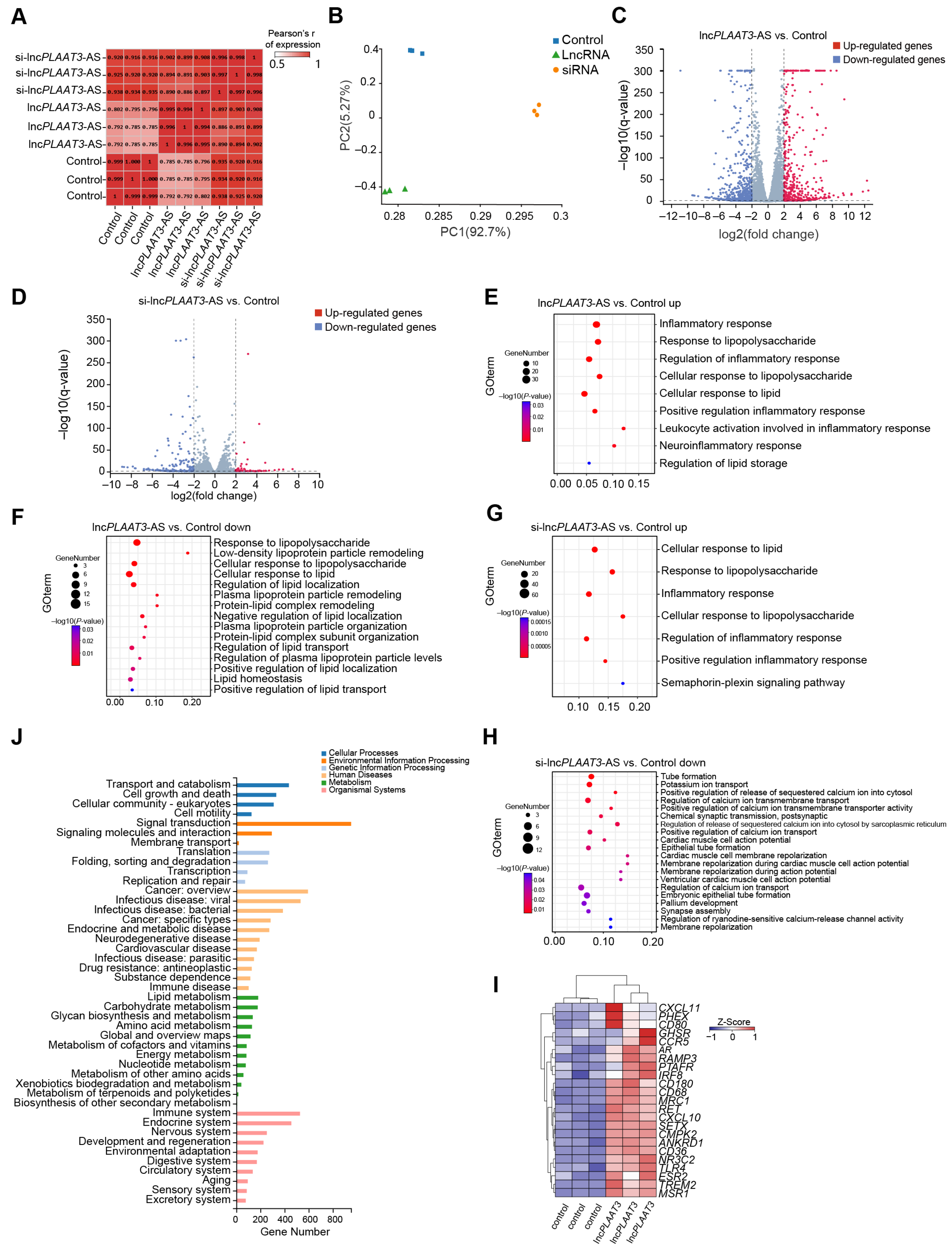
3.3. Differences in Gene Expression in Adipocytes after lncPLAAT3 Overexpression or Knockdown
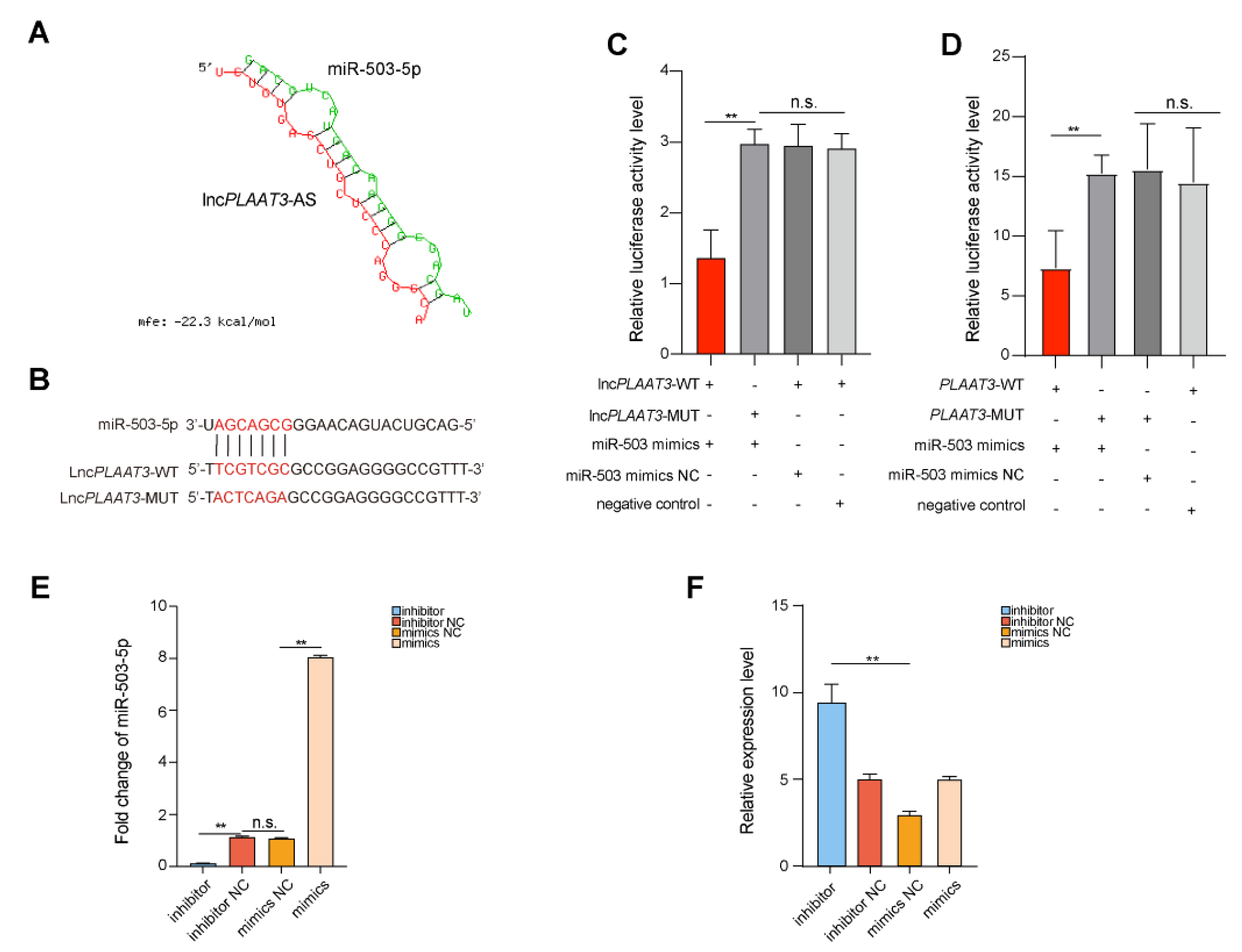
3.4. LncPLAAT3-AS Targets miR-503-5p during Adipocyte Differentiation
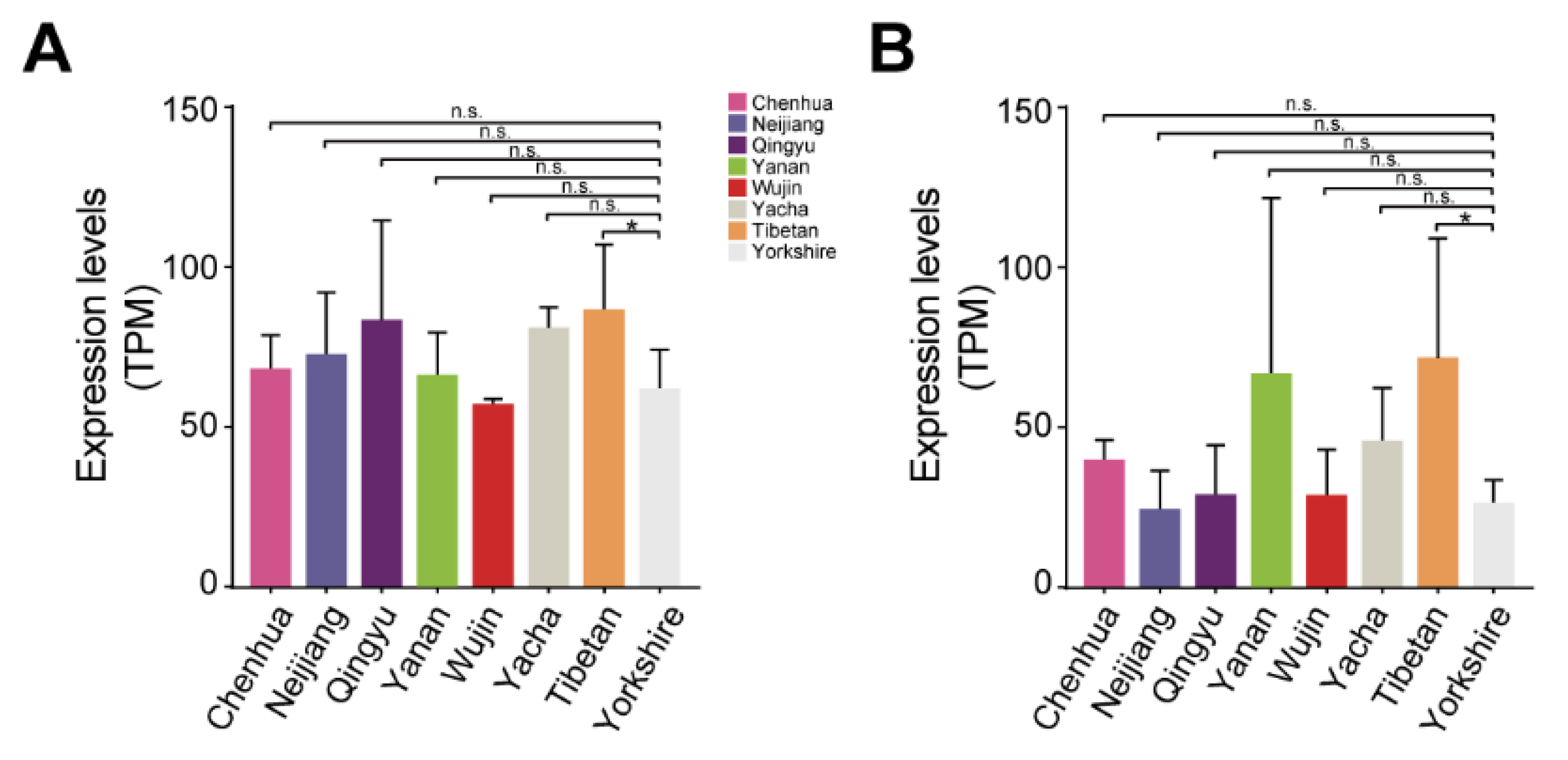
3.5. Differences in lncPLAAT3-AS and PLAAT3 Proliferation among Different Pig Breeds
3.6. SNPs of lncPLAAT3-AS and PLAAT3
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Liao, Q.; Liu, C.; Yuan, X.; Kang, S.; Miao, R.; Xiao, H.; Zhao, G.; Luo, H.; Bu, D.; Zhao, H.; et al. Large-scale prediction of long non-coding RNA functions in a coding–non-coding gene co-expression network. Nucleic Acids Res. 2011, 39, 3864–3878. [Google Scholar] [CrossRef] [PubMed]
- He, H.; Wang, Y.; Ye, P.; Yi, D.; Cheng, Y.; Tang, H.; Zhu, Z.; Wang, X.; Jin, S. Long noncoding RNA ZFPM2-AS1 acts as a miRNA sponge and promotes cell invasion through regulation of miR-139/GDF10 in hepatocellular carcinoma. J. Exp. Clin. Cancer Res. 2020, 39, 159. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Li, H.; Zhang, M.; Li, R. LncRNA GAS6 antisense RNA 1 facilitates the tumorigenesis of clear cell renal cell carcinoma by regulating the AMP-activated protein kinase/mTOR signaling pathway. Oncol. Lett. 2021, 22, 727. [Google Scholar] [CrossRef]
- Zhang, H.; Guan, R.; Zhang, Z.; Li, D.; Xu, J.; Gong, Y.; Chen, X.; Lu, W. LncRNA Nqo1-AS1 Attenuates Cigarette Smoke-Induced Oxidative Stress by Upregulating its Natural Antisense Transcript Nqo1. Front. Pharmacol. 2021, 12, 729062. [Google Scholar] [CrossRef] [PubMed]
- Kuo, F.-C.; Neville, M.J.; Sabaratnam, R.; Wesolowska-Andersen, A.; Phillips, D.; Wittemans, L.B.; van Dam, A.D.; Loh, N.Y.; Todorčević, M.; Denton, N.; et al. HOTAIR interacts with PRC2 complex regulating the regional preadipocyte transcriptome and human fat distribution. Cell Rep. 2022, 40, 111136. [Google Scholar] [CrossRef]
- Meurens, F.; Summerfield, A.; Nauwynck, H.; Saif, L.; Gerdts, V. The pig: A model for human infectious diseases. Trends Microbiol. 2012, 20, 50–57. [Google Scholar] [CrossRef]
- Rosen, E.D.; MacDougald, O.A. Adipocyte differentiation from the inside out. Nat. Rev. Mol. Cell Biol. 2006, 7, 885–896. [Google Scholar] [CrossRef]
- Groenen, M.A.M.; Archibald, A.L.; Uenishi, H.; Tuggle, C.K.; Takeuchi, Y.; Rothschild, M.F.; Rogel-Gaillard, C.; Park, C.; Milan, D.; Megens, H.-J.; et al. Analyses of pig genomes provide insight into porcine demography and evolution. Nature 2012, 491, 393–398. [Google Scholar] [CrossRef] [Green Version]
- Pabst, R. The pig as a model for immunology research. Cell Tissue Res. 2020, 380, 287–304. [Google Scholar] [CrossRef]
- Jaworski, K.; Ahmadian, M.; Duncan, R.E.; Sarkadi-Nagy, E.; Varady, K.A.; Hellerstein, M.K.; Lee, H.-Y.; Samuel, V.T.; Shulman, G.I.; Kim, K.-H.; et al. AdPLA ablation increases lipolysis and prevents obesity induced by high-fat feeding or leptin deficiency. Nat. Med. 2009, 15, 159–168. [Google Scholar] [CrossRef]
- Liu, P.; Jin, L.; Zhao, L.; Long, K.; Song, Y.; Tang, Q.; Ma, J.; Wang, X.; Tang, G.; Jiang, Y.; et al. Identification of a novel antisense long non-coding RNA PLA2G16-AS that regulates the expression of PLA2G16 in pigs. Gene 2018, 671, 78–84. [Google Scholar] [CrossRef]
- Du, J.; Xu, Y.; Zhang, P.; Zhao, X.; Gan, M.; Li, Q.; Ma, J.; Tang, G.; Jiang, Y.; Wang, J.; et al. MicroRNA-125a-5p Affects Adipocytes Proliferation, Differentiation and Fatty Acid Composition of Porcine Intramuscular Fat. Int. J. Mol. Sci. 2018, 19, 501. [Google Scholar] [CrossRef] [Green Version]
- Mills, J.D.; Kawahara, Y.; Janitz, M. Strand-Specific RNA-Seq Provides Greater Resolution of Transcriptome Profiling. Curr. Genom. 2013, 14, 173–181. [Google Scholar] [CrossRef]
- Papasavva, P.; Papaioannou, N.; Patsali, P.; Kurita, R.; Nakamura, Y.; Sitarou, M.; Christou, S.; Kleanthous, M.; Lederer, C. Distinct miRNA Signatures and Networks Discern Fetal from Adult Erythroid Differentiation and Primary from Immortalized Erythroid Cells. Int. J. Mol. Sci. 2021, 22, 3626. [Google Scholar] [CrossRef]
- Tao, X.; Liang, Y.; Yang, X.; Pang, J.; Zhong, Z.; Chen, X.; Yang, Y.; Zeng, K.; Kang, R.; Lei, Y.; et al. Transcriptomic profiling in muscle and adipose tissue identifies genes related to growth and lipid deposition. PLoS ONE 2017, 12, e0184120. [Google Scholar] [CrossRef] [Green Version]
- Shang, P.; Li, W.; Liu, G.; Zhang, J.; Li, M.; Wu, L.; Wang, K.; Chamba, Y. Identification of lncRNAs and Genes Responsible for Fatness and Fatty Acid Composition Traits between the Tibetan and Yorkshire Pigs. J. Genom. 2019, 2019, 5070975-12. [Google Scholar] [CrossRef]
- Garin-Shkolnik, T.; Rudich, A.; Hotamisligil, G.S.; Rubinstein, M. FABP4 Attenuates PPARγ and Adipogenesis and Is Inversely Correlated With PPARγ in Adipose Tissues. Diabetes 2014, 63, 900–911. [Google Scholar] [CrossRef] [Green Version]
- Gondret, F.; Riquet, J.; Tacher, S.; Demars, J.; Sanchez, M.; Billon, Y.; Robic, A.; Bidanel, J.; Milan, D. Towards candidate genes affecting body fatness at the SSC7 QTL by expression analyses. J. Anim. Breed. Genet. 2011, 129, 316–324. [Google Scholar] [CrossRef]
- Pepino, M.Y.; Kuda, O.; Samovski, D.; Abumrad, N.A. Structure-function of CD36 and importance of fatty acid signal transduction in fat metabolism. Annu. Rev. Nutr. 2014, 34, 281–303. [Google Scholar] [CrossRef] [Green Version]
- Guo, C.; Yuan, L.; Liu, X.; Du, A.; Huang, Y.; Zhang, L. Effect of ARB on expression of CD68 and MCP-1 in adipose tissue of rats on long-term high fat diet. J. Huazhong Univ. Sci. Technol. 2008, 28, 257–260. [Google Scholar] [CrossRef]
- Chan, P.-C.; Liao, M.-T.; Lu, C.-H.; Tian, Y.-F.; Hsieh, P.-S. Targeting inhibition of CCR5 on improving obesity-associated insulin resistance and impairment of pancreatic insulin secretion in high fat-fed rodent models. Eur. J. Pharmacol. 2021, 891, 173703. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Li, P.; Li, H.; Wang, S.; Ding, L.; Wang, H.; Ye, H.; Jin, Y.; Hou, J.; Fang, X.; et al. TREM2 regulates obesity-induced insulin resistance via adipose tissue remodeling in mice of high-fat feeding. J. Transl. Med. 2019, 17, 300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, D.; She, J.; Hu, X.; Zhang, M.; Sun, R.; Qin, S. The ELF3-regulated lncRNA UBE2CP3 is over-stabilized by RNA-RNA interactions and drives gastric cancer metastasis via miR-138-5p/ITGA2 axis. Oncogene 2021, 40, 5403–5415. [Google Scholar] [CrossRef] [PubMed]
- Sabarinathan, R.; Tafer, H.; Seemann, S.E.; Hofacker, I.L.; Stadler, P.F.; Gorodkin, J. RNAsnp: Efficient detection of local RNA secondary structure changes induced by SNPs. Hum. Mutat. 2013, 34, 546–556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, L.; He, J.; Zhao, Y.; Niu, L.; Chen, L.; Tang, G.; Jiang, Y.; Hao, X.; Bai, L.; Li, X.; et al. MicroRNA-126b-5p Exacerbates Development of Adipose Tissue and Diet-Induced Obesity. Int. J. Mol. Sci. 2021, 22, 10261. [Google Scholar] [CrossRef]
- Hu, M.; Kuang, R.; Guo, Y.; Ma, R.; Hou, Y.; Xu, Y.; Qi, X.; Wang, D.; Zhou, H.; Xiong, Y.; et al. Epigenomics analysis of miRNA cis-regulatory elements in pig muscle and fat tissues. Genomics 2022, 114, 110276. [Google Scholar] [CrossRef]
- Klingenberg, M.; Matsuda, A.; Diederichs, S.; Patel, T. Non-coding RNA in hepatocellular carcinoma: Mechanisms, biomarkers and therapeutic targets. J. Hepatol. 2017, 67, 603–618. [Google Scholar] [CrossRef] [Green Version]
- Lang, Y.-Y.; Xu, X.-Y.; Liu, Y.-L.; Ye, C.-F.; Hu, N.; Yao, Q.; Cheng, W.-S.; Cheng, Z.-G.; Liu, Y. Ghrelin Relieves Obesity-Induced Myocardial Injury by Regulating the Epigenetic Suppression of miR-196b Mediated by lncRNA HOTAIR. Obes. Facts 2022, 15, 540–549. [Google Scholar] [CrossRef]
- Kuda, O.; Jenkins, C.M.; Skinner, J.R.; Moon, S.H.; Su, X.; Gross, R.W.; Abumrad, N.A. CD36 protein is involved in store-operated calcium flux, phospholipase A2 activation, and production of prostaglandin E2. J. Biol. Chem. 2011, 286, 17785–17795. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Chen, L.; Tian, S.; Lin, Y.; Tang, Q.; Zhou, X.; Li, D.; Yeung, C.K.; Che, T.; Jin, L.; et al. Comprehensive variation discovery and recovery of missing sequence in the pig genome using multiple de novo assemblies. Genome Res. 2016, 27, 865–874. [Google Scholar] [CrossRef]
- Squillaro, T.; Peluso, G.; Galderisi, U.; Di Bernardo, G. Long non-coding RNAs in regulation of adipogenesis and adipose tissue function. eLife 2020, 9, e59053. [Google Scholar] [CrossRef]
- Kosinska-Selbi, B.; Mielczarek, M.; Szyda, J. Review: Long non-coding RNA in livestock. Animal 2020, 14, 2003–2013. [Google Scholar] [CrossRef]
- Bridges, M.C.; Daulagala, A.C.; Kourtidis, A. LNCcation: lncRNA localization and function. J. Cell Biol. 2021, 220, 110276. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequence (5′ to 3′) | Product Size (bp) |
|---|---|---|
| PLA2G16 | F: ATATGTGGTCCACCTGGCTCCCC | 395 |
| R: ATTGCTTTTGCCGCTTGTTTCTG | ||
| PLA2G16-AS | F: GGACTCTGCGGCCATTTAAC | 213 |
| R: GCTTTGGGACAATGAGTCGC | ||
| GAPDH | F: CCCCTTCATTGACCTCCACT | 192 |
| R: CCATTTGATGTTGGCGGGAT | ||
| CCND1 | F: GCATGTTCGTGGCCTCTAAG | 228 |
| R: CGTGTTTGCGGATGATCTGT | ||
| CDK4 | F: TCAGCACAGTTCGTGAGGTG | 77 |
| R: GTCCATCAGCCGGACAACAT | ||
| P53 | F: GGGACGGAACAGCTTTGA | 161 |
| R: TTTGCACTGGCGAGGAG | ||
| PPARγ | F: CTCCAAGAATACCAAAGTGCGA | 150 |
| R: GCCTGATGCTTTATCCCCACA | ||
| C/EBPα | F: CAAGAACAGCAACGAGTACCG | 124 |
| R: GTCACTGGTCAACTCCAGCAC | ||
| FABP4 | F: GAAGTGGGAGTGGGCTTT | 190 |
| R: TTATGGTGCTCTTGACTTTCCT | ||
| ACS | F: GCAGGCAGGCTCAGTTT | 129 |
| R: CTCTGTTCAGGGGAGGGT | ||
| ACADL | F: TGTCTCCAGCTGCATGAAACGA | 107 |
| R: AGCTGCACACAGTCATAAGCCA | ||
| DGAT | F: CCTACCGCGATCTCTACTACTT | 126 |
| R: GGGTGAAGAACAGCATCTCAA | ||
| FAS | F: CCAACCAGCAACACCAA | 100 |
| R: CAGGTACGGGAATGAGGA | ||
| ssc-miR-503 | UAGCAGCGGGAACAGUACUGCAG | |
| U6 | F: CGCTTCGGCAGCACATATAC | 87 |
| R: TTCACGAATTTGCGTGTCAT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Z.; Chai, J.; Wang, Y.; Gu, Y.; Long, K.; Li, M.; Jin, L. LncPLAAT3-AS Regulates PLAAT3-Mediated Adipocyte Differentiation and Lipogenesis in Pigs through miR-503-5p. Genes 2023, 14, 161. https://doi.org/10.3390/genes14010161
Wang Z, Chai J, Wang Y, Gu Y, Long K, Li M, Jin L. LncPLAAT3-AS Regulates PLAAT3-Mediated Adipocyte Differentiation and Lipogenesis in Pigs through miR-503-5p. Genes. 2023; 14(1):161. https://doi.org/10.3390/genes14010161
Chicago/Turabian StyleWang, Zhiming, Jin Chai, Yuhao Wang, Yiren Gu, Keren Long, Mingzhou Li, and Long Jin. 2023. "LncPLAAT3-AS Regulates PLAAT3-Mediated Adipocyte Differentiation and Lipogenesis in Pigs through miR-503-5p" Genes 14, no. 1: 161. https://doi.org/10.3390/genes14010161