
Current Analytical Methods and Research Trends Are Used to Identify Domestic Pig and Wild Boar DNA in Meat and Meat Products

Abstract
:1. Introduction
2. Pig and Wild Boar Phylogenetics
3. Authentication of Meat from Domestic Pig
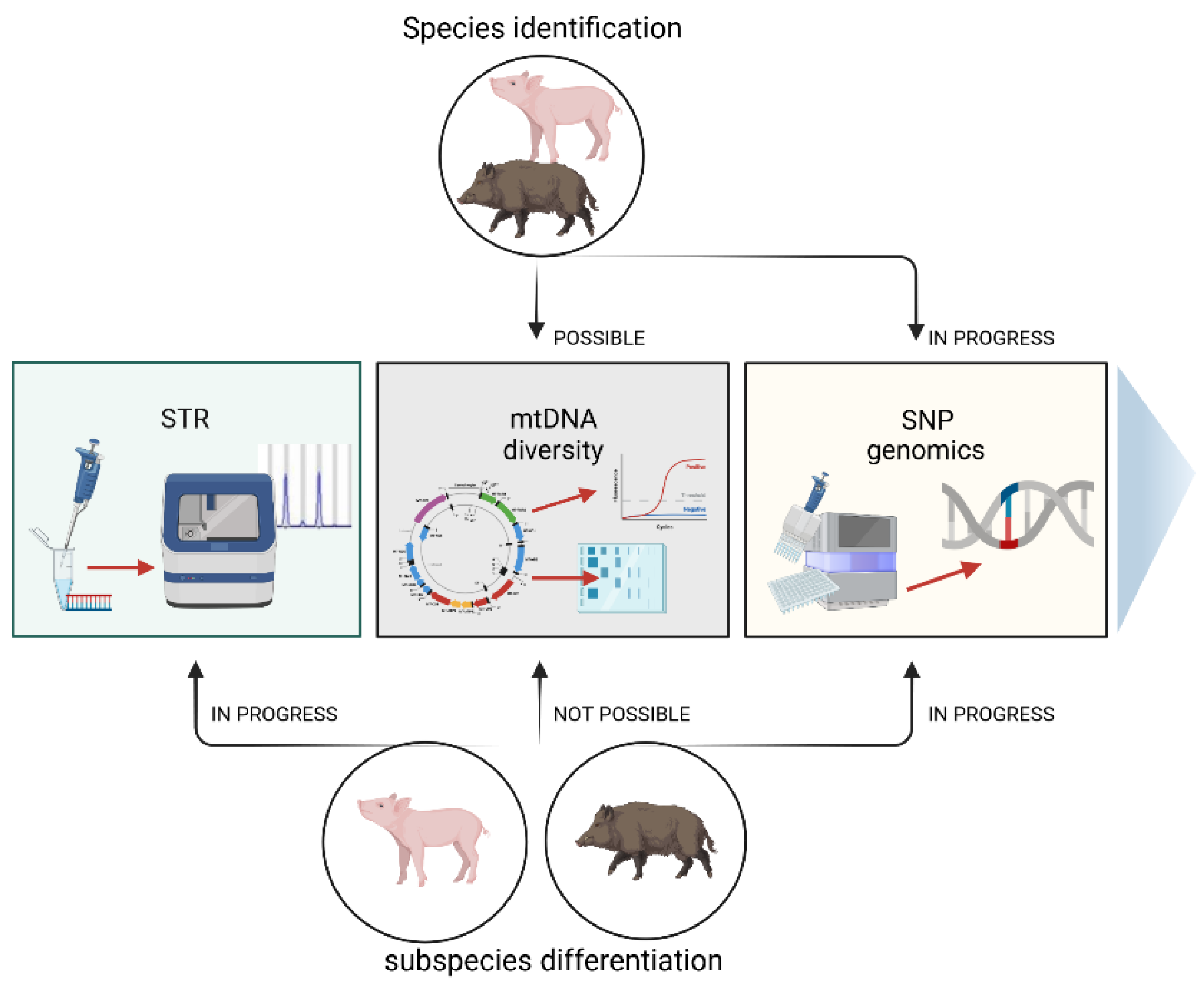
4. Methods Used for Species Identification—Possibilities and Application
4.1. PCR
4.2. PCR RAPD
4.3. PCR-RFLP
4.4. Real-Time PCR (Quantification and Qualification)
4.5. Digital PCR
4.6. LAMP
4.7. DNA Barcoding and NGS
5. Genetic Markers Used for Pig and Wild-Boar Distinguishing
5.1. Mitochondrial DNA Genome Loci
5.2. Melanocortin 1 Receptor Gene (MC1R)
5.3. Nuclear Receptor Subfamily 6, Group A, Member 1 (NR6A1)
5.4. Short Tandem Repeats (STRs)
5.5. Other Potential Genetic Markers
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bassols, A.; Costa, C.; Eckersall, P.D.; Osada, J.; Sabrià, J.; Tibau, J. The pig as an animal model for human pathologies: A proteomics perspective. Proteom.—Clin. Appl. 2014, 8, 715–731. [Google Scholar] [CrossRef]
- Ropodi, A.; Pavlidis, D.; Mohareb, F.; Panagou, E.; Nychas, G.-J. Multispectral image analysis approach to detect adulteration of beef and pork in raw meats. Food Res. Int. 2014, 67, 12–18. [Google Scholar] [CrossRef]
- Alamprese, C.; Casale, M.; Sinelli, N.; Lanteri, S.; Casiraghi, E. Detection of minced beef adulteration with turkey meat by UV–vis, NIR and MIR spectroscopy. LWT 2013, 53, 225–232. [Google Scholar] [CrossRef]
- Cai, S.; Kong, F.; Xu, S. Detection of porcine-derived ingredients from adulterated meat based on real-time loop-mediated isothermal amplification. Mol. Cell Probes 2020, 53, 101609. [Google Scholar] [CrossRef] [PubMed]
- Niño, A.M.; Granja, R.H.; Wanschel, A.C.; Salerno, A.G. The challenges of ractopamine use in meat production for export to European Union and Russia. Food Control 2017, 72, 289–292. [Google Scholar] [CrossRef]
- King, H.; Bedale, W. Hazard Analysis and Risk-Based Preventive Controls: Improving Food Safety in Human Food Manufacturing for Food Businesses; Academic Press: Cambridge, MA, USA, 2017; ISBN 978-0-12-809475-4. [Google Scholar]
- Groenen, M.A.M.; Archibald, A.L.; Uenishi, H.; Tuggle, C.K.; Takeuchi, Y.; Rothschild, M.F.; Rogel-Gaillard, C.; Park, C.; Milan, D.; Megens, H.-J.; et al. Analyses of pig genomes provide insight into porcine demography and evolution. Nature 2012, 491, 393–398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, B.; Cui, L.; Perez-Enciso, M.; Traspov, A.; Crooijmans, R.P.M.A.; Zinovieva, N.; Schook, L.B.; Archibald, A.; Gatphayak, K.; Knorr, C.; et al. Genome-wide SNP data unveils the globalization of domesticated pigs. Genet. Sel. Evol. 2017, 49, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larson, G.; Burger, J. A population genetics view of animal domestication. Trends Genet. 2013, 29, 197–205. [Google Scholar] [CrossRef]
- Scandura, M.; Iacolina, L.; Crestanello, B.; Pecchioli, E.; DI Benedetto, M.F.; Russo, V.; Davoli, R.; Apollonio, M.; Bertorelle, G. Ancient vs. recent processes as factors shaping the genetic variation of the European wild boar: Are the effects of the last glaciation still detectable? Mol. Ecol. 2008, 17, 1745–1762. [Google Scholar] [CrossRef]
- Fontanesi, L.; Ribani, A.; Scotti, E.; Utzeri, V.; Veličković, N.; Dall’Olio, S. Differentiation of meat from European wild boars and domestic pigs using polymorphisms in the MC1R and NR6A1 genes. Meat Sci. 2014, 98, 781–784. [Google Scholar] [CrossRef]
- Iacolina, L.; Pertoldi, C.; Amills, M.; Kusza, S.; Megens, H.-J.; Bâlteanu, V.A.; Bakan, J.; Cubric-Curik, V.; Oja, R.; Saarma, U.; et al. Hotspots of recent hybridization between pigs and wild boars in Europe. Sci. Rep. 2018, 8, 17372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mary, N.; Iannuccelli, N.; Petit, G.; Bonnet, N.; Pinton, A.; Barasc, H.; Amélie, F.; Calgaro, A.; Grosbois, V.; Servin, B.; et al. Genome-wide analysis of hybridization in wild boar populations reveals adaptive introgression from domestic pig. Evol. Appl. 2022, 15, 1115–1128. [Google Scholar] [CrossRef] [PubMed]
- Dzialuk, A.; Zastempowska, E.; Skórzewski, R.; Twarużek, M.; Grajewski, J. High domestic pig contribution to the local gene pool of free-living European wild boar: A case study in Poland. Mammal Res. 2017, 63, 65–71. [Google Scholar] [CrossRef] [Green Version]
- Babicz, M.; Pastwa, M.; Skrzypczak, E.; Buczyński, J.T. Variability in themelanocortin 1 receptor(MC1R) gene in wild boars and local pig breeds in Poland. Anim. Genet. 2013, 44, 357–358. [Google Scholar] [CrossRef]
- Musto, M. DNA Quality and Integrity of Nuclear and Mitochondrial Sequences from Beef Meat as Affected by Different Cooking Methods. Food Technol. Biotechnol. 2011, 49, 523–528. [Google Scholar]
- Natonek-Wiśniewska, M.; Krzyścin, P.; Piestrzyńska-Kajtoch, A. The species identification of bovine, porcine, ovine and chicken components in animal meals, feeds and their ingredients, based on COX I analysis and ribosomal DNA sequences. Food Control 2013, 34, 69–78. [Google Scholar] [CrossRef]
- Tan, L.L.; Ahmed, S.A.; Ng, S.K.; Citartan, M.; Raabe, C.A.; Rozhdestvensky, T.S.; Tang, T.H. Rapid detection of porcine DNA in processed food samples using a streamlined DNA extraction method combined with the SYBR Green real-time PCR assay. Food Chem. 2019, 309, 125654. [Google Scholar] [CrossRef]
- Prusakova, O.V.; Glukhova, X.A.; Afanas’Eva, G.V.; Trizna, Y.A.; Nazarova, L.F.; Beletsky, I.P. A simple and sensitive two-tube multiplex PCR assay for simultaneous detection of ten meat species. Meat Sci. 2018, 137, 34–40. [Google Scholar] [CrossRef]
- Natonek-Wiśniewska, M.; Krzyścin, P. Evaluation of the suitability of mitochondrial DNA for species identification of microtraces and forensic traces. Acta Biochim. Pol. 2017, 64, 705–708. [Google Scholar] [CrossRef] [Green Version]
- Man-an, S.K.A.; Fadilah, A.R.; Mardhiyyah, S. Contemporary Issues and Development in the Global Halal Industry; Ab.Manan, S.K., Abd Rahman, F., Sahri, M., Eds.; Springer: Singapore, 2016; ISBN 978-981-10-1450-5. [Google Scholar]
- Mane, B.G.; Tanwar, V.K.; Girish, P.S.; Sharma, D.; Dixit, V.P. RAPD Markers for Differentiation of Meat Species. Indian J. Vet. Res. 2008, 17, 9–13. [Google Scholar]
- Nakyinsige, K.; Man, Y.B.C.; Sazili, A.Q. Halal authenticity issues in meat and meat products. Meat Sci. 2012, 91, 207–214. [Google Scholar] [CrossRef] [PubMed]
- Chappal-war, A.M.; Pathak, V.; Goswami, M.; Sharma, B.; Singh, P.; Mishra, R. Recent Novel Techniques Applied for Detection of Meat Adulteration and Fraudulent Practices. Indian J. Vet. Public Health 2020, 7, 1–6. [Google Scholar]
- Rahmati, S.; Julkapli, N.M.; Yehye, W.A.; Basirun, W.J. Identification of meat origin in food products–A review. Food Control 2016, 68, 379–390. [Google Scholar] [CrossRef]
- Hossain, M.A.M.; Ali, E.; Hamid, S.B.A.; Asing; Mustafa, S.; Desa, M.N.M.; Zaidul, I.S.M. Double Gene Targeting Multiplex Polymerase Chain Reaction–Restriction Fragment Length Polymorphism Assay Discriminates Beef, Buffalo, and Pork Substitution in Frankfurter Products. J. Agric. Food Chem. 2016, 64, 6343–6354. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Oceja, A.; Nuñez, C.; Baeta, M.; Gamarra, D.; de Pancorbo, M. Species identification in meat products: A new screening method based on high resolution melting analysis of cyt b gene. Food Chem. 2017, 237, 701–706. [Google Scholar] [CrossRef]
- Thanakiatkrai, P.; Kitpipit, T. Meat species identification by two direct-triplex real-time PCR assays using low resolution melting. Food Chem. 2017, 233, 144–150. [Google Scholar] [CrossRef]
- Natonek-Wiśniewska, M.; Krzyścin, P. Development of Easy and Effective Real-Time PCR tests to Identify Bovine, Porcine, and Ovine Components in Food. Zywnosc Nauk. Technol. Jakosc/Food Sci. Technol. Qual. 2015, 22, 73–84. [Google Scholar] [CrossRef]
- Shehata, H.R.; Li, J.; Chen, S.; Redda, H.; Cheng, S.; Tabujara, N.; Li, H.; Warriner, K.; Hanner, R. Droplet digital polymerase chain reaction (ddPCR) assays integrated with an internal control for quantification of bovine, porcine, chicken and turkey species in food and feed. PLoS ONE 2017, 12, e0182872. [Google Scholar] [CrossRef]
- Köppel, R.; Ganeshan, A.; Weber, S.; Pietsch, K.; Graf, C.; Hochegger, R.; Griffiths, K.; Burkhardt, S. Duplex digital PCR for the determination of meat proportions of sausages containing meat from chicken, turkey, horse, cow, pig and sheep. Eur. Food Res. Technol. 2019, 245, 853–862. [Google Scholar] [CrossRef]
- Lee, S.-Y.; Kim, M.-J.; Hong, Y.; Kim, H.-Y. Development of a rapid on-site detection method for pork in processed meat products using real-time loop-mediated isothermal amplification. Food Control 2016, 66, 53–61. [Google Scholar] [CrossRef]
- Ran, G.; Ren, L.; Han, X.; Liu, X.; Li, Z.; Pang, D.; Ouyang, H.; Tang, X. Development of a Rapid Method for the Visible Detection of Pork DNA in Halal Products by Loop-Mediated Isothermal Amplification. Food Anal. Methods 2015, 9, 565–570. [Google Scholar] [CrossRef]
- Riztyan; Takasaki, K.; Yamakoshi, Y.; Futo, S. Single-Laboratory Validation of Rapid and Easy DNA Strip for Porcine DNA Detection in Beef Meatballs. J. AOAC Int. 2018, 101, 1653–1656. [Google Scholar] [CrossRef] [PubMed]
- Tasrip, N.A.; Mokhtar Khairil, N.F.; Hanapi, U.K.; Abdul Manaf, Y.N.; Ali, M.E.; Cheah, Y.K. No Title Loop Mediated Isothermal Amplification; a Review on Its Application and Strategy in Animal Species Authentication of Meat Based Food Products. Int. Food Res. J. 2019, 26, 1–10. [Google Scholar]
- Naaum, A.M.; Shehata, H.R.; Chen, S.; Li, J.; Tabujara, N.; Awmack, D.; Lutze-Wallace, C.; Hanner, R. Complementary molecular methods detect undeclared species in sausage products at retail markets in Canada. Food Control 2018, 84, 339–344. [Google Scholar] [CrossRef]
- Haynes, E.; Jimenez, E.; Pardo, M.A.; Helyar, S.J. The future of NGS (Next Generation Sequencing) analysis in testing food authenticity. Food Control 2019, 101, 134–143. [Google Scholar] [CrossRef]
- Arslan, A.; Ilhak, I.; Calicioglu, M.; Karahan, M. Identification of meats using random amplified polymorphic dna (rapd) technique. J. Muscle Foods 2005, 16, 37–45. [Google Scholar] [CrossRef]
- Jin, L.-G.; Cho, J.-Y.; Seong, K.-B.; Park, J.-Y.; Kong, I.-S.; Hong, Y.-K. 18S rRNA gene sequences and random amplified polymorphic DNA used in discriminating Manchurian trout from other freshwater salmonids. Fish. Sci. 2006, 72, 903–905. [Google Scholar] [CrossRef]
- Natonek-Wiśniewska, M.; Ząbek, T.; Słota, E. Species Identification of Mammalian Mt DNA Using PCR-RFLP. Ann. Anim. Sci. 2007, 7, 305–311. [Google Scholar]
- Natonek-Wiśniewska, M.; Słota, E. A New Method for Species Identification of Poultry Based on 12S-RRNA Fragment Polymorphism. Ann. Anim. Sci. 2009, 9, 127–132. [Google Scholar]
- Sultana, S.; Hossain, M.A.M.; Naquiah, N.N.A.; Ali, E. Novel multiplex PCR-RFLP assay discriminates bovine, porcine and fish gelatin substitution in Asian pharmaceuticals capsule shells. Food Addit. Contam. Part A 2018, 35, 1662–1673. [Google Scholar] [CrossRef]
- Sun, Y.-L.; Lin, C.-S. Establishment and Application of a Fluorescent Polymerase Chain Reaction−Restriction Fragment Length Polymorphism (PCR-RFLP) Method for Identifying Porcine, Caprine, and Bovine Meats. J. Agric. Food Chem. 2003, 51, 1771–1776. [Google Scholar] [CrossRef] [PubMed]
- Guan, F.; Jin, Y.-T.; Zhao, J.; Xu, A.-C.; Luo, Y.-Y. A PCR Method That Can Be Further Developed into PCR-RFLP Assay for Eight Animal Species Identification. J. Anal. Methods Chem. 2018, 2018, 5890140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gargouri, H.; Moalla, N.; Kacem, H.H. PCR–RFLP and species-specific PCR efficiency for the identification of adulteries in meat and meat products. Eur. Food Res. Technol. 2021, 247, 2183–2192. [Google Scholar] [CrossRef]
- Sultana, S.; Hossain, M.M.; Azlan, A.; Johan, M.R.; Chowdhury, Z.Z.; Ali, E. TaqMan probe based multiplex quantitative PCR assay for determination of bovine, porcine and fish DNA in gelatin admixture, food products and dietary supplements. Food Chem. 2020, 325, 126756. [Google Scholar] [CrossRef] [PubMed]
- Janudin, A.A.S.; Chin, N.A.; Ahmed, M.U. An Eva Green Real-Time PCR Assay for Porcine DNA Analysis in Raw and Processed Foods. Malays. J. Halal Res. 2022, 5, 33–39. [Google Scholar] [CrossRef]
- Lubis, H.; Salihah, N.T.; Norizan, N.A.; Hossain, M.M.; Ahmed, M.U. Fast and Sensitive Real-time PCR-based Detection of Porcine DNA in Food Samples by Using EvaGreen Dye. Food Sci. Technol. Res. 2018, 24, 803–810. [Google Scholar] [CrossRef]
- Munir, M.A.; Inayatullah, A. Comparison of real time PCR and conventional PCR by identifying genomic DNA of bovine and porcine. J. Kim. Ter. Indones. 2021, 23, 63–71. [Google Scholar] [CrossRef]
- Basanisi, M.G.; La Bella, G.; Nobili, G.; Coppola, R.; Damato, A.M.; Cafiero, M.A.; La Salandra, G. Application of the novel Droplet digital PCR technology for identification of meat species. Int. J. Food Sci. Technol. 2019, 55, 1145–1150. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, M.U.; Hasan, Q.; Hossain, M.M.; Saito, M.; Tamiya, E. Meat species identification based on the loop mediated isothermal amplification and electrochemical DNA sensor. Food Control 2010, 21, 599–605. [Google Scholar] [CrossRef]
- Girish, P.; Barbuddhe, S.; Kumari, A.; Rawool, D.B.; Karabasanavar, N.S.; Muthukumar, M.; Vaithiyanathan, S. Rapid detection of pork using alkaline lysis- Loop Mediated Isothermal Amplification (AL-LAMP) technique. Food Control 2019, 110, 107015. [Google Scholar] [CrossRef]
- Kumar, Y.; Bansal, S.; Jaiswal, P. Loop-Mediated Isothermal Amplification (LAMP): A Rapid and Sensitive Tool for Quality Assessment of Meat Products. Compr. Rev. Food Sci. Food Saf. 2017, 16, 1359–1378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kane, D.E.; Hellberg, R.S. Identification of species in ground meat products sold on the U.S. commercial market using DNA-based methods. Food Control 2016, 59, 158–163, Corrigendum in 2016, 63, 267. [Google Scholar] [CrossRef]
- Tillmar, A.O.; Dell’Amico, B.; Welander, J.; Holmlund, G. A Universal Method for Species Identification of Mammals Utilizing Next Generation Sequencing for the Analysis of DNA Mixtures. PLoS ONE 2013, 8, e83761. [Google Scholar] [CrossRef] [Green Version]
- Dobrovolny, S.; Blaschitz, M.; Weinmaier, T.; Pechatschek, J.; Cichna-Markl, M.; Indra, A.; Hufnagl, P.; Hochegger, R. Development of a DNA metabarcoding method for the identification of fifteen mammalian and six poultry species in food. Food Chem. 2018, 272, 354–361. [Google Scholar] [CrossRef]
- Xing, R.-R.; Wang, N.; Hu, R.-R.; Zhang, J.-K.; Han, J.-X.; Chen, Y. Application of next generation sequencing for species identification in meat and poultry products: A DNA metabarcoding approach. Food Control 2019, 101, 173–179. [Google Scholar] [CrossRef]
- Cottenet, G.; Blancpain, C.; Chuah, P.F.; Cavin, C. Evaluation and application of a next generation sequencing approach for meat species identification. Food Control 2019, 110, 107003. [Google Scholar] [CrossRef]
- Safdar, M.; Junejo, Y.; Arman, K.; Abasıyanık, M. A highly sensitive and specific tetraplex PCR assay for soybean, poultry, horse and pork species identification in sausages: Development and validation. Meat Sci. 2014, 98, 296–300. [Google Scholar] [CrossRef] [PubMed]
- Fang, M.; Andersson, L. Mitochondrial diversity in European and Chinese pigs is consistent with population expansions that occurred prior to domestication. Proc. R. Soc. B Boil. Sci. 2006, 273, 1803–1810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bruford, M.W.; Bradley, D.G.; Luikart, G. DNA markers reveal the complexity of livestock domestication. Nat. Rev. Genet. 2003, 4, 900–910. [Google Scholar] [CrossRef] [PubMed]
- Koseniuk, A.; Słota, E. Mitochondrial control region diversity in Polish sheep breeds. Arch. Anim. Breed. 2016, 59, 227–233. [Google Scholar] [CrossRef] [Green Version]
- Fajardo, V.; González, I.; Martín, I.; Rojas, M.; Hernández, P.E.; Garcia, T.; Martín, R. Differentiation of European wild boar (Sus scrofa scrofa) and domestic swine (Sus scrofa domestica) meats by PCR analysis targeting the mitochondrial D-loop and the nuclear melanocortin receptor 1 (MC1R) genes. Meat Sci. 2008, 78, 314–322. [Google Scholar] [CrossRef] [PubMed]
- Klungland, H.; Våge, D.I. Pigmentary Switces in Domestic Animal Species. Ann. N. Y. Acad. Sci. 2003, 994, 331–338. [Google Scholar] [CrossRef] [PubMed]
- Lu, D.; Willard, D.; Patel, I.R.; Kadwell, S.; Overton, L.; Kost, T.; Luther, M.; Chen, W.; Woychik, R.P.; Wilkison, W.O.; et al. Agouti Protein Isan Antagonist of the Melanocyte-Stimulating-Hormone Receptor. Nature 1994, 371, 799–802. [Google Scholar] [CrossRef]
- Ollmann, M.M.; Lamoreux, M.L.; Wilson, B.D.; Barsh, G.S. Interaction of Agouti protein with the melanocortin 1 receptor in vitro and in vivo. Genes Dev. 1998, 12, 316–330. [Google Scholar] [CrossRef] [Green Version]
- Kijas, J.M.H.; Wales, R.; Törnsten, A.; Chardon, P.; Moller, M.; Andersson, L. Melanocortin Receptor 1 (MC1R) Mutations and Coat Color in Pigs. Genetics 1998, 150, 1177–1185. [Google Scholar] [CrossRef] [PubMed]
- Koutsogiannouli, E.A.; Moutou, K.A.; Sarafidou, T.; Stamatis, C.; Mamuris, Z. Detection of hybrids between wild boars (Sus scrofa scrofa) and domestic pigs (Sus scrofa f. domestica) in Greece, using the PCR-RFLP method on melanocortin-1 receptor (MC1R) mutations. Mamm. Biol. 2010, 75, 69–73. [Google Scholar] [CrossRef]
- Koseniuk, A.; Smołucha, G.; Natonek-Wiśniewska, M.; Radko, A.; Rubiś, D. Differentiating Pigs from Wild Boars Based on NR6A1 and MC1R Gene Polymorphisms. Animals 2021, 11, 2123. [Google Scholar] [CrossRef] [PubMed]
- Mikawa, S.; Morozumi, T.; Shimanuki, S.-I.; Hayashi, T.; Uenishi, H.; Domukai, M.; Okumura, N.; Awata, T. Fine mapping of a swine quantitative trait locus for number of vertebrae and analysis of an orphan nuclear receptor, germ cell nuclear factor (NR6A1). Genome Res. 2007, 17, 586–593. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaltenbrunner, M.; Mayer, W.; Kerkhoff, K.; Epp, R.; Rüggeberg, H.; Hochegger, R.; Cichna-Markl, M. Differentiation between wild boar and domestic pig in food by targeting two gene loci by real-time PCR. Sci. Rep. 2019, 9, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Beugin, M.-P.; Baubet, E.; De Citres, C.D.; Kaerle, C.; Muselet, L.; Klein, F.; Queney, G. A set of 20 multiplexed single nucleotide polymorphism (SNP) markers specifically selected for the identification of the wild boar (Sus scrofa scrofa) and the domestic pig (Sus scrofa domesticus). Conserv. Genet. Resour. 2017, 9, 671–675. [Google Scholar] [CrossRef]
- Radko, A.; Smołucha, G.; Koseniuk, A. Microsatellite DNA Analysis for Diversity Study, Individual Identification and Parentage Control in Pig Breeds in Poland. Genes 2021, 12, 595. [Google Scholar] [CrossRef] [PubMed]
- Nikolov, I.; Stoeckle, B.; Markov, G.; Kuehn, R. Substantial hybridisation between wild boars (Sus scrofa scrofa) and East Balkan pigs (Sus scrofa f. domestica) in natural environment as a result of semi-wild rearing in Bulgaria. Czech J. Anim. Sci. 2017, 62, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Lorenzini, R.; Fanelli, R.; Tancredi, F.; Siclari, A.; Garofalo, L. Matching STR and SNP genotyping to discriminate between wild boar, domestic pigs and their recent hybrids for forensic purposes. Sci. Rep. 2020, 10, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Goedbloed, D.J.; Megens, H.; Van Hooft, P.; Herrero-Medrano, J.M.; Lutz, W.; Alexandri, P.; Crooijmans, R.P.M.A.; Groenen, M.; Van Wieren, S.E.; Ydenberg, R.C.; et al. Genome-wide single nucleotide polymorphism analysis reveals recent genetic introgression from domestic pigs into Northwest European wild boar populations. Mol. Ecol. 2012, 22, 856–866. [Google Scholar] [CrossRef] [PubMed]
- Rubin, C.-J.; Megens, H.-J.; Barrio, A.M.; Maqbool, K.; Sayyab, S.; Schwochow, D.; Wang, C.; Carlborg, Ö.; Jern, P.; Jørgensen, C.B.; et al. Strong signatures of selection in the domestic pig genome. Proc. Natl. Acad. Sci. USA 2012, 109, 19529–19536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karim, L.; Takeda, H.; Lin, L.; Druet, T.; Arias, J.A.C.; Baurain, D.; Cambisano, N.; Davis, S.R.; Farnir, F.; Grisart, B.; et al. Variants modulating the expression of a chromosome domain encompassing PLAG1 influence bovine stature. Nat. Genet. 2011, 43, 405–413. [Google Scholar] [CrossRef] [PubMed]
- Gudbjartsson, D.F.; Walters, G.B.; Thorleifsson, G.; Stefansson, H.; Halldorsson, B.V.; Zusmanovich, P.; Sulem, P.; Thorlacius, S.; Gylfason, A.; Steinberg, S.; et al. Many sequence variants affecting diversity of adult human height. Nat. Genet. 2008, 40, 609–615. [Google Scholar] [CrossRef]
- Vaysse, A.; Ratnakumar, A.; Derrien, T.; Axelsson, E.; Pielberg, G.R.; Sigurdsson, S.; Fall, T.; Seppälä, E.H.; Hansen, M.S.T.; Lawley, C.T.; et al. Identification of Genomic Regions Associated with Phenotypic Variation between Dog Breeds using Selection Mapping. PLoS Genet. 2011, 7, e1002316. [Google Scholar] [CrossRef] [Green Version]
- Pryce, J.E.; Hayes, B.; Bolormaa, S.; Goddard, M. Polymorphic Regions Affecting Human Height Also Control Stature in Cattle. Genetics 2011, 187, 981–984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Signer-Hasler, H.; Flury, C.; Haase, B.; Burger, D.; Simianer, H.; Leeb, T.; Rieder, S. A Genome-Wide Association Study Reveals Loci Influencing Height and Other Conformation Traits in Horses. PLoS ONE 2012, 7, e37282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomas, G.; Moffatt, P.; Salois, P.; Gaumond, M.-H.; Gingras, R.; Godin, E.; Miao, D.; Goltzman, D.; Lanctôt, C. Osteocrin, a Novel Bone-specific Secreted Protein That Modulates the Osteoblast Phenotype. J. Biol. Chem. 2003, 278, 50563–50571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Banzet, S.; Koulmann, N.; Sanchez, H.; Serrurier, B.; Peinnequin, A.; Bigard, A. Musclin gene expression is strongly related to fast-glycolytic phenotype. Biochem. Biophys. Res. Commun. 2007, 353, 713–718. [Google Scholar] [CrossRef] [PubMed]
- Nishizawa, H.; Matsuda, M.; Yamada, Y.; Kawai, K.; Suzuki, E.; Makishima, M.; Kitamura, T.; Shimomura, I. Musclin, a Novel Skeletal Muscle-derived Secretory Factor. J. Biol. Chem. 2004, 279, 19391–19395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilkinson, S.; Lu, Z.H.; Megens, H.-J.; Archibald, A.L.; Haley, C.; Jackson, I.J.; Groenen, M.A.M.; Crooijmans, R.P.M.A.; Ogden, R.; Wiener, P. Signatures of Diversifying Selection in European Pig Breeds. PLoS Genet. 2013, 9, e1003453. [Google Scholar] [CrossRef] [Green Version]
- Gondret, F.; Riquet, J.; Tacher, S.; Demars, J.; Sanchez, M.; Billon, Y.; Robic, A.; Bidanel, J.; Milan, D. Towards candidate genes affecting body fatness at the SSC7 QTL by expression analyses. J. Anim. Breed. Genet. 2011, 129, 316–324. [Google Scholar] [CrossRef]
- Zhang, Y.; Gao, T.; Hu, S.; Lin, B.; Yan, D.; Xu, Z.; Zhang, Z.; Mao, Y.; Mao, H.; Wang, L.; et al. The Functional SNPs in the 5’ Regulatory Region of the Porcine PPARD Gene Have Significant Association with Fat Deposition Traits. PLoS ONE 2015, 10, e0143734. [Google Scholar] [CrossRef]
- Muñoz, M.; Bozzi, R.; García-Casco, J.; Núñez, Y.; Ribani, A.; Franci, O.; García, F.; Škrlep, M.; Schiavo, G.; Bovo, S.; et al. Genomic diversity, linkage disequilibrium and selection signatures in European local pig breeds assessed with a high density SNP chip. Sci. Rep. 2019, 9, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Groenen, M.A.M. A decade of pig genome sequencing: A window on pig domestication and evolution. Genet. Sel. Evol. 2016, 48, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Moretti, R.; Criscione, A.; Turri, F.; Bordonaro, S.; Marletta, D.; Castiglioni, B.; Chessa, S. A 20-SNP Panel as a Tool for Genetic Authentication and Traceability of Pig Breeds. Animals 2022, 12, 1335. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Technique/Method | Advantages | Limitations and Difficulties | References |
|---|---|---|---|
| PCR | Simple to develop and easy to conduct Can detect small amounts of DNA High specificity species identification Analysis of monoplexes or multiplexes depending on needs Low detection limit Ability to detect processed and heat-treated samples | No quantitation Lower specificity when amplicon length is short More difficult PCR optimisation in case of multiplex PCR Unequal amplification efficiency which causes variable sensitivity Contaminants could produce false-negative results Lower DNA yield for heat-treated samples | [17,18,19,20,21] |
| PCR RAPD | Detects multi-species, and no previous knowledge of DNA is required Can detect a high level of polymorphism (for example, between similar species) Costs per assay are low | Unable to differentiate species in mixtures Mistakes during the analysis of degraded or autoclaved materials Difficulties in obtaining reproducible results | [22,23,24] |
| PCR-RFLP | Species-specific restriction pattern | The results can be challenging to interpret in the case of entirely unknown species components of the studied object Inadequate for highly processed or meat mixtures | [25,26] |
| Real-Time PCR (quantification and qualification) | Possible quantitative result Very high sensitivity High sample throughput | Probe-based methods are expensive and time-intensive Dye-based methods are less accurate Quantification PCR requires appropriate reference material Precision of measure is different for different product types qPCR instruments are very costly HRM analysis requires HRM-capable real-time PCR machines and specialised software algorithms | [27,28,29] |
| Digital PCR | Better detection limit and accuracy | Little sample volume per reaction Small dynamic range if the number of partitions is limited The risk of falsely low quantification due to molecular dropout Restricted multiplex detection | [30,31] |
| LAMP | Rapid High specificity High amplification efficiency No need for thermal cycling Low operational cost | Less versatile Little to no multiplexing Less sensitive to inhibitors than PCR in case of complex samples. The method is more complicated than classical PCR owing to the presence of more primers | [32,33,34,35] |
| DNA barcoding and NGS | High sensitivity and specificity Single-step DNA sequencing and quantification by NGS High throughput detection | High-quality DNA necessary Long analysis time from DNA extraction to the finish of bioinformatics analysis Complex library preparation protocol for NGS Trace quantity of foreign DNA may cause inaccurate estimation of species Costly equipment to analyse and very highly skilled personnel | [36,37] |
| Technique | Marker/Gene | Tested on Sample of | References | |
|---|---|---|---|---|
| Muscle | Meat | |||
| PCR RFLP and real-time PCR | MC1R, NR6A1, mtDNA regions | yes | yes | [11,14,63,68,70,71,72] |
| PCR multiplex | STRs | yes | no | [10,75] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Natonek-Wiśniewska, M.; Piestrzynska-Kajtoch, A.; Koseniuk, A.; Krzyścin, P. Current Analytical Methods and Research Trends Are Used to Identify Domestic Pig and Wild Boar DNA in Meat and Meat Products. Genes 2022, 13, 1825. https://doi.org/10.3390/genes13101825
Natonek-Wiśniewska M, Piestrzynska-Kajtoch A, Koseniuk A, Krzyścin P. Current Analytical Methods and Research Trends Are Used to Identify Domestic Pig and Wild Boar DNA in Meat and Meat Products. Genes. 2022; 13(10):1825. https://doi.org/10.3390/genes13101825
Chicago/Turabian StyleNatonek-Wiśniewska, Małgorzata, Agata Piestrzynska-Kajtoch, Anna Koseniuk, and Piotr Krzyścin. 2022. "Current Analytical Methods and Research Trends Are Used to Identify Domestic Pig and Wild Boar DNA in Meat and Meat Products" Genes 13, no. 10: 1825. https://doi.org/10.3390/genes13101825