
Psychrophilic Yeasts: Insights into Their Adaptability to Extremely Cold Environments

Abstract
:1. Introduction
2. Material and Methods
2.1. Data Collection and Genome Annotation
2.2. Analyses of Metabolic Pathway Enrichment
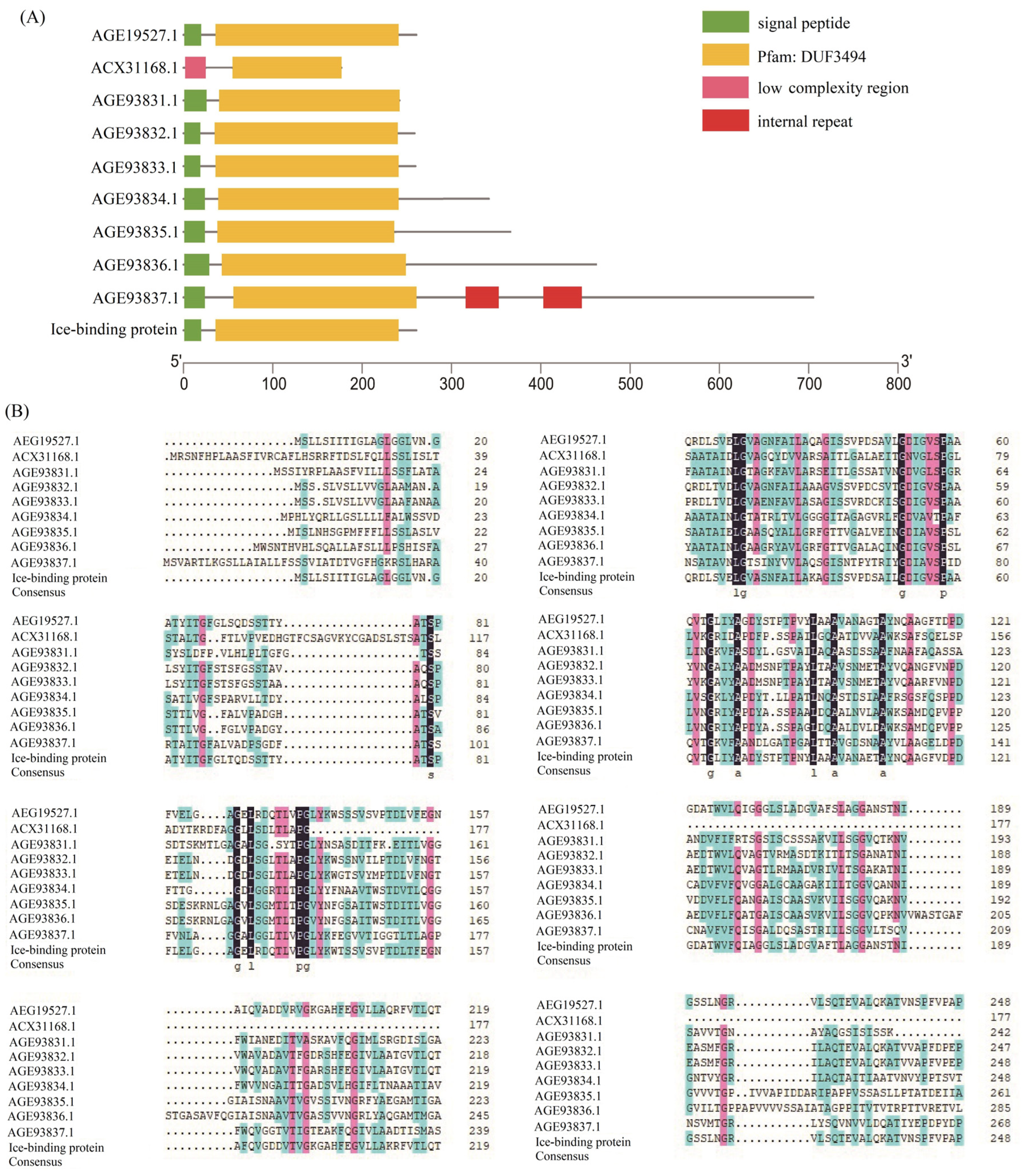
2.3. Homologous Gene Identification and Domain Analysis
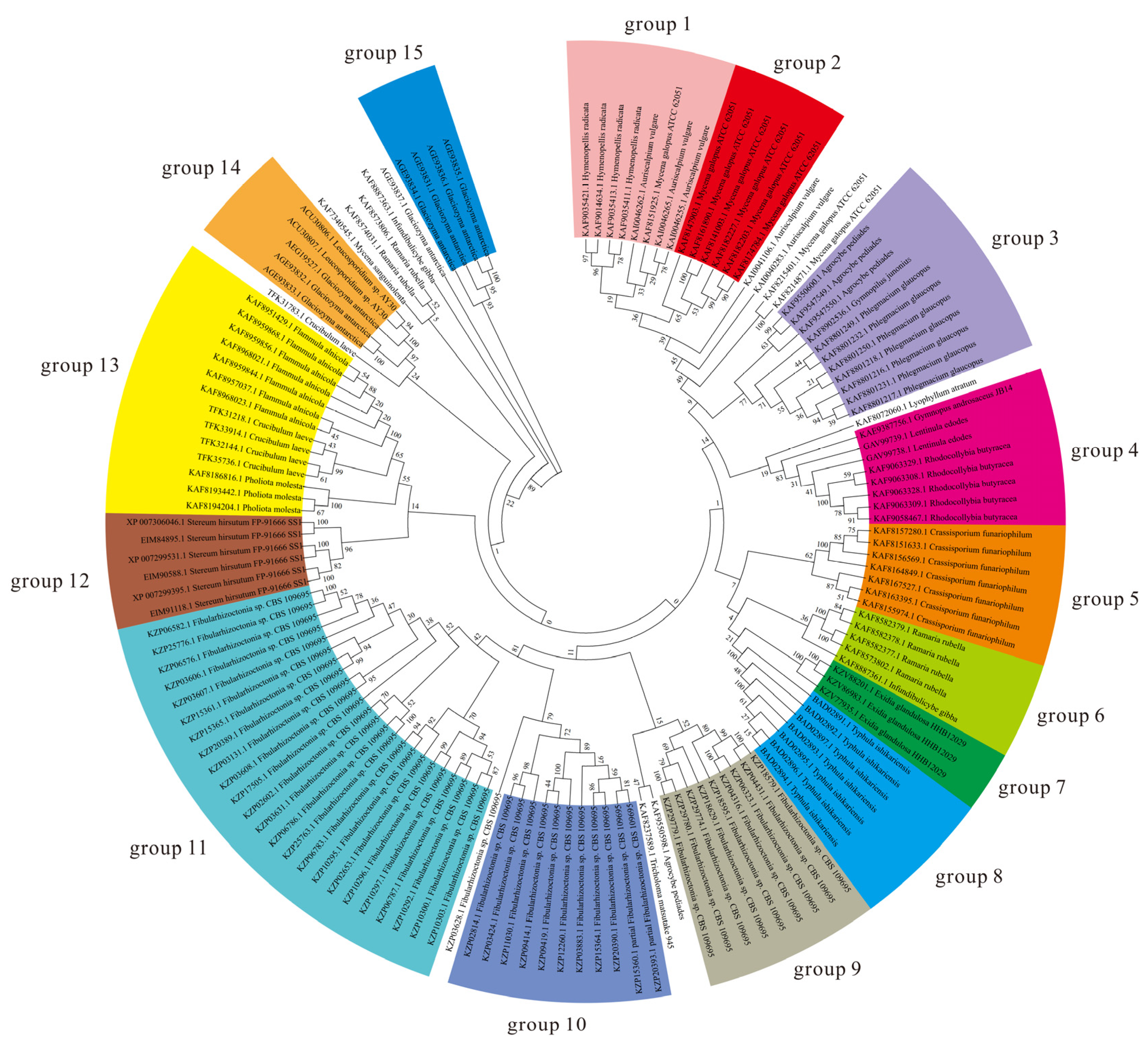
2.4. Phylogenomic Analysis
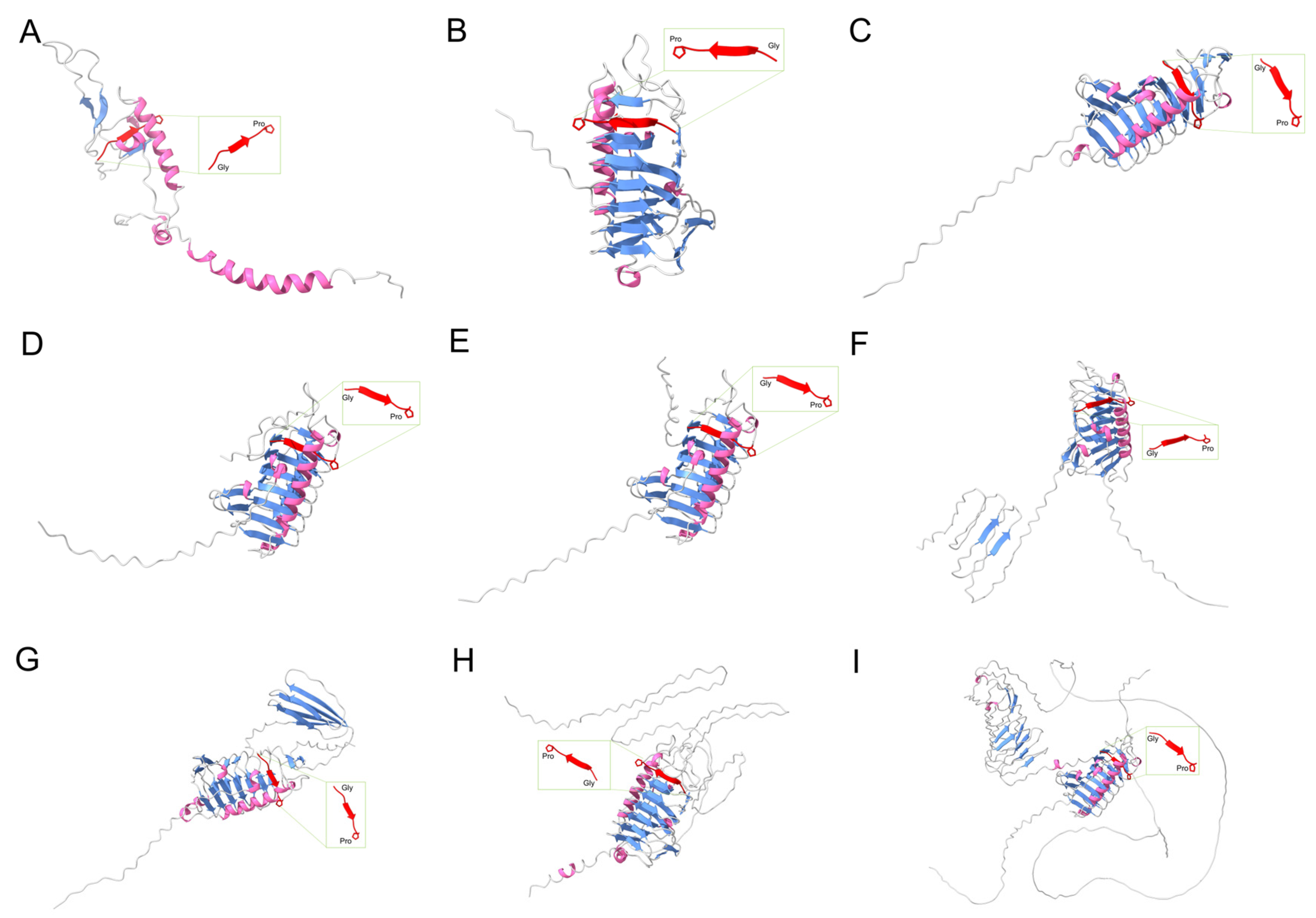
2.5. Modeling of AFPs and Potential Ice-Binding Site Predictions
3. Results
3.1. Genome-Wide Characterization of Psychrophilic Yeasts
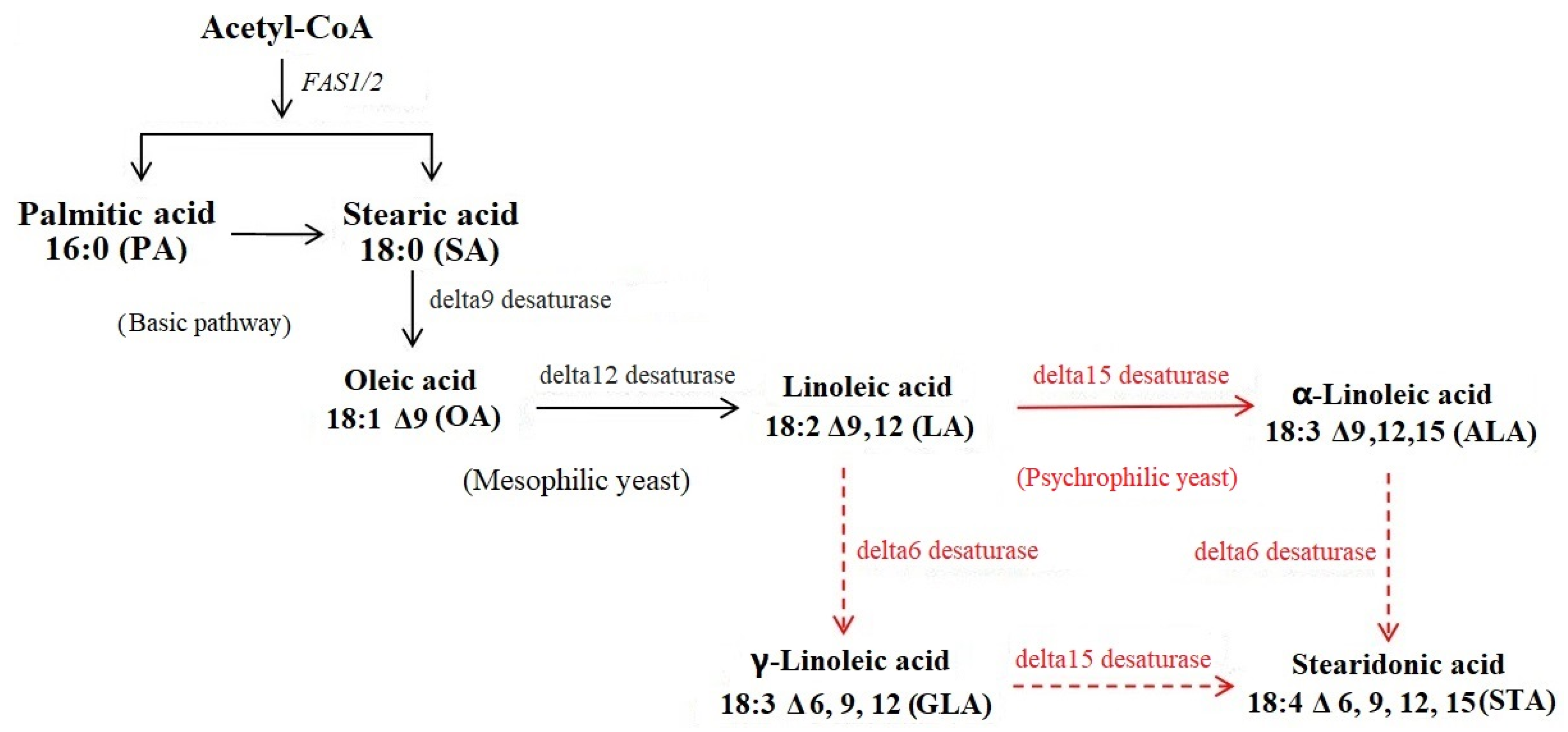
3.2. Unique Polyunsaturated Fatty Acid (PUFA) Pathways That Benefit Low-Temperature Adaptation
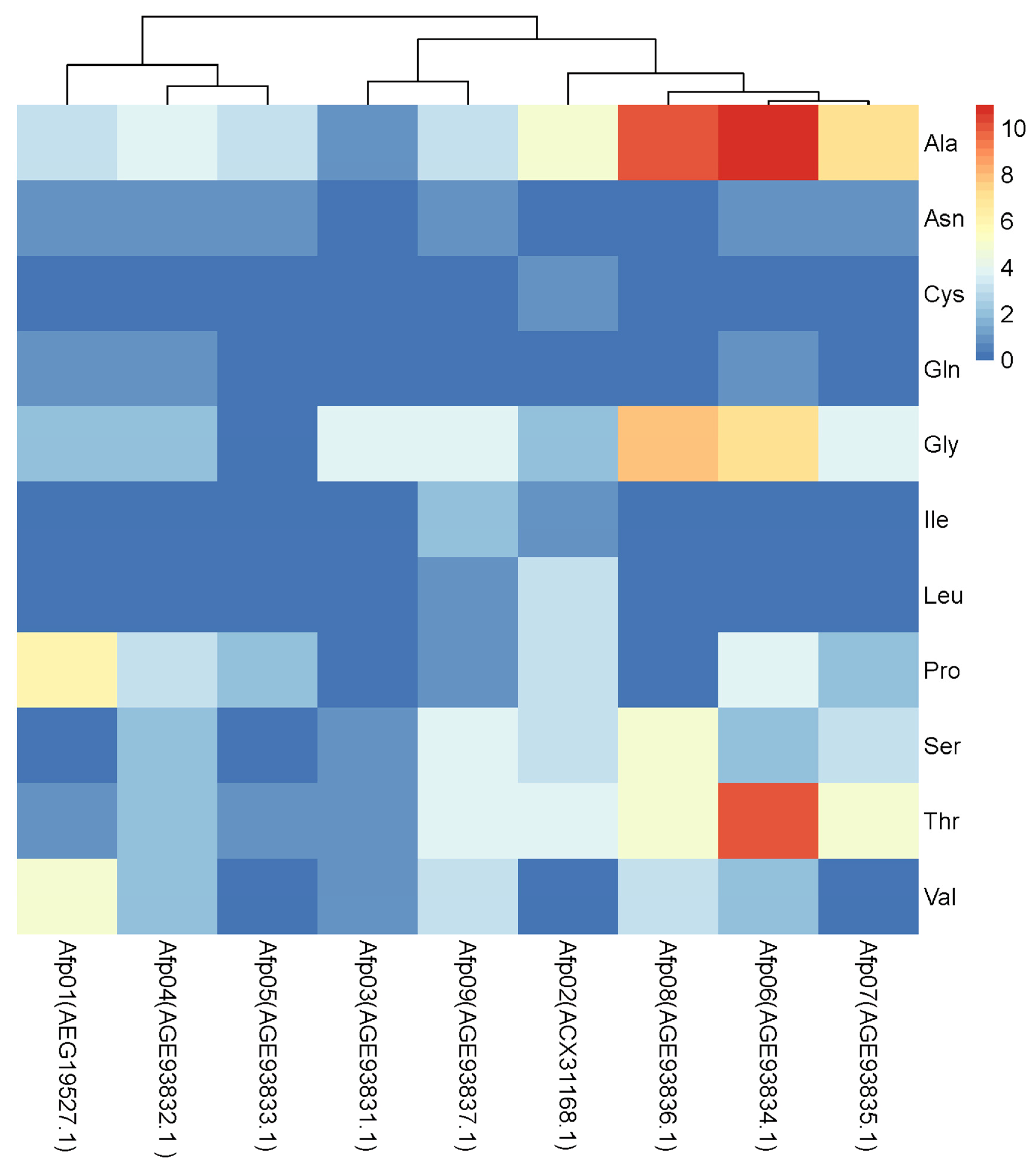
3.3. Various AFPs Contribute to Low-Temperature Protection
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Margesin, R.; Miteva, V. Diversity and ecology of psychrophilic microorganisms. Res. Microbiol. 2011, 162, 346–361. [Google Scholar] [CrossRef] [PubMed]
- Hassan, N.; Rafiq, M.; Hayat, M.; Ali Shah, A.; Hasan, F. Psychrophilic and psychrotrophic fungi: A comprehensive review. Rev. Environ. Sci. Bio. 2016, 15, 147–172. [Google Scholar] [CrossRef]
- De Maayer, P.; Anderson, D.; Cary, C.; Cowan, D.A. Some like it cold: Understanding the survival strategies of psychrophiles. EMBO Rep. 2014, 15, 508–517. [Google Scholar] [CrossRef]
- Cavicchioli, R.; Siddiqui, S.K.; Andrews, D.; Sowers, R.K. Low-temperature extremophiles and their applications. Curr. Opin. Biotech. 2002, 13, 253–261. [Google Scholar] [CrossRef] [PubMed]
- Buzzini, P.; Branda, E.; Goretti, M.; Turchetti, B. Psychrophilic yeasts from worldwide glacial habitats: Diversity, adaptation strategies and biotechnological potential. FEMS Microbiol. Ecol. 2012, 82, 217–241. [Google Scholar] [CrossRef]
- Péter, G.; Rosa, C. Biodiversity and Ecophysiology of Yeasts; Springer: Berlin/Heidelberg, Germany, 2006. [Google Scholar]
- Alias, N.; Ahmad Mazian, M.; Salleh, A.B.; Basri, M.; Rahman, R.N. Molecular Cloning and Optimization for High Level Expression of Cold-Adapted Serine Protease from Antarctic Yeast Glaciozyma antarctica PI12. Enzym. Res. 2014, 2014, 197938. [Google Scholar] [CrossRef] [Green Version]
- Grzymski, J.J.; Carter, B.J.; DeLong, E.F.; Feldman, R.A.; Ghadiri, A.; Murray, A.E. Comparative genomics of DNA fragments from six Antarctic marine planktonic bacteria. Appl. Environ. Microbiol. 2006, 72, 1532–1541. [Google Scholar] [CrossRef] [Green Version]
- Konstantinidis, K.T.; Braff, J.; Karl, D.M.; DeLong, E.F. Comparative metagenomic analysis of a microbial community residing at a depth of 4000 meters at station ALOHA in the North Pacific subtropical gyre. Appl. Environ. Microbiol. 2009, 75, 5345–5355. [Google Scholar] [CrossRef] [Green Version]
- Simon, C.; Wiezer, A.; Strittmatter, A.W.; Daniel, R. Phylogenetic diversity and metabolic potential revealed in a glacier ice metagenome. Appl. Environ. Microbiol. 2009, 75, 7519–7526. [Google Scholar] [CrossRef] [Green Version]
- Berlemont, R.; Delsaute, M.; Pipers, D.; D’Amico, S.; Feller, G.; Galleni, M.; Power, P. Insights into bacterial cellulose biosynthesis by functional metagenomics on Antarctic soil samples. ISME J. 2009, 3, 1070–1081. [Google Scholar] [CrossRef]
- Cieslinski, H.; Dlugolecka, A.; Kur, J.; Turkiewicz, M. An MTA phosphorylase gene discovered in the metagenomic library derived from Antarctic top soil during screening for lipolytic active clones confers strong pink fluorescence in the presence of rhodamine B. FEMS Microbiol. Lett. 2009, 299, 232–240. [Google Scholar] [PubMed] [Green Version]
- Heath, C.; Hu, X.P.; Cary, S.C.; Cowan, D. Identification of a novel alkaliphilic esterase active at low temperatures by screening a metagenomic library from antarctic desert soil. Appl. Environ. Microbiol. 2009, 75, 4657–4659. [Google Scholar] [CrossRef] [Green Version]
- Yusof, N.A.; Kamaruddin, S.; Bakar, F.D.A.; Mahadi, N.M.; Murad, A.M.A. Structural and functional insights into TRiC chaperonin from a psychrophilic yeast, Glaciozyma antarctica. Cell Stress Chaperon. 2019, 24, 351–368. [Google Scholar] [CrossRef] [PubMed]
- Firdaus-Raih, M.; Hashim, N.H.F.; Bharudin, I.; Abu Bakar, M.F.; Huang, K.K.; Alias, H.; Lee, B.K.B.; Mat Isa, M.N.; Mat-Sharani, S.; Sulaiman, S.; et al. The Glaciozyma antarctica genome reveals an array of systems that provide sustained responses towards temperature variations in a persistently cold habitat. PLoS ONE 2018, 13, e0189947. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goordial, J.; Raymond-Bouchard, I.; Riley, R.; Ronholm, J.; Shapiro, N.; Woyke, T.; LaButti, K.M.; Tice, H.; Amirebrahimi, M.; Grigoriev, I.V.; et al. Improved High-Quality Draft Genome Sequence of the Eurypsychrophile Rhodotorula sp. JG1b, Isolated from Permafrost in the Hyperarid Upper-Elevation McMurdo Dry Valleys, Antarctica. Genome Announc. 2016, 4, e00069-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Touchette, D.; Altshuler, I.; Gostincar, C.; Zalar, P.; Raymond-Bouchard, I.; Zajc, J.; McKay, C.P.; Gunde-Cimerman, N.; Whyte, L.G. Novel Antarctic yeast adapts to cold by switching energy metabolism and increasing small RNA synthesis. ISME J. 2022, 16, 221–232. [Google Scholar] [CrossRef]
- Nachtweide, S.; Stanke, M. Multi-Genome Annotation with AUGUSTUS. Methods Mol. Biol. 2019, 1962, 139–160. [Google Scholar]
- Aramaki, T.; Blanc-Mathieu, R.; Endo, H.; Ohkubo, K.; Kanehisa, M.; Goto, S.; Ogata, H. KofamKOALA: KEGG Ortholog assignment based on profile HMM and adaptive score threshold. Bioinformatics 2020, 36, 2251–2252. [Google Scholar] [CrossRef] [Green Version]
- Moriya, Y.; Itoh, M.; Okuda, S.; Yoshizawa, A.C.; Kanehisa, M. KAAS: An automatic genome annotation and pathway reconstruction server. Nucleic Acids Res. 2007, 35, W182–W185. [Google Scholar] [CrossRef] [Green Version]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Zidek, A.; Potapenko, A.; et al. Highly accurate protein structure prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef]
- Doxey, A.C.; McConkey, B.J. AFPredictor: A computational screening protocol for antifreeze/ice-structuring proteins. Nat. Protoc. Exch. 2006. [Google Scholar] [CrossRef]
- Mo, Y. Probing the nature of hydrogen bonds in DNA base pairs. J. Mol. Model 2006, 12, 665–672. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; Zhou, Z.; Zhang, L. An overview of fungal glycan-based therapeutics. Prog. Mol. Biol. Transl. Sci. 2019, 163, 135–163. [Google Scholar] [PubMed]
- Lee, J.M.; Lee, H.; Kang, S.; Park, W.J. Fatty Acid Desaturases, Polyunsaturated Fatty Acid Regulation, and Biotechnological Advances. Nutrients 2016, 8, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McDonough, V.M.; Roth, T.M. Growth temperature affects accumulation of exogenous fatty acids and fatty acid composition in Schizosaccharomyces pombe. Antonie Van Leeuwenhoek 2004, 86, 349–354. [Google Scholar] [CrossRef]
- Martin, C.E.; Oh, C.S.; Jiang, Y. Regulation of long chain unsaturated fatty acid synthesis in yeast. Biochim. Biophys. Acta 2007, 1771, 271–285. [Google Scholar] [CrossRef]
- Rossi, M.; Buzzini, P.; Cordisco, L.; Amaretti, A.; Sala, M.; Raimondi, S.; Ponzoni, C.; Pagnoni, U.M.; Matteuzzi, D. Growth, lipid accumulation, and fatty acid composition in obligate psychrophilic, facultative psychrophilic, and mesophilic yeasts. FEMS Microbiol. Ecol. 2009, 69, 363–372. [Google Scholar] [CrossRef] [Green Version]
- Cheung, R.C.F.; Ng, T.B.; Wong, J.K. Antifreeze Proteins from Diverse Organisms and their Applications: An Overview. Curr. Protein Pept. Sci. 2017, 18, 262–283. [Google Scholar] [CrossRef]
- Sicheri, F.; Yang, D.S. Ice-binding structure and mechanism of an antifreeze protein from winter flounder. Nature 1995, 375, 427–431. [Google Scholar] [CrossRef] [Green Version]
- María, P.; Carboni-Oerlemans, C.; Tuin, B.; Bargeman, G.; Van Der Meer, A.; Van Gemert, R. Biotechnological applications of Candida antarctica lipase A: State-of-the-art. J. Mol. Catal. B Enzym. 2005, 37, 36–46. [Google Scholar] [CrossRef]
- Tronelli, D.; Maugini, E.; Bossa, F.; Pascarella, S. Structural adaptation to low temperatures--analysis of the subunit interface of oligomeric psychrophilic enzymes. FEBS J. 2007, 274, 4595–4608. [Google Scholar] [CrossRef] [PubMed]
- Gerday, C.; Aittaleb, M.; Bentahir, M.; Chessa, J.P.; Claverie, P.; Collins, T.; D’Amico, S.; Dumont, J.; Garsoux, G.; Georlette, D.; et al. Cold-adapted enzymes: From fundamentals to biotechnology. Trends Biotechnol. 2000, 18, 103–107. [Google Scholar] [CrossRef] [PubMed]
- Vance, T.D.R.; Bayer-Giraldi, M.; Davies, P.L.; Mangiagalli, M. Ice-binding proteins and the ’domain of unknown function’ 3494 family. FEBS J. 2019, 286, 855–873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munoz, P.A.; Marquez, S.L.; Gonzalez-Nilo, F.D.; Marquez-Miranda, V.; Blamey, J.M. Structure and application of antifreeze proteins from Antarctic bacteria. Microb. Cell Fact. 2017, 16, 138. [Google Scholar] [CrossRef] [Green Version]
- Xiang, H.; Yang, X.; Ke, L.; Hu, Y. The properties, biotechnologies, and applications of antifreeze proteins. Int. J. Biol. Macromol. 2020, 153, 661–675. [Google Scholar] [CrossRef]
- Zhao, X.; Ma, Z.; Yin, M. Using Support Vector Machine and Evolutionary Profiles to Predict Antifreeze Protein Sequences. Int. J. Mol. Med. 2012, 13, 2196–2207. [Google Scholar] [CrossRef]
- Kandaswamy, K.K.; Chou, K.C.; Martinetz, T.; Moller, S.; Suganthan, P.N.; Sridharan, S.; Pugalenthi, G. AFP-Pred: A random forest approach for predicting antifreeze proteins from sequence-derived properties. J. Theor. Biol. 2011, 270, 56–62. [Google Scholar] [CrossRef]
- Davies, P.L.; Baardsnes, J.; Kuiper, M.J.; Walker, V.K. Structure and function of antifreeze proteins. Philos Trans. R. Soc. Lond. B Biol. Sci. 2002, 357, 927–935. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains | Isolations | Accession No. | Sequence Length (Mb) | GC Content (%) | BGCs | CAEs | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| Terpene | Nrps | Nrps-like | Lipase | Glucoamylase | α-Amylase | |||||
| L. scottii | Southern Ocean (sea water) | GCA_003054985.1 | 26.75 | 0.59 | 1 | 2 | 2 | 1 | 0 | 1 |
| M. psychrophila | China: Hailuogou, Sichuan (alpine glacier soil) | GCA_001889225.1 | 27.77 | 0.54 | 1 | 0 | 1 | 0 | 0 | 1 |
| M. frigida | Not applicable | GCA_001600395.1 | 28.62 | 0.54 | 1 | 0 | 2 | 0 | 0 | 1 |
| M. blollopis | Antarctica: East Antarctica, Skarvsnes ice-free area | GCA_000950635.1 | 30.48 | 0.54 | 1 | 0 | 2 | 0 | 0 | 1 |
| G. antarctica | Sea ice near Casey Research Station, Antarctica | GCA_002917775.1 | 20.03 | 0.60 | 2 | 1 | 1 | 0 | 0 | 1 |
| R. frigidialcoholis | Missing | GCA_001541205.1 | 19.39 | 0.61 | 2 | 1 | 1 | 1 | 0 | 1 |
| C. neoformans * | Not applicable | GCF_000091045.1 | 19.05 | 0.49 | 2 | 0 | 2 | 1 | 1 | 1 |
| M. australis | Antarctica: Admiralty Bay, King George island | GCA_002073855.1 | 14.35 | 0.47 | 1 | 0 | 1 | 1 | 1 | 0 |
| C. psychrophila | Antarctica | GCA_900186205.1 | 11.24 | 0.37 | 0 | 0 | 1 | 1 | 1 | 0 |
| M. caribbica | Reclamation soil | GCA_000755205.1 | 10.61 | 0.44 | 1 | 0 | 1 | 1 | 1 | 0 |
| S. cerevisiae * | Not applicable | GCF_000146045.2 | 12.16 | 0.38 | 1 | 0 | 0 | 1 | 1 | 0 |
| S. pombe * | Not applicable | GCF_000002945.1 | 12.60 | 0.36 | 0 | 0 | 1 | 1 | 1 | 1 |
| Strains | Number of PUFAs | |||
|---|---|---|---|---|
| Delta 6 Desaturase | Delta 9 Desaturase | Delta 12 Desaturase | Delta 15 Desaturase | |
| L. scottii | 1 | 1 | 1 | 0 |
| M. psychrophila | 0 | 1 | 1 | 1 |
| M. frigida | 0 | 1 | 1 | 1 |
| M. blollopis | 0 | 1 | 1 | 1 |
| G. antarctica | 1 | 1 | 1 | 1 |
| R. frigidialcoholis | 1 | 1 | 1 | 1 |
| C. neoformans * | 0 | 1 | 1 | 0 |
| M. australis | 1 | 1 | 1 | 1 |
| C. psychrophila | 1 | 1 | 1 | 1 |
| M. caribbica | 1 | 1 | 1 | 1 |
| S. cerevisiae * | 0 | 1 | 0 | 0 |
| S. pombe * | 0 | 1 | 0 | 0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, H.; Zheng, G.; Chen, Z.; Ding, X.; Wu, J.; Zhang, H.; Jia, S. Psychrophilic Yeasts: Insights into Their Adaptability to Extremely Cold Environments. Genes 2023, 14, 158. https://doi.org/10.3390/genes14010158
Liu H, Zheng G, Chen Z, Ding X, Wu J, Zhang H, Jia S. Psychrophilic Yeasts: Insights into Their Adaptability to Extremely Cold Environments. Genes. 2023; 14(1):158. https://doi.org/10.3390/genes14010158
Chicago/Turabian StyleLiu, Haisheng, Guiliang Zheng, Zhongwei Chen, Xiaoya Ding, Jinran Wu, Haili Zhang, and Shulei Jia. 2023. "Psychrophilic Yeasts: Insights into Their Adaptability to Extremely Cold Environments" Genes 14, no. 1: 158. https://doi.org/10.3390/genes14010158