
Steatosis and Metabolic Disorders Associated with Synergistic Activation of the CAR/RXR Heterodimer by Pesticides

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Luciferase Assay
2.3. Animal Experiment
2.4. Gene Expression Studies
2.5. Liver Neutral Lipid Analysis
2.6. Histology
2.7. Plasma Analysis
2.8. Determination of Combined Effects and Statistical Analysis
3. Results
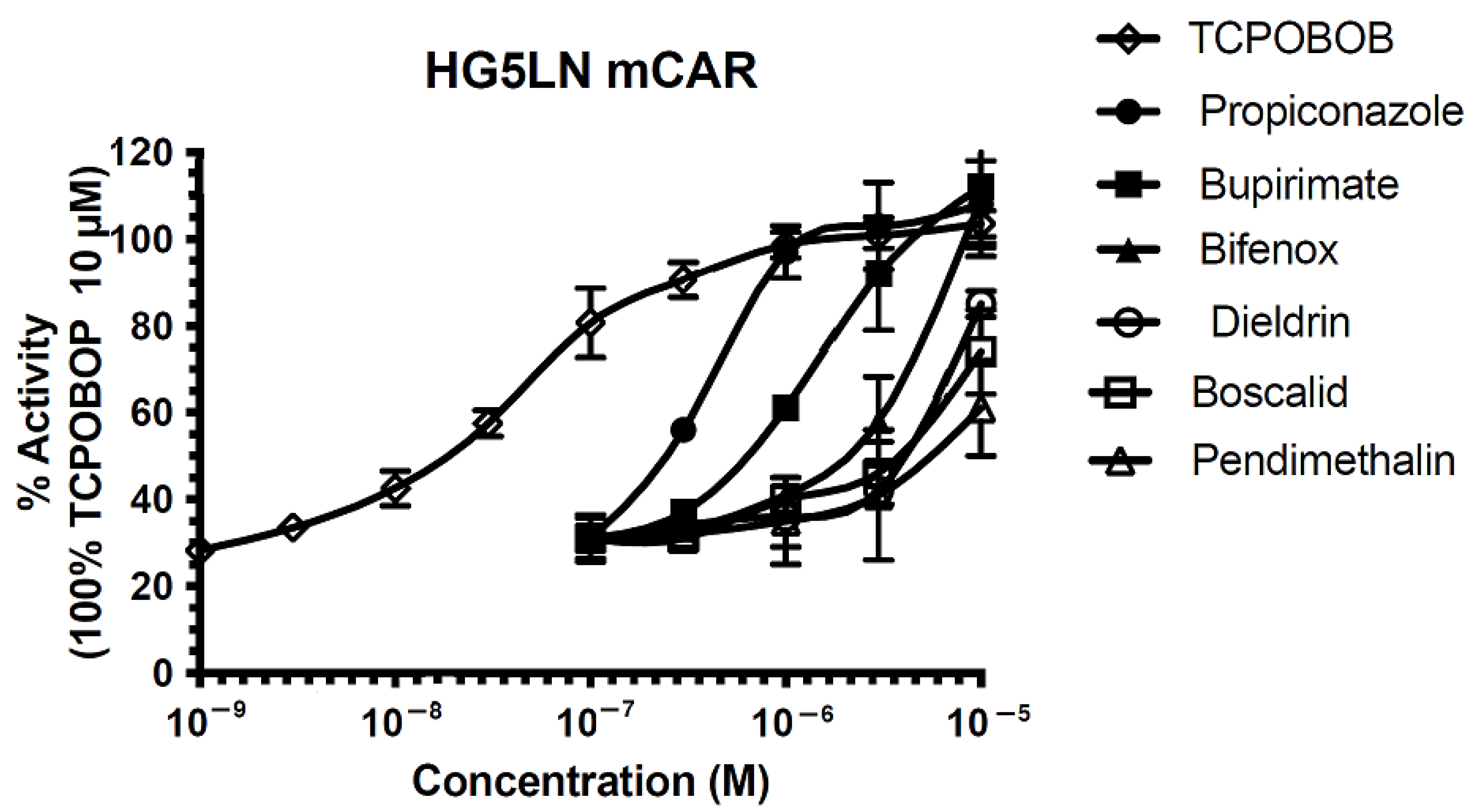
3.1. Modulation of Transcriptional Activity of mCAR by Pesticides
3.2. Synergic and Combined Effects on CAR Activation
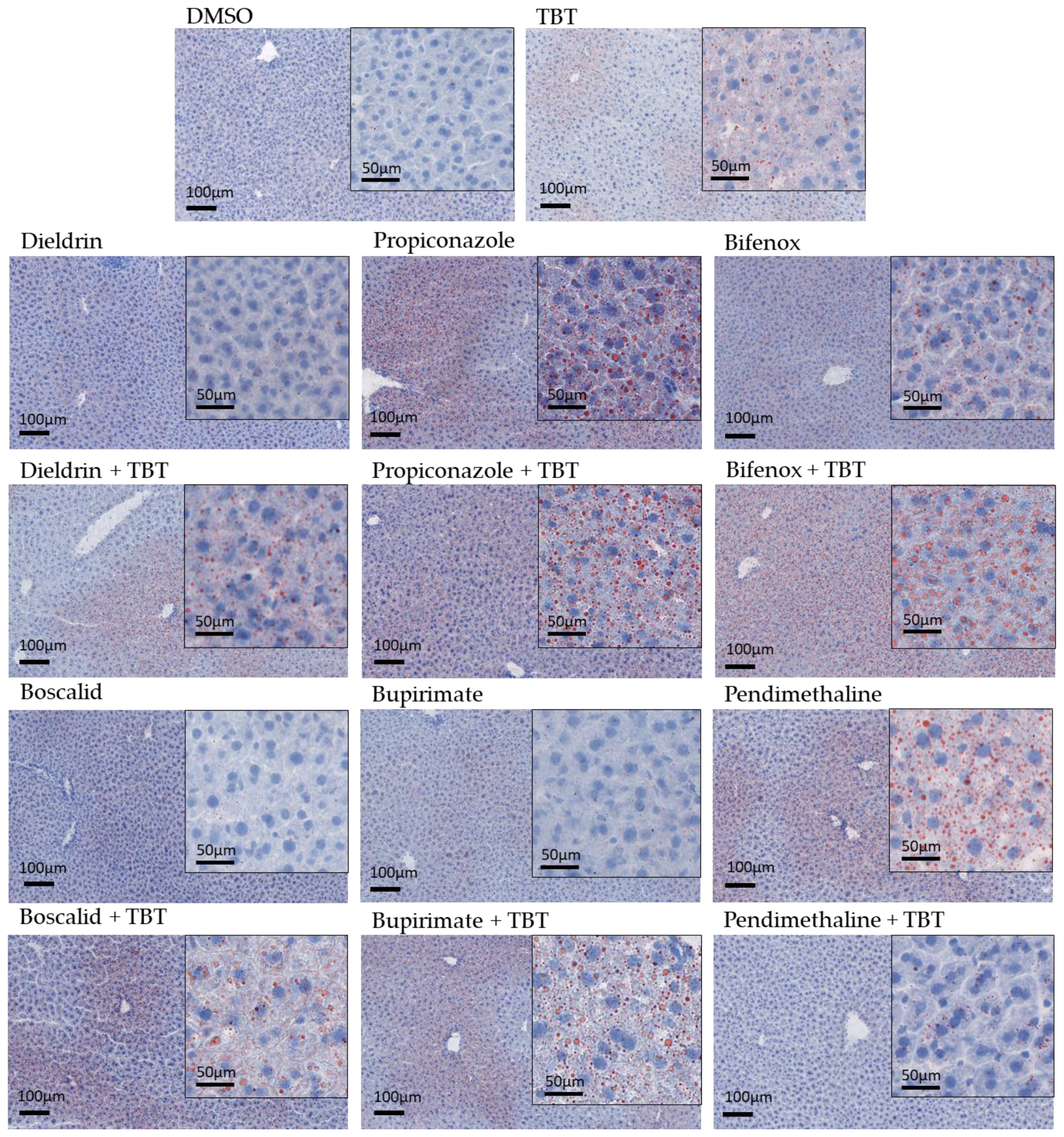
3.3. NAFLD Induction
3.4. Changes in Plasmatic Biochemical Profiles
3.5. Pesticides Elicit Transcriptional Changes in Key Genes of the Liver’s Metabolic Pathways

4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Younossi, Z.M.; Koenig, A.B.; Abdelatif, D.; Fazel, Y.; Henry, L.; Wymer, M. Global Epidemiology of Nonalcoholic Fatty Liver Disease—Meta-Analytic Assessment of Prevalence, Incidence, and Outcomes. Hepatology 2016, 64, 73–84. [Google Scholar] [CrossRef] [PubMed]
- Kabbany, M.N.; Selvakumar, P.K.C.; Watt, K.; Lopez, R.; Akras, Z.; Zein, N.; Carey, W.; Alkhouri, N. Prevalence of Nonalcoholic Steatohepatitis-Associated Cirrhosis in the United States: An Analysis of National Health and Nutrition Examination Survey Data. Off. J. Am. Coll. Gastroenterol 2017, 112, 581–587. [Google Scholar] [CrossRef] [PubMed]
- Salt, W.B. Nonalcoholic Fatty Liver Disease (NAFLD): A Comprehensive Review. J. Insur. Med.—N. Y. Denver 2004, 36, 27–41. [Google Scholar]
- Nakamura, A.; Terauchi, Y. Lessons from Mouse Models of High-Fat Diet-Induced NAFLD. Int. J. Mol. Sci. 2013, 14, 21240–21257. [Google Scholar] [CrossRef]
- Kristiansen, M.N.B.; Veidal, S.S.; Christoffersen, C.; Jelsing, J.; Rigbolt, K.T.G. Molecular Characterization of Microvesicular and Macrovesicular Steatosis Shows Widespread Differences in Metabolic Pathways. Lipids 2019, 54, 109–115. [Google Scholar] [CrossRef] [PubMed]
- Fromenty, B.; Berson, A.; Pessayre, D. Microvesicular Steatosis and Steatohepatitis: Role of Mitochondrial Dysfunction and Lipid Peroxidation. J. Hepatol. 1997, 26, 13–22. [Google Scholar] [CrossRef] [PubMed]
- Smith, B.W.; Adams, L.A. Non-Alcoholic Fatty Liver Disease. Crit. Rev. Clin. Lab. Sci. 2011, 48, 97–113. [Google Scholar] [CrossRef] [PubMed]
- Aller, R.; Izaola, O.; Ruiz-Rebollo, L.; Pacheco, D.; de Luis, D.A. Predictive Factors of Non-Alcoholic Steatohepatitis: Relationship with Metabolic Syndrome. Nutr. Hosp. 2015, 31, 2496–2502. [Google Scholar]
- Liebe, R.; Esposito, I.; Bock, H.H.; vom Dahl, S.; Stindt, J.; Baumann, U.; Luedde, T.; Keitel, V. Diagnosis and Management of Secondary Causes of Steatohepatitis. J. Hepatol. 2021, 74, 1455–1471. [Google Scholar] [CrossRef]
- Wahlang, B.; Beier, J.I.; Clair, H.B.; Bellis-Jones, H.J.; Falkner, K.C.; McClain, C.J.; Cave, M.C. Toxicant-Associated Steatohepatitis. Toxicol. Pathol. 2013, 41, 343–360. [Google Scholar] [CrossRef]
- Arciello, M.; Gori, M.; Maggio, R.; Barbaro, B.; Tarocchi, M.; Galli, A.; Balsano, C. Environmental Pollution: A Tangible Risk for NAFLD Pathogenesis. Int. J. Mol. Sci. 2013, 14, 22052–22066. [Google Scholar] [CrossRef] [PubMed]
- Mulligan, C.; Kondakala, S.; Yang, E.-J.; Stokes, J.V.; Stewart, J.A.; Kaplan, B.L.F.; Howell, G.E. Exposure to an Environmentally Relevant Mixture of Organochlorine Compounds and Polychlorinated Biphenyls Promotes Hepatic Steatosis in Male Ob/Ob Mice. Environ. Toxicol. 2017, 32, 1399–1411. [Google Scholar] [CrossRef] [PubMed]
- Al-Eryani, L.; Wahlang, B.; Falkner, K.C.; Guardiola, J.J.; Clair, H.B.; Prough, R.A.; Cave, M. Identification of Environmental Chemicals Associated with the Development of Toxicant-Associated Fatty Liver Disease in Rodents. Toxicol. Pathol. 2015, 43, 482–497. [Google Scholar] [CrossRef] [PubMed]
- Pirozzi, A.V.A.; Stellavato, A.; La Gatta, A.; Lamberti, M.; Schiraldi, C. Mancozeb, a Fungicide Routinely Used in Agriculture, Worsens Nonalcoholic Fatty Liver Disease in the Human HepG2 Cell Model. Toxicol. Lett. 2016, 249, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Cave, M.C.; Clair, H.B.; Hardesty, J.E.; Falkner, K.C.; Feng, W.; Clark, B.J.; Sidey, J.; Shi, H.; Aqel, B.A.; McClain, C.J.; et al. Nuclear Receptors and Nonalcoholic Fatty Liver Disease. Biochim. Biophys. Acta 2016, 1859, 1083–1099. [Google Scholar] [CrossRef]
- Ballestri, S.; Nascimbeni, F.; Romagnoli, D.; Baldelli, E.; Lonardo, A. The Role of Nuclear Receptors in the Pathophysiology, Natural Course, and Drug Treatment of NAFLD in Humans. Adv. Ther. 2016, 33, 291–319. [Google Scholar] [CrossRef]
- Wahlang, B.; Jin, J.; Beier, J.I.; Hardesty, J.E.; Daly, E.F.; Schnegelberger, R.D.; Falkner, K.C.; Prough, R.A.; Kirpich, I.A.; Cave, M.C. Mechanisms of Environmental Contributions to Fatty Liver Disease. Curr. Environ. Health Rep. 2019, 6, 80–94. [Google Scholar] [CrossRef]
- Mo, L.; He, J. Nuclear Hormone Receptors PXR and CAR and Metabolic Diseases. Horm. Mol. Biol. Clin. Investig. 2014, 19, 129–140. [Google Scholar] [CrossRef]
- Wang, Y.-M.; Ong, S.S.; Chai, S.C.; Chen, T. Role of CAR and PXR in Xenobiotic Sensing and Metabolism. Expert. Opin. Drug. Metab. Toxicol. 2012, 8, 803–817. [Google Scholar] [CrossRef]
- Banerjee, M.; Robbins, D.; Chen, T. Targeting Xenobiotic Receptors PXR and CAR in Human Diseases. Drug. Discov. Today 2015, 20, 618–628. [Google Scholar] [CrossRef]
- Moreau, A.; Vilarem, M.J.; Maurel, P.; Pascussi, J.M. Xenoreceptors CAR and PXR Activation and Consequences on Lipid Metabolism, Glucose Homeostasis, and Inflammatory Response. Mol. Pharm. 2008, 5, 35–41. [Google Scholar] [CrossRef]
- Marmugi, A.; Lukowicz, C.; Lasserre, F.; Montagner, A.; Polizzi, A.; Ducheix, S.; Goron, A.; Gamet-Payrastre, L.; Gerbal-Chaloin, S.; Pascussi, J.M.; et al. Activation of the Constitutive Androstane Receptor Induces Hepatic Lipogenesis and Regulates Pnpla3 Gene Expression in a LXR-Independent Way. Toxicol. Appl. Pharmacol. 2016, 303, 90–100. [Google Scholar] [CrossRef]
- Knebel, C.; Buhrke, T.; Süssmuth, R.; Lampen, A.; Marx-Stoelting, P.; Braeuning, A. Pregnane X Receptor Mediates Steatotic Effects of Propiconazole and Tebuconazole in Human Liver Cell Lines. Arch. Toxicol. 2019, 93, 1311–1322. [Google Scholar] [CrossRef]
- Wada, T.; Gao, J.; Xie, W. PXR and CAR in Energy Metabolism. Trends Endocrinol. Metab. 2009, 20, 273–279. [Google Scholar] [CrossRef] [PubMed]
- Delfosse, V.; Huet, T.; Harrus, D.; Granell, M.; Bourguet, M.; Gardia-Parège, C.; Chiavarina, B.; Grimaldi, M.; Le Mével, S.; Blanc, P.; et al. Mechanistic Insights into the Synergistic Activation of the RXR-PXR Heterodimer by Endocrine Disruptor Mixtures. Proc. Natl. Acad. Sci. USA 2021, 118, e2020551118. [Google Scholar] [CrossRef] [PubMed]
- Delfosse, V.; Dendele, B.; Huet, T.; Grimaldi, M.; Boulahtouf, A.; Gerbal-Chaloin, S.; Beucher, B.; Roecklin, D.; Muller, C.; Rahmani, R.; et al. Synergistic Activation of Human Pregnane X Receptor by Binary Cocktails of Pharmaceutical and Environmental Compounds. Nat. Commun. 2015, 6, 8089. [Google Scholar] [CrossRef]
- Kanayama, T.; Kobayashi, N.; Mamiya, S.; Nakanishi, T.; Nishikawa, J. Organotin Compounds Promote Adipocyte Differentiation as Agonists of the Peroxisome Proliferator-Activated Receptor γ/Retinoid X Receptor Pathway. Mol. Pharmacol. 2005, 67, 766–774. [Google Scholar] [CrossRef] [PubMed]
- Antizar-Ladislao, B. Environmental Levels, Toxicity and Human Exposure to Tributyltin (TBT)-Contaminated Marine Environment. A Review. Environ. Int. 2008, 34, 292–308. [Google Scholar] [CrossRef]
- Nakanishi, T. Endocrine Disruption Induced by Organotin Compounds; Organotins Function as a Powerful Agonist for Nuclear Receptors Rather than an Aromatase Inhibitor. J. Toxicol. Sci. 2008, 33, 269–276. [Google Scholar] [CrossRef]
- Le Maire, A.; Grimaldi, M.; Roecklin, D.; Dagnino, S.; Vivat-Hannah, V.; Balaguer, P.; Bourguet, W. Activation of RXR-PPAR Heterodimers by Organotin Environmental Endocrine Disruptors. EMBO Rep. 2009, 10, 367–373. [Google Scholar] [CrossRef]
- Seimandi, M.; Lemaire, G.; Pillon, A.; Perrin, A.; Carlavan, I.; Voegel, J.J.; Vignon, F.; Nicolas, J.-C.; Balaguer, P. Differential Responses of PPARalpha, PPARdelta, and PPARgamma Reporter Cell Lines to Selective PPAR Synthetic Ligands. Anal. Biochem. 2005, 344, 8–15. [Google Scholar] [CrossRef]
- Chomczynski, P.; Sacchi, N. Single-Step Method of RNA Isolation by Acid Guanidinium Thiocyanate-Phenol-Chloroform Extraction. Anal. Biochem. 1987, 162, 156–159. [Google Scholar] [CrossRef]
- Lukowicz, C.; Ellero-Simatos, S.; Régnier, M.; Polizzi, A.; Lasserre, F.; Montagner, A.; Lippi, Y.; Jamin, E.L.; Martin, J.-F.; Naylies, C.; et al. Metabolic Effects of a Chronic Dietary Exposure to a Low-Dose Pesticide Cocktail in Mice: Sexual Dimorphism and Role of the Constitutive Androstane Receptor. Environ. Health Perspect. 2018, 126, 067007. [Google Scholar] [CrossRef] [PubMed]
- Kleiner, D.E.; Brunt, E.M.; Natta, M.V.; Behling, C.; Contos, M.J.; Cummings, O.W.; Ferrell, L.D.; Liu, Y.-C.; Torbenson, M.S.; Unalp-Arida, A.; et al. Design and Validation of a Histological Scoring System for Nonalcoholic Fatty Liver Disease. Hepatology 2005, 41, 1313–1321. [Google Scholar] [CrossRef] [PubMed]
- Foucquier, J.; Guedj, M. Analysis of Drug Combinations: Current Methodological Landscape. Pharmacol. Res. Perspect. 2015, 3, e00149. [Google Scholar] [CrossRef] [PubMed]
- Timsit, Y.E.; Negishi, M. CAR and PXR: The Xenobiotic-Sensing Receptors. Steroids 2007, 72, 231–246. [Google Scholar] [CrossRef]
- Kojetin, D.J.; Matta-Camacho, E.; Hughes, T.S.; Srinivasan, S.; Nwachukwu, J.C.; Cavett, V.; Nowak, J.; Chalmers, M.J.; Marciano, D.P.; Kamenecka, T.M.; et al. Structural Mechanism for Signal Transduction in RXR Nuclear Receptor Heterodimers. Nat. Commun. 2015, 6, 8013. [Google Scholar] [CrossRef] [PubMed]
- Aranda, A.; Pascual, A. Nuclear Hormone Receptors and Gene Expression. Physiol. Rev. 2001, 81, 1269–1304. [Google Scholar] [CrossRef]
- Dawson, M.I.; Xia, Z. The Retinoid X Receptors and Their Ligands. Biochim. Biophys. Acta 2012, 1821, 21–56. [Google Scholar] [CrossRef]
- Cherian, M.T.; Chai, S.C.; Chen, T. Small-Molecule Modulators of the Constitutive Androstane Receptor. Expert. Opin. Drug. Metab. Toxicol. 2015, 11, 1099–1114. [Google Scholar] [CrossRef]
- Xu, R.X.; Lambert, M.H.; Wisely, B.B.; Warren, E.N.; Weinert, E.E.; Waitt, G.M.; Williams, J.D.; Collins, J.L.; Moore, L.B.; Willson, T.M.; et al. A Structural Basis for Constitutive Activity in the Human CAR/RXRalpha Heterodimer. Mol. Cell. 2004, 16, 919–928. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Chen, B.; Lu, J.; Xie, W. Deciphering the Roles of the Constitutive Androstane Receptor in Energy Metabolism. Acta Pharmacol. Sin. 2015, 36, 62–70. [Google Scholar] [CrossRef] [PubMed]
- Ipsen, D.H.; Lykkesfeldt, J.; Tveden-Nyborg, P. Molecular Mechanisms of Hepatic Lipid Accumulation in Non-Alcoholic Fatty Liver Disease. Cell. Mol. Life Sci. 2018, 75, 3313–3327. [Google Scholar] [CrossRef]
- Rives, C.; Fougerat, A.; Ellero-Simatos, S.; Loiseau, N.; Guillou, H.; Gamet-Payrastre, L.; Wahli, W. Oxidative Stress in NAFLD: Role of Nutrients and Food Contaminants. Biomolecules 2020, 10, 1702. [Google Scholar] [CrossRef]
- Shulman, A.I.; Mangelsdorf, D.J. Retinoid X Receptor Heterodimers in the Metabolic Syndrome. N. Engl. J. Med. 2005, 353, 604–615. [Google Scholar] [CrossRef]
- Yamazaki, Y.; Kakizaki, S.; Horiguchi, N.; Sohara, N.; Sato, K.; Takagi, H.; Mori, M.; Negishi, M. The Role of the Nuclear Receptor Constitutive Androstane Receptor in the Pathogenesis of Non-alcoholic Steatohepatitis. Gut 2007, 56, 565–574. [Google Scholar] [CrossRef] [PubMed]
- Jackson, E.; Shoemaker, R.; Larian, N.; Cassis, L. Adipose Tissue as a Site of Toxin Accumulation. Compr. Physiol. 2017, 7, 1085–1135. [Google Scholar] [CrossRef]
- Zhang, S.; Peng, X.; Yang, S.; Li, X.; Huang, M.; Wei, S.; Liu, J.; He, G.; Zheng, H.; Yang, L.; et al. The Regulation, Function, and Role of Lipophagy, a Form of Selective Autophagy, in Metabolic Disorders. Cell Death Dis. 2022, 13, 132. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequence F 5’-3’ | Primer Sequence R 3’-5’ |
|---|---|---|
| Acox1 | AGACCCTGAAGAAATCATGTGG | AGGAACATGCCCAAGTGAAG |
| Cd36 | GTTAAACAAAGAGGTCCTTACACATACAG | AGTGAAGGCTCAAAGATGGC |
| Cyp2b10 | TTTCTGCCCTTCTCAACAGGAA | TGGACGTGAAGAAAAGGAACAAC |
| Eci | GTTCACCATCAGCCTGGAGAAG | AGAAGATACCCGGGCATTCC |
| Fasn | AGTCAGCTATGAAGCAATTGTGGA | CACCCAGACGCCAGTGTTC |
| Gck | TCGCAGGTGGAGAGCGA | TCGCAGTCGGCGACAGA |
| Mvd | CGGTCAACATCGCAGTTATCAA | GTGCAGCGTGACGCTCAG |
| Plin3 | GGCTGGACAGACTGCAGGA | TCTTGAGCCCCAGACACTGTAG |
| Scd1 | CAGTGCCGCGCATCTCTAT | CAGCGGTACTCACTGGCAGA |
| Tbp | ACTTCGTGCAAGAAATGCTGAA | GCAGTTGTCCGTGGCTCTCT |
| Treatment | Liver/Body Weight |
|---|---|
| TBT | 1.33 ± 0.06 a |
| Dieldrin | 1.06 ± 0.05 |
| Dieldrin + TBT | 1.17 ± 0.10 a |
| Propiconazole | 1.23 ± 0.05 a |
| Propiconazole + TBT | 1.47 ± 0.09 a |
| Bifenox | 1.19 ± 0.05 a |
| Bifenox + TBT | 1.46 ± 0.08 abc |
| Boscalid | 1.11 ± 0.12 |
| Boscalid + TBT | 1.25 ± 0.15 a |
| Bupirimate | 1.04 ± 0.15 |
| Bupirimate + TBT | 1.34 ± 0.09 a |
| Pendimethalin | 1.29 ± 0.09 a |
| Pendimethalin + TBT | 1.10 ± 0.09 |
| Treatment | Steatosis Degree | Location | Microvesicular Morphology | |||
|---|---|---|---|---|---|---|
| <5% | Stage 1 | Mean Area % | Zone 3 | Azonal | ||
| DMSO | 6 | 0 | 0.50 ± 0.43 | 0 | 0 | 0 |
| TBT | 5 | 1 | 1.95 ± 2.19 | 1 | 0 | 1 |
| Dieldrin | 3 | 3 | 4.11 ± 2.62 | 3 | 0 | 3 |
| Dieldrin + TBT | 5 | 1 | 5.27 ± 7.59 | 1 | 0 | 1 |
| Propiconazole | 2 | 4 | 6.51 ± 2.51 | 3 | 1 | 4 |
| Propiconazole + TBT | 2 | 3 | 12.81 ± 9.79 a | 0 | 3 | 3 |
| Bifenox | 3 | 3 | 5.91 ± 3.56 | 2 | 1 | 3 |
| Bifenox + TBT | 3 | 3 | 6.49 ± 5.99 a | 2 | 1 | 3 |
| Boscalid | 3 | 0 | 2.08 ± 1.58 | 0 | 1 | 1 |
| Boscalid + TBT | 4 | 2 | 4.19 ± 4.76 | 2 | 0 | 1 |
| Bupirimate | 6 | 0 | 1.33 ± 0.71 | 0 | 0 | 0 |
| Bupirimate + TBT | 3 | 3 | 7.31 ± 7.85 a | 3 | 0 | 3 |
| Pendimethalin | 2 | 4 | 5.78 ± 3.00 a | 4 | 0 | 4 |
| Pendimethalin + TBT | 5 | 1 | 2.14 ± 1.72 | 1 | 0 | 1 |
| Treatment | Triglycerides | Cholesterol Esters |
|---|---|---|
| TBT | 1.52 ± 0.74 | 0.57 ± 0.13 |
| Dieldrin | 1.53 ± 0.60 | 1.08 ± 0.23 |
| Dieldrin + TBT | 2.43 ± 4.51 a | 1.03 ± 0.47 |
| Propiconazole | 2.79 ± 0.72 | 1.19 ± 0.43 |
| Propiconazole + TBT | 4.96 ± 2.68 a | 1.38 ± 0.10 |
| Bifenox | 5.47 ± 1.65 | 1.48 ± 0.23 |
| Bifenox + TBT | 8.13 ± 4.74 a | 1.11 ± 0.57 |
| Boscalid | 2.10 ± 1.05 | 1.33 ± 0.73 |
| Boscalid + TBT | 3.16 ± 1.40 a | 0.75 ± 0.27 |
| Bupirimate | 2.76 ± 1.19 | 0.98 ± 0.14 |
| Bupirimate + TBT | 4.51 ± 1.61 a | 1.04 ± 0.42 |
| Pendimethalin | 3.36 ± 1.55 a | 0.86 ± 0.18 |
| Pendimethalin + TBT | 2.66 ± 1.45 | 0.95 ± 0.07 |
| Treatment | TG | FFA | Cholesterol | HDL | LDL |
|---|---|---|---|---|---|
| TBT | 1.17 ± 0.24 | 0.69 ± 0.20 | 1.12 ± 0.08 | 1.19 ± 0.1 | 1.01 ± 0.27 |
| Dieldrin | 1.47 ± 0.62 | 1.05 ± 0.40 | 1.17 ± 0.19 | 1.16 ± 0.13 | 1.11 ± 0.30 |
| Additivity | 1.64 | 0.73 | 1.29 | 1.35 | 1.12 |
| Dieldrin + TBT | 1.21 ± 0.38 | 0.90 ± 0.11 | 1.26 ± 0.18 | 1.29 ± 0.13 a | 1.4 ± 0.36 |
| Propiconazole | 1.06 ± 0.14 | 0.91 ± 0.29 | 1.07 ± 0.14 | 1.12 ± 0.13 | 0.88 ± 0.31 |
| Additivity | 1.23 | 0.6 | 1.19 | 1.31 | 0.89 |
| Propiconazole + TBT | 1.16 ± 0.23 | 0.54 ± 0.07 a | 1.31 ± 0.10 a# | 1.34 ± 0.14 a | 1.34 ± 0.78 |
| Bifenox | 0.87 ± 0.23 | 0.89 ± 0.30 | 1.21 ± 0.09 | 1.23 ± 0.14 | 0.94 ± 0.12 |
| Additivity | 1.04 | 0.57 | 1.33 | 1.42 | 0.95 |
| Bifenox + TBT | 0.81 ± 0.33 | 0.77 ± 0.17 a | 1.21 ± 0.09 | 1.27 ± 0.10 a | 0.8 ± 0.28 |
| Boscalid | 1.27 ± 0.33 | 1.01 ± 0.19 | 1.03 ± 0.18 | 1.11 ± 0.20 | 0.86 ± 0.12 |
| Additivity | 1.44 | 0.7 | 1.15 | 1.31 | 0.87 |
| Boscalid + TBT | 1.51 ± 0.60 | 0.95 ± 0.26 | 1.24 ± 0.06 # | 1.3 ± 0.09 a | 1.02 ± 0.13 |
| Bupirimate | 1.32 ± 0.55 | 1.23 ± 0.26 | 1.09 ± 0.16 | 1.09 ± 0.21 | 1.14 ± 0.20 |
| Additivity | 1.49 | 0.92 | 1.21 | 1.28 | 1.15 |
| Bupirimate + TBT | 1.16 ± 0.32 | 0.84 ± 0.21 | 1.35 ± 0.09 a | 1.36 ± 0.07 a# | 1.24 ± 0.15 |
| Pendimethalin | 1.12 ± 0.53 | 0.84 ± 0.34 | 1.24 ± 0.07 | 1.34 ± 0.08 | 1.08 ± 0.37 |
| Additivity | 1.29 | 0.52 | 1.36 | 1.54 | 1.09 |
| Pendimethalin + TBT | 1.09 ± 0.63 | 1.01 ± 0.51 | 0.9 ± 0.39 | 0.94 ± 0.42 | 0.96 ± 0.52 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dauwe, Y.; Mary, L.; Oliviero, F.; Grimaldi, M.; Balaguer, P.; Gayrard, V.; Mselli-Lakhal, L. Steatosis and Metabolic Disorders Associated with Synergistic Activation of the CAR/RXR Heterodimer by Pesticides. Cells 2023, 12, 1201. https://doi.org/10.3390/cells12081201
Dauwe Y, Mary L, Oliviero F, Grimaldi M, Balaguer P, Gayrard V, Mselli-Lakhal L. Steatosis and Metabolic Disorders Associated with Synergistic Activation of the CAR/RXR Heterodimer by Pesticides. Cells. 2023; 12(8):1201. https://doi.org/10.3390/cells12081201
Chicago/Turabian StyleDauwe, Yannick, Lucile Mary, Fabiana Oliviero, Marina Grimaldi, Patrick Balaguer, Véronique Gayrard, and Laïla Mselli-Lakhal. 2023. "Steatosis and Metabolic Disorders Associated with Synergistic Activation of the CAR/RXR Heterodimer by Pesticides" Cells 12, no. 8: 1201. https://doi.org/10.3390/cells12081201