
The Transcriptome of Chicken Migratory Primordial Germ Cells Reveals Intrinsic Sex Differences and Expression of Hallmark Germ Cell Genes
 ,
,  ,
,  and
and {kind=link}
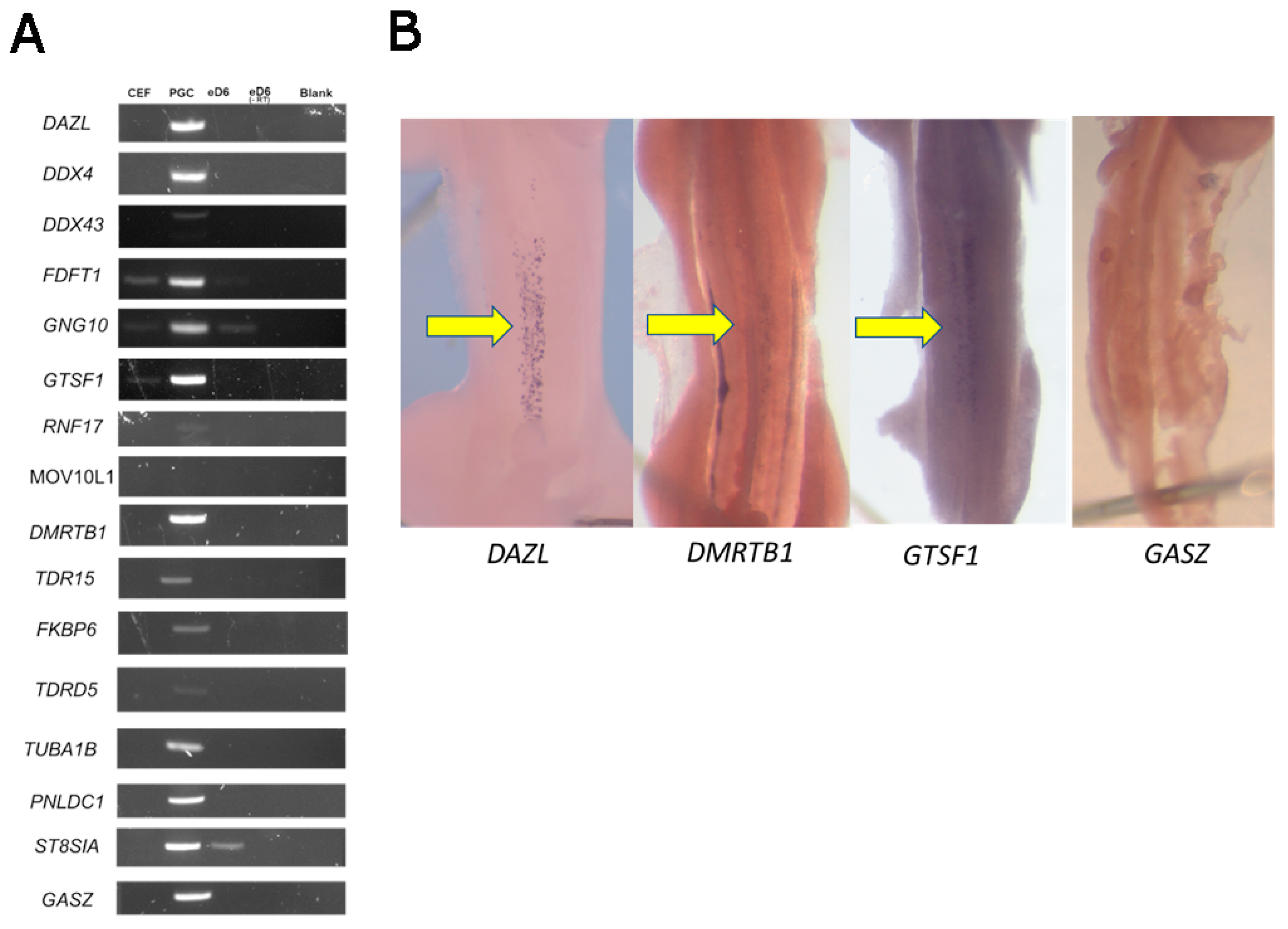{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. PGC Culture Medium
2.2. Derivation and Culturing of Chicken PGCs
2.3. RNA Isolation and Quantification
2.4. Generation of RNA-Seq Data and Genome Mapping
2.5. Transcriptome Comparison with Other Cell Types
2.6. Transcriptome Comparison with In Ovo PGCs (E2.5)
2.7. RT-PCR to Validate Germ Cell Expression
2.8. Preparation of Chicken Embryonic Fibroblasts (CEFs)
2.9. In Situ Hybridization and Probe Synthesis
2.10. Differential Gene Expression
2.11. Quantitative Reverse Transcription PCR (RT-qPCR)
3. Results
3.1. RNA Transcriptome Analysis of Cultured PGCs
3.2. Comparison of In Vitro Cultured PGCs with In Ovo PGCs at Stage 16 HH
3.3. Germ Cell-Specific Gene Expression
3.4. Validation of Germ Cell-Specific Expression
3.5. Identification of Differentially Expressed Genes (DEGs) between Male and Female PGCs


3.6. Female Differentially Expressed Genes
3.7. Male Differentially Expressed Genes
3.8. Validation of Differentially Expressed Genes by Quantitative RT-PCR
3.9. The Absence of Total Dosage Compensation in Chicken PGCs
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Glover, J.D.; Whyte, J.; Brzeszczynska, J.; Taylor, L.; Helen; Sang, M.; Mcgrew, M.J. Culturing avian primordial germ cells and novel transposon vectors for transgenesis. Transgenic Res. 2014, 23, 193. [Google Scholar]
- Woodcock, M.E.; Idoko-Akoh, A.; McGrew, M.J. Gene editing in birds takes flight. Mamm. Genome 2017, 28, 315–323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whyte, J.; Blesbois, E.; McGrew, M.J. Increased sustainability in poultry production: New tools and resources for genetic management. In Sustainable Poultry Production in Europe; CABI: Wallingford, UK, 2016; pp. 214–231. [Google Scholar] [CrossRef]
- Tsunekawa, N.; Naito, M.; Sakai, Y.; Nishida, T.; Noce, T. Isolation of chicken vasa homolog gene and tracing the origin of primordial germ cells. Development 2000, 127, 2741–2750. [Google Scholar] [CrossRef]
- Yang, S.Y.; Lee, H.J.; Lee, H.C.; Hwang, Y.S.; Park, Y.H.; Ono, T.; Han, J.Y. The dynamic development of germ cells during chicken embryogenesis. Poult. Sci. 2018, 97, 650–657. [Google Scholar] [CrossRef] [PubMed]
- Itoh, Y.; Melamed, E.; Yang, X.; Kampf, K.; Wang, S.; Yehya, N.; Van Nas, A.; Replogle, K.; Band, M.R.; Clayton, D.F.; et al. Dosage compensation is less effective in birds than in mammals. J. Biol. 2007, 6, 2. [Google Scholar] [CrossRef] [Green Version]
- Zhao, D.; McBride, D.; Nandi, S.; McQueen, H.A.; McGrew, M.J.; Hocking, P.M.; Lewis, P.D.; Sang, H.M.; Clinton, M. Somatic sex identity is cell autonomous in the chicken. Nature 2010, 464, 237–242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naito, M.; Matsubara, Y.; Harumi, T.; Tagami, T.; Kagami, H.; Sakurai, M.; Kuwana, T. Differentiation of donor primordial germ cells into functional gametes in the gonads of mixed-sex germline chimaeric chickens produced by transfer of primordial germ cells isolated from embryonic blood. Reproduction 1999, 117, 291–298. [Google Scholar] [CrossRef] [Green Version]
- Nakajima, Y.; Hattori, T.; Asano, A.; Ishikawa, N.; Tajima, A. Migration and differentiation of gonadal germ cells under cross-sex germline chimeras condition in domestic chickens. J. Reprod. Dev. 2014, 60, 406–410. [Google Scholar] [CrossRef] [Green Version]
- van de Lavoir, M.-C.; Diamond, J.H.; Leighton, P.A.; Mather-Love, C.; Heyer, B.S.; Bradshaw, R.; Kerchner, A.; Hooi, L.T.; Gessaro, T.M.; Swanberg, S.E.; et al. Germline transmission of genetically modified primordial germ cells. Nature 2006, 441, 766–769. [Google Scholar] [CrossRef] [PubMed]
- Macdonald, J.; Glover, J.D.; Taylor, L.; Sang, H.M.; McGrew, M.J. Characterisation and germline transmission of cultured avian primordial germ cells. PLoS ONE 2010, 5, e15518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tagami, T.; Kagami, H.; Matsubara, Y.; Harumi, T.; Naito, M.; Takeda, K.; Hanada, H.; Nirasawa, K. Differentiation of female primordial germ cells in the male testes of chicken (Gallus gallus domesticus). Mol. Reprod. Dev. 2006, 74, 68–75. [Google Scholar] [CrossRef]
- Soler, L.; Alves, S.; Brionne, A.; Jacques, A.; Guérin, V.; Cherif-Feildel, M.; Combes-Soia, L.; Fouchécourt, S.; Thélie, A.; Blesbois, E.; et al. Protein expression reveals a molecular sexual identity of avian primordial germ cells at pre-gonadal stages. Sci. Rep. 2021, 11, 19236. [Google Scholar] [CrossRef]
- Ballantyne, M.; Taylor, L.; Hu, T.; Meunier, D.; Nandi, S.; Sherman, A.; Flack, B.; Henshall, J.M.; Hawken, R.J.; McGrew, M.J. Avian Primordial Germ Cells Are Bipotent for Male or Female Gametogenesis. Front. Cell. Dev. Biol. 2021, 9, 726827. [Google Scholar] [CrossRef]
- Liu, C.; Chang, I.-K.; Khazanehdari, K.A.; Thomas, S.; Varghese, P.; Baskar, V.; Alkhatib, R.; Li, W.; Kinne, J.; McGrew, M.J.; et al. Uniparental chicken offsprings derived from oogenesis of chicken primordial germ cells (ZZ). Biol. Reprod. 2017, 96, 686–693. [Google Scholar] [CrossRef] [Green Version]
- Ichikawa, K.; Nakamura, Y.; Bono, H.; Ezaki, R.; Matsuzaki, M.; Horiuchi, H. Prediction of sex-determination mechanisms in avian primordial germ cells using RNA-seq analysis. Sci. Rep. 2022, 12, 13528. [Google Scholar] [CrossRef] [PubMed]
- Whyte, J.; Glover, J.D.; Woodcock, M.; Brzeszczynska, J.; Taylor, L.; Sherman, A.; Kaiser, P.; McGrew, M.J. FGF, Insulin, and SMAD Signaling Cooperate for Avian Primordial Germ Cell Self-Renewal. Stem Cell. Rep. 2015, 5, 1171–1182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katta, M.A.V.S.K.; Khan, A.W.; Doddamani, D.; Thudi, M.; Varshney, R.K. NGS-QCbox and Raspberry for Parallel, Automated and Rapid Quality Control Analysis of Large-Scale Next Generation Sequencing (Illumina) Data. PLoS ONE 2015, 10, e0139868. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liao, Y.; Smyth, G.K.; Shi, W. featureCounts: An efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics 2014, 30, 923–930. [Google Scholar] [CrossRef] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]
- Mi, H.; Muruganujan, A.; Huang, X.; Ebert, D.; Mills, C.; Guo, X.; Thomas, P.D. Protocol Update for large-scale genome and gene function analysis with the PANTHER classification system (v.14.0). Nat. Protoc. 2019, 14, 703–721. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING v11: Protein-protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kramer, A.; Green, J.; Pollard, J., Jr.; Tugendreich, S. Causal analysis approaches in Ingenuity Pathway Analysis. Bioinformatics 2014, 30, 523–530. [Google Scholar] [CrossRef] [PubMed]
- Henrique, D.; Adam, J.; Myat, A.; Chitnis, A.; Lewis, J.; Ish-Horowicz, D. Expression of a Delta homologue in prospective neurons in the chick. Nature 1995, 375, 787–790. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Zhao, R.; Zuo, Q.; Yuan, X.; Jin, K.; Jin, J.; Ding, Y.; Zhang, C.; Li, T.; Jiang, J.; Li, J.; et al. Production of viable chicken by allogeneic transplantation of primordial germ cells induced from somatic cells. Nat. Commun. 2021, 12, 2989. [Google Scholar] [CrossRef]
- Woodcock, M.E.; Gheyas, A.A.; Mason, A.S.; Nandi, S.; Taylor, L.; Sherman, A.; Smith, J.; Burt, D.W.; Hawken, R.; McGrew, M.J. Reviving rare chicken breeds using genetically engineered sterility in surrogate host birds. Proc. Natl. Acad. Sci. USA 2019, 116, 20930–20937. [Google Scholar] [CrossRef] [Green Version]
- Melamed, E.; Arnold, A.P. Regional differences in dosage compensation on the chicken Z chromosome. Genome Biol. 2007, 8, R202. [Google Scholar] [CrossRef] [Green Version]
- Ellegren, H.; Hultin-Rosenberg, L.; Brunström, B.; Dencker, L.; Kultima, K.; Scholz, B. Faced with inequality: Chicken do not have a general dosage compensation of sex-linked genes. BMC Biol. 2007, 5, 40. [Google Scholar] [CrossRef] [Green Version]
- Itoh, Y.; Replogle, K.; Kim, Y.-H.; Wade, J.; Clayton, D.F.; Arnold, A.P. Sex bias and dosage compensation in the zebra finch versus chicken genomes: General and specialized patterns among birds. Genome Res. 2010, 20, 512–518. [Google Scholar] [CrossRef] [Green Version]
- Mank, J.E.; Ellegren, H. All dosage compensation is local: Gene-by-gene regulation of sex-biased expression on the chicken Z chromosome. Heredity 2008, 102, 312–320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zimmer, F.; Harrison, P.W.; Dessimoz, C.; Mank, J.E. Compensation of Dosage-Sensitive Genes on the Chicken Z Chromosome. Genome Biol. Evol. 2016, 8, 1233–1242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, S.; Fei, T.; Zhang, L.; Zhang, R.; Chen, F.; Ning, Y.; Han, Y.; Feng, X.-H.; Meng, A.; Chen, Y.-G. Smad7 antagonizes transforming growth factor beta signaling in the nucleus by interfering with functional Smad-DNA complex formation. Mol. Cell. Biol. 2007, 27, 4488–4499. [Google Scholar] [CrossRef] [Green Version]
- Tang, Y.; Liu, Z.; Zhao, L.; Clemens, T.L.; Cao, X. Smad7 stabilizes beta-catenin binding to E-cadherin complex and promotes cell-cell adhesion. J. Biol. Chem. 2008, 283, 23956–23963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Landström, M.; Heldin, N.-E.; Bu, S.; Hermansson, A.; Itoh, S.; ten Dijke, P.; Heldin, C.-H. Smad7 mediates apoptosis induced by transforming growth factor β in prostatic carcinoma cells. Curr. Biol. 2000, 10, 535–538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, X.; Liao, H.; Cheng, M.; Shi, X.; Lin, X.; Feng, X.-H.; Chen, Y.-G. Smad7 Protein Interacts with Receptor-regulated Smads (R-Smads) to Inhibit Transforming Growth Factor-β (TGF-β)/Smad Signaling. J. Biol. Chem. 2016, 291, 382–392. [Google Scholar] [CrossRef] [PubMed] [Green Version]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Doddamani, D.; Woodcock, M.; Taylor, L.; Nandi, S.; McTeir, L.; Davey, M.G.; Smith, J.; McGrew, M.J. The Transcriptome of Chicken Migratory Primordial Germ Cells Reveals Intrinsic Sex Differences and Expression of Hallmark Germ Cell Genes. Cells 2023, 12, 1151. https://doi.org/10.3390/cells12081151
Doddamani D, Woodcock M, Taylor L, Nandi S, McTeir L, Davey MG, Smith J, McGrew MJ. The Transcriptome of Chicken Migratory Primordial Germ Cells Reveals Intrinsic Sex Differences and Expression of Hallmark Germ Cell Genes. Cells. 2023; 12(8):1151. https://doi.org/10.3390/cells12081151
Chicago/Turabian StyleDoddamani, Dadakhalandar, Mark Woodcock, Lorna Taylor, Sunil Nandi, Lynn McTeir, Megan G. Davey, Jacqueline Smith, and Mike J. McGrew. 2023. "The Transcriptome of Chicken Migratory Primordial Germ Cells Reveals Intrinsic Sex Differences and Expression of Hallmark Germ Cell Genes" Cells 12, no. 8: 1151. https://doi.org/10.3390/cells12081151