
Serotonin Receptors and Their Involvement in Melanization of Sensory Cells in Ciona intestinalis

 , , and
, , and 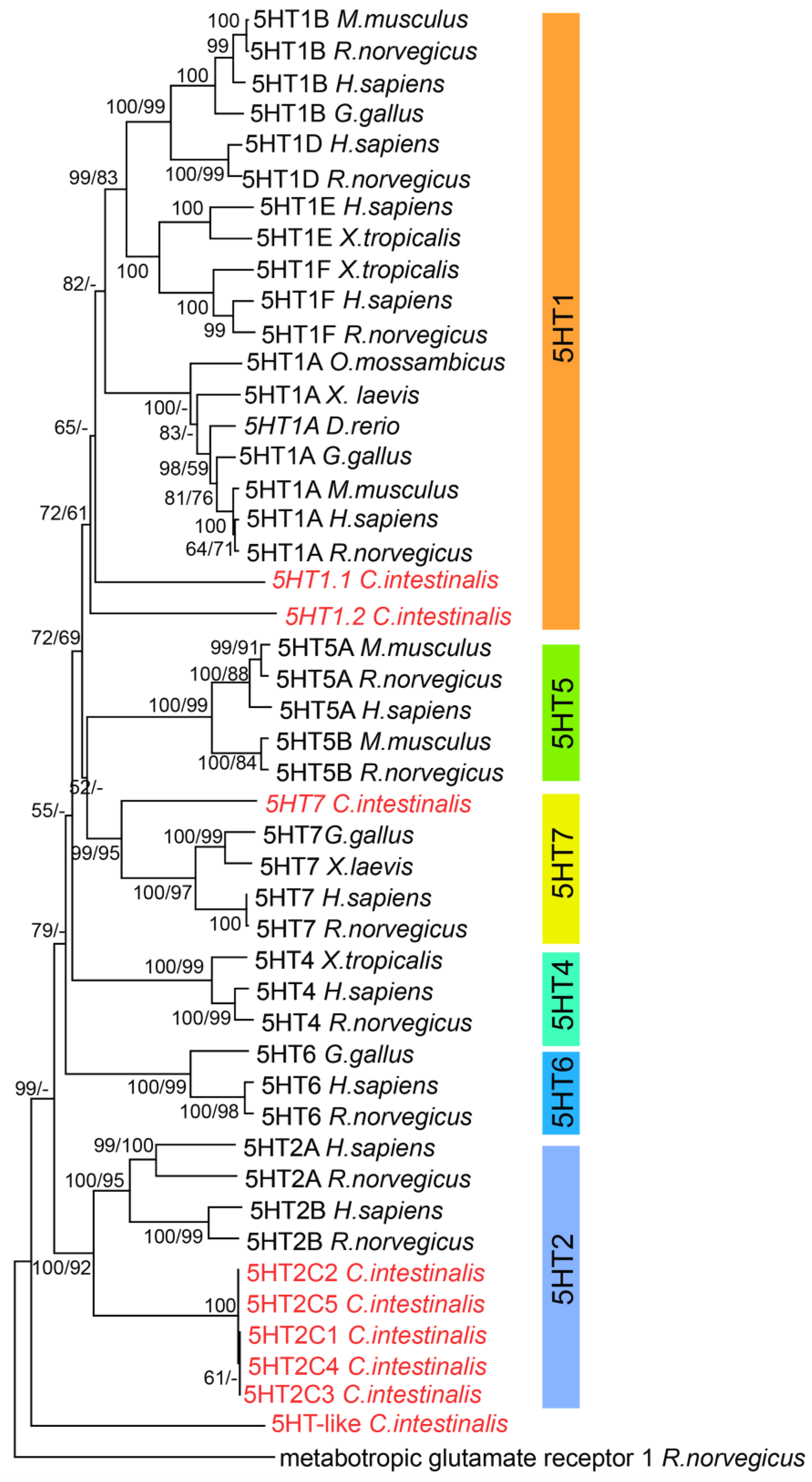{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Maintenance
2.2. RNA Preparation
2.3. Identification of 5-HT Receptor Sequences and Molecular Cloning
2.4. Phylogenetic Analysis of 5-HT Receptors (5HTRs)
2.5. WAY-100635 Exposure during Embryogenesis
2.6. Whole Mount in Situ Hybridization
3. Results
3.1. Phylogenetic Analysis of 5-HT Receptors (5HTRs)
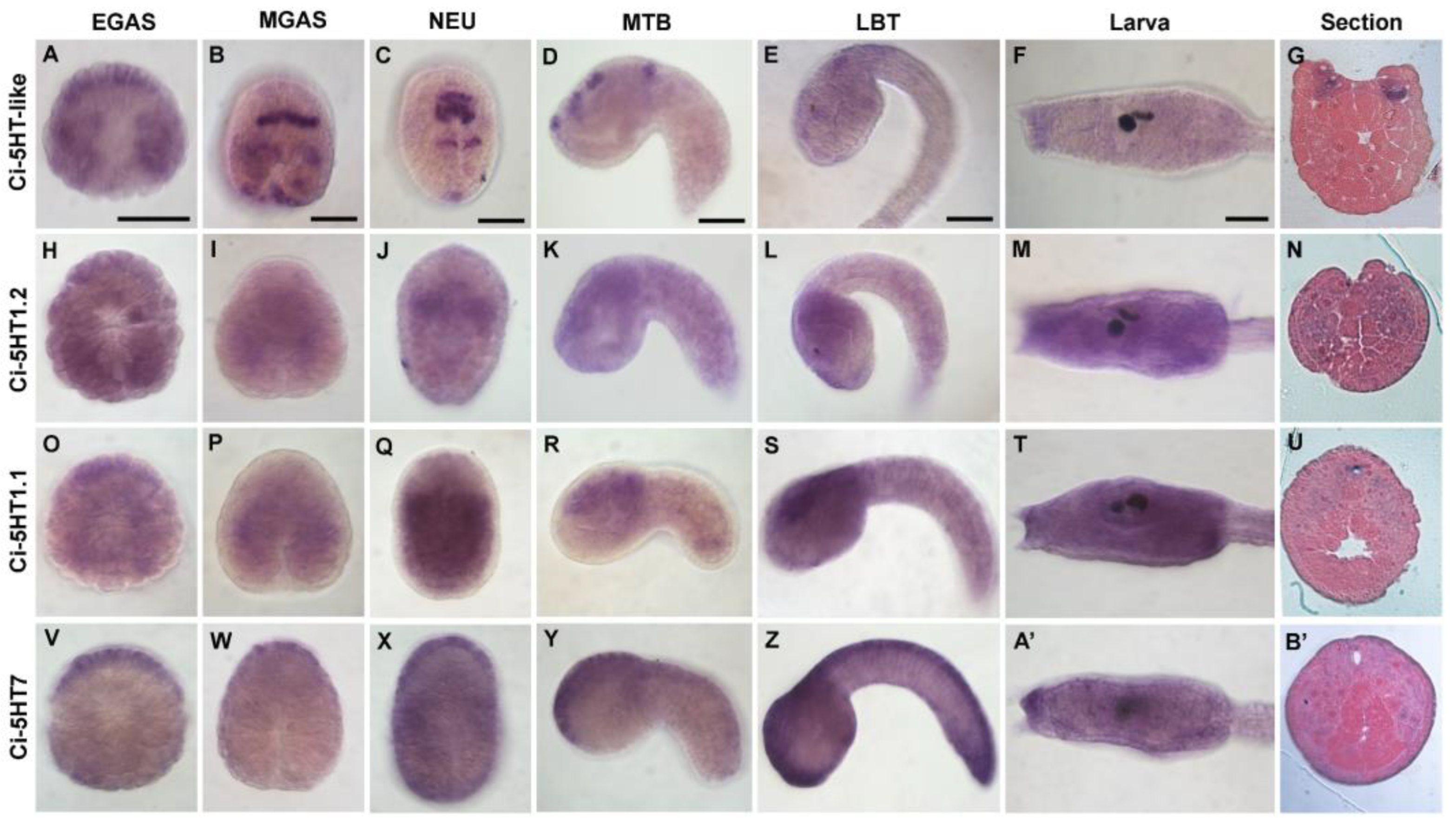
3.2. Expression Profile of 5HTRs
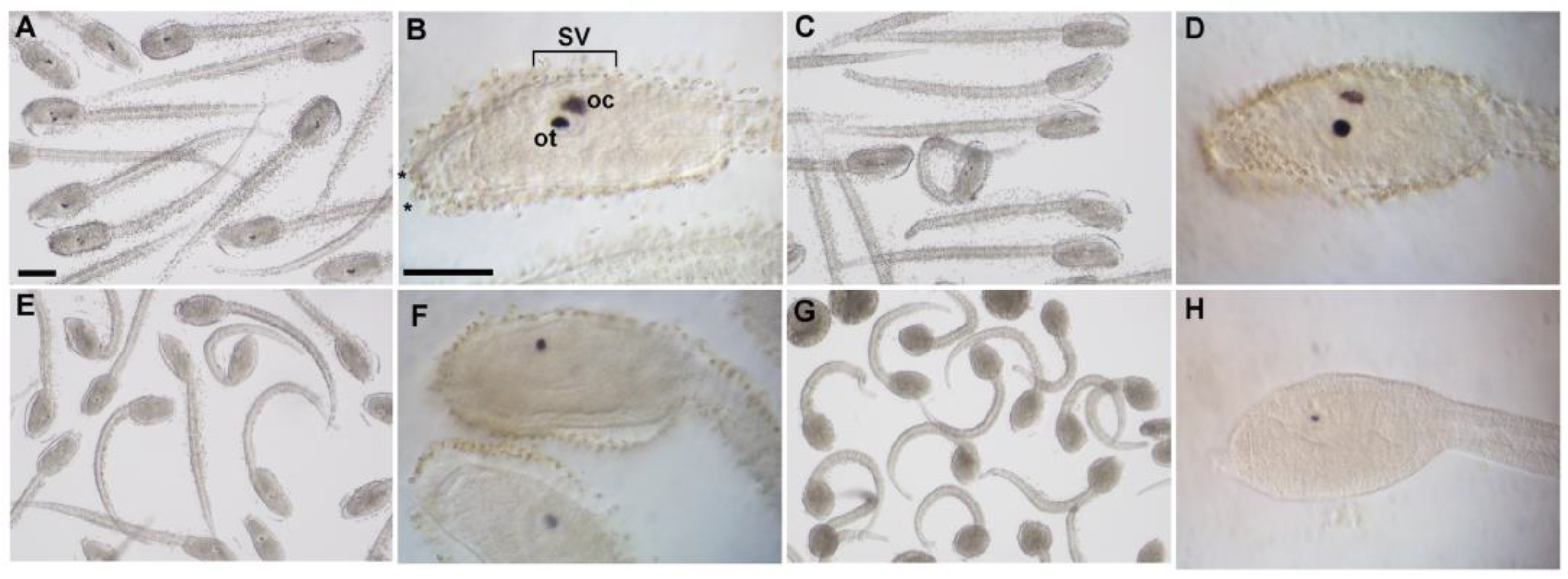
3.3. WAY-100635 Exposure Decreased Melanin Content in Ascidian Pigmented Sensory Organs
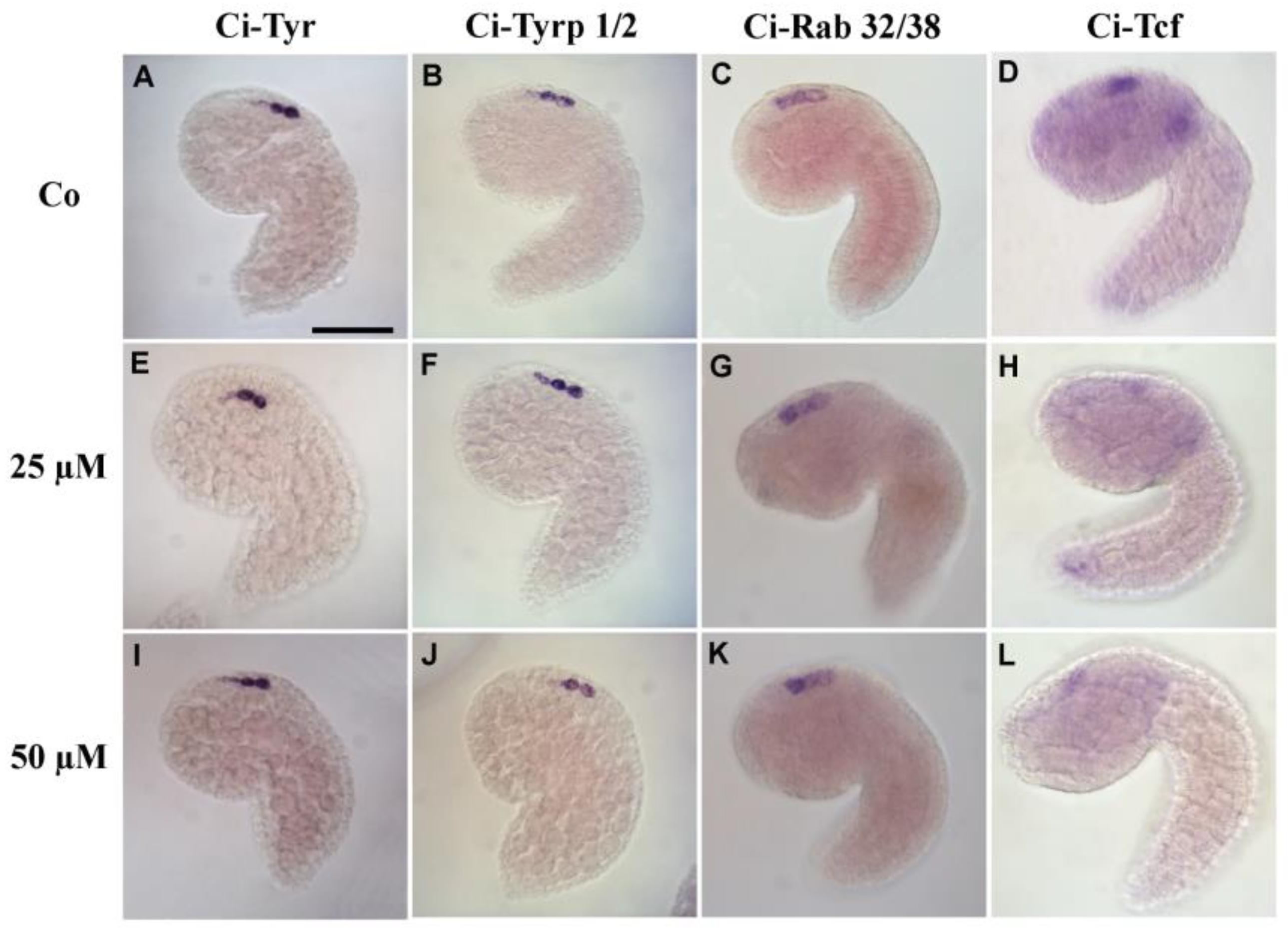
3.4. 5-HT Is Involved in Pigment Cell Terminal Differentiation
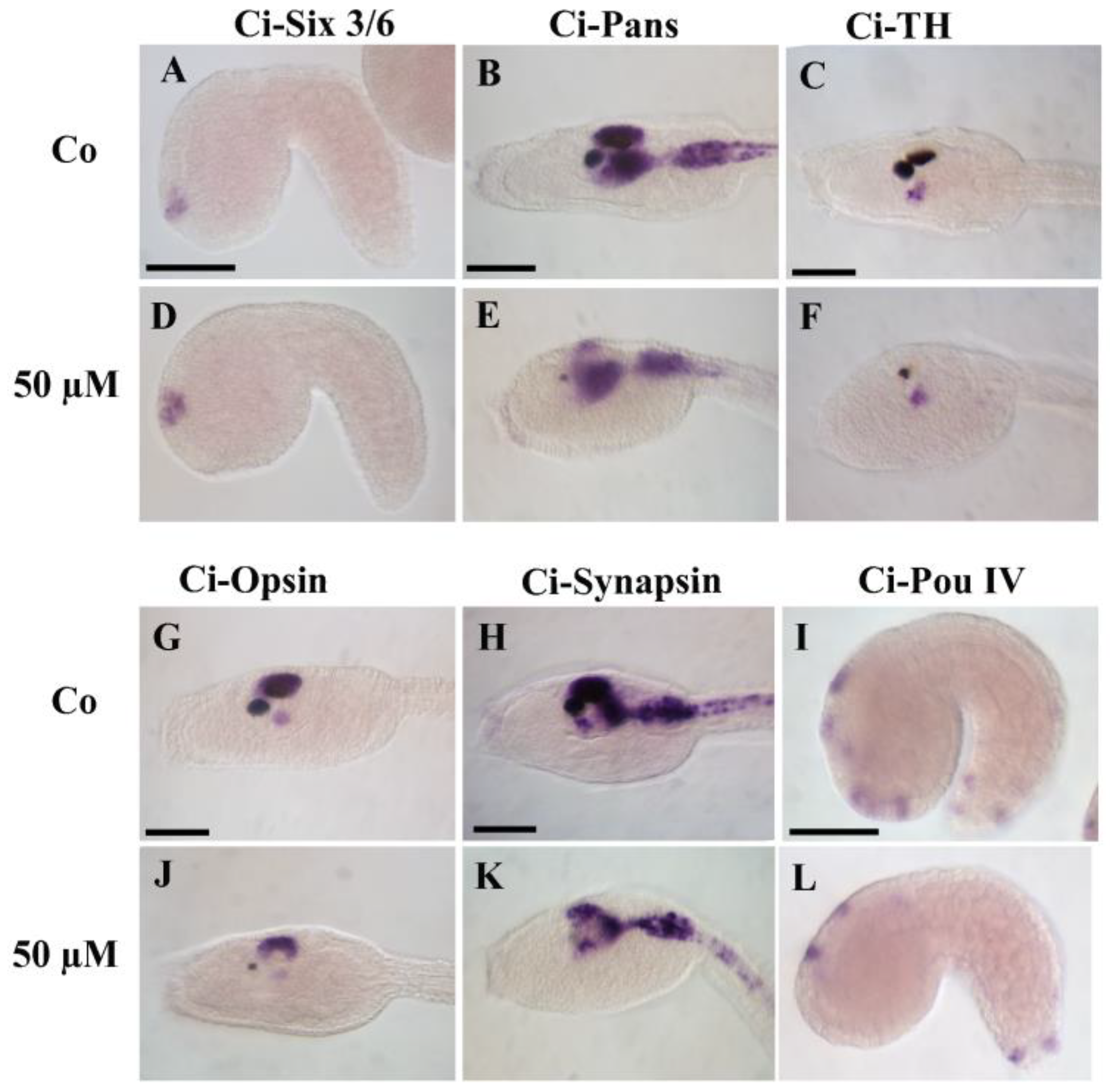
3.5. Pharmacological Inactivation of 5HTRs Disrupts the Development of Epidermal Sensory Neurons
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Berger, M.; Gray, J.A.; Roth, B.L. The expanded biology of serotonin. Annu. Rev. Med. 2009, 60, 355–366. [Google Scholar] [CrossRef] [PubMed]
- Lv, J.; Liu, F. The role of serotonin beyond the central nervous system during embryogenesis. Front. Cell. Neurosci. 2017, 11, 74. [Google Scholar] [CrossRef] [PubMed]
- Canesi, L.; Miglioli, A.; Balbi, T.; Fabbri, E. Physiological roles of serotonin in bivalves: Possible interference by environmental chemicals resulting in neuroendocrine disruption. Front. Endocrinol. 2022, 13, 792589. [Google Scholar] [CrossRef]
- Tecott, L.H. Serotonin and the Orchestration of Energy Balance. Cell Metab. 2007, 6, 352–361. [Google Scholar] [CrossRef] [PubMed]
- Gaspar, P.; Cases, O.; Maroteaux, L. The developmental role of serotonin: News from mouse molecular genetics. Nat. Rev. Neurosci. 2003, 4, 1002–1012. [Google Scholar] [CrossRef] [PubMed]
- Romero-Reyes, J.; Molina-Hernández, A.; Díaz, N.F.; Camacho-Arroyo, I. Role of serotonin in vertebrate embryo development. Reprod. Biol. 2021, 21, 100475. [Google Scholar] [CrossRef] [PubMed]
- Buznikov, G.A.; Peterson, R.E.; Nikitina, L.A.; Bezuglov, V.V.; Lauder, J.M. The pre-nervous serotonergic system of developing sea urchin embryos and larvae: Pharmacologic and immunocytochemical evidence. Neurochem. Res. 2005, 30, 825–837. [Google Scholar] [CrossRef] [PubMed]
- Nikishin, D.A.; Milošević, I.; Gojković, M.; Rakić, L.; Bezuglov, V.V.; Shmukler, Y.B. Expression and functional activity of neurotransmitter system components in sea urchins’ early development. Zygote 2016, 24, 206–218. [Google Scholar] [CrossRef]
- Sullivan, J.M.; Benton, J.L.; Beltz, B.S. Serotonin Depletion In Vivo Inhibits the Branching of Olfactory Projection Neurons in the Lobster Deutocerebrum. J. Neurosci. 2000, 20, 7716–7721. [Google Scholar] [CrossRef]
- Pytliak, M.; Vargová, V.; Mechírová, V.; Felšci, M. Serotonin receptors-from molecular biology to clinical applications. Physiol. Res. 2011, 60, 15–25. [Google Scholar] [CrossRef]
- Mohammad-Zadeh, L.F.; Moses, L.; Gwaltney-Brant, S.M. Serotonin: A review. J. Vet. Pharmacol. Ther. 2008, 31, 187–199. [Google Scholar] [CrossRef] [PubMed]
- Saitoh, O.; Yuruzume, E.; Nakata, H. Identification of planarian serotonin receptor by ligand binding and PCR studies. Neuroreport 1996, 8, 173–178. [Google Scholar] [CrossRef]
- Blenau, W.; Thamm, M. Distribution of serotonin (5-HT) and its receptors in the insect brain with focus on the mushroom bodies. Lessons from Drosophila melanogaster and Apis mellifera. Arthropod Struct. Dev. 2011, 40, 381–394. [Google Scholar] [CrossRef]
- Chen, W.; Gao, X.; Wang, H.; Xie, G.; An, S.; Du, Y.; Zhao, X. Identification and pharmacological characterization of two serotonin type 7 receptor isoforms from Mythimna separata. Int. J. Mol. Sci. 2023, 24, 655. [Google Scholar] [CrossRef]
- Ayala, F.J.; Rzhetsky, A.; Ayala, F.J. Origin of the metazoan phyla: Molecular clocks confirm paleontological estimates. Proc. Natl. Acad. Sci. USA 1998, 95, 606–611. [Google Scholar] [CrossRef] [PubMed]
- Nichols, D.E.; Nichols, C.D. Serotonin Receptors. Chem. Rev. 2008, 108, 1614–1641. [Google Scholar] [CrossRef] [PubMed]
- Tierney, A.J. Structure and function of invertebrate 5-HT receptors: A review. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2001, 128, 791–804. [Google Scholar] [CrossRef] [PubMed]
- Tierney, A.J. Invertebrate serotonin receptors: A molecular perspective on classification and pharmacology. J. Exp. Biol. 2018, 221, jeb184838. [Google Scholar] [CrossRef]
- Colas, J.F.; Launay, J.M.; Kellermann, O.; Rosay, P.; Maroteaux, L. Drosophila 5-HT2 serotonin receptor: Coexpression with fushi-tarazu during segmentation. Proc. Natl. Acad. Sci. USA 1995, 92, 5441–5445. [Google Scholar] [CrossRef]
- Nichols, C.D. 5-HT2 receptors in Drosophila are expressed in the brain and modulate aspects of circadian behaviors. Dev. Neurobiol. 2007, 67, 752–763. [Google Scholar] [CrossRef]
- Saudou, F.; Boschert, U.; Amlaiky, N.; Plassat, J.L.; Hen, R. A family of Drosophila serotonin receptors with distinct intracellular signalling properties and expression patterns. EMBO J. 1992, 11, 7–17. [Google Scholar] [CrossRef]
- Angers, A.; Storozhuk, M.V.; Duchaîne, T.; Castellucci, V.F.; DesGroseillers, L. Cloning and functional expression of an Aplysia 5-HT receptor negatively coupled to adenylate cyclase. J. Neurosci. 1998, 18, 5586–5593. [Google Scholar] [CrossRef] [PubMed]
- Gerhardt, C.C.; Leysen, J.E.; Planta, R.J.; Vreugdenhil, E.; Van Heerikhuizen, H. Functional characterisation of a 5-HT2 receptor cDNA cloned from Lymnaea stagnalis. Eur. J. Pharmacol. 1996, 311, 249–258. [Google Scholar] [CrossRef] [PubMed]
- Sugamori, K.S.; Sunahara, R.K.; Guan, H.C.; Bulloch, A.G.M.; Tensen, C.P.; Seeman, P.; Niznik, H.B.; Van Tol, H.H.M. Serotonin receptor cDNA cloned from Lymnaea stagnalis. Proc. Natl. Acad. Sci. USA 1993, 90, 11–15. [Google Scholar] [CrossRef]
- Hamdan, F.F.; Ungrin, M.D.; Abramovitz, M.; Ribeiro, P. Characterization of a novel serotonin receptor from Caenorhabditis elegans: Cloning and expression of two splice variants. J. Neurochem. 1999, 72, 1372–1383. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Duran, E.; Diaz, F.; Xiao, H.; Messer, W.S.; Komuniecki, R. Alternative-splicing of serotonin receptor isoforms in the pharynx and muscle of the parasitic nematode, Ascaris suum. Mol. Biochem. Parasitol. 1999, 101, 95–106. [Google Scholar] [CrossRef]
- Olde, B.; McCombie, W.R. Molecular cloning and functional expression of a serotonin receptor from Caenorhabditis elegans. J. Mol. Neurosci. 1997, 8, 53–62. [Google Scholar] [CrossRef]
- Burke, R.D.; Angerer, L.M.; Elphick, M.R.; Humphrey, G.W.; Yaguchi, S.; Kiyama, T.; Liang, S.; Mu, X.; Agca, C.; Klein, W.H.; et al. A genomic view of the sea urchin nervous system. Dev. Biol. 2006, 300, 434–460. [Google Scholar] [CrossRef]
- Katow, H.; Yaguchi, S.; Kiyomoto, M.; Washio, M. The 5-HT receptor cell is a new member of secondary mesenchyme cell descendants and forms a major blastocoelar network in sea urchin larvae. Mech. Dev. 2004, 121, 325–337. [Google Scholar] [CrossRef]
- Wang, T.; Yang, Z.; Zhou, N.; Sun, L.; Lv, Z.; Wu, C. Identification and functional characterisation of 5-HT4 receptor in sea cucumber Apostichopus japonicus (Selenka). Sci. Rep. 2017, 7, 40247. [Google Scholar] [CrossRef]
- Nordström, K.J.V.; Fredriksson, R.; Schiöth, H.B. The amphioxus (Branchiostoma floridae) genome contains a highly diversified set of G protein-coupled receptors. BMC Evol. Biol. 2008, 8, 9. [Google Scholar] [CrossRef] [PubMed]
- Sherwood, N.M.; Tello, J.A.; Roch, G.J. Neuroendocrinology of protochordates: Insights from Ciona genomics. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2006, 144, 254–271. [Google Scholar] [CrossRef] [PubMed]
- Stolfi, A.; Christiaen, L. Genetic and Genomic Toolbox of the Chordate Ciona intestinalis. Genetics 2012, 192, 55–66. [Google Scholar] [CrossRef] [PubMed]
- Tsuda, M.; Sakurai, D.; Goda, M. Direct evidence for the role of pigment cells in the brain of ascidian larvae by laser ablation. J. Exp. Biol. 2003, 206, 1409–1417. [Google Scholar] [CrossRef]
- Horie, T.; Sakurai, D.; Ohtsuki, H.; Terakita, A.; Shichida, Y.; Usukura, J.; Kusakabe, T.; Tsuda, M. Pigmented and nonpigmented ocelli in the brain vesicle of the ascidian larva. J. Comp. Neurol. 2008, 509, 88–102. [Google Scholar] [CrossRef]
- Waki, K.; Imai, K.S.; Satou, Y. Genetic pathways for differentiation of the peripheral nervous system in ascidians. Nat. Commun. 2015, 6, 8719. [Google Scholar] [CrossRef]
- D’Aniello, E.; Paganos, P.; Anishchenko, E.; D’Aniello, S.; Arnone, M.I. Comparative neurobiology of biogenic amines in animal models in Deuterostomes. Front. Ecol. Evol. 2020, 8, 322. [Google Scholar] [CrossRef]
- Stach, T. Comparison of the serotonergic nervous system among Tunicata: Implications for its evolution within Chordata. Org. Divers. Evol. 2005, 5, 15–24. [Google Scholar] [CrossRef]
- Pennati, R.; Groppelli, S.; Sotgia, C.; Candiani, S.; Pestarino, M.; De Bernardi, F. Serotonin localization in Phallusia mammillata larvae and effects of 5-HT antagonists during larval development. Dev. Growth Differ. 2001, 43, 647–656. [Google Scholar] [CrossRef]
- Pennati, R.; Groppelli, S.; Sotgia, C.; Zega, G.; Pestarino, M.; De Bernardi, F. WAY-100635, an antagonist of 5-HT1A receptor, causes malformations of the CNS in ascidian embryos. Dev. Genes Evol. 2003, 213, 187–192. [Google Scholar] [CrossRef]
- Fletcher, A.; Forster, E.A.; Bill, D.J.; Brown, G.; Cliffe, I.A.; Hartley, J.E.; Jones, D.E.; McLenachan, A.; Stanhope, K.J.; Critchley, D.J.P.; et al. Electrophysiological, biochemical, neurohormonal and behavioural studies with WAY-100635, a potent, selective and silent 5-HT1A receptor antagonist. Behav. Brain Res. 1995, 73, 337–353. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Fu, M.; Pei, S.; Zhou, L.; Shang, J. R-fluoxetine increases melanin synthesis through a 5-HT1A/2A receptor and p38 MAPK signaling pathways. Int. J. Mol. Sci. 2019, 20, 80. [Google Scholar] [CrossRef]
- Wu, H.L.; Pang, S.L.; Liu, Q.Z.; Wang, Q.; Cai, M.X.; Shang, J. 5-HT1A/1B receptors as targets for optimizing pigmentary responses in C57BL/6 mouse skin to stress. PLoS ONE 2014, 9, e89663. [Google Scholar] [CrossRef]
- Pennati, R.; Candiani, S.; Biggiogero, M.; Zega, G.; Groppelli, S.; Oliveri, D.; Parodi, M.; Di Bernardi, F.; Pestarino, M. Developmental expression of tryptophan hydroxylase gene in Ciona intestinalis. Dev. Genes Evol. 2007, 217, 307–313. [Google Scholar] [CrossRef] [PubMed]
- Razy-Krajka, F.; Brown, E.R.; Horie, T.; Callebert, J.; Sasakura, Y.; Joly, J.S.; Kusakabe, T.G.; Vernier, P. Monoaminergic modulation of photoreception in ascidian: Evidence for a proto-hypothalamo-retinal territory. BMC Biol. 2012, 10, 45. [Google Scholar] [CrossRef]
- Mercurio, S.; Sugni, M. Comparing in vivo and in vitro approaches to study the hormonal regulation of sea urchin reproduction. J. Mar. Biol. Assoc. United Kingd. 2016, 96, 1363–1372. [Google Scholar] [CrossRef]
- Silvia, M.; Paolo, T.; Nobile, M.; Denise, F.; Cinta, P.; Michela, S. Unraveling estradiol metabolism and involvement in the reproductive cycle of non-vertebrate animals: The sea urchin model. Steroids 2015, 104, 25–36. [Google Scholar] [CrossRef] [PubMed]
- Mercurio, S.; Cauteruccio, S.; Manenti, R.; Candiani, S.; Scarì, G.; Licandro, E.; Pennati, R. miR-7 knockdown by peptide nucleic acids in the ascidian ciona intestinalis. Int. J. Mol. Sci. 2019, 20, 5127. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular evolutionary genetics analysis version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Patocka, N.; Sharma, N.; Rashid, M.; Ribeiro, P. Serotonin Signaling in Schistosoma mansoni: A Serotonin-Activated G Protein-Coupled Receptor Controls Parasite Movement. PLoS Pathog. 2014, 10, e1003878. [Google Scholar] [CrossRef]
- Mercurio, S.; Messinetti, S.; Manenti, R.; Ficetola, G.F.; Pennati, R. Embryotoxicity characterization of the flame retardant tris(1-chloro-2-propyl)phosphate (TCPP) in the invertebrate chordate Ciona intestinalis. J. Exp. Zool. Part A Ecol. Integr. Physiol. 2021, 335, 339–347. [Google Scholar] [CrossRef] [PubMed]
- Satou, Y.; Yamada, L.; Mochizuki, Y.; Takatori, N.; Kawashima, T.; Sasaki, A.; Hamaguchi, M.; Awazu, S.; Yagi, K.; Sasakura, Y.; et al. A cDNA resource from the basal chordate Ciona intestinalis. Genesis 2002, 33, 153–154. [Google Scholar] [CrossRef] [PubMed]
- Hotta, K.; Mitsuhara, K.; Takahashi, H.; Inaba, K.; Oka, K.; Gojobori, T.; Ikeo, K. A web-based interactive developmental table for the Ascidian Ciona intestinalis, including 3D real-image embryo reconstructions: I. From fertilized egg to hatching larva. Dev. Dyn. 2007, 236, 1790–1805. [Google Scholar] [CrossRef]
- Esposito, R.; D’Aniello, S.; Squarzoni, P.; Pezzotti, M.R.; Ristoratore, F.; Spagnuolo, A. New insights into the evolution of metazoan tyrosinase gene family. PLoS ONE 2012, 7, e35731. [Google Scholar] [CrossRef]
- Haupaix, N.; Abitua, P.B.; Sirour, C.; Yasuo, H.; Levine, M.; Hudson, C. Ephrin-mediated restriction of ERK1/2 activity delimits the number of pigment cells in the Ciona CNS. Dev. Biol. 2014, 394, 170–180. [Google Scholar] [CrossRef]
- Olivo, P.; Palladino, A.; Ristoratore, F.; Spagnuolo, A. Brain Sensory Organs of the Ascidian Ciona robusta: Structure, Function and Developmental Mechanisms. Front. Cell Dev. Biol. 2021, 9, 701779. [Google Scholar] [CrossRef] [PubMed]
- Racioppi, C.; Kamal, A.K.; Razy-Krajka, F.; Gambardella, G.; Zanetti, L.; Di Bernardo, D.; Sanges, R.; Christiaen, L.A.; Ristoratore, F. Fibroblast growth factor signalling controls nervous system patterning and pigment cell formation in Ciona intestinalis. Nat. Commun. 2014, 5, 4830. [Google Scholar] [CrossRef]
- Racioppi, C.; Coppola, U.; Christiaen, L.; Ristoratore, F. Transcriptional regulation of Rab32/38, a specific marker of pigment cell formation in Ciona robusta. Dev. Biol. 2019, 448, 111–118. [Google Scholar] [CrossRef]
- Squarzoni, P.; Parveen, F.; Zanetti, L.; Ristoratore, F.; Spagnuolo, A. FGF/MAPK/Ets signaling renders pigment cell precursors competent to respond to Wnt signal by directly controlling Ci-Tcf transcription. Development 2011, 138, 1421–1432. [Google Scholar] [CrossRef]
- Garstang, M.G.; Osborne, P.W.; Ferrier, D.E.K. TCF/Lef regulates the Gsx ParaHox gene in central nervous system development in chordates. BMC Evol. Biol. 2016, 16, 57. [Google Scholar] [CrossRef]
- Mazet, F.; Hutt, J.A.; Milloz, J.; Millard, J.; Graham, A.; Shimeld, S.M. Molecular evidence from Ciona intestinalis for the evolutionary origin of vertebrate sensory placodes. Dev. Biol. 2005, 282, 494–508. [Google Scholar] [CrossRef] [PubMed]
- Alfano, C.; Teresa Russo, M.; Spagnuolo, A. Developmental expression and transcriptional regulation of Ci-Pans, a novel neural marker gene of the ascidian, Ciona intestinalis. Gene 2007, 406, 36–41. [Google Scholar] [CrossRef]
- Zega, G.; Candiani, S.; Groppelli, S.; De Bernardi, F.; Pennati, R. Neurotoxic effect of the herbicide paraquat on ascidian larvae. Environ. Toxicol. Pharmacol. 2010, 29, 24–31. [Google Scholar] [CrossRef]
- Kusakabe, T.; Kusakabe, R.; Kawakami, I.; Satou, Y.; Satoh, N.; Tsuda, M. Ci-opsin1, a vertebrate-type opsin gene, expressed in the larval ocellus of the ascidian Ciona intestinalis. FEBS Lett. 2001, 506, 69–72. [Google Scholar] [CrossRef]
- Fricker, A.D.; Rios, C.; Devi, L.A.; Gomes, I. Serotonin receptor activation leads to neurite outgrowth and neuronal survival. Mol. Brain Res. 2005, 138, 228–235. [Google Scholar] [CrossRef] [PubMed]
- Rojas, P.S.; Neira, D.; Muñoz, M.; Lavandero, S.; Fiedler, J.L. Serotonin (5-HT) regulates neurite outgrowth through 5-HT 1A and 5-HT 7 receptors in cultured hippocampal neurons. J. Neurosci. Res. 2014, 92, 1000–1009. [Google Scholar] [CrossRef] [PubMed]
- Candiani, S.; Moronti, L.; Pennati, R.; De Bernardi, F.; Benfenati, F.; Pestarino, M. The synapsin gene family in basal chordates: Evolutionary perspectives in metazoans. BMC Evol. Biol. 2010, 10, 32. [Google Scholar] [CrossRef]
- Candiani, S.; Pennati, R.; Oliveri, D.; Locascio, A.; Branno, M.; Castagnola, P.; Pestarino, M.; De Bernardi, F. Ci-POU-IV expression identifies PNS neurons in embryos and larvae of the ascidian Ciona intestinalis. Dev. Genes Evol. 2005, 215, 41–45. [Google Scholar] [CrossRef]
- Ori, M.; De- Lucchini, S.; Marras, G.; Nardi, I. Unraveling new roles for serotonin receptor 2B in development: Key findings from Xenopus. Int. J. Dev. Biol. 2013, 57, 707–714. [Google Scholar] [CrossRef]
- Yuan, Q.; Lin, F.; Zheng, X.; Sehgal, A. Serotonin modulates circadian entrainment in Drosophila. Neuron 2005, 47, 115–127. [Google Scholar] [CrossRef]
- Del Marmol, V.; Beermann, F. Tyrosinase and related proteins in mammalian pigmentation. FEBS Lett. 1996, 381, 165–168. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, K.; Satou, Y.; Yamamoto, H.; Satoh, N. A Genome-Wide Survey of Genes for Enzymes Involved in Pigment Synthesis in an Ascidian, Ciona intestinalis. Zool. Sci. 2005, 22, 723–734. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.J.; Park, M.K.; Kim, S.Y.; Park Choo, H.Y.; Lee, A.Y.; Lee, C.H. Serotonin induces melanogenesis via serotonin receptor 2A. Br. J. Dermatol. 2011, 165, 1344–1348. [Google Scholar] [CrossRef] [PubMed]
- Yue, Y.; Zhong, M.; An, X.; Feng, Q.; Lai, Y.; Yu, M.; Zhang, X.; Liao, Z.; Chen, M.; Dong, J.; et al. Serotonin (5-HT) 2A receptor involvement in melanin synthesis and transfer via activating the PKA/CREB signaling pathway. Int. J. Mol. Sci. 2022, 23, 6111. [Google Scholar] [CrossRef]
- Abitua, P.B.; Wagner, E.; Navarrete, I.A.; Levine, M. Identification of a rudimentary neural crest in a non-vertebrate chordate. Nature 2012, 492, 104–107. [Google Scholar] [CrossRef]
- Beyer, T.; Danilchik, M.; Thumberger, T.; Vick, P.; Tisler, M.; Schneider, I.; Bogusch, S.; Andre, P.; Ulmer, B.; Walentek, P.; et al. Serotonin signaling is required for Wnt-dependent GRP specification and leftward flow in Xenopus. Curr. Biol. 2012, 22, 33–39. [Google Scholar] [CrossRef]
- Hui, J.; Zhang, J.; Kim, H.; Tong, C.; Ying, Q.; Li, Z.; Mao, X.; Shi, G.; Yan, J.; Zhang, Z.; et al. Fluoxetine regulates neurogenesis in vitro through modulation of GSK-3B/B-Catenin signaling. Int. J. Neuropsychopharmacol. 2015, 18, pyu099. [Google Scholar] [CrossRef]
- Polter, A.M.; Li, X. Glycogen synthase kinase-3 is an intermediate modulator of serotonin neurotransmission. Front. Mol. Neurosci. 2011, 4, 31. [Google Scholar] [CrossRef]
- Joyce Tang, W.; Chen, J.S.; Zeller, R.W. Transcriptional regulation of the peripheral nervous system in Ciona intestinalis. Dev. Biol. 2013, 378, 183–193. [Google Scholar] [CrossRef]
- Roure, A.; Darras, S. Msxb is a core component of the genetic circuitry specifying the dorsal and ventral neurogenic midlines in the ascidian embryo. Dev. Biol. 2016, 409, 277–287. [Google Scholar] [CrossRef]
- Pasini, A.; Amiel, A.; Rothbächer, U.; Roure, A.; Lemaire, P.; Darras, S. Formation of the ascidian epidermal sensory neurons: Insights into the origin of the chordate peripheral nervous system. PLoS Biol. 2006, 4, 1173–1186. [Google Scholar] [CrossRef] [PubMed]
- Pasini, A.; Manenti, R.; Rothbächer, U.; Lemaire, P. Antagonizing Retinoic Acid and FGF/MAPK Pathways Control Posterior Body Patterning in the Invertebrate Chordate Ciona intestinalis. PLoS ONE 2012, 7, e46193. [Google Scholar] [CrossRef] [PubMed]
- Wagner, E.; Levine, M. FGF signaling establishes the anterior border of the Ciona neural tube. Development 2012, 139, 2351–2359. [Google Scholar] [CrossRef]
- Kim, K.; Gibboney, S.; Razy-Krajka, F.; Lowe, E.K.; Wang, W.; Stolfi, A. Regulation of Neurogenesis by FGF Signaling and Neurogenin in the Invertebrate Chordate Ciona. Front. Cell Dev. Biol. 2020, 8, 477. [Google Scholar] [CrossRef]
- Osman, S.; Lundkvist, C.; Pike, V.W.; Halldin, C.; McCarron, J.A.; Swahn, C.G.; Ginovart, N.; Luthra, S.K.; Bench, C.J.; Grasby, P.M.; et al. Characterization of the radioactive metabolites of the 5-HT(1A) receptor radioligand, [O-methyl-11C]WAY-100635, in monkey and human plasma by HPLC: Comparison of the behaviour of an identified radioactive metabolite with parent radioligand in monkey using. Nucl. Med. Biol. 1996, 23, 627–634. [Google Scholar] [CrossRef] [PubMed]
- Chemel, B.R.; Roth, B.L.; Armbruster, B.; Watts, V.J.; Nichols, D.E. WAY-100635 is a potent dopamine D4 receptor agonist. Psychopharmacology 2006, 188, 244–251. [Google Scholar] [CrossRef]
- Kamesh, N.; Aradhyam, G.K.; Manoj, N. The repertoire of G protein-coupled receptors in the sea squirt Ciona intestinalis. BMC Evol. Biol. 2008, 8, 129. [Google Scholar] [CrossRef]
- Delsuc, F.; Brinkmann, H.; Chourrout, D.; Philippe, H. Tunicates and not cephalochordates are the closest living relatives of vertebrates. Nature 2006, 439, 965–968. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mercurio, S.; Bozzo, M.; Pennati, A.; Candiani, S.; Pennati, R. Serotonin Receptors and Their Involvement in Melanization of Sensory Cells in Ciona intestinalis. Cells 2023, 12, 1150. https://doi.org/10.3390/cells12081150
Mercurio S, Bozzo M, Pennati A, Candiani S, Pennati R. Serotonin Receptors and Their Involvement in Melanization of Sensory Cells in Ciona intestinalis. Cells. 2023; 12(8):1150. https://doi.org/10.3390/cells12081150
Chicago/Turabian StyleMercurio, Silvia, Matteo Bozzo, Alessandro Pennati, Simona Candiani, and Roberta Pennati. 2023. "Serotonin Receptors and Their Involvement in Melanization of Sensory Cells in Ciona intestinalis" Cells 12, no. 8: 1150. https://doi.org/10.3390/cells12081150