
Methemoglobinemia, Increased Deformability and Reduced Membrane Stability of Red Blood Cells in a Cat with a CYB5R3 Splice Defect

 , ,
, ,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals, Samples, and Clinical Examination
2.2. Blood Test Analyses
2.3. Molecular Genetic Studies and Protein Structure Modeling
3. Results
3.1. Clinical Examination and Blood Analyses
3.2. Erythrocytic Reduced Glutathione Concentration and CYB5R Activity
3.3. Erythrocyte Morphology, Deformability, and Ion Content
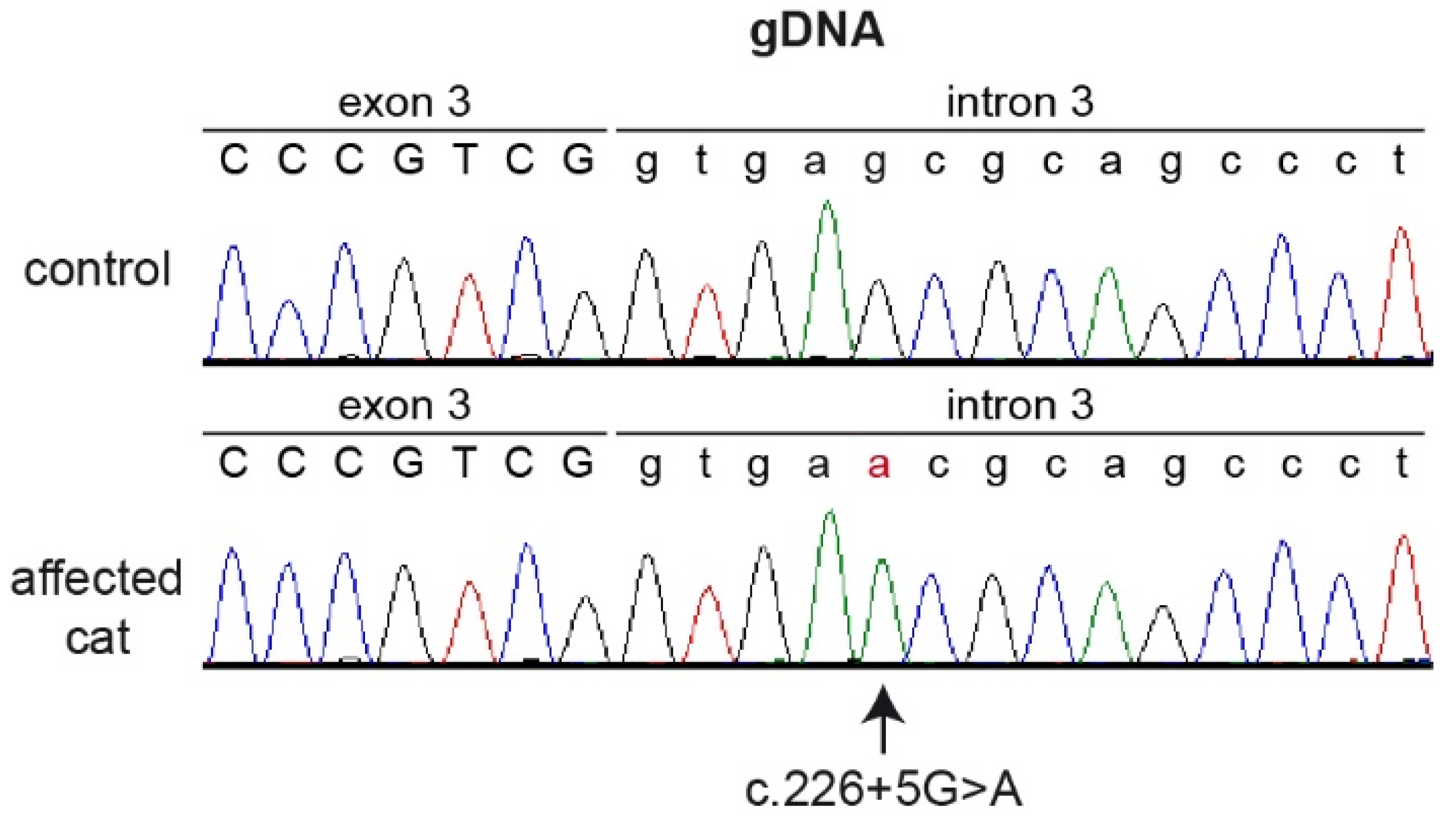
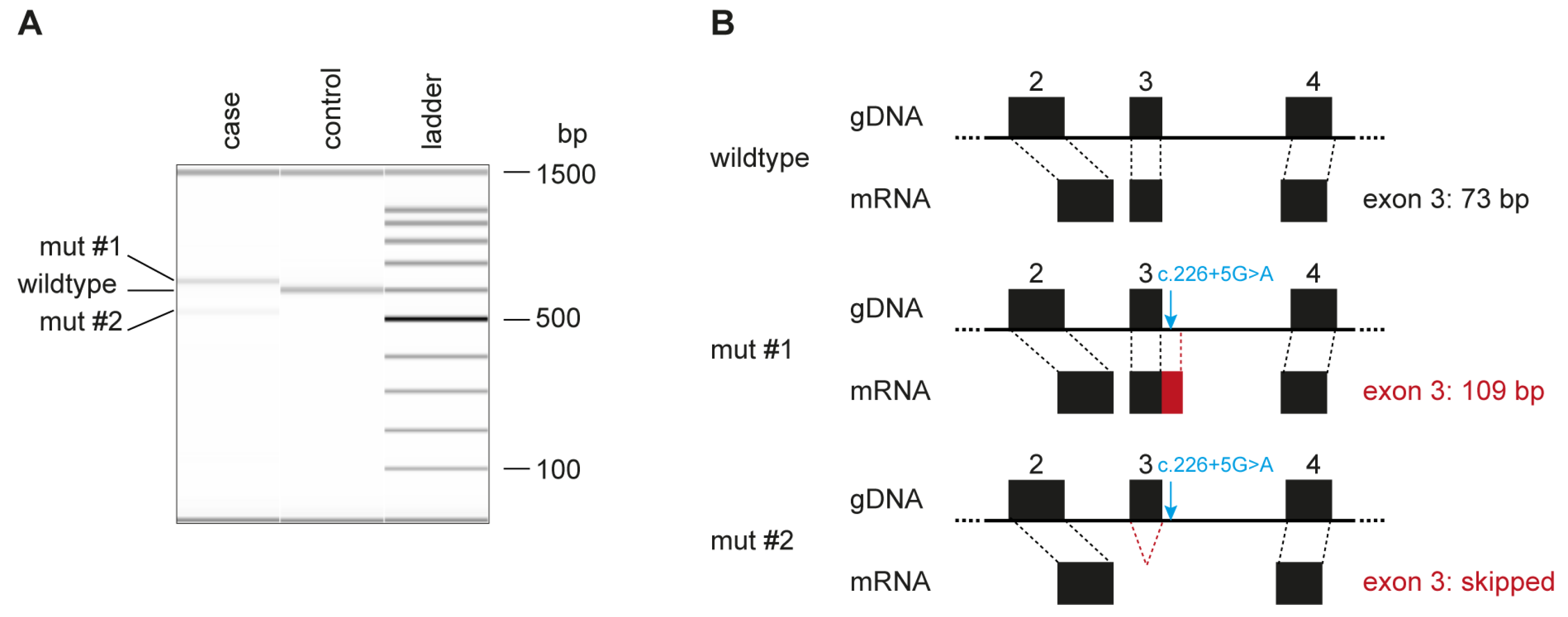
3.4. Genetic Analysis
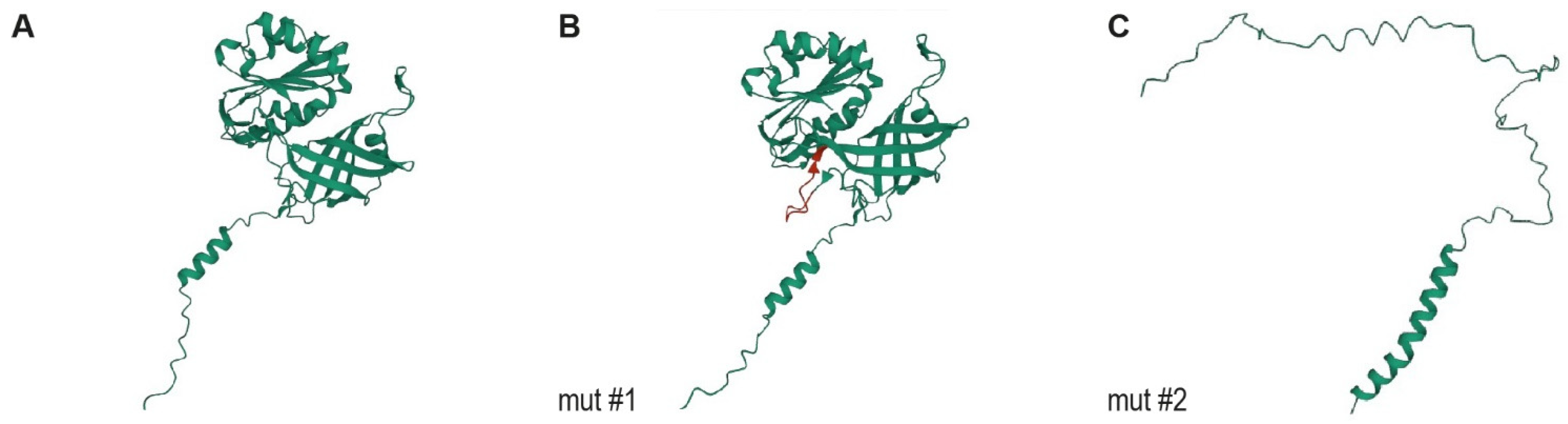
3.5. Protein Modelling
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hall, R.; Yuan, S.; Wood, K.; Katona, M.; Straub, A.C. Cytochrome b5 reductases: Redox regulators of cell homeostasis. J. Biol. Chem. 2022, 298, 102654. [Google Scholar] [CrossRef] [PubMed]
- Hall, A.H.; Kulig, K.W.; Rumack, B.H. Drug- and chemical-induced methaemoglobinaemia. Clinical features and management. Med. Toxicol. 1986, 1, 253–260. [Google Scholar] [CrossRef]
- Mansouri, A.; Lurie, A.A. Concise review: Methemoglobinemia. Am. J. Hematol. 1993, 42, 7–12. [Google Scholar] [CrossRef] [PubMed]
- Jaffe, E.R. Methaemoglobinaemia. Clin. Haematol. 1981, 10, 99–122. [Google Scholar] [CrossRef] [PubMed]
- Percy, M.J.; Lappin, T.R. Recessive congenital methaemoglobinaemia: Cytochrome b(5) reductase deficiency. Br. J. Haematol. 2008, 141, 298–308. [Google Scholar] [CrossRef] [PubMed]
- Harvey, J.W.; Stockham, S.L.; Scott, M.A.; Johnson, P.J.; Donald, J.J.; Chandler, C.J. Methemoglobinemia and eccentrocytosis in equine erythrocyte flavin adenine dinucleotide deficiency. Vet. Pathol. 2003, 40, 632–642. [Google Scholar] [CrossRef]
- Gupta, V.; Kulkarni, A.; Warang, P.; Devendra, R.; Chiddarwar, A.; Kedar, P. Mutation update: Variants of the CYB5R3 gene in recessive congenital methemoglobinemia. Hum. Mutat. 2020, 41, 737–748. [Google Scholar] [CrossRef]
- Jaffe, E.R. Enzymopenic hereditary methemoglobinemia: A clinical/biochemical classification. Blood Cells 1986, 12, 81–90. [Google Scholar]
- Wright, R.O.; Lewander, W.J.; Woolf, A.D. Methemoglobinemia: Etiology, pharmacology, and clinical management. Ann. Emerg. Med. 1999, 34, 646–656. [Google Scholar] [CrossRef]
- Ewenczyk, C.; Leroux, A.; Roubergue, A.; Laugel, V.; Afenjar, A.; Saudubray, J.M.; Beauvais, P.; Billette de Villemeur, T.; Vidailhet, M.; Roze, E. Recessive hereditary methaemoglobinaemia, type II: Delineation of the clinical spectrum. Brain 2008, 131, 760–761. [Google Scholar] [CrossRef] [Green Version]
- Siendones, E.; Ballesteros, M.; Navas, P. Cellular and Molecular Mechanisms of Recessive Hereditary Methaemoglobinaemia Type II. J. Clin. Med. 2018, 7, 341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deorukhkar, A.; Kulkarni, A.; Kedar, P. Three novel mutations in CYB5R3 gene causing NADH-cytochrome b5 reductase enzyme deficiency leads to recessive congenital methaemoglobinemia. Mol. Biol. Rep. 2022, 49, 2141–2147. [Google Scholar] [CrossRef] [PubMed]
- Ullah, A.; Shah, A.A.; Syed, F.; Mahmood, A.; Ur Rehman, H.; Khurshid, B.; Samad, A.; Ahmad, W.; Basit, S. Molecular Dynamic Simulation Analysis of a Novel Missense Variant in CYB5R3 Gene in Patients with Methemoglobinemia. Medicina 2023, 59, 379. [Google Scholar] [CrossRef]
- Harvey, J.W. Pathogenesis, laboratory diagnosis, and clinical implications of erythrocyte enzyme deficiencies in dogs, cats, and horses. Vet. Clin. Pathol. 2006, 35, 144–156. [Google Scholar] [CrossRef] [PubMed]
- Harvey, J.W.; Kaneko, J.J. Erythrocyte enzyme activities and glutathione levels of the horse, cat, dog and man. Comp. Biochem. Physiol. B 1975, 52, 507–510. [Google Scholar] [CrossRef] [PubMed]
- Harvey, J.W.; Ling, G.V.; Kaneko, J.J. Methemoglobin reductase deficiency in a dog. J. Am. Vet. Med. Assoc. 1974, 164, 1030–1033. [Google Scholar]
- Jaffey, J.A.; Harmon, M.R.; Villani, N.A.; Creighton, E.K.; Johnson, G.S.; Giger, U.; Dodam, J.R. Long-term Treatment with Methylene Blue in a Dog with Hereditary Methemoglobinemia Caused by Cytochrome b5 Reductase Deficiency. J. Vet. Intern. Med. 2017, 31, 1860–1865. [Google Scholar] [CrossRef] [Green Version]
- Jaffey, J.A.; Reading, N.S.; Abdulmalik, O.; Kreisler, R.; Bullock, G.; Wiest, A.; Villani, N.A.; Mhlanga-Mutangadura, T.; Johnson, G.S.; Cohn, L.A.; et al. Clinical, metabolic, and molecular genetic characterization of hereditary methemoglobinemia caused by cytochrome b5 reductase deficiency in 30 dogs. Sci. Rep. 2020, 10, 21399. [Google Scholar] [CrossRef]
- McKenna, J.A.; Sacco, J.; Son, T.T.; Trepanier, L.A.; Callan, M.B.; Harvey, J.W.; Arndt, J.W. Congenital methemoglobinemia in a dog with a promoter deletion and a nonsynonymous coding variant in the gene encoding cytochrome b(5). J. Vet. Intern. Med. 2014, 28, 1626–1631. [Google Scholar] [CrossRef] [Green Version]
- Shino, H.; Otsuka-Yamasaki, Y.; Sato, T.; Ooi, K.; Inanami, O.; Sato, R.; Yamasaki, M. Familial Congenital Methemoglobinemia in Pomeranian Dogs Caused by a Missense Variant in the NADH-Cytochrome B5 Reductase Gene. J. Vet. Intern. Med. 2018, 32, 165–171. [Google Scholar] [CrossRef] [Green Version]
- Zhou, S.; Tearle, R.; Jozani, R.J.; Winra, B.; Schaaf, O.; Nicholson, A.; Peaston, A. Genetic cause for congenital methemoglobinemia in an Australian Pomeranian dog. J. Vet. Intern. Med. 2019, 33, 868–873. [Google Scholar] [CrossRef] [PubMed]
- Harvey, J.W.; Dahl, M.; High, M.E. Methemoglobin reductase deficiency in a cat. J. Am. Vet. Med. Assoc. 1994, 205, 1290–1291. [Google Scholar] [PubMed]
- Jaffey, J.A.; Reading, N.S.; Giger, U.; Abdulmalik, O.; Buckley, R.M.; Johnstone, S.; Lyons, L.A.; Lives Cat Genome, C. Clinical, metabolic, and genetic characterization of hereditary methemoglobinemia caused by cytochrome b5 reductase deficiency in cats. J. Vet. Intern. Med. 2019, 33, 2725–2731. [Google Scholar] [CrossRef] [Green Version]
- Rumbeiha, W.K.; Oehme, F.W. Methylene blue can be used to treat methemoglobinemia in cats without inducing Heinz body hemolytic anemia. Vet. Hum. Toxicol. 1992, 34, 120–122. [Google Scholar] [PubMed]
- Vasiliadou, E.; Karakitsou, V.; Kazakos, G.; Oikonomidis, I.L.; Tsouloufi, T.K.; Kosmas, P.; Abdulmalik, O.Y.; Koutinas, C.; Giger, U.; Mylonakis, M.E. Hereditary methemoglobinemia in a cyanotic cat presented for ovariohysterectomy. Can. Vet. J. 2019, 60, 502–506. [Google Scholar]
- Harvey, J.W. Congenital erythrocyte enzyme deficiencies. Vet. Clin. N. Am. Small Anim. Pract. 1996, 26, 1003–1011. [Google Scholar] [CrossRef]
- Hegesh, E.; Calmanovici, N.; Avron, M. New method for determining ferrihemoglobin reductase (NADH-methemoglobin reductase) in erythrocytes. J. Lab. Clin. Med. 1968, 72, 339–344. [Google Scholar]
- Beutler, E. Red Cell Metabolism. A Manual of Biochemical Methods; Grune & Stratton: New York, NY, USA; San Francisco, CA, USA; London, UK, 1975. [Google Scholar]
- Tietze, F. Enzymic method for quantitative determination of nanogram amounts of total and oxidized glutathione: Applications to mammalian blood and other tissues. Anal. Biochem. 1969, 27, 502–522. [Google Scholar] [CrossRef]
- Bogdanova, A.Y.; Ogunshola, O.O.; Bauer, C.; Gassmann, M. Pivotal role of reduced glutathione in oxygen-induced regulation of the Na(+)/K(+) pump in mouse erythrocyte membranes. J. Membr. Biol. 2003, 195, 33–42. [Google Scholar] [CrossRef] [Green Version]
- van Cromvoirt, A.M.; Fenk, S.; Sadafi, A.; Melnikova, E.V.; Lagutkin, D.A.; Dey, K.; Petrushanko, I.Y.; Hegemann, I.; Goede, J.S.; Bogdanova, A. Donor Age and Red Cell Age Contribute to the Variance in Lorrca Indices in Healthy Donors for Next Generation Ektacytometry: A Pilot Study. Front. Physiol. 2021, 12, 639722. [Google Scholar] [CrossRef]
- Jagannathan, V.; Drogemuller, C.; Leeb, T.; Dog Biomedical Variant Database Consortium (DBVDC). A comprehensive biomedical variant catalogue based on whole genome sequences of 582 dogs and eight wolves. Anim. Genet. 2019, 50, 695–704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cingolani, P.; Platts, A.; le Wang, L.; Coon, M.; Nguyen, T.; Wang, L.; Land, S.J.; Lu, X.; Ruden, D.M. A program for annotating and predicting the effects of single nucleotide polymorphisms, SnpEff: SNPs in the genome of Drosophila melanogaster strain w1118; iso-2; iso-3. Fly 2012, 6, 80–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mirdita, M.; Schutze, K.; Moriwaki, Y.; Heo, L.; Ovchinnikov, S.; Steinegger, M. ColabFold: Making protein folding accessible to all. Nat. Methods 2022, 19, 679–682. [Google Scholar] [CrossRef] [PubMed]
- Ayres, D.A. Pulse oximetry and CO-oximetry. In Advanced Monitoring and Procedured for Small Animal Emergency and Critical Care; Burkitt Creedon, J.M., Davis, H., Eds.; Willey-Blackwell: Hoboken, NJ, USA, 2012; pp. 274–285. [Google Scholar]
- Huisjes, R.; Bogdanova, A.; van Solinge, W.W.; Schiffelers, R.M.; Kaestner, L.; van Wijk, R. Squeezing for Life—Properties of Red Blood Cell Deformability. Front. Physiol. 2018, 9, 656. [Google Scholar] [CrossRef] [Green Version]
- Stalder, L.; Muhlemann, O. The meaning of nonsense. Trends Cell Biol. 2008, 18, 315–321. [Google Scholar] [CrossRef]
- Otsuka-Yamasaki, Y.; Inanami, O.; Shino, H.; Sato, R.; Yamasaki, M. Characterization of a novel nicotinamide adenine dinucleotide-cytochrome b5 reductase mutation associated with canine hereditary methemoglobinemia. J. Vet. Med. Sci. 2021, 83, 315–321. [Google Scholar] [CrossRef]
- Haymond, S.; Cariappa, R.; Eby, C.S.; Scott, M.G. Laboratory assessment of oxygenation in methemoglobinemia. Clin. Chem. 2005, 51, 434–444. [Google Scholar] [CrossRef]
- Arashiki, N.; Kimata, N.; Manno, S.; Mohandas, N.; Takakuwa, Y. Membrane peroxidation and methemoglobin formation are both necessary for band 3 clustering: Mechanistic insights into human erythrocyte senescence. Biochemistry 2013, 52, 5760–5769. [Google Scholar] [CrossRef] [Green Version]
- Petrushanko, I.; Bogdanov, N.; Bulygina, E.; Grenacher, B.; Leinsoo, T.; Boldyrev, A.; Gassmann, M.; Bogdanova, A. Na-K-ATPase in rat cerebellar granule cells is redox sensitive. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2006, 290, R916–R925. [Google Scholar] [CrossRef] [Green Version]
- Rossi, R.; Giustarini, D.; Milzani, A.; Dalle-Donne, I. Membrane skeletal protein S-glutathionylation and hemolysis in human red blood cells. Blood Cells Mol. Dis. 2006, 37, 180–187. [Google Scholar] [CrossRef]
- Ogasawara, Y.; Komiyama, M.; Funakoshi, M.; Ishii, K. Disruption of glutathione homeostasis causes accumulation of S-glutathionyl proteins in response to exposure to reactive oxygen species in human erythrocytes. Biol. Pharm. Bull. 2010, 33, 1925–1931. [Google Scholar] [CrossRef] [Green Version]
- Giustarini, D.; Dalle-Donne, I.; Milzani, A.; Braconi, D.; Santucci, A.; Rossi, R. Membrane Skeletal Protein S-Glutathionylation in Human Red Blood Cells as Index of Oxidative Stress. Chem. Res. Toxicol. 2019, 32, 1096–1102. [Google Scholar] [CrossRef]
- Bohmer, A.; Pich, A.; Schmidt, M.; Haghikia, A.; Tsikas, D. Evidence by chromatography and mass spectrometry that inorganic nitrite induces S-glutathionylation of hemoglobin in human red blood cells. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2016, 1019, 72–82. [Google Scholar] [CrossRef] [PubMed]
- Hanggi, P.; Makhro, A.; Gassmann, M.; Schmugge, M.; Goede, J.S.; Speer, O.; Bogdanova, A. Red blood cells of sickle cell disease patients exhibit abnormally high abundance of N-methyl D-aspartate receptors mediating excessive calcium uptake. Br. J. Haematol. 2014, 167, 252–264. [Google Scholar] [CrossRef] [PubMed]
- Hertz, L.; Huisjes, R.; Llaudet-Planas, E.; Petkova-Kirova, P.; Makhro, A.; Danielczok, J.G.; Egee, S.; Del Mar Manu-Pereira, M.; van Wijk, R.; Vives Corrons, J.L.; et al. Is Increased Intracellular Calcium in Red Blood Cells a Common Component in the Molecular Mechanism Causing Anemia? Front. Physiol. 2017, 8, 673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Makhro, A.; Hegemann, I.; Seiler, E.; Simionato, G.; Claveriam, V.; Bogdanov, N.; Sasselli, C.; Torgeson, P.; Kaestner, L.; Manz, M.G.; et al. A pilot clinical phase II trial MemSID: Acute and durable changes of red blood cells of sickle cell disease patients on memantine treatment. eJHaem 2020, 1, 23–34. [Google Scholar]
- Kampf, S.; Seiler, E.; Bujok, J.; Hofmann-Lehmann, R.; Riond, B.; Makhro, A.; Bogdanova, A. Aging Markers in Equine Red Blood Cells. Front. Physiol. 2019, 10, 893. [Google Scholar] [CrossRef] [Green Version]
- Makhro, A.; Hanggi, P.; Goede, J.S.; Wang, J.; Bruggemann, A.; Gassmann, M.; Schmugge, M.; Kaestner, L.; Speer, O.; Bogdanova, A. N-methyl-D-aspartate receptors in human erythroid precursor cells and in circulating red blood cells contribute to the intracellular calcium regulation. Am. J. Physiol. Cell Physiol. 2013, 305, C1123–C1138. [Google Scholar] [CrossRef] [Green Version]
- Bernstein, R.E. Potassium and sodium balance in mammalian red cells. Science 1954, 120, 459–460. [Google Scholar] [CrossRef]
- Sega, M.F.; Chu, H.; Christian, J.; Low, P.S. Interaction of deoxyhemoglobin with the cytoplasmic domain of murine erythrocyte band 3. Biochemistry 2012, 51, 3264–3272. [Google Scholar] [CrossRef] [Green Version]
- Chu, H.; McKenna, M.M.; Krump, N.A.; Zheng, S.; Mendelsohn, L.; Thein, S.L.; Garrett, L.J.; Bodine, D.M.; Low, P.S. Reversible binding of hemoglobin to band 3 constitutes the molecular switch that mediates O2 regulation of erythrocyte properties. Blood 2016, 128, 2708–2716. [Google Scholar] [CrossRef] [PubMed]
- Chu, H.; Breite, A.; Ciraolo, P.; Franco, R.S.; Low, P.S. Characterization of the deoxyhemoglobin binding site on human erythrocyte band 3: Implications for O2 regulation of erythrocyte properties. Blood 2008, 111, 932–938. [Google Scholar] [CrossRef] [Green Version]
- Waugh, S.M.; Low, P.S. Hemichrome binding to band 3: Nucleation of Heinz bodies on the erythrocyte membrane. Biochemistry 1985, 24, 34–39. [Google Scholar] [CrossRef]
- Narla, J.; Mohandas, N. Red cell membrane disorders. Int. J. Lab. Hematol. 2017, 39 (Suppl. S1), 47–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stefanovic, M.; Puchulu-Campanella, E.; Kodippili, G.; Low, P.S. Oxygen regulates the band 3-ankyrin bridge in the human erythrocyte membrane. Biochem. J. 2013, 449, 143–150. [Google Scholar] [CrossRef] [Green Version]
- Zhou, S.; Giannetto, M.; DeCourcey, J.; Kang, H.; Kang, N.; Li, Y.; Zheng, S.; Zhao, H.; Simmons, W.R.; Wei, H.S.; et al. Oxygen tension-mediated erythrocyte membrane interactions regulate cerebral capillary hyperemia. Sci. Adv. 2019, 5, eaaw4466. [Google Scholar] [CrossRef] [Green Version]
- Alaarg, A.; Schiffelers, R.M.; van Solinge, W.W.; van Wijk, R. Red blood cell vesiculation in hereditary hemolytic anemia. Front. Physiol. 2013, 4, 365. [Google Scholar] [CrossRef] [PubMed]
- Jaffe, E.R.; Neurmann, G. A Comparision of the Effect of Menadione, Methylene Blue and Ascorbic Acid on the Reduction of Methemoglobin in Vivo. Nature 1964, 202, 607–608. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | At Time of Anesthesia Induction | After Supplementation * | After Methylene Blue ** | After 2 Weeks *** | After ~4 Months *** | After ~6 Months *** | Reference Interval **** |
|---|---|---|---|---|---|---|---|
| Hematocrit, % | 29 | 33 | 46 | 45 | 33–45 | ||
| Hemoglobin, g/L | 116 | 121 | 160 | 113–155 | |||
| MCV, fL | 36 | 40 | 39 | 40–48 | |||
| MCHC, g/dL | 40 | 37 | 35 | 33–36 | |||
| Reticulocytes, 106/L | 14,076 | <45,000 | |||||
| O2-Hb,% | 23.7 | 64.5 | 91.1 | 47.1 | 63.4 | 60.4 | >90% # |
| CO-Hb, % | 4.4 | 2.7 | 3.6 | 1.7 | 2.8 | 2.2 | <1% # |
| metHb, % | 49.6 | 39.9 | 3.1 | 32.4 | 33.8 | 35.2 | 0.3–4.0 |
| GSH, µmol/g Hb | 11.9 | 10.1 | 9.4 | 4.8 ± 0.9 | |||
| CYB5R activity, | 0.27 | 0.46 ± 0.10 | 2.07 ± 0.81 | ||||
| IU/g Hb | 13.0 | 22.2 | 100 | ||||
| % |
| Parameters # | Control Cats (n = 5) | Affected Cat |
|---|---|---|
| EImin, AU | 0.131 ± 0.033 | 0.142 ± 0.008 |
| Omin, mOsm | 187.8 ± 5.6 | 186.3 ± 2.5 |
| EImax, AU | 0.517 ± 0.052 | 0.547 ± 0.002 ** |
| OEImax, mOsm | 359.4 ± 18.9 | 355.3 ± 2.5 |
| EIhyper, AU | 0.259 ± 0.027 | 0.274 ± 0.001 ** |
| Ohyper, mOsm | 484.6 ± 7.6 | 462.0 ± 2.0 ** |
| Area, AU | 124.9 ± 14.2 | 118.4 ± 0.25 |
| Na, mmol/kg dw | 269.9 ± 13.7 | 264.8 |
| K+, mmol/kg dw | 13.7 ± 2.9 | 12.6 |
| Filtering Step | Heterozygous Variants | Homozygous Variants |
|---|---|---|
| Variants in the case genome | 6,189,794 | 6,101,321 |
| Private variants | 90,816 | 23,992 |
| Variants with SnpEff impact high, moderate or low | 951 | 252 |
| Private variants in CYB5R3 | 0 | 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jenni, S.; Ludwig-Peisker, O.; Jagannathan, V.; Lapsina, S.; Stirn, M.; Hofmann-Lehmann, R.; Bogdanov, N.; Schetle, N.; Giger, U.; Leeb, T.; et al. Methemoglobinemia, Increased Deformability and Reduced Membrane Stability of Red Blood Cells in a Cat with a CYB5R3 Splice Defect. Cells 2023, 12, 991. https://doi.org/10.3390/cells12070991
Jenni S, Ludwig-Peisker O, Jagannathan V, Lapsina S, Stirn M, Hofmann-Lehmann R, Bogdanov N, Schetle N, Giger U, Leeb T, et al. Methemoglobinemia, Increased Deformability and Reduced Membrane Stability of Red Blood Cells in a Cat with a CYB5R3 Splice Defect. Cells. 2023; 12(7):991. https://doi.org/10.3390/cells12070991
Chicago/Turabian StyleJenni, Sophia, Odette Ludwig-Peisker, Vidhya Jagannathan, Sandra Lapsina, Martina Stirn, Regina Hofmann-Lehmann, Nikolay Bogdanov, Nelli Schetle, Urs Giger, Tosso Leeb, and et al. 2023. "Methemoglobinemia, Increased Deformability and Reduced Membrane Stability of Red Blood Cells in a Cat with a CYB5R3 Splice Defect" Cells 12, no. 7: 991. https://doi.org/10.3390/cells12070991