
Analysis of Immunophenotypic Changes during Ex Vivo Human Erythropoiesis and Its Application in the Study of Normal and Defective Erythropoiesis
 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Purification and Culture of CD34+ Cells
2.2. Benzidine Staining
2.3. Colony-Forming Assays
2.4. May–Grunwald–Giemsa Staining
2.5. Hoechst and SytoTM 16 Staining
2.6. FACS Analysis
2.7. Lentivirus Preparation and Cell Transduction
2.8. RNA Extraction and qPCR Analysis
2.9. Statistical Data Analysis
3. Results
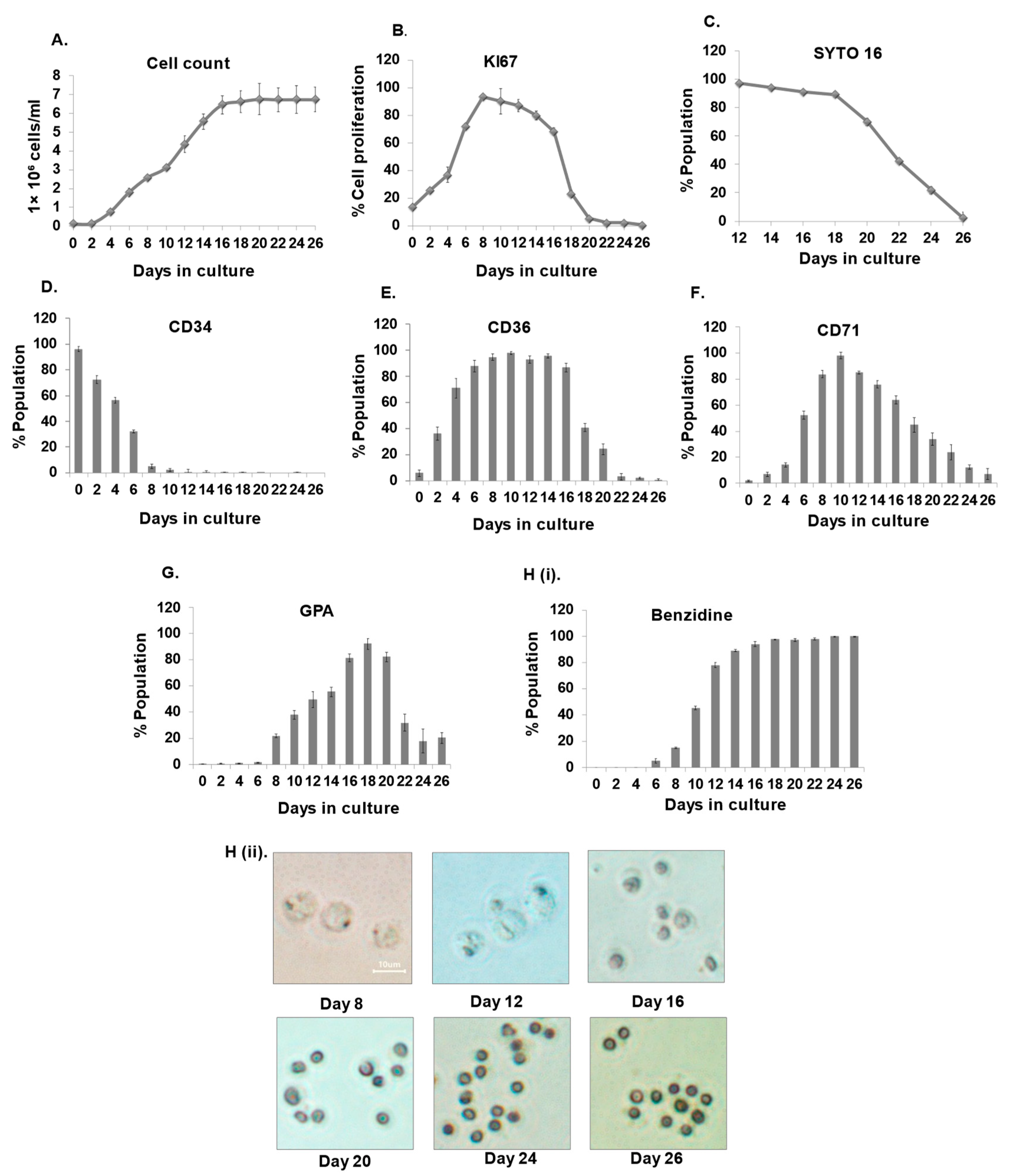
3.1. Establishment of Erythroid Differentiation
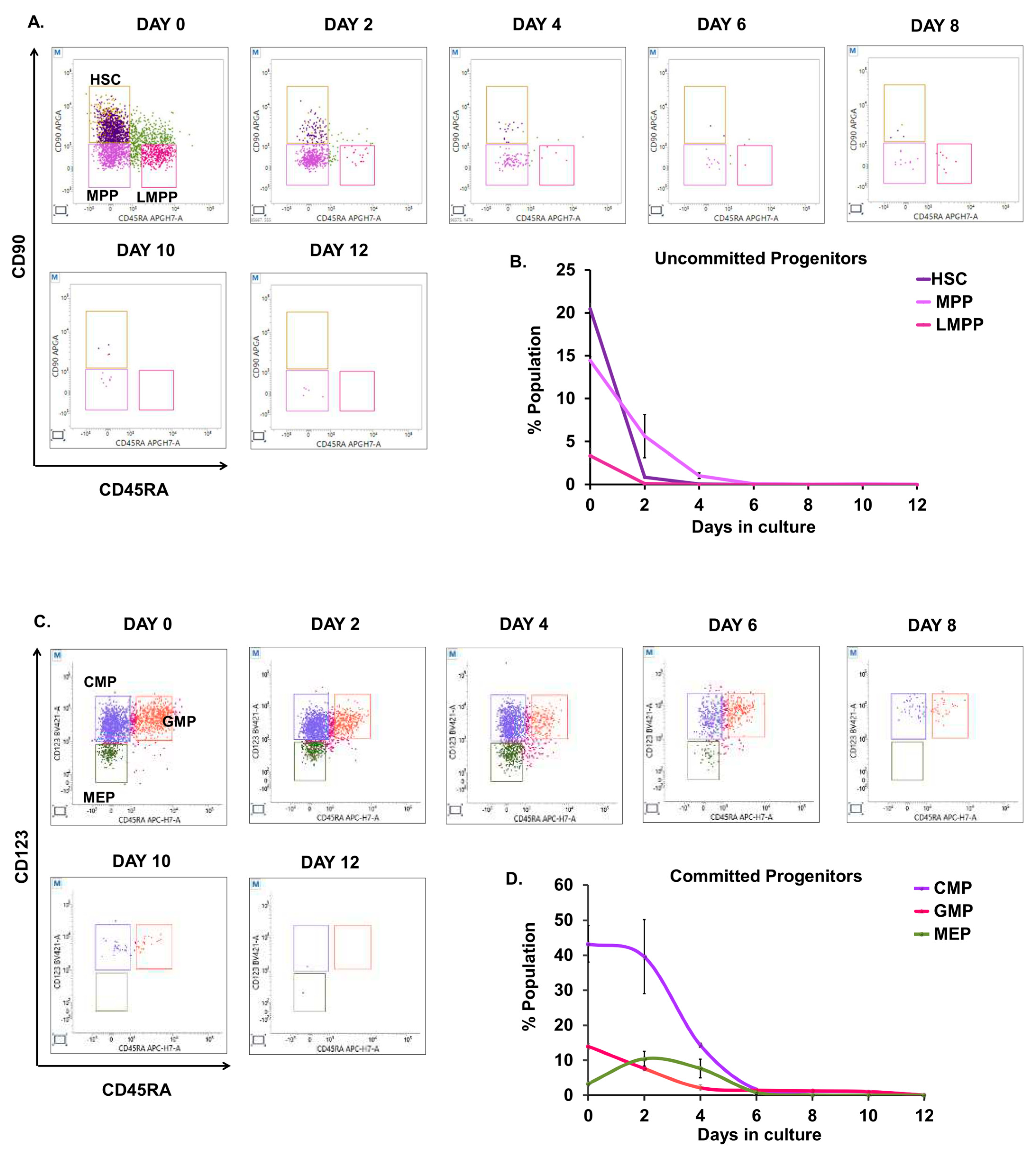
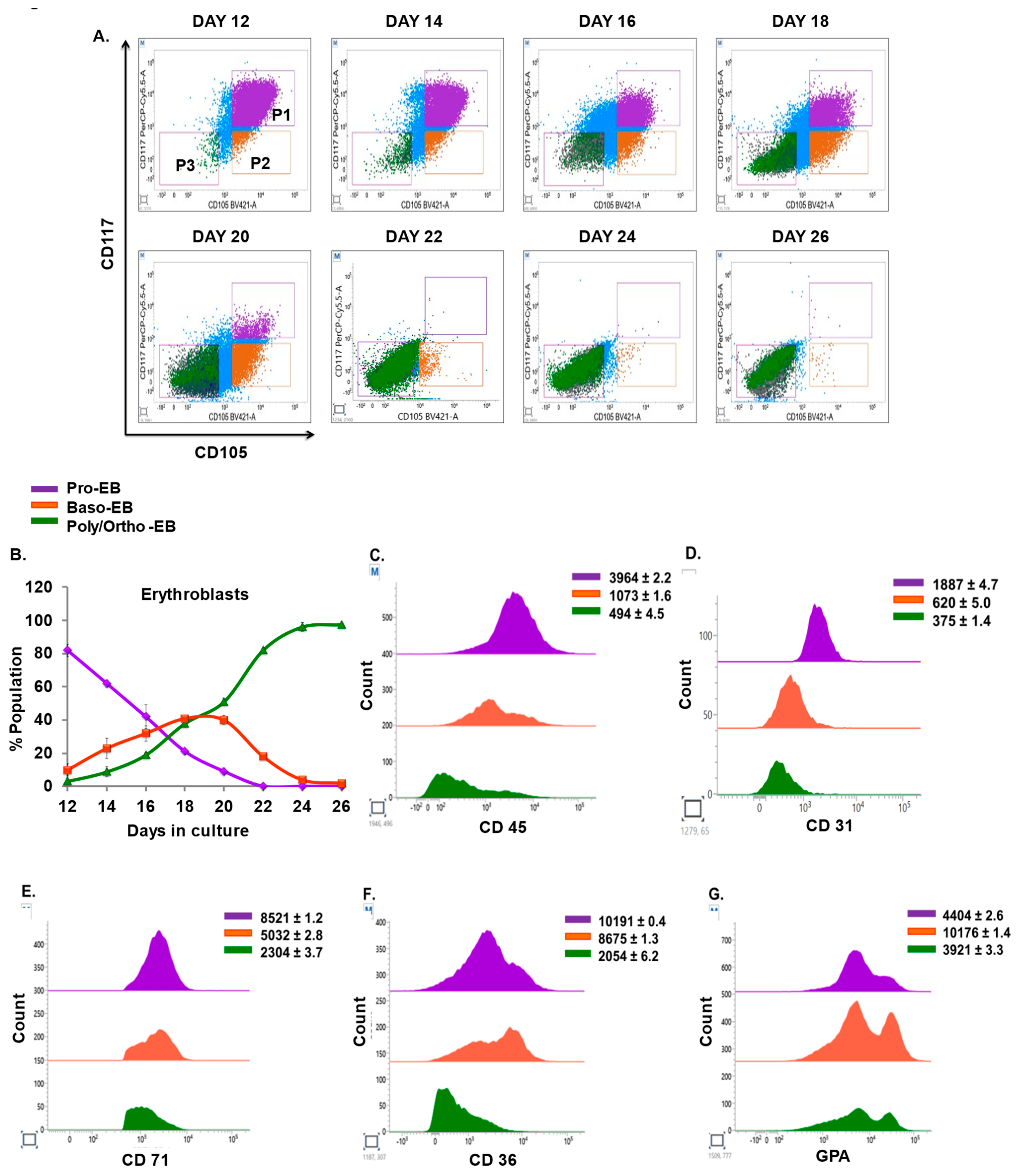
3.2. Kinetics of Committed and Uncommitted Progenitors during Erythroid Differentiation
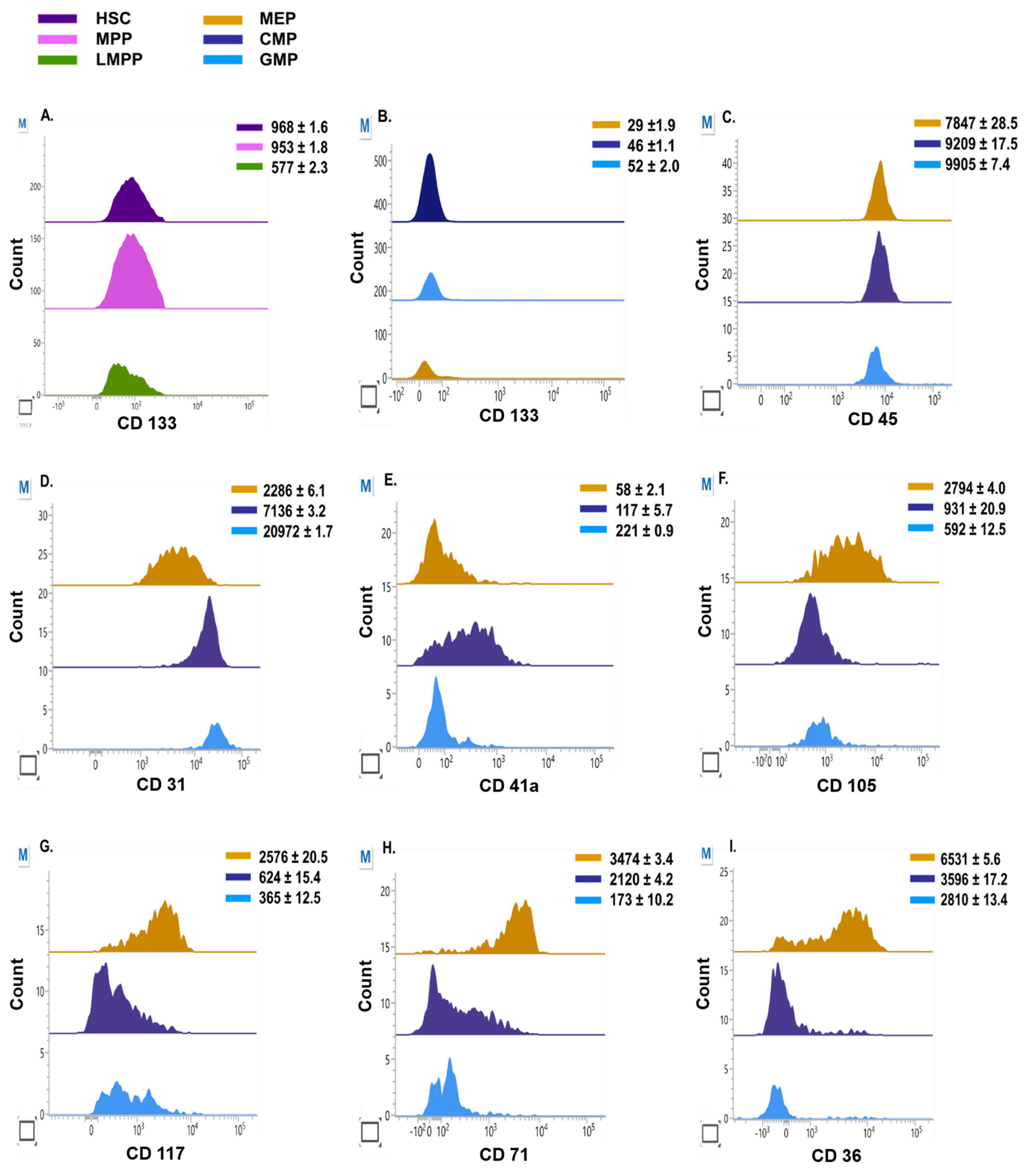
3.3. Lineage Commitment of Hematopoietic Precursors during Erythroid Differentiation
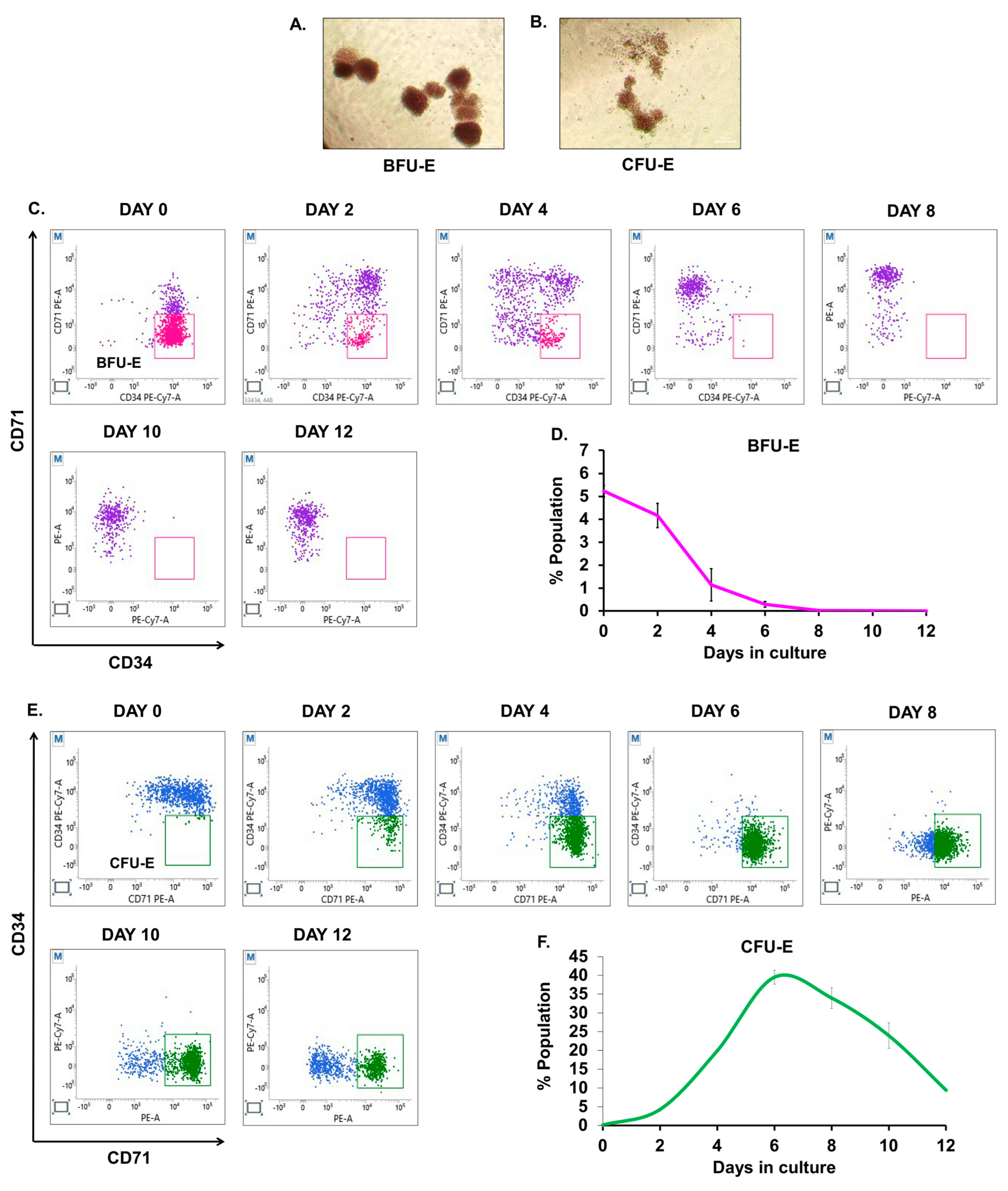
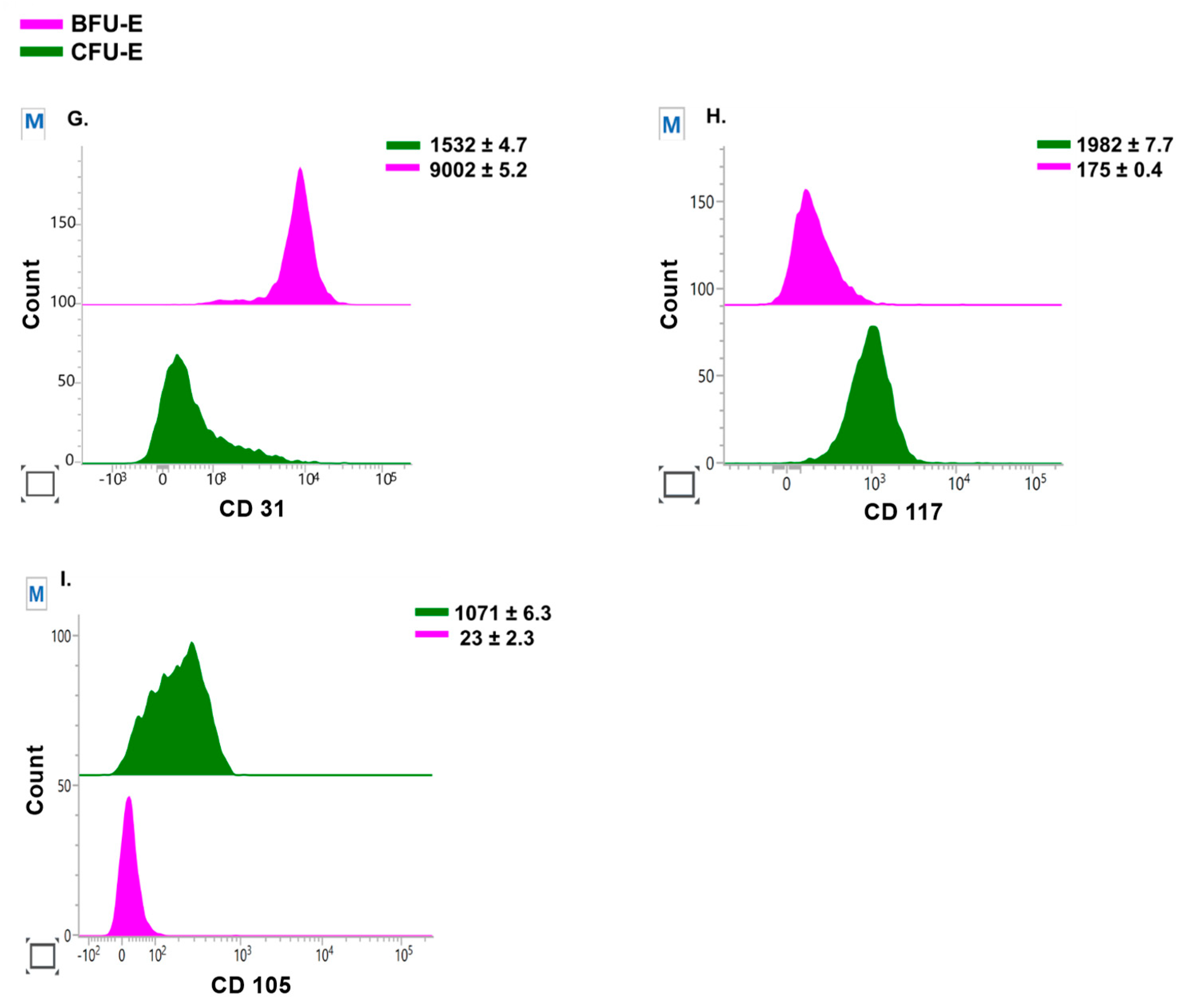
3.4. Quantitative Analysis of BFU-E and CFU-E during Ex Vivo Culture
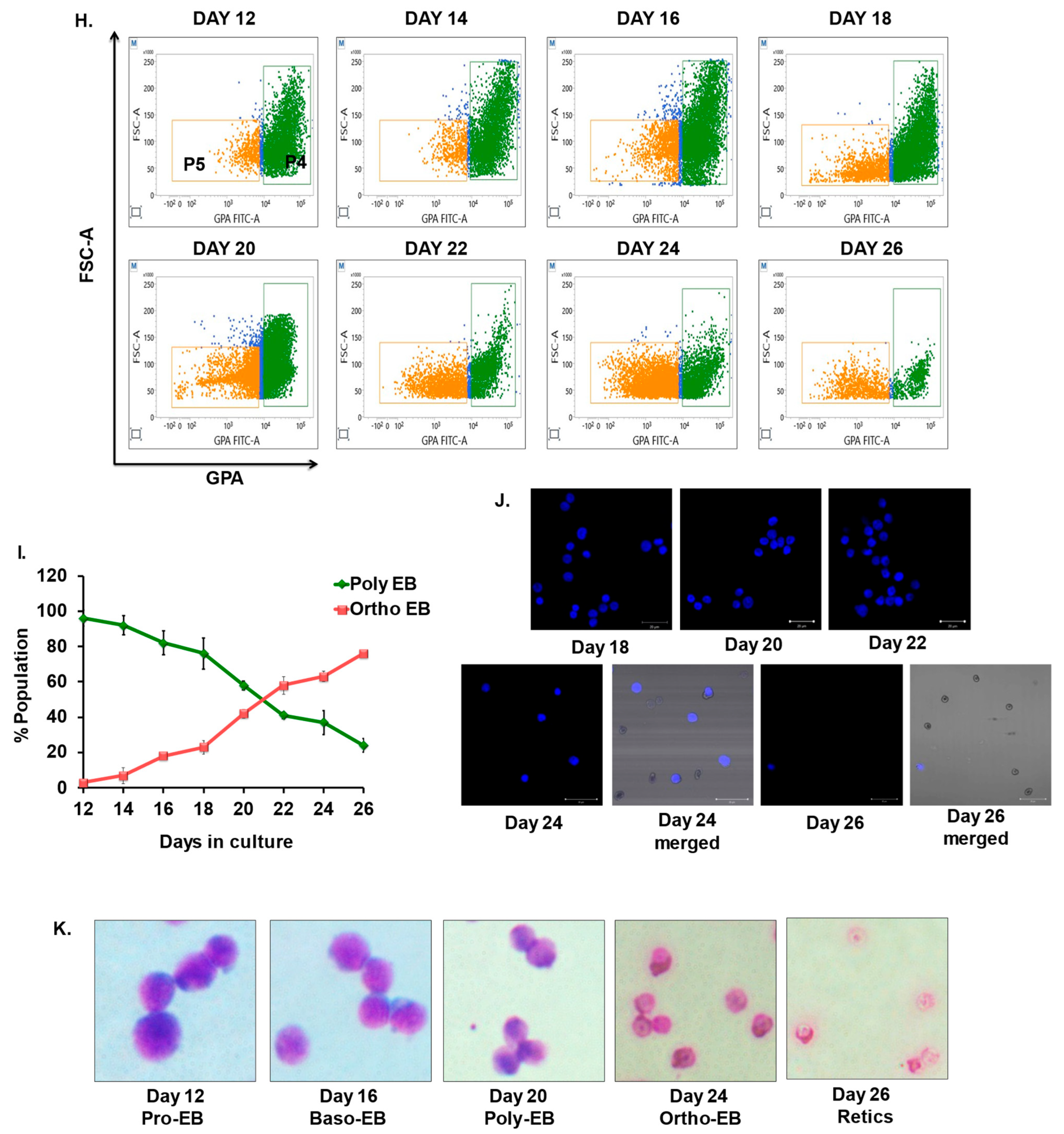
3.5. Kinetics and Characterization of Terminal Erythroblasts
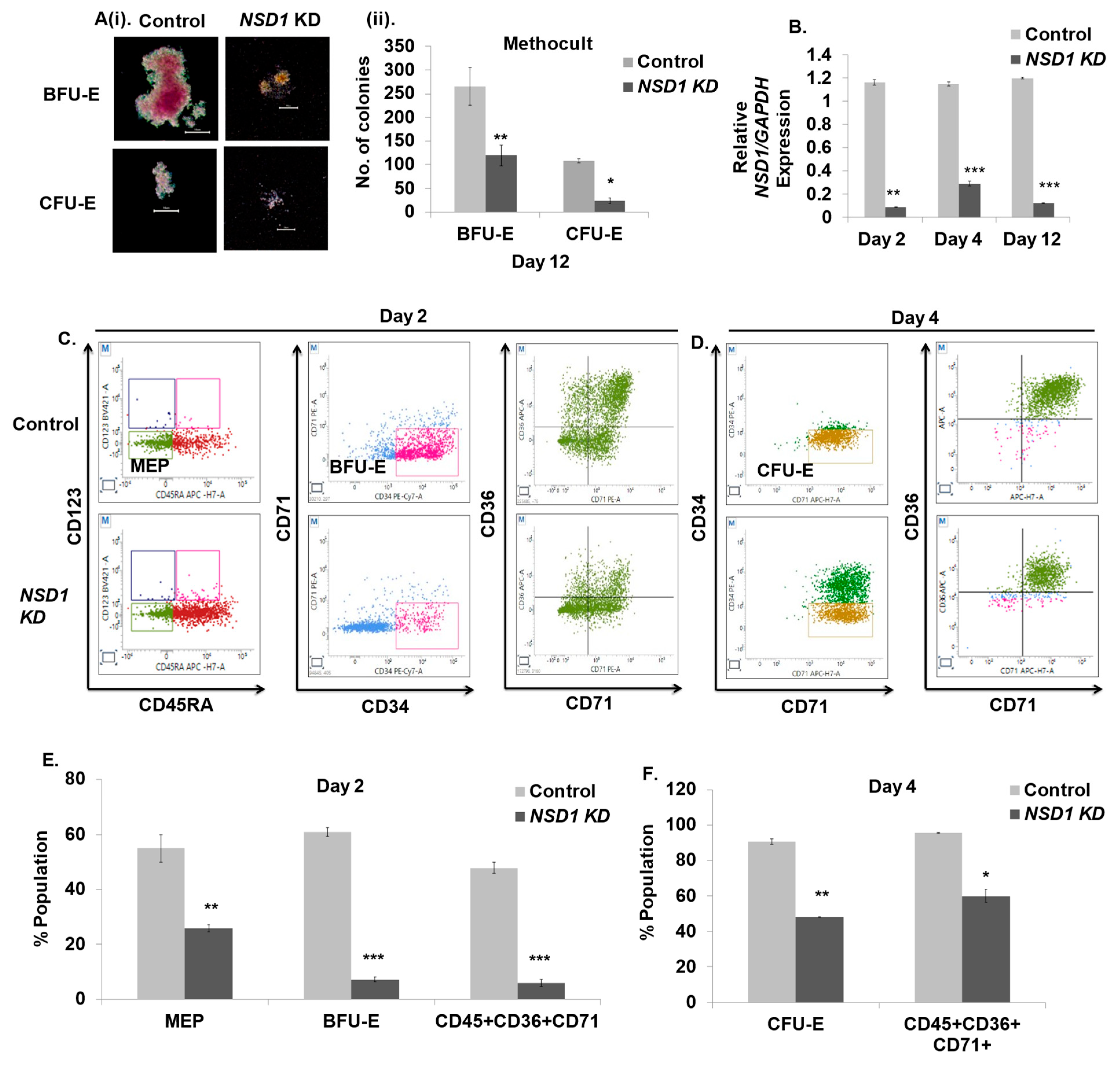
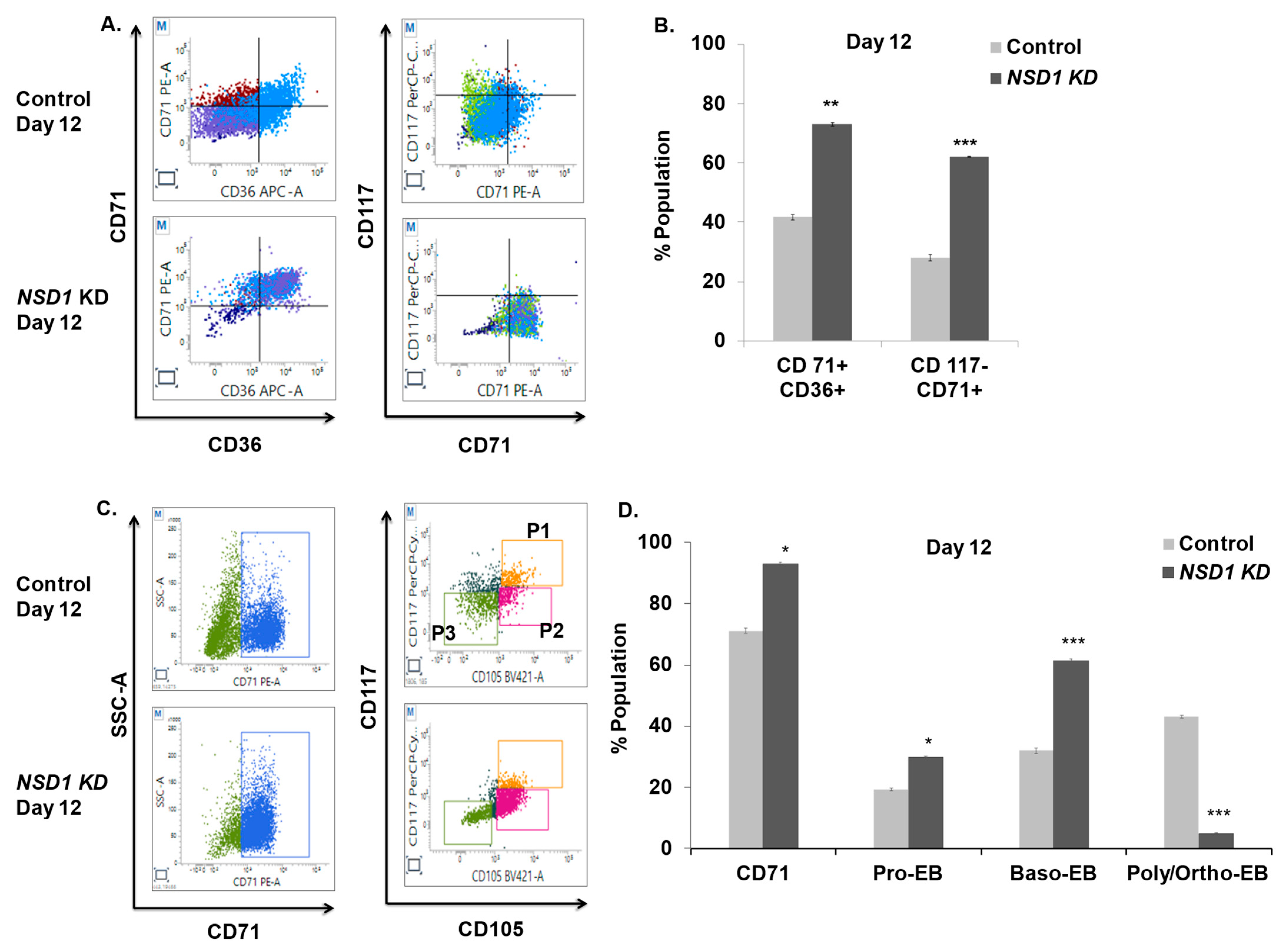
3.6. NSD1 Knockdown Alters Early Human Erythroid Progenitors
3.7. NSD1 Knockdown Blocks Terminal Differentiation in Maturing Erythroblasts
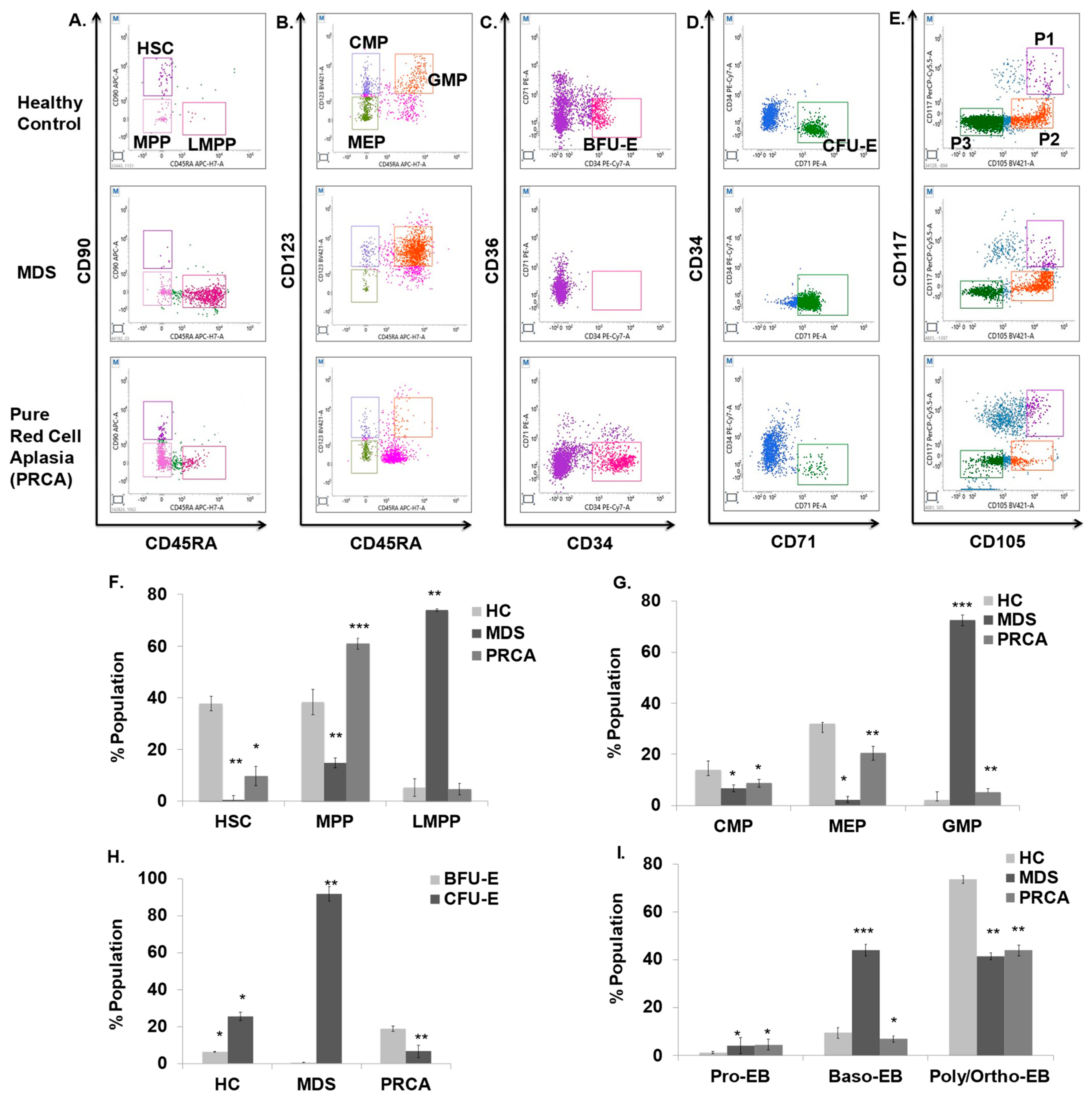
3.8. MDS and PRCA Patients Display Altered Erythropoiesis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Palis, J. Primitive and definitive erythropoiesis in mammals. Front. Physiol. 2014, 5, 3. [Google Scholar] [CrossRef]
- Seita, J.; Weissman, I.L. Hematopoietic Stem Cell: Self-Renewal versus Differentiation. Wiley Interdiscip. Rev. Syst. Biol. Med. 2010, 2, 640–653. [Google Scholar] [CrossRef] [PubMed]
- Cheng, H.; Zheng, Z.; Cheng, T. New Paradigms on Hematopoietic Stem Cell Differentiation. Protein Cell 2020, 11, 34–44. [Google Scholar] [CrossRef]
- Dulmovits, B.M.; Hom, J.; Narla, A.; Mohandas, N.; Blanc, L. Characterization, Regulation, and Targeting of Erythroid Progenitors in Normal and Disordered Human Erythropoiesis. Curr. Opin. Hematol. 2017, 24, 159–166. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Zhang, J.; Ginzburg, Y.; Li, H.; Xue, F.; De Franceschi, L.; Chasis, J.A.; Mohandas, N.; An, X. Quantitative Analysis of Murine Terminal Erythroid Differentiation in Vivo: Novel Method to Study Normal and Disordered Erythropoiesis. E-Blood 2013, 121, e43–e49. [Google Scholar] [CrossRef] [PubMed]
- Giarratana, M.C.; Kobari, L.; Lapillonne, H.; Chalmers, D.; Kiger, L.; Cynober, T.; Marden, M.C.; Wajcman, H.; Douay, L. Ex Vivo Generation of Fully Mature Human Red Blood Cells from Hematopoietic Stem Cells. Nat. Biotechnol. 2005, 23, 69–74. [Google Scholar] [CrossRef] [PubMed]
- Palii, C.G.; Pasha, R.; Brand, M. Lentiviral-Mediated Knockdown during Ex Vivo Erythropoiesis of Human Hematopoietic Stem Cells. J. Vis. Exp. 2011, 16, 2813. [Google Scholar] [CrossRef]
- Flygare, J.; Estrada, V.R.; Shin, C.; Gupta, S.; Lodish, H.F. HIF1α Synergizes with Glucocorticoids to Promote BFU-E Progenitor Self-Renewal. Blood 2011, 117, 3435–3444. [Google Scholar] [CrossRef]
- Terszowski, G.; Waskow, C.; Conradt, P.; Lenze, D.; Koenigsmann, J.; Carstanjen, D.; Horak, I.; Rodewald, H.R. Prospective Isolation and Global Gene Expression Analysis of the Erythrocyte Colony-Forming Unit (CFU-E). Blood 2005, 105, 1937–1945. [Google Scholar] [CrossRef]
- Koulnis, M.; Pop, R.; Porpiglia, E.; Shearstone, J.R.; Hidalgo, D.; Socolovsky, M. Identification and Analysis of Mouse Erythroid Progenitors Using the CD71/TER119 Flow-Cytometric Assay. J. Vis. Exp. 2011, 5, 2809. [Google Scholar] [CrossRef]
- Chen, K.; Liu, J.; Heck, S.; Chasis, J.A.; An, X.; Mohandas, N. Resolving the Distinct Stages in Erythroid Differentiation Based on Dynamic Changes in Membrane Protein Expression during Erythropoiesis. Biol. Sci. 2009, 106, 17413–17418. [Google Scholar] [CrossRef]
- Gillespie, M.A.; Palii, C.G.; Sanchez-Taltavull, D.; Shannon, P.; Longabaugh, W.J.R.; Downes, D.J.; Sivaraman, K.; Espinoza, H.M.; Hughes, J.R.; Price, N.D.; et al. Absolute Quantification of Transcription Factors Reveals Principles of Gene Regulation in Erythropoiesis. Mol. Cell 2020, 78, 960–974.e11. [Google Scholar] [CrossRef]
- Gautier, E.F.; Ducamp, S.; Leduc, M.; Salnot, V.; Guillonneau, F.; Dussiot, M.; Hale, J.; Giarratana, M.C.; Raimbault, A.; Douay, L.; et al. Comprehensive Proteomic Analysis of Human Erythropoiesis. Cell Rep. 2016, 16, 1470–1484. [Google Scholar] [CrossRef]
- Yan, H.; Ali, A.; Blanc, L.; Narla, A.; Lane, J.M.; Gao, E.; Papoin, J.; Hale, J.; Hillyer, C.D.; Taylor, N.; et al. Comprehensive Phenotyping of Erythropoiesis in Human Bone Marrow: Evaluation of Normal and Ineffective Erythropoiesis. Am. J. Hematol. 2021, 96, 1064–1076. [Google Scholar] [CrossRef]
- Li, J.; Hale, J.; Bhagia, P.; Xue, F.; Chen, L.; Jaffray, J.; Yan, H.; Lane, J.; Gallagher, P.G.; Mohandas, N.; et al. Isolation and Transcriptome Analyses of Human Erythroid Progenitors: BFU-E and CFU-E. Blood 2014, 124, 3636–3645. [Google Scholar] [CrossRef]
- Hu, J.; Liu, J.; Xue, F.; Halverson, G.; Reid, M.; Guo, A.; Chen, L.; Raza, A.; Galili, N.; Jaffray, J.; et al. Isolation and Functional Characterization of Human Erythroblasts at Distinct Stages: Implications for Understanding of Normal and Disordered Erythropoiesis in Vivo Key Points. Blood 2013, 121, 3245–3253. [Google Scholar] [CrossRef]
- Fajtova, M.; Kovarikova, A.; Svec, P.; Kankuri, E.; Sedlak, J. Immunophenotypic Profile of Nucleated Erythroid Progenitors during Maturation in Regenerating Bone Marrow. Leuk. Lymphoma 2013, 54, 2523–2530. [Google Scholar] [CrossRef]
- Malik, P.; Fisher, T.C.; Barsky, L.L.W.; Zeng, L.; Izadi, P.; Hiti, A.L.; Weinberg, K.I.; Coates, T.D.; Meiselman, H.J.; Kohn, D.B. Rapid Communication. An In Vitro Model of Human Red Blood Cell Production from Hematopoietic Progenitor Cells. Blood 1998, 91, 2664–2671. [Google Scholar] [CrossRef]
- Monaco, M.C.G.; Maric, D.; Salvucci, O.; Passeri, C.A.L.; Accorsi, P.; Major, E.O.; Berardi, A.C. Identification of Circulating CD31+CD45+ Cell Populations with the Potential to Differentiate into Erythroid Cells. Stem Cell Res. Ther. 2021, 12, 236. [Google Scholar] [CrossRef]
- Leonards, K.; Almosailleakh, M.; Tauchmann, S.; Bagger, F.O.; Thirant, C.; Juge, S.; Bock, T.; Méreau, H.; Bezerra, M.F.; Tzankov, A.; et al. Nuclear Interacting SET Domain Protein 1 Inactivation Impairs GATA1-Regulated Erythroid Differentiation and Causes Erythroleukemia. Nat. Commun. 2020, 11, 2870. [Google Scholar] [CrossRef]
- Ogawa, S. Genetics of MDS. Blood 2019, 133, 1049–1059. [Google Scholar] [CrossRef]
- Means, R.T. Pure Red Cell Aplasia. Blood 2016, 128, 2504–2509. [Google Scholar] [CrossRef]
- Čikoš, Š.; Bukovská, A.; Koppel, J. Relative Quantification of MRNA: Comparison of Methods Currently Used for Real-Time PCR Data Analysis. BMC Mol. Biol. 2007, 8, 113. [Google Scholar] [CrossRef]
- Brott, D.A.; Maher, R.J.; Parrish, C.R.; Richardson, R.J.; Smith, A.K. Flow Cytometric Characterization of Perfused Human Bone Marrow Cultures: Identification of the Major Cell Lineages and Correlation with the CFU-GM Assay. Cytom. Part A 2003, 53, 22–27. [Google Scholar] [CrossRef]
- Boyer, S.W.; Rajendiran, S.; Beaudin, A.E.; Smith-Berdan, S.; Muthuswamy, P.K.; Perez-Cunningham, J.; Martin, E.W.; Cheung, C.; Tsang, H.; Landon, M.; et al. Clonal and Quantitative In Vivo Assessment of Hematopoietic Stem Cell Differentiation Reveals Strong Erythroid Potential of Multipotent Cells. Stem Cell Rep. 2019, 12, 801–815. [Google Scholar] [CrossRef]
- Sun, J.; Ramos, A.; Chapman, B.; Johnnidis, J.B.; Le, L.; Ho, Y.J.; Klein, A.; Hofmann, O.; Camargo, F.D. Clonal Dynamics of Native Haematopoiesis. Nature 2014, 514, 322–327. [Google Scholar] [CrossRef]
- Busch, K.; Klapproth, K.; Barile, M.; Flossdorf, M.; Holland-Letz, T.; Schlenner, S.M.; Reth, M.; Höfer, T.; Rodewald, H.R. Fundamental Properties of Unperturbed Haematopoiesis from Stem Cells in Vivo. Nature 2015, 518, 542–546. [Google Scholar] [CrossRef]
- Takahashi, M.; Matsuoka, Y.; Sumide, K.; Nakatsuka, R.; Fujioka, T.; Kohno, H.; Sasaki, Y.; Matsui, K.; Asano, H.; Kaneko, K.; et al. CD133 Is a Positive Marker for a Distinct Class of Primitive Human Cord Blood-Derived CD34-Negative Hematopoietic Stem Cells. Leukemia 2014, 28, 1308–1315. [Google Scholar] [CrossRef]
- Cimato, T.R.; Conway, A.; Nichols, J.; Wallace, P.K. CD133 Expression in Circulating Hematopoietic Progenitor Cells. Cytom. B Clin. Cytom. 2019, 96, 39–45. [Google Scholar] [CrossRef]
- Mello, F.V.; Land, M.G.P.; Costa, E.S.; Teodósio, C.; Sanchez, M.L.; Bárcena, P.; Peres, R.T.; Pedreira, C.E.; Alves, L.R.; Orfao, A. Maturation-Associated Gene Expression Profiles during Normal Human Bone Marrow Erythropoiesis. Cell Death Discov. 2019, 5, 69. [Google Scholar] [CrossRef]
- Westers, T.M.; Cremers, E.M.P.; Oelschlaegel, U.; Johansson, U.; Bettelheim, P.; Matarraz, S.; Orfao, A.; Moshaver, B.; Brodersen, L.E.; Loken, M.R.; et al. Immunophenotypic Analysis of Erythroid Dysplasia in Myelodysplastic Syndromes. A Report from the IMDSFlow Working Group. Haematologica 2017, 102, 308–319. [Google Scholar] [CrossRef]
- Kim, H.; Cho, H.J.; Kim, S.W.; Liu, B.; Choi, Y.J.; Lee, J.; Sohn, Y.D.; Lee, M.Y.; Houge, M.A.; Yoon, Y.S. CD31+ Cells Represent Highly Angiogenic and Vasculogenic Cells in Bone Marrow: Novel Role of Nonendothelial CD31+ Cells in Neovascularization and Their Therapeutic Effects on Ischemic Vascular Disease. Circ. Res. 2010, 107, 602–614. [Google Scholar] [CrossRef]
- Yang, C.T.; French, A.; Goh, P.A.; Pagnamenta, A.; Mettananda, S.; Taylor, J.; Knight, S.; Nathwani, A.; Roberts, D.J.; Watt, S.M.; et al. Human Induced Pluripotent Stem Cell Derived Erythroblasts Can Undergo Definitive Erythropoiesis and Co-Express Gamma and Beta Globins. Br. J. Haematol. 2014, 166, 435–448. [Google Scholar] [CrossRef]
- Zhou, F.; Li, X.; Wang, W.; Zhu, P.; Zhou, J.; He, W.; Ding, M.; Xiong, F.; Zheng, X.; Li, Z.; et al. Tracing Haematopoietic Stem Cell Formation at Single-Cell Resolution. Nature 2016, 533, 487–492. [Google Scholar] [CrossRef]
- MacHherndl-Spandl, S.; Suessner, S.; Danzer, M.; Proell, J.; Gabriel, C.; Lauf, J.; Sylie, R.; Klein, H.U.; Béné, M.C.; Weltermann, A.; et al. Molecular Pathways of Early CD105-Positive Erythroid Cells as Compared with CD34-Positive Common Precursor Cells by Flow Cytometric Cell-Sorting and Gene Expression Profiling. Blood Cancer J. 2013, 3, e100. [Google Scholar] [CrossRef]
- Meurer, S.K.; Weiskirchen, R. Endoglin: An ‘Accessory’ Receptor Regulating Blood Cell Development and Inflammation. Int. J. Mol. Sci. 2020, 21, 9247. [Google Scholar] [CrossRef]
- Sperling, A.S.; Gibson, C.J.; Ebert, B.L. The Genetics of Myelodysplastic Syndrome: From Clonal Haematopoiesis to Secondary Leukaemia. Nat. Rev. Cancer 2017, 17, 5–19. [Google Scholar] [CrossRef]
- Fujishima, N.; Kohmaru, J.; Koyota, S.; Kuba, K.; Saga, T.; Omokawa, A.; Moritoki, Y.; Ueki, S.; Ishida, F.; Nakao, S.; et al. Clonal Hematopoiesis in Adult Pure Red Cell Aplasia. Sci. Rep. 2021, 11, 2253. [Google Scholar] [CrossRef]
- Cremers, E.M.P.; Westers, T.M.; Alhan, C.; Cali, C.; Visser-Wisselaar, H.A.; Chitu, D.A.; van der Velden, V.H.J.; te Marvelde, J.G.; Klein, S.K.; Muus, P.; et al. Implementation of Erythroid Lineage Analysis by Flow Cytometry in Diagnostic Models for Myelodysplastic Syndromes. Haematologica 2017, 102, 320–326. [Google Scholar] [CrossRef]
- Xu, F.; Wu, L.; He, Q.; Zhang, Z.; Chang, C.; Li, X. Immunophenotypic Analysis of Erythroid Dysplasia and Its Diagnostic Application in Myelodysplastic Syndromes. Intern. Med. J. 2012, 42, 401–411. [Google Scholar] [CrossRef]
- Jerez, J.; Ocqueteau, M. Immunophenotype of Erythroid Precursors in Patient with Pure Red Cell Aplasia (PRCA): Utility of Analysis of Erythroid Maturation. Ann. Hematol. Oncol. 2021, 8, 346. [Google Scholar]
- Huang, Z.; Dore, L.C.; Li, Z.; Orkin, S.H.; Feng, G.; Lin, S.; Crispino, J.D. GATA-2 Reinforces Megakaryocyte Development in the Absence of GATA-1. Mol. Cell. Biol. 2009, 29, 5168–5180. [Google Scholar] [CrossRef] [PubMed]
- Notta, F.; Zandi, S.; Takayama, N.; Dobson, S.; Gan, O.I.; Wilson, G.; Kaufmann, K.B.; McLeod, J.; Laurenti, E.; Dunant, C.F.; et al. Distinct Routes of Lineage Development Reshape the Human Blood Hierarchy across Ontogeny. Science 2016, 351, aab2116. [Google Scholar] [CrossRef] [PubMed]
- Pronk, C.J.H.; Rossi, D.J.; Månsson, R.; Attema, J.L.; Norddahl, G.L.; Chan, C.K.F.; Sigvardsson, M.; Weissman, I.L.; Bryder, D. Elucidation of the Phenotypic, Functional, and Molecular Topography of a Myeloerythroid Progenitor Cell Hierarchy. Cell Stem Cell 2007, 1, 428–442. [Google Scholar] [CrossRef]
- Mori, Y.; Chen, J.Y.; Pluvinage, J.V.; Seita, J.; Weissman, I.L. Prospective Isolation of Human Erythroid Lineage-Committed Progenitors. Proc. Natl. Acad. Sci. USA 2015, 112, 9638–9643. [Google Scholar] [CrossRef]
- Morishita, M.; di Luccio, E. Cancers and the NSD Family of Histone Lysine Methyltransferases. Biochim. Biophys. Acta 2011, 1816, 158–163. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibody | Source | Catalogue Number |
|---|---|---|
| APC-R700 Mouse anti-Human CD45 | BD HorizonTM | 566042 |
| PE Mouse anti-Human CD117 | BD BiosciencesTM | 555714 |
| Brilliant Violet 605TM Mouse anti-Human CD38 | BioLegendTM | 356642 |
| PE-Cy7TM Mouse anti-Human CD34 | BD BiosciencesTM | 348791 |
| Brilliant Violet 510TM anti-human Lineage Cocktail (CD3, CD14, CD16, CD19, CD20, CD56) | BioLegendTM | 348807 |
| APC Mouse anti-Human CD90 | BD BiosciencesTM | 559869 |
| APC-H7 Mouse anti-Human CD45RA | BD BiosciencesTM | 560674 |
| Brilliant Violet 421TM Mouse anti-Human CD123 | BioLegendTM | 306018 |
| Antibody | Source | Catalogue Number |
|---|---|---|
| APC-R700 Mouse anti-Human CD45 | BD HorizonTM | 566042 |
| FITC anti-Human Lineage cocktail1 (Lin1) (CD3, CD14, CD16, CD19, CD20, CD56) | BD FastImmuneTM | 340546 |
| CD235a (Glycophorin A) Mouse anti-Human Super bright 600 | eBioscienceTM | 63-9987-41 |
| Brilliant Violet 421TM Mouse anti-Human CD123 | BioLegendTM | 306018 |
| APC Mouse anti-Human CD36 | BD PharmingenTM | 550956 |
| PE-Cy7TM Mouse anti-Human CD34 | BD BiosciencesTM | 348791 |
| PE Mouse anti-Human CD 71 | BD PharmingenTM | 555537 |
| Antibody | Source | Catalogue Number |
|---|---|---|
| PE Mouse anti-Human CD 71 | BD PharmingenTM | 555537 |
| PE-Cyanine 5 Mouse anti-Human CD 117 | eBioscienceTM | 15-1178-41 |
| Brilliant violet 421TM anti-Human CD105 | BioLegendTM | 323219 |
| FITC Mouse anti-Human CD235a | BD PharmingenTM | 559943 |
| Oligos | Sequence |
|---|---|
| hTRC Scrambled | Forward 5′-CCGGCCGCAGGTATGCACGCGTCTCGAGACGCGTGCATACCTGCGGTTTTTG-3′ Reverse 5′-GGCGTCCATACGTGCGCAGAGCTCTGCGCACGTATGGACGCCAAAAACTTAA-3′ |
| hNSD1 CDS | Forward 5′-CCGGTCCAGTGAGAACTCGTTAATACTCGAGTATTAACGAGTTCTCACTGGATTTTTG-3′ Reverse 5′-AATTCAAAAATCCAGTGAGAACTCGTTAATACTCGAGTATTAACGAGTTCTCACTGGA-3′ |
| hNSD1 UTR | Forward 5′-CCGGGTGCTAATTTCACGGTATAAACTCGAGTTTATACCGTGAAATTAGCACTTTTTG-3′ Reverse 5′-AATTCAAAAAGTGCTAATTTCACGGTATAAACTCGAGTTTATACCGTGAAATTAGCAC-3′ |
| Target | Sequence |
|---|---|
| hNSD1 | For: 5′AGG TAC AGG AGC AGG TGC ACA-3′ |
| Rev: 5′AGC ACT AGA TCG ACC TCG GGC-3′ | |
| hGAPDH | For: 5′GTGGTCTCCCTGACTTTCAACAGC-3′ |
| Rev: 5′A TGAGGTCCACCTGCTTGCTG-3′ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Katiyar, S.; Shah, A.; Rahman, K.; Tripathy, N.K.; Kashyap, R.; Nityanand, S.; Chaturvedi, C.P. Analysis of Immunophenotypic Changes during Ex Vivo Human Erythropoiesis and Its Application in the Study of Normal and Defective Erythropoiesis. Cells 2023, 12, 1303. https://doi.org/10.3390/cells12091303
Katiyar S, Shah A, Rahman K, Tripathy NK, Kashyap R, Nityanand S, Chaturvedi CP. Analysis of Immunophenotypic Changes during Ex Vivo Human Erythropoiesis and Its Application in the Study of Normal and Defective Erythropoiesis. Cells. 2023; 12(9):1303. https://doi.org/10.3390/cells12091303
Chicago/Turabian StyleKatiyar, Shobhita, Arunim Shah, Khaliqur Rahman, Naresh Kumar Tripathy, Rajesh Kashyap, Soniya Nityanand, and Chandra Prakash Chaturvedi. 2023. "Analysis of Immunophenotypic Changes during Ex Vivo Human Erythropoiesis and Its Application in the Study of Normal and Defective Erythropoiesis" Cells 12, no. 9: 1303. https://doi.org/10.3390/cells12091303