
The Effect of the Donor’s and Recipient’s Sex on Red Blood Cells Evaluated Using Transfusion Simulations

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
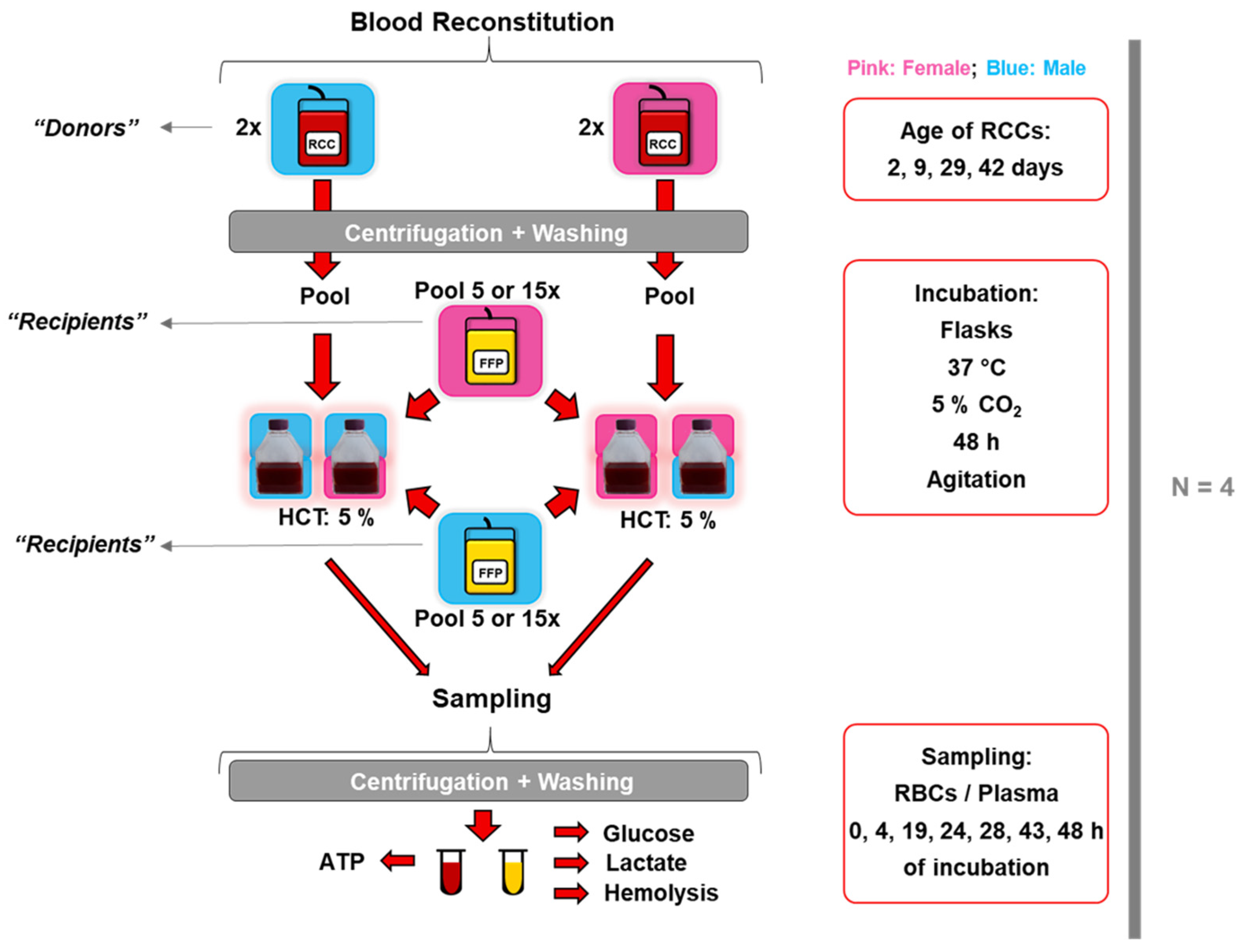
2. Materials and Methods
2.1. Blood Products
2.2. In Vitro Transfusional Models
2.3. Sampling
2.4. Standard Blood Parameters
2.5. Morphology
2.6. Intracellular ATP Quantification
2.7. Extracellular Glucose and Lactate Quantification
2.8. Hemolysis Rate
2.9. Data Treatment and Statistics
3. Results
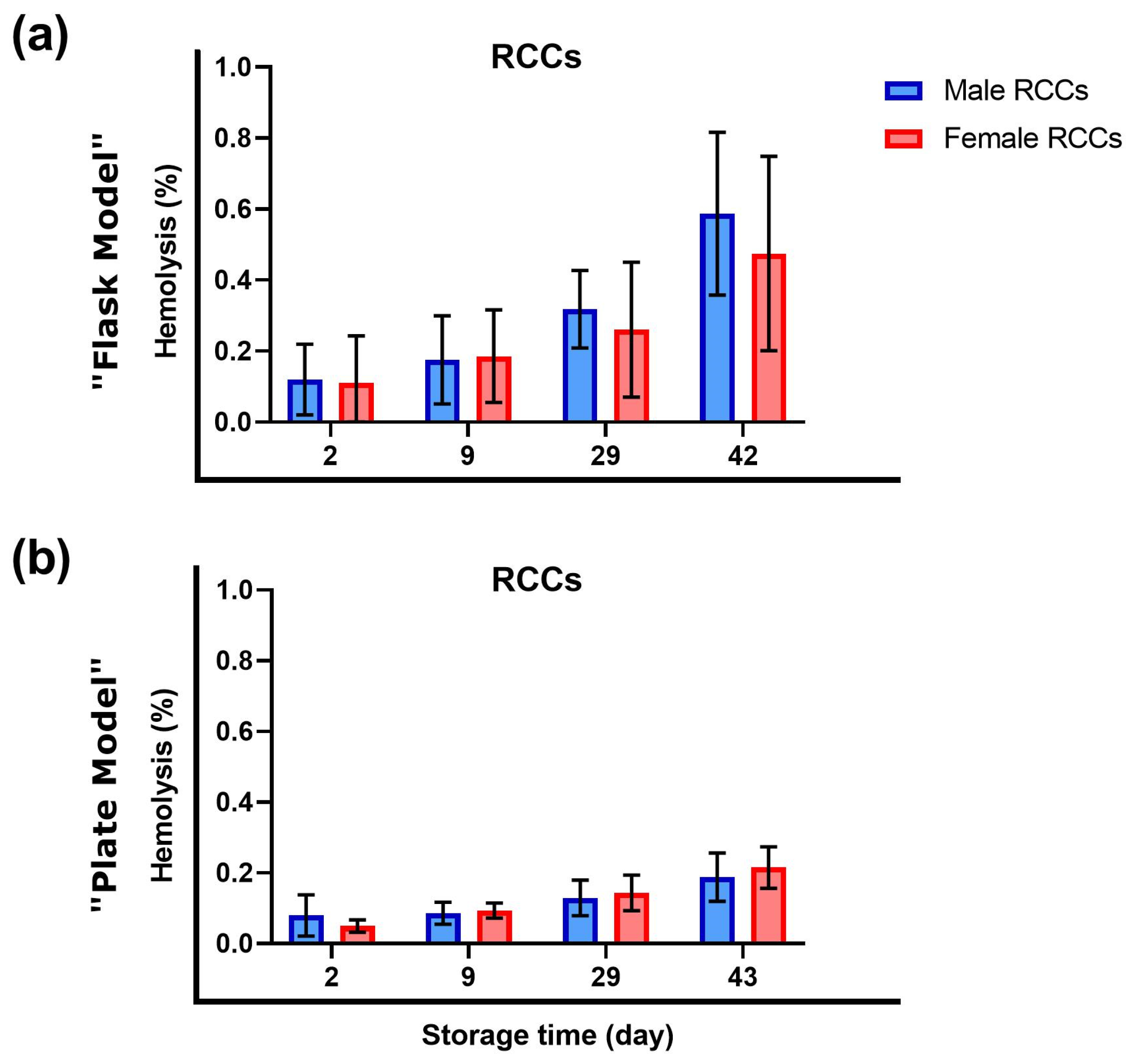
3.1. Sex Did Not Influence Hemolysis during Storage
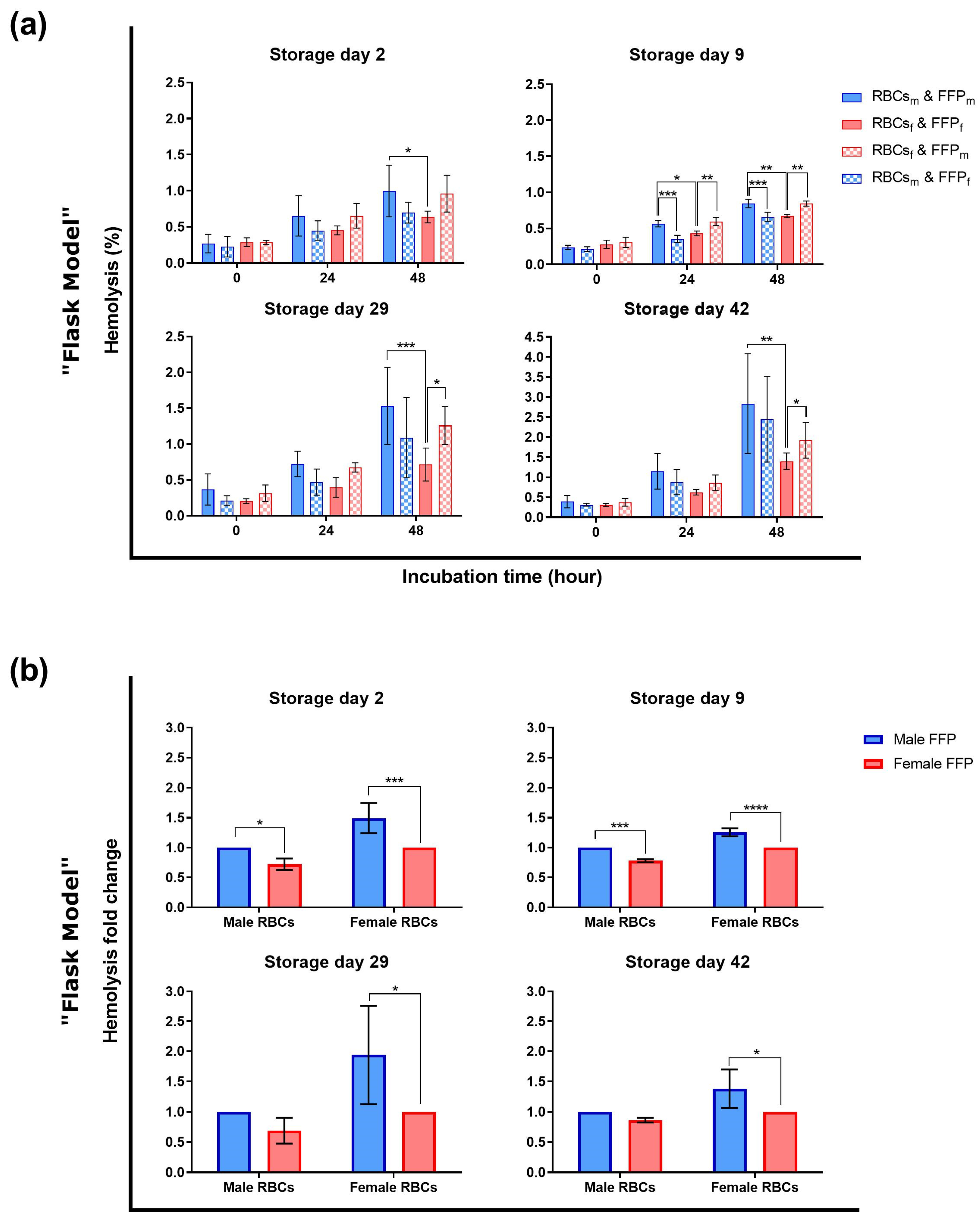
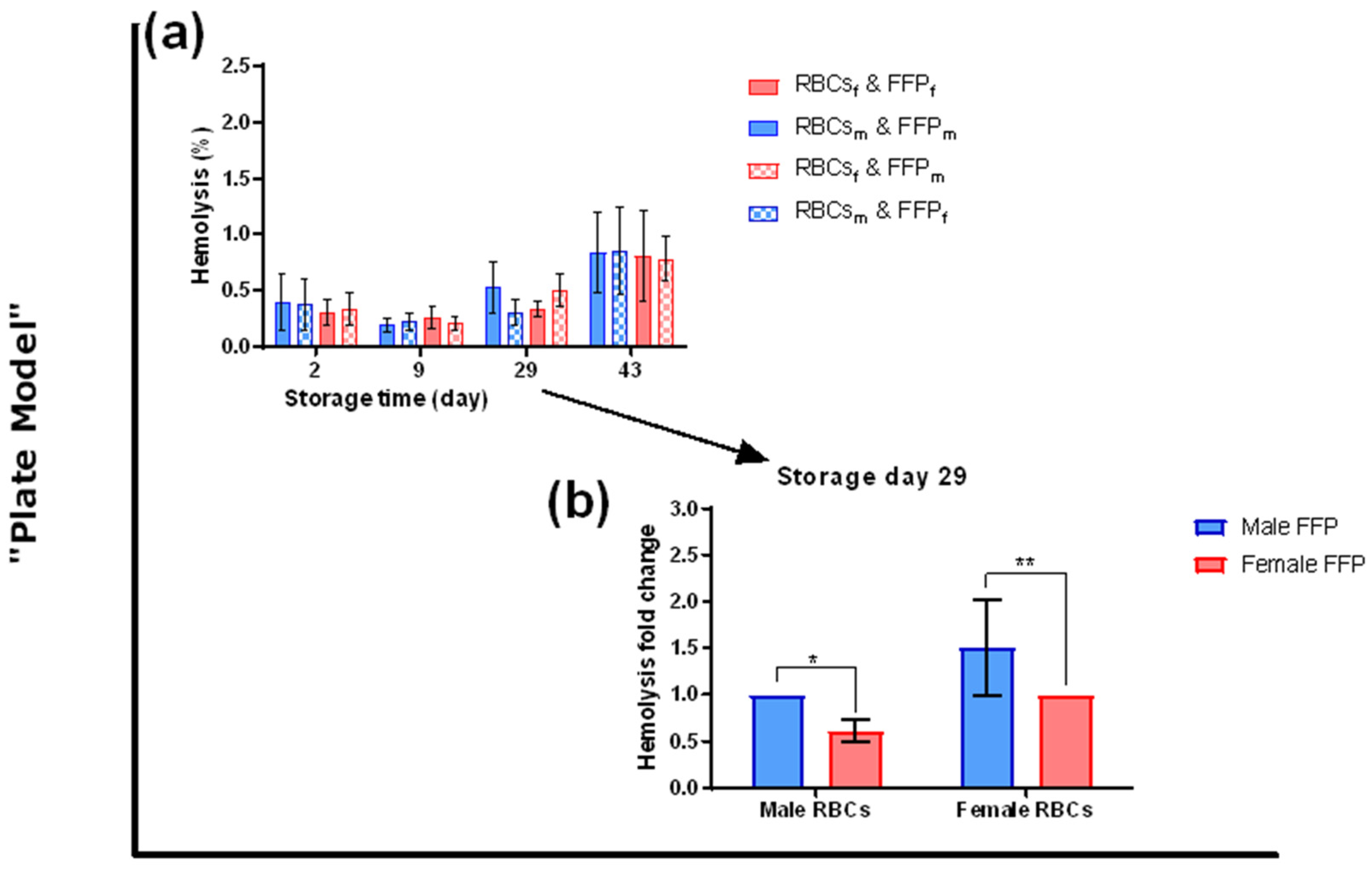
3.2. The Sex Influences Hemolysis of Red Blood Cells during Transfusion Simulations
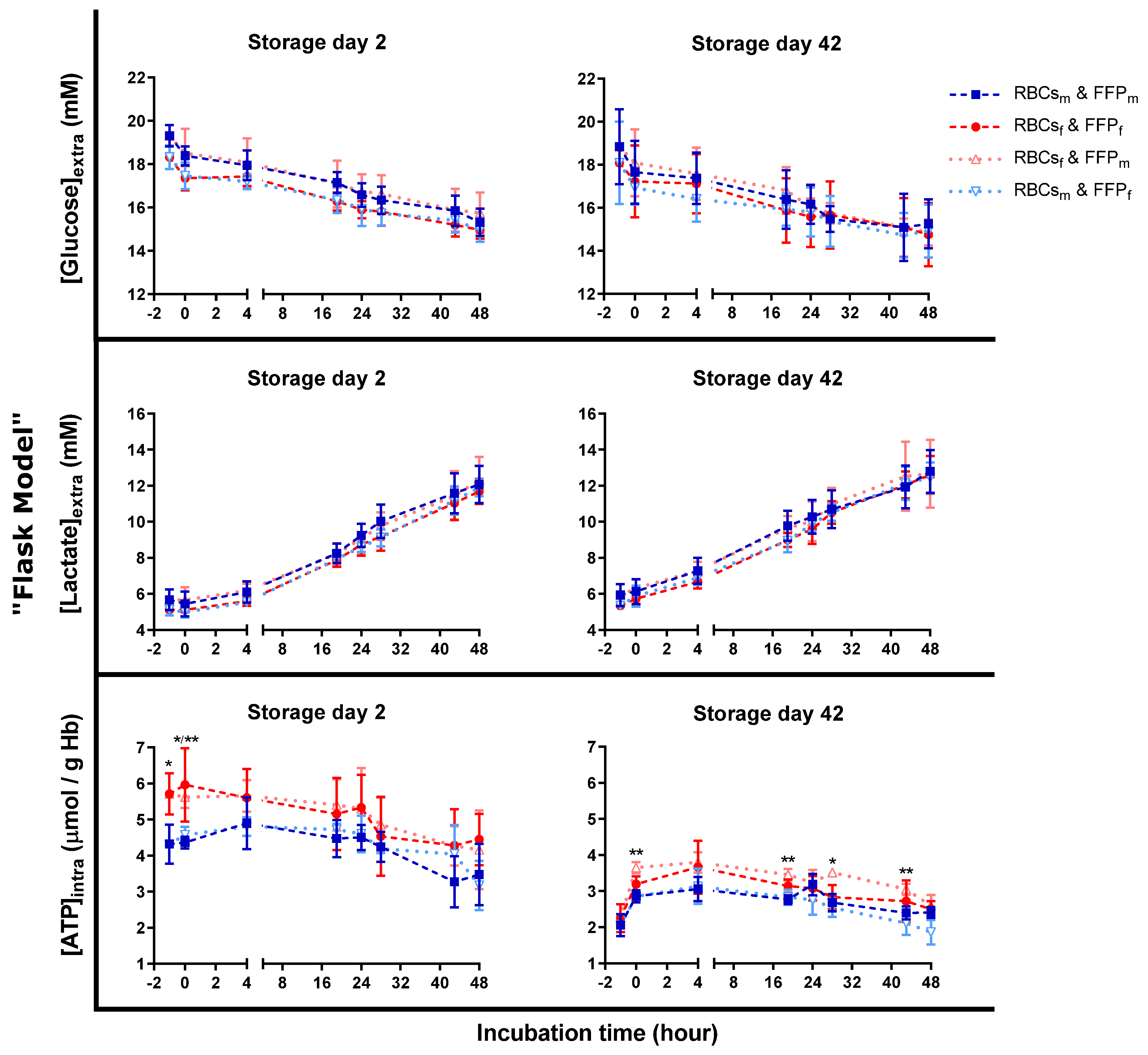
3.3. Similar Metabolic Activity during Transfusion Simulations
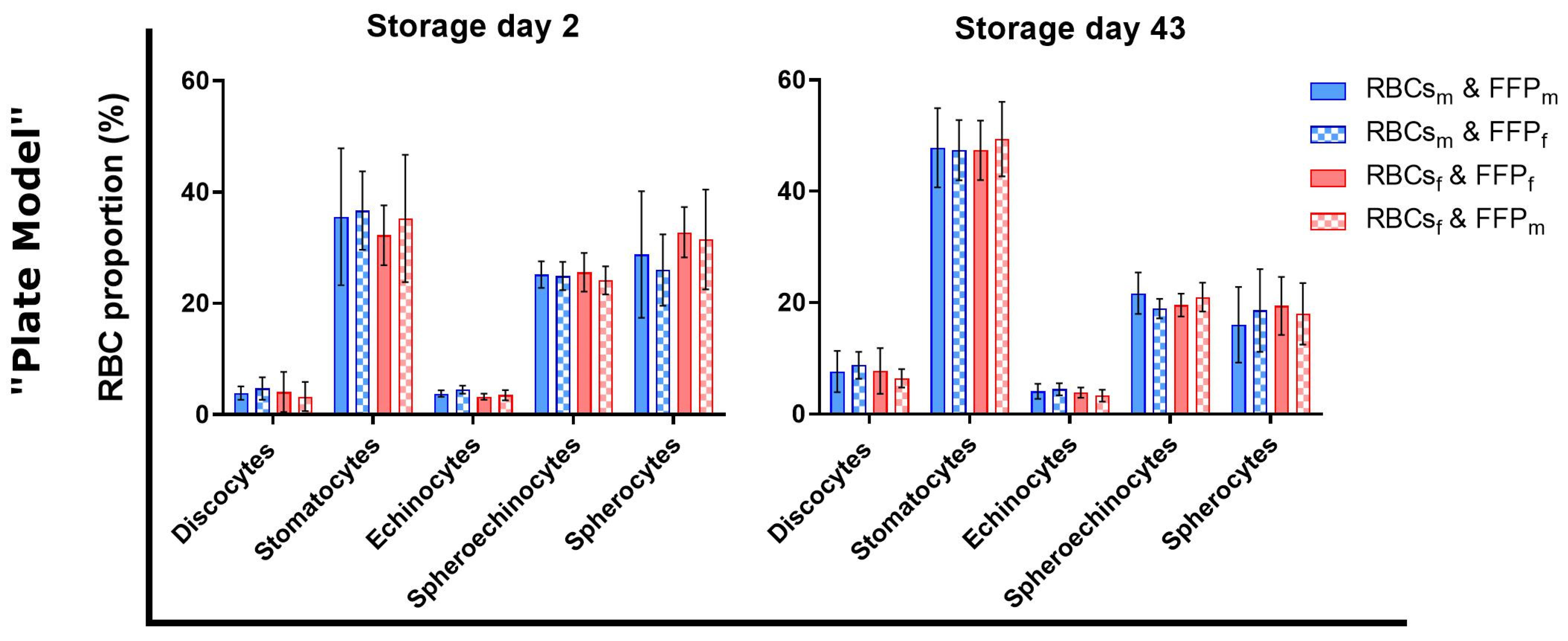
3.4. Sex Did Not Affect Red Blood Cell Morphology during Transfusion Simulations
4. Discussion
4.1. Hemolysis in Red Cell Concentrates
4.2. Hemolysis during Transfusion Simulations
4.2.1. Metabolic and Morphological Data Cannot Explain the Hemolytic Differences
4.2.2. Plasma Composition Could Explain the Hemolytic Profile
4.3. Limits of the Study
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kim, H.T.; Zhang, M.J.; Woolfrey, A.E.; St Martin, A.; Chen, J.; Saber, W.; Perales, M.A.; Armand, P.; Eapen, M. Donor and recipient sex in allogeneic stem cell transplantation: What really matters. Haematologica 2016, 101, 1260–1266. [Google Scholar] [CrossRef] [PubMed]
- Katz-Greenberg, G.; Shah, S. Sex and Gender Differences in Kidney Transplantation. Semin. Nephrol. 2022, 42, 219–229. [Google Scholar] [CrossRef]
- Weiss, E.S.; Allen, J.G.; Patel, N.D.; Russell, S.D.; Baumgartner, W.A.; Shah, A.S.; Conte, J.V. The impact of donor-recipient sex matching on survival after orthotopic heart transplantation: Analysis of 18 000 transplants in the modern era. Circ. Heart Fail. 2009, 2, 401–408. [Google Scholar] [CrossRef] [PubMed]
- Khush, K.K.; Kubo, J.T.; Desai, M. Influence of donor and recipient sex mismatch on heart transplant outcomes: Analysis of the International Society for Heart and Lung Transplantation Registry. J. Heart Lung Transplant. 2012, 31, 459–466. [Google Scholar] [CrossRef]
- Lin, Y.; Saw, C.L.; Hannach, B.; Goldman, M. Transfusion-related acute lung injury prevention measures and their impact at Canadian Blood Services. Transfusion 2012, 52, 567–574. [Google Scholar] [CrossRef]
- Lieberman, L.; Petraszko, T.; Yi, Q.L.; Hannach, B.; Skeate, R. Transfusion-related lung injury in children: A case series and review of the literature. Transfusion 2014, 54, 57–64. [Google Scholar] [CrossRef] [PubMed]
- Zeller, M.P.; Rochwerg, B.; Jamula, E.; Li, N.; Hillis, C.; Acker, J.P.; Runciman, R.J.R.; Lane, S.J.; Ahmed, N.; Arnold, D.M.; et al. Sex-mismatched red blood cell transfusions and mortality: A systematic review and meta-analysis. Vox Sang. 2019, 114, 505–516. [Google Scholar] [CrossRef]
- Middelburg, R.A.; Briet, E.; van der Bom, J.G. Mortality after transfusions, relation to donor sex. Vox Sang. 2011, 101, 221–229. [Google Scholar] [CrossRef]
- Chasse, M.; Tinmouth, A.; English, S.W.; Acker, J.P.; Wilson, K.; Knoll, G.; Shehata, N.; van Walraven, C.; Forster, A.J.; Ramsay, T.; et al. Association of Blood Donor Age and Sex With Recipient Survival After Red Blood Cell Transfusion. JAMA Intern. Med. 2016, 176, 1307–1314. [Google Scholar] [CrossRef]
- Heddle, N.M.; Cook, R.J.; Liu, Y.; Zeller, M.; Barty, R.; Acker, J.P.; Eikelboom, J.; Arnold, D.M. The association between blood donor sex and age and transfusion recipient mortality: An exploratory analysis. Transfusion 2019, 59, 482–491. [Google Scholar] [CrossRef]
- Desmarets, M.; Bardiaux, L.; Benzenine, E.; Dussaucy, A.; Binda, D.; Tiberghien, P.; Quantin, C.; Monnet, E. Effect of storage time and donor sex of transfused red blood cells on 1-year survival in patients undergoing cardiac surgery: An observational study. Transfusion 2016, 56, 1213–1222. [Google Scholar] [CrossRef] [PubMed]
- Holzmann, M.J.; Sartipy, U.; Olsson, M.L.; Dickman, P.; Edgren, G. Sex-Discordant Blood Transfusions and Survival After Cardiac Surgery: A Nationwide Cohort Study. Circulation 2016, 134, 1692–1694. [Google Scholar] [CrossRef] [PubMed]
- Edgren, G.; Ullum, H.; Rostgaard, K.; Erikstrup, C.; Sartipy, U.; Holzmann, M.J.; Nyren, O.; Hjalgrim, H. Association of Donor Age and Sex With Survival of Patients Receiving Transfusions. JAMA Intern. Med. 2017, 177, 854–860. [Google Scholar] [CrossRef] [PubMed]
- Edgren, G.; Murphy, E.L.; Brambilla, D.J.; Westlake, M.; Rostgaard, K.; Lee, C.; Cable, R.G.; Triulzi, D.; Bruhn, R.; St Lezin, E.M.; et al. Association of Blood Donor Sex and Prior Pregnancy With Mortality Among Red Blood Cell Transfusion Recipients. JAMA 2019, 321, 2183–2192. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, T.; Prudent, M.; D’Alessandro, A. Red blood cell storage lesion: Causes and potential clinical consequences. Blood Transfus. 2019, 17, 27–52. [Google Scholar] [CrossRef]
- Langst, E.; Tissot, J.D.; Prudent, M. Storage of red blood cell concentrates: Clinical impact. Transfus. Clin. Biol. 2021, 28, 397–402. [Google Scholar] [CrossRef]
- Delobel, J.; Prudent, M.; Tissot, J.D.; Lion, N. Proteomics of the red blood cell carbonylome during blood banking of erythrocyte concentrates. Proteom. Clin. Appl. 2016, 10, 257–266. [Google Scholar] [CrossRef]
- Delobel, J.; Prudent, M.; Rubin, O.; Crettaz, D.; Tissot, J.D.; Lion, N. Subcellular fractionation of stored red blood cells reveals a compartment-based protein carbonylation evolution. J. Proteom. 2012, 76, 181–193. [Google Scholar] [CrossRef]
- Kriebardis, A.G.; Antonelou, M.H.; Stamoulis, K.E.; Economou-Petersen, E.; Margaritis, L.H.; Papassideri, I.S. Progressive oxidation of cytoskeletal proteins and accumulation of denatured hemoglobin in stored red cells. J. Cell. Mol. Med. 2007, 11, 148–155. [Google Scholar] [CrossRef]
- Reisz, J.A.; Wither, M.J.; Dzieciatkowska, M.; Nemkov, T.; Issaian, A.; Yoshida, T.; Dunham, A.J.; Hill, R.C.; Hansen, K.C.; D’Alessandro, A. Oxidative modifications of glyceraldehyde 3-phosphate dehydrogenase regulate metabolic reprogramming of stored red blood cells. Blood 2016, 128, e32–e42. [Google Scholar] [CrossRef]
- Delobel, J.; Prudent, M.; Crettaz, D.; ElHajj, Z.; Riederer, B.M.; Tissot, J.D.; Lion, N. Cysteine redox proteomics of the hemoglobin-depleted cytosolic fraction of stored red blood cells. Proteom. Clin. Appl. 2016, 10, 883–893. [Google Scholar] [CrossRef] [PubMed]
- Kriebardis, A.G.; Antonelou, M.H.; Stamoulis, K.E.; Economou-Petersen, E.; Margaritis, L.H.; Papassideri, I.S. RBC-derived vesicles during storage: Ultrastructure, protein composition, oxidation, and signaling components. Transfusion 2008, 48, 1943–1953. [Google Scholar] [CrossRef] [PubMed]
- Prudent, M.; Crettaz, D.; Delobel, J.; Seghatchian, J.; Tissot, J.D.; Lion, N. Differences between calcium-stimulated and storage-induced erythrocyte-derived microvesicles. Transfus. Apher. Sci. 2015, 53, 153–158. [Google Scholar] [CrossRef] [PubMed]
- Bosman, G.J.; Lasonder, E.; Groenen-Dopp, Y.A.; Willekens, F.L.; Werre, J.M. The proteome of erythrocyte-derived microparticles from plasma: New clues for erythrocyte aging and vesiculation. J. Proteom. 2012, 76, 203–210. [Google Scholar] [CrossRef]
- Tzounakas, V.L.; Anastasiadi, A.T.; Lekka, M.E.; Papageorgiou, E.G.; Stamoulis, K.; Papassideri, I.S.; Kriebardis, A.G.; Antonelou, M.H. Deciphering the Relationship Between Free and Vesicular Hemoglobin in Stored Red Blood Cell Units. Front. Physiol. 2022, 13, 840995. [Google Scholar] [CrossRef]
- Bardyn, M.; Rappaz, B.; Jaferzadeh, K.; Crettaz, D.; Tissot, J.D.; Moon, I.; Turcatti, G.; Lion, N.; Prudent, M. Red blood cells ageing markers: A multi-parametric analysis. Blood Transfus. 2017, 15, 239–248. [Google Scholar] [CrossRef]
- Roussel, C.; Dussiot, M.; Marin, M.; Morel, A.; Ndour, P.A.; Duez, J.; Le Van Kim, C.; Hermine, O.; Colin, Y.; Buffet, P.A.; et al. Spherocytic shift of red blood cells during storage provides a quantitative whole cell-based marker of the storage lesion. Transfusion 2017, 57, 1007–1018. [Google Scholar] [CrossRef]
- Barshtein, G.; Gural, A.; Manny, N.; Zelig, O.; Yedgar, S.; Arbell, D. Storage-induced damage to red blood cell mechanical properties can be only partially reversed by rejuvenation. Transfus. Med. Hemother. 2014, 41, 197–204. [Google Scholar] [CrossRef]
- Kanias, T.; Lanteri, M.C.; Page, G.P.; Guo, Y.; Endres, S.M.; Stone, M.; Keating, S.; Mast, A.E.; Cable, R.G.; Triulzi, D.J.; et al. Ethnicity, sex, and age are determinants of red blood cell storage and stress hemolysis: Results of the REDS-III RBC-Omics study. Blood Adv. 2017, 1, 1132–1141. [Google Scholar] [CrossRef]
- Jordan, A.; Chen, D.; Yi, Q.L.; Kanias, T.; Gladwin, M.T.; Acker, J.P. Assessing the influence of component processing and donor characteristics on quality of red cell concentrates using quality control data. Vox Sang. 2016, 111, 8–15. [Google Scholar] [CrossRef]
- Bardyn, M.; Crettaz, D.; Borlet, M.; Langst, E.; Martin, A.; Abonnenc, M.; Tissot, J.D.; Dunham, A.; Yoshida, T.; Prudent, M. Hypoxia and hypocapnia storage of gamma-irradiated red cell concentrates. Blood Transfus. 2021, 19, 300–308. [Google Scholar] [CrossRef] [PubMed]
- Grau, M.; Cremer, J.M.; Schmeichel, S.; Kunkel, M.; Bloch, W. Comparisons of Blood Parameters, Red Blood Cell Deformability and Circulating Nitric Oxide Between Males and Females Considering Hormonal Contraception: A Longitudinal Gender Study. Front. Physiol. 2018, 9, 1835. [Google Scholar] [CrossRef] [PubMed]
- Tzounakas, V.L.; Anastasiadi, A.T.; Drossos, P.V.; Karadimas, D.G.; Valsami, S.E.; Stamoulis, K.E.; Papassideri, I.S.; Politou, M.; Antonelou, M.H.; Kriebardis, A.G. Sex-related aspects of the red blood cell storage lesion. Blood Transfus. 2021, 19, 224–236. [Google Scholar] [CrossRef]
- D’Alessandro, A.; Fu, X.; Kanias, T.; Reisz, J.A.; Culp-Hill, R.; Guo, Y.; Gladwin, M.T.; Page, G.; Kleinman, S.; Lanteri, M.; et al. Donor sex, age and ethnicity impact stored red blood cell antioxidant metabolism through mechanisms in part explained by glucose 6-phosphate dehydrogenase levels and activity. Haematologica 2021, 106, 1290–1302. [Google Scholar] [CrossRef]
- Bardyn, M.; Maye, S.; Lesch, A.; Delobel, J.; Tissot, J.D.; Cortes-Salazar, F.; Tacchini, P.; Lion, N.; Girault, H.H.; Prudent, M. The antioxidant capacity of erythrocyte concentrates is increased during the first week of storage and correlated with the uric acid level. Vox Sang. 2017, 112, 638–647. [Google Scholar] [CrossRef] [PubMed]
- Vona, R.; Gambardella, L.; Ortona, E.; Santulli, M.; Malorni, W.; Care, A.; Pietraforte, D.; Straface, E. Functional Estrogen Receptors of Red Blood Cells. Do They Influence Intracellular Signaling? Cell. Physiol. Biochem. 2019, 53, 186–199. [Google Scholar] [CrossRef]
- Swanepoel, A.C.; Emmerson, O.; Pretorius, E. Effect of Progesterone and Synthetic Progestins on Whole Blood Clot Formation and Erythrocyte Structure. Microsc. Microanal. 2017, 23, 607–617. [Google Scholar] [CrossRef] [PubMed]
- Islamzada, E.; Matthews, K.; Lamoureux, E.S.; Duffy, S.P.; Scott, M.D.; Ma, H. Degradation of red blood cell deformability during cold storage in blood bags. EJHaem 2022, 3, 63–71. [Google Scholar] [CrossRef]
- Matthews, K.; Myrand-Lapierre, M.E.; Ang, R.R.; Duffy, S.P.; Scott, M.D.; Ma, H. Microfluidic deformability analysis of the red cell storage lesion. J. Biomech. 2015, 48, 4065–4072. [Google Scholar] [CrossRef]
- Kanias, T.; Sinchar, D.; Osei-Hwedieh, D.; Baust, J.J.; Jordan, A.; Zimring, J.C.; Waterman, H.R.; de Wolski, K.S.; Acker, J.P.; Gladwin, M.T. Testosterone-dependent sex differences in red blood cell hemolysis in storage, stress, and disease. Transfusion 2016, 56, 2571–2583. [Google Scholar] [CrossRef]
- Langst, E.; Crettaz, D.; Delobel, J.; Renella, R.; Bardyn, M.; Turcatti, G.; Tissot, J.D.; Prudent, M. In vitro-transfusional model for red-blood-cell study: The advantage of lowering hematocrit. Blood Transfus. 2022. ahead of Print. [Google Scholar] [CrossRef]
- Han, V.; Serrano, K.; Devine, D.V. A comparative study of common techniques used to measure haemolysis in stored red cell concentrates. Vox Sang. 2010, 98, 116–123. [Google Scholar] [CrossRef]
- Bardyn, M.; Chen, J.; Dussiot, M.; Crettaz, D.; Schmid, L.; Langst, E.; Amireault, P.; Tissot, J.D.; Jolicoeur, M.; Prudent, M. Restoration of Physiological Levels of Uric Acid and Ascorbic Acid Reroutes the Metabolism of Stored Red Blood Cells. Metabolites 2020, 10, 226. [Google Scholar] [CrossRef] [PubMed]
- Jeswani, G.; Alexander, A.; Saraf, S.; Saraf, S.; Qureshi, A.; Ajazuddin. Recent approaches for reducing hemolytic activity of chemotherapeutic agents. J. Control. Release 2015, 211, 10–21. [Google Scholar] [CrossRef]
- Mykhailova, O.; Olafson, C.; Turner, T.R.; D’Alessandro, A.; Acker, J.P. Donor-dependent aging of young and old red blood cell subpopulations: Metabolic and functional heterogeneity. Transfusion 2020, 60, 2633–2646. [Google Scholar] [CrossRef]
- Szczesny-Malysiak, E.; Mohaissen, T.; Bulat, K.; Kaczmarska, M.; Wajda, A.; Marzec, K.M. Sex-dependent membranopathy in stored human red blood cells. Haematologica 2021, 106, 2779–2782. [Google Scholar] [CrossRef]
- Bardyn, M.; Martin, A.; Dognitz, N.; Abonnenc, M.; Dunham, A.; Yoshida, T.; Prudent, M. Oxygen in Red Blood Cell Concentrates: Influence of Donors’ Characteristics and Blood Processing. Front. Physiol. 2020, 11, 616457. [Google Scholar] [CrossRef] [PubMed]
- D’Alessandro, A.; Nemkov, T.; Sun, K.; Liu, H.; Song, A.; Monte, A.A.; Subudhi, A.W.; Lovering, A.T.; Dvorkin, D.; Julian, C.G.; et al. AltitudeOmics: Red Blood Cell Metabolic Adaptation to High Altitude Hypoxia. J. Proteome Res. 2016, 15, 3883–3895. [Google Scholar] [CrossRef]
- D’Alessandro, A.; Xia, Y. Erythrocyte adaptive metabolic reprogramming under physiological and pathological hypoxia. Curr. Opin. Hematol. 2020, 27, 155–162. [Google Scholar] [CrossRef]
- Yoshida, T.; Blair, A.; D’Alessandro, A.; Nemkov, T.; Dioguardi, M.; Silliman, C.C.; Dunham, A. Enhancing uniformity and overall quality of red cell concentrate with anaerobic storage. Blood Transfus. 2017, 15, 172–181. [Google Scholar] [CrossRef]
- Reinhart, W.H.; Chien, S. Echinocyte-stomatocyte transformation and shape control of human red blood cells: Morphological aspects. Am. J. Hematol. 1987, 24, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Vlaar, A.P.; Kulik, W.; Nieuwland, R.; Peters, C.P.; Tool, A.T.; van Bruggen, R.; Juffermans, N.P.; de Korte, D. Accumulation of bioactive lipids during storage of blood products is not cell but plasma derived and temperature dependent. Transfusion 2011, 51, 2358–2366. [Google Scholar] [CrossRef] [PubMed]
- Minetti, G.; Egee, S.; Morsdorf, D.; Steffen, P.; Makhro, A.; Achilli, C.; Ciana, A.; Wang, J.; Bouyer, G.; Bernhardt, I.; et al. Red cell investigations: Art and artefacts. Blood Rev. 2013, 27, 91–101. [Google Scholar] [CrossRef] [PubMed]
- Unfer, T.C.; Maurer, L.H.; Kemerich, D.M.; Figueiredo, C.G.; Duarte, M.M.; Gelain, D.P.; Moreira, J.C.; Emanuelli, T. Non-genomic, direct modulatory effect of 17beta-estradiol, progesterone and their synthetic derivatives on the activity of human erythrocyte CuZn superoxide dismutase. Free Radic. Res. 2013, 47, 219–232. [Google Scholar] [CrossRef]
- Mary, A.; Bonne, C.; Modat, G. Erythrocyte deformability in an in vitro model of hypoxia/reoxygenation—Protective effects of superoxide dismutase and catalase. Clin. Hemorheol. Microcirc. 1992, 12, 287–296. [Google Scholar] [CrossRef]
- Silliman, C.C.; Dzieciatkowska, M.; Moore, E.E.; Kelher, M.R.; Banerjee, A.; Liang, X.; Land, K.J.; Hansen, K.C. Proteomic analyses of human plasma: Venus versus Mars. Transfusion 2012, 52, 417–424. [Google Scholar] [CrossRef]
- Ishikawa, M.; Tajima, Y.; Murayama, M.; Senoo, Y.; Maekawa, K.; Saito, Y. Plasma and serum from nonfasting men and women differ in their lipidomic profiles. Biol. Pharm. Bull. 2013, 36, 682–685. [Google Scholar] [CrossRef]
- Ishikawa, M.; Maekawa, K.; Saito, K.; Senoo, Y.; Urata, M.; Murayama, M.; Tajima, Y.; Kumagai, Y.; Saito, Y. Plasma and serum lipidomics of healthy white adults shows characteristic profiles by subjects’ gender and age. PLoS ONE 2014, 9, e91806. [Google Scholar] [CrossRef]
- Spiegel, S.; Milstien, S. Sphingosine-1-phosphate: An enigmatic signalling lipid. Nat. Rev. Mol. Cell Biol. 2003, 4, 397–407. [Google Scholar] [CrossRef]
- Serhan, C.N.; Chiang, N.; Van Dyke, T.E. Resolving inflammation: Dual anti-inflammatory and pro-resolution lipid mediators. Nat. Rev. Immunol. 2008, 8, 349–361. [Google Scholar] [CrossRef]
- Kikas, P.; Chalikias, G.; Tziakas, D. Cardiovascular Implications of Sphingomyelin Presence in Biological Membranes. Eur. Cardiol. 2018, 13, 42–45. [Google Scholar] [CrossRef] [PubMed]
- Xu, F.; Zou, L.; Lin, Q.; Ong, C.N. Use of liquid chromatography/tandem mass spectrometry and online databases for identification of phosphocholines and lysophosphatidylcholines in human red blood cells. Rapid Commun. Mass Spectrom. 2009, 23, 3243–3254. [Google Scholar] [CrossRef] [PubMed]
- Merrill, A.H., Jr.; Wang, E.; Innis, W.S.; Mullins, R. Increases in serum sphingomyelin by 17 beta-estradiol. Lipids 1985, 20, 252–254. [Google Scholar] [CrossRef] [PubMed]
- Joles, J.A.; Bijleveld, C.; van Tol, A.; Geelen, M.J.; Koomans, H.A. Estrogen replacement during hypoalbuminemia may enhance atherosclerotic risk. J. Am. Soc. Nephrol. 1997, 8, 1870–1876. [Google Scholar] [CrossRef]
- Alessandri, J.M.; Extier, A.; Al-Gubory, K.H.; Harbeby, E.; Lallemand, M.S.; Linard, A.; Lavialle, M.; Guesnet, P. Influence of gender on DHA synthesis: The response of rat liver to low dietary alpha-linolenic acid evidences higher omega3 ∆4-desaturation index in females. Eur. J. Nutr. 2012, 51, 199–209. [Google Scholar] [CrossRef] [PubMed]
- Malka, R.; Nathan, D.M.; Higgins, J.M. Mechanistic modeling of hemoglobin glycation and red blood cell kinetics enables personalized diabetes monitoring. Sci. Transl. Med. 2016, 8, 359ra130. [Google Scholar] [CrossRef]
- Lacroix, J.; Hebert, P.C.; Fergusson, D.A.; Tinmouth, A.; Cook, D.J.; Marshall, J.C.; Clayton, L.; McIntyre, L.; Callum, J.; Turgeon, A.F.; et al. Age of transfused blood in critically ill adults. N. Engl. J. Med. 2015, 372, 1410–1418. [Google Scholar] [CrossRef]
- Heddle, N.M.; Cook, R.J.; Arnold, D.M.; Liu, Y.; Barty, R.; Crowther, M.A.; Devereaux, P.J.; Hirsh, J.; Warkentin, T.E.; Webert, K.E.; et al. Effect of Short-Term vs. Long-Term Blood Storage on Mortality after Transfusion. N. Engl. J. Med. 2016, 375, 1937–1945. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Laengst, E.; Crettaz, D.; Tissot, J.-D.; Prudent, M. The Effect of the Donor’s and Recipient’s Sex on Red Blood Cells Evaluated Using Transfusion Simulations. Cells 2023, 12, 1454. https://doi.org/10.3390/cells12111454
Laengst E, Crettaz D, Tissot J-D, Prudent M. The Effect of the Donor’s and Recipient’s Sex on Red Blood Cells Evaluated Using Transfusion Simulations. Cells. 2023; 12(11):1454. https://doi.org/10.3390/cells12111454
Chicago/Turabian StyleLaengst, Emmanuel, David Crettaz, Jean-Daniel Tissot, and Michel Prudent. 2023. "The Effect of the Donor’s and Recipient’s Sex on Red Blood Cells Evaluated Using Transfusion Simulations" Cells 12, no. 11: 1454. https://doi.org/10.3390/cells12111454