
Identification of Neurocan and Phosphacan as Early Biomarkers for Open Neural Tube Defects
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
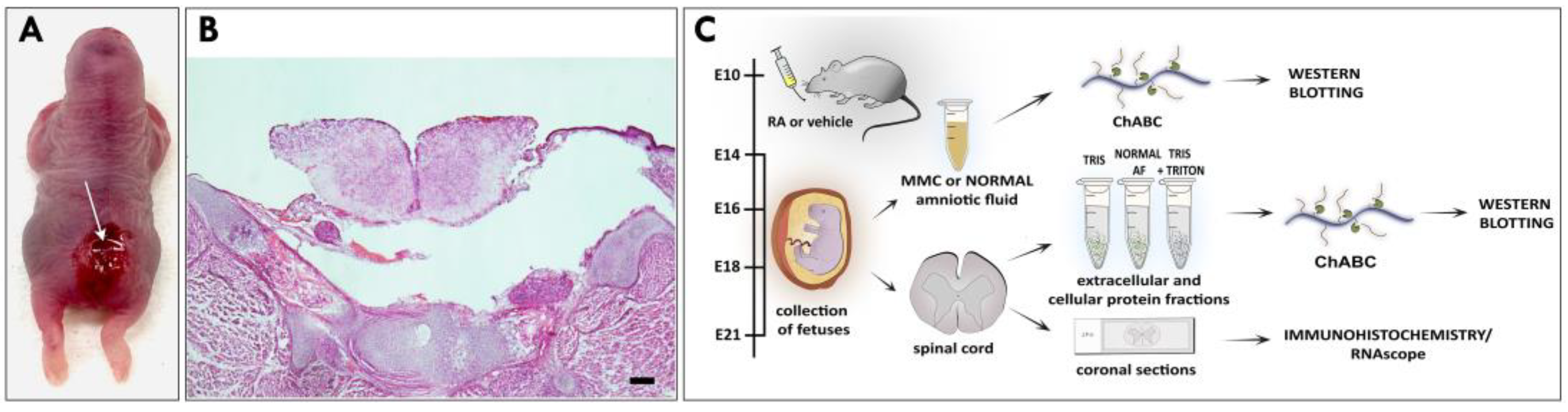
2.1. Retinoic-Acid-Induced Animal Model of MMC
2.2. AF Preparation
2.3. Protein Isolation
2.3.1. Total Spinal Cord Protein Extracts
2.3.2. Sequential Spinal Cord Protein Extracts
2.3.3. MMC Spinal Cord Protein Extracts in Normal AF
2.4. Western Blotting
2.5. Histology and Immunofluorescence
2.6. RNAscope and Coimmunostaining
2.7. Statistical Analysis
3. Results
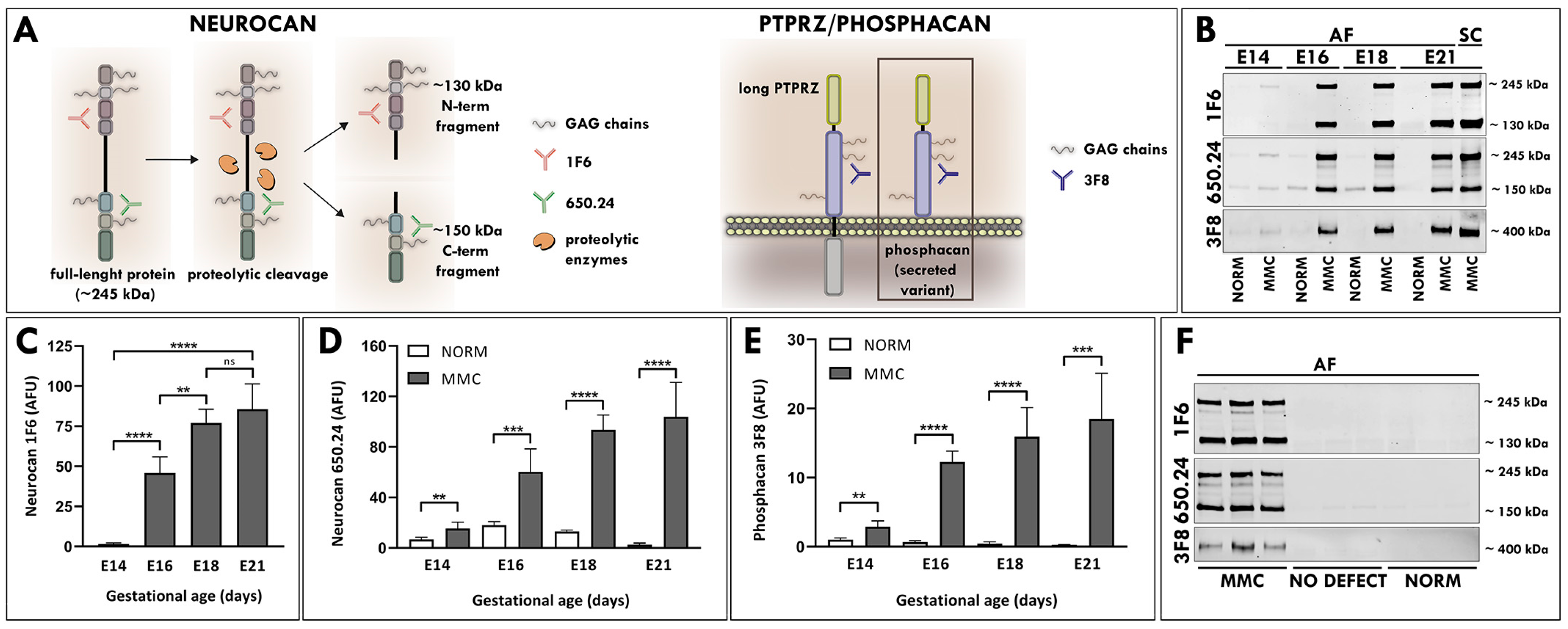
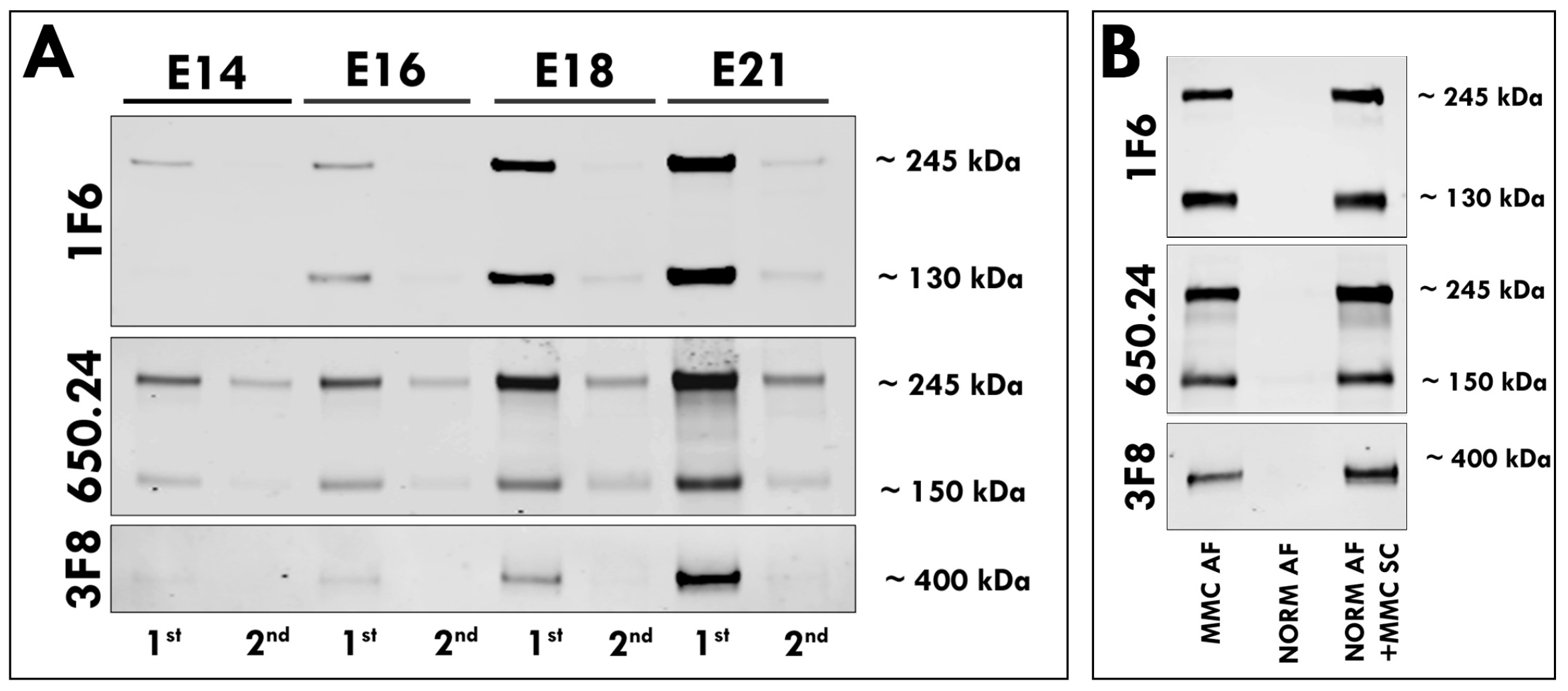
3.1. Elevated Levels of Neurocan and Phosphacan in the AF of MMC Fetuses
3.2. Secretion of Neurocan and Phosphacan from the MMC Spinal Cord
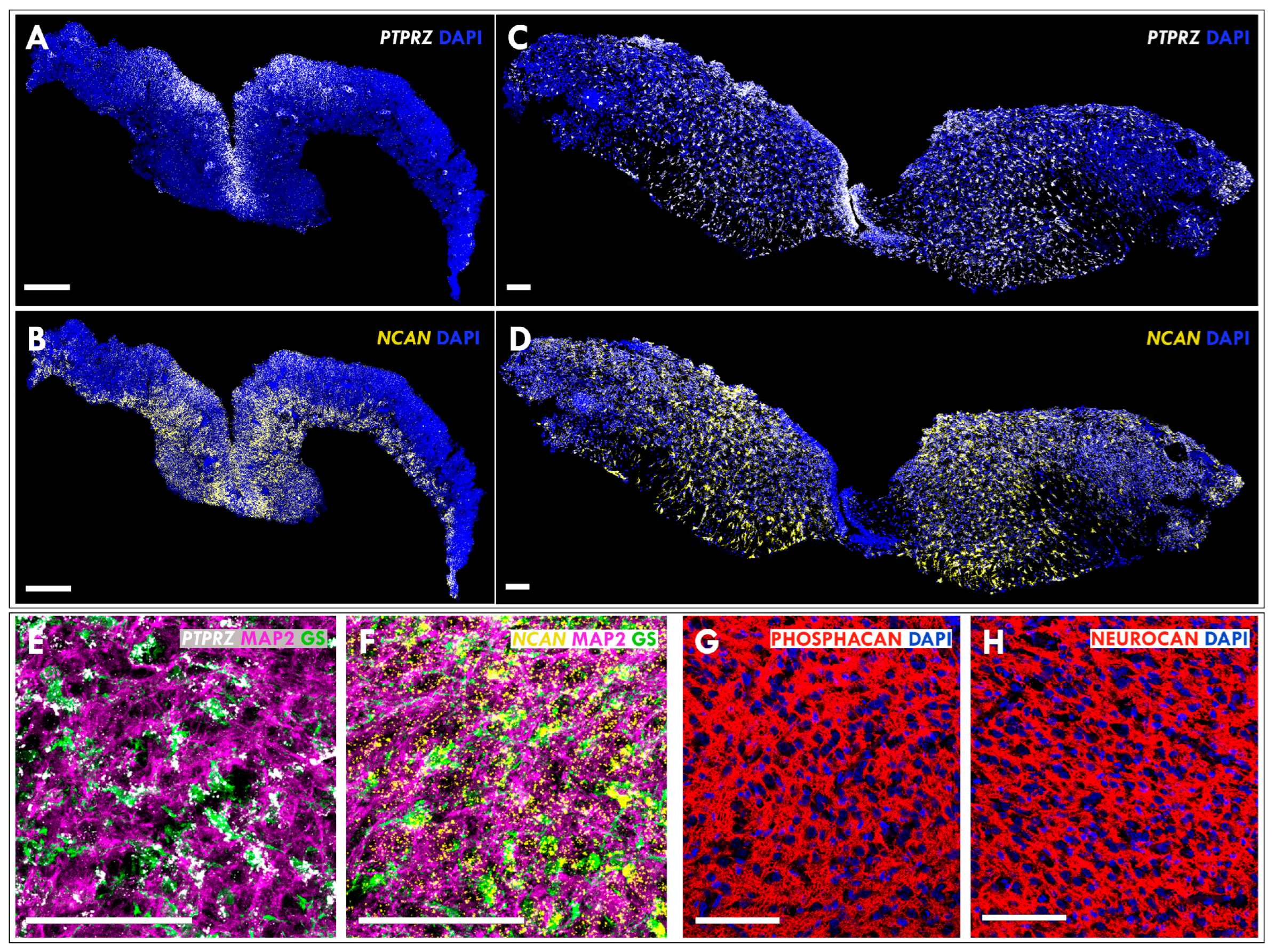
3.3. Expression of Neurocan and Phosphacan in the Developing MMC Spinal Cord
4. Discussion
5. Conclusions
6. Patents
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Copp, A.J.; Greene, N.D.; Murdoch, J.N. The genetic basis of mammalian neurulation. Nat. Rev. Genet. 2003, 4, 784–793. [Google Scholar] [CrossRef] [PubMed]
- Parker, S.E.; Mai, C.T.; Canfield, M.A.; Rickard, R.; Wang, Y.; Meyer, R.E.; Anderson, P.; Mason, C.A.; Collins, J.S.; Kirby, R.S.; et al. Updated National Birth Prevalence estimates for selected birth defects in the United States, 2004–2006. Birth Defects Res. A Clin. Mol. Teratol. 2010, 88, 1008–1016. [Google Scholar] [CrossRef] [PubMed]
- Canfield, M.A.; Mai, C.T.; Wang, Y.; O’Halloran, A.; Marengo, L.K.; Olney, R.S.; Borger, C.L.; Rutkowski, R.; Fornoff, J.; Irwin, N.; et al. The association between race/ethnicity and major birth defects in the United States, 1999–2007. Am. J. Public Health 2014, 104, e14–e23. [Google Scholar] [CrossRef] [PubMed]
- Kaufman, B.A. Neural tube defects. Pediatr. Clin. North Am. 2004, 51, 389–419. [Google Scholar] [CrossRef]
- Hunt, G.M. Open spina bifida: Outcome for a complete cohort treated unselectively and followed into adulthood. Dev. Med. Child Neurol. 1990, 32, 108–118. [Google Scholar] [CrossRef]
- Dias, M.S.; McLone, D.G. Hydrocephalus in the child with dysraphism. Neurosurg. Clin. N. Am. 1993, 4, 715–726. [Google Scholar] [CrossRef]
- Hunt, G.M.; Poulton, A. Open spina bifida: A complete cohort reviewed 25 years after closure. Dev. Med. Child Neurol. 1995, 37, 19–29. [Google Scholar] [CrossRef]
- Tomlinson, P.; Sugarman, I.D. Complications with shunts in adults with spina bifida. BMJ 1995, 311, 286–287. [Google Scholar] [CrossRef] [Green Version]
- Hunt, G.M.; Oakeshott, P. Outcome in people with open spina bifida at age 35: Prospective community based cohort study. BMJ 2003, 326, 1365–1366. [Google Scholar] [CrossRef] [Green Version]
- Bowman, R.M.; McLone, D.G.; Grant, J.A.; Tomita, T.; Ito, J.A. Spina bifida outcome: A 25-year prospective. Pediatr. Neurosurg. 2001, 34, 114–120. [Google Scholar] [CrossRef]
- Sandler, A.D. Children with spina bifida: Key clinical issues. Pediatr. Clin. N. Am. 2010, 57, 879–892. [Google Scholar] [CrossRef] [PubMed]
- Palomaki, G.E.; Bupp, C.; Gregg, A.R.; Norton, M.E.; Oglesbee, D.; Best, R.G.; ACMG Biochemical Genetics Subcommittee of the Laboratory Quality Assurance Committee documents@ acmg. Net. Laboratory screening and diagnosis of open neural tube defects, 2019 revision: A technical standard of the American College of Medical Genetics and Genomics (ACMG). Genet. Med. 2020, 22, 462–474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Norem, C.T.; Schoen, E.J.; Walton, D.L.; Krieger, R.C.; O’Keefe, J.; To, T.T.; Ray, G.T. Routine ultrasonography compared with maternal serum alpha-fetoprotein for neural tube defect screening. Obstet. Gynecol. 2005, 106, 747–752. [Google Scholar] [CrossRef] [PubMed]
- Rose, N.M.M. Fetal Neural Tube Defects: Diagnosis, Management, and Treatment; Global Library of Women’s Medicine’s: London, UK, 2009. [Google Scholar]
- Dashe, J.S.; Twickler, D.M.; Santos-Ramos, R.; McIntire, D.D.; Ramus, R.M. Alpha-fetoprotein detection of neural tube defects and the impact of standard ultrasound. Am. J. Obstet. Gynecol. 2006, 195, 1623–1628. [Google Scholar] [CrossRef]
- Margolis, R.K.; Rauch, U.; Maurel, P.; Margolis, R.U. Neurocan and phosphacan: Two major nervous tissue-specific chondroitin sulfate proteoglycans. Perspect. Dev. Neurobiol. 1996, 3, 273–290. [Google Scholar]
- Meyer-Puttlitz, B.; Junker, E.; Margolis, R.U.; Margolis, R.K. Chondroitin sulfate proteoglycans in the developing central nervous system. II. Immunocytochemical localization of neurocan and phosphacan. J. Comp. Neurol. 1996, 366, 44–54. [Google Scholar] [CrossRef]
- Deepa, S.S.; Carulli, D.; Galtrey, C.; Rhodes, K.; Fukuda, J.; Mikami, T.; Sugahara, K.; Fawcett, J.W. Composition of perineuronal net extracellular matrix in rat brain: A different disaccharide composition for the net-associated proteoglycans. J. Biol. Chem. 2006, 281, 17789–17800. [Google Scholar] [CrossRef] [Green Version]
- Grumet, M.; Friedlander, D.R.; Sakurai, T. Functions of brain chondroitin sulfate proteoglycans during developments: Interactions with adhesion molecules. Perspect. Dev. Neurobiol. 1996, 3, 319–330. [Google Scholar]
- Danzer, E.; Schwarz, U.; Wehrli, S.; Radu, A.; Adzick, N.S.; Flake, A.W. Retinoic acid induced myelomeningocele in fetal rats: Characterization by histopathological analysis and magnetic resonance imaging. Exp. Neurol. 2005, 194, 467–475. [Google Scholar] [CrossRef]
- Barbe, M.F.; Adiga, R.; Gordiienko, O.; Pleshko, N.; Selzer, M.E.; Krynska, B. Micro-computed tomography assessment of vertebral column defects in retinoic acid-induced rat model of myelomeningocele. Birth Defects Res. A Clin. Mol. Teratol. 2014, 100, 453–462. [Google Scholar] [CrossRef]
- Zieba, J.; Miller, A.; Gordiienko, O.; Smith, G.M.; Krynska, B. Clusters of amniotic fluid cells and their associated early neuroepithelial markers in experimental myelomeningocele: Correlation with astrogliosis. PLoS ONE 2017, 12, e0174625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zieba, J.; Walczak, M.; Gordiienko, O.; Gerstenhaber, J.A.; Smith, G.M.; Krynska, B. Altered Amniotic Fluid Levels of Hyaluronic Acid in Fetal Rats with Myelomeningocele: Understanding Spinal Cord Injury. J. Neurotrauma. 2019, 36, 1965–1973. [Google Scholar] [CrossRef] [PubMed]
- Maurel, P.; Rauch, U.; Flad, M.; Margolis, R.K.; Margolis, R.U. Phosphacan, a chondroitin sulfate proteoglycan of brain that interacts with neurons and neural cell-adhesion molecules, is an extracellular variant of a receptor-type protein tyrosine phosphatase. Proc. Natl. Acad. Sci. USA 1994, 91, 2512–2516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anlauf, E.; Derouiche, A. Glutamine synthetase as an astrocytic marker: Its cell type and vesicle localization. (1664–2392 (Print)). Front. Endocrinol. 2013, 4, 144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dehmelt, L.; Halpain, S. The MAP2/Tau family of microtubule-associated proteins. (1474-760X (Electronic)). Genome Biol. 2005, 6, 204. [Google Scholar]
- Petzold, A.; Stiefel, D.; Copp, A.J. Amniotic fluid brain-specific proteins are biomarkers for spinal cord injury in experimental myelomeningocele. J. Neurochem. 2005, 95, 594–598. [Google Scholar] [CrossRef] [Green Version]
- Danzer, E.; Zhang, L.; Radu, A.; Bebbington, M.W.; Liechty, K.W.; Adzick, N.S.; Flake, A.W. Amniotic fluid levels of glial fibrillary acidic protein in fetal rats with retinoic acid induced myelomeningocele: A potential marker for spinal cord injury. Am. J. Obstet. Gynecol. 2011, 204, 178.e1–178.e11. [Google Scholar] [CrossRef]
- Geer, L.A.; Pycke, B.F.; Sherer, D.M.; Abulafia, O.; Halden, R.U. Use of amniotic fluid for determining pregnancies at risk of preterm birth and for studying diseases of potential environmental etiology. Environ. Res. 2015, 136, 470–481. [Google Scholar] [CrossRef] [Green Version]
- Tsangaris, G.T.; Anagnostopoulos, A.K.; Tounta, G.; Antsaklis, A.; Mavrou, A.; Kolialexi, A. Application of proteomics for the identification of biomarkers in amniotic fluid: Are we ready to provide a reliable prediction? EPMA J. 2011, 2, 149–155. [Google Scholar] [CrossRef] [Green Version]
- Cameron, M.; Moran, P. Prenatal screening and diagnosis of neural tube defects. Prenat. Diagn. 2009, 29, 402–411. [Google Scholar] [CrossRef]
- Weiss, R.R.; Macri, J.N.; Elligers, K.; Princler, G.L.; McIntire, R.; Waldman, T.A. Amniotic fluid alpha-fetoprotein as a marker in prenatal diagnosis of neural tube defects. Obstet. Gynecol. 1976, 47, 148–151. [Google Scholar] [PubMed]
- Bradley, L.A.; Palomaki, G.E.; McDowell, G.A. Technical standards and guidelines: Prenatal screening for open neural tube defects. Genet. Med. 2005, 7, 355–369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Butler, H.J.B. An Atlas for Staging Mammalian and Chick Embryos, 1st ed.; CRC Press: Boca Raton, FL, USA, 1987. [Google Scholar]
- Minta, K.; Cullen, N.C.; Nimer, F.A.; Thelin, E.P.; Piehl, F.; Clarin, M.; Tullberg, M.; Jeppsson, A.; Portelius, E.; Zetterberg, H.; et al. Dynamics of extracellular matrix proteins in cerebrospinal fluid and serum and their relation to clinical outcome in human traumatic brain injury. Clin. Chem. Lab. Med. 2019, 57, 1565–1573. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Janik, K.; Smith, G.M.; Krynska, B. Identification of Neurocan and Phosphacan as Early Biomarkers for Open Neural Tube Defects. Cells 2023, 12, 1084. https://doi.org/10.3390/cells12071084
Janik K, Smith GM, Krynska B. Identification of Neurocan and Phosphacan as Early Biomarkers for Open Neural Tube Defects. Cells. 2023; 12(7):1084. https://doi.org/10.3390/cells12071084
Chicago/Turabian StyleJanik, Karolina, George M. Smith, and Barbara Krynska. 2023. "Identification of Neurocan and Phosphacan as Early Biomarkers for Open Neural Tube Defects" Cells 12, no. 7: 1084. https://doi.org/10.3390/cells12071084