
Copy Number Variation in Inflammatory Breast Cancer
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. IBC Patient Population
2.2. TCGA Study Population
2.3. Pathology Review and IBC Specimen Preparation
2.4. Dual DNA/RNA Extraction from Paraffin-Embedded IBC Biopsy Material
2.5. Quantification of RNA and DNA from Formalin-Fixed Paraffin-Embedded (FFPE) IBC Biopsy Material
2.6. Human Transcriptome Array
2.7. OncoScan Gene Chip
2.8. Quality Control and Biological Concordance Analysis
2.9. Statistical Analysis
3. Results
3.1. Inflammatory Breast Cancer Patients
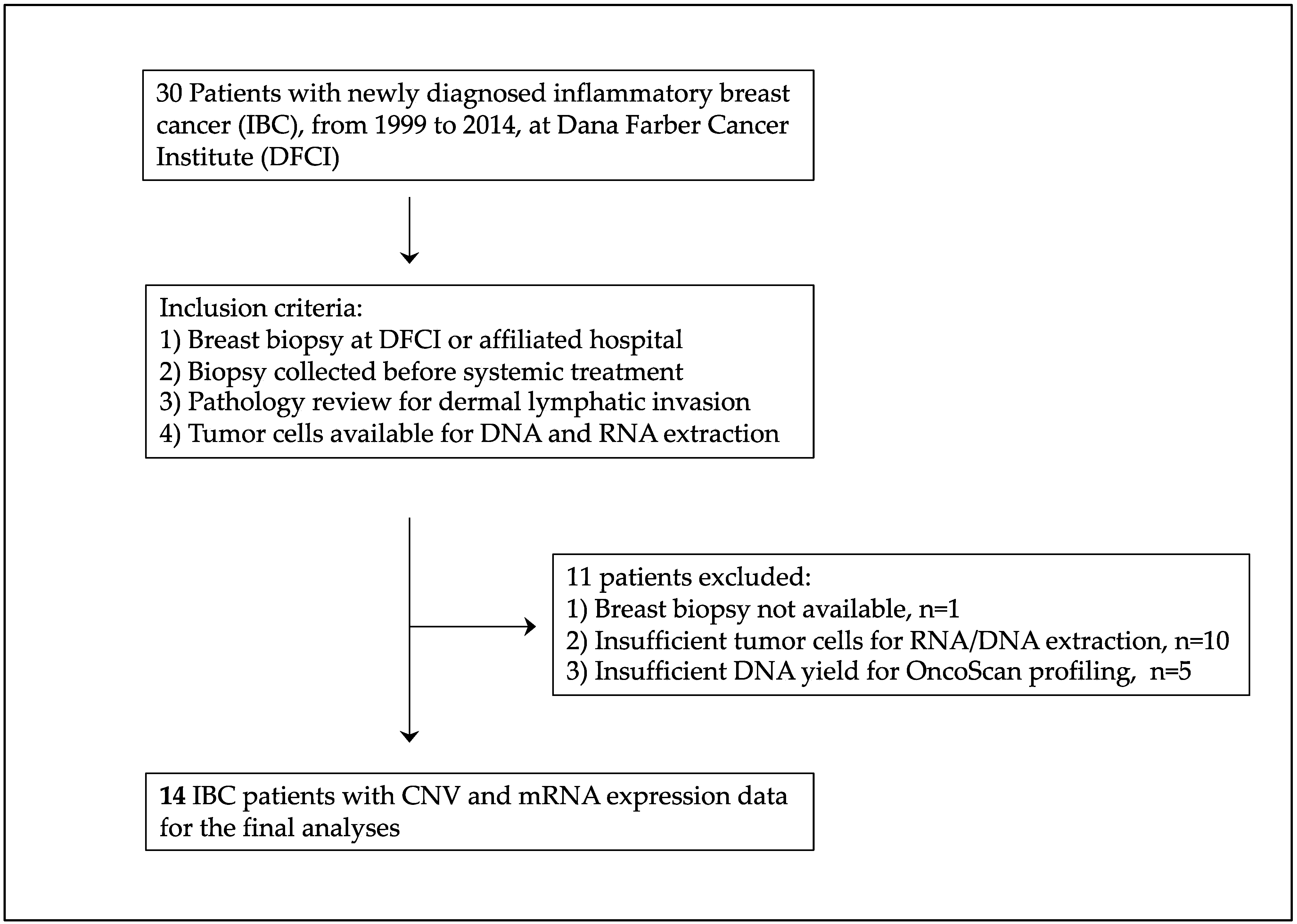
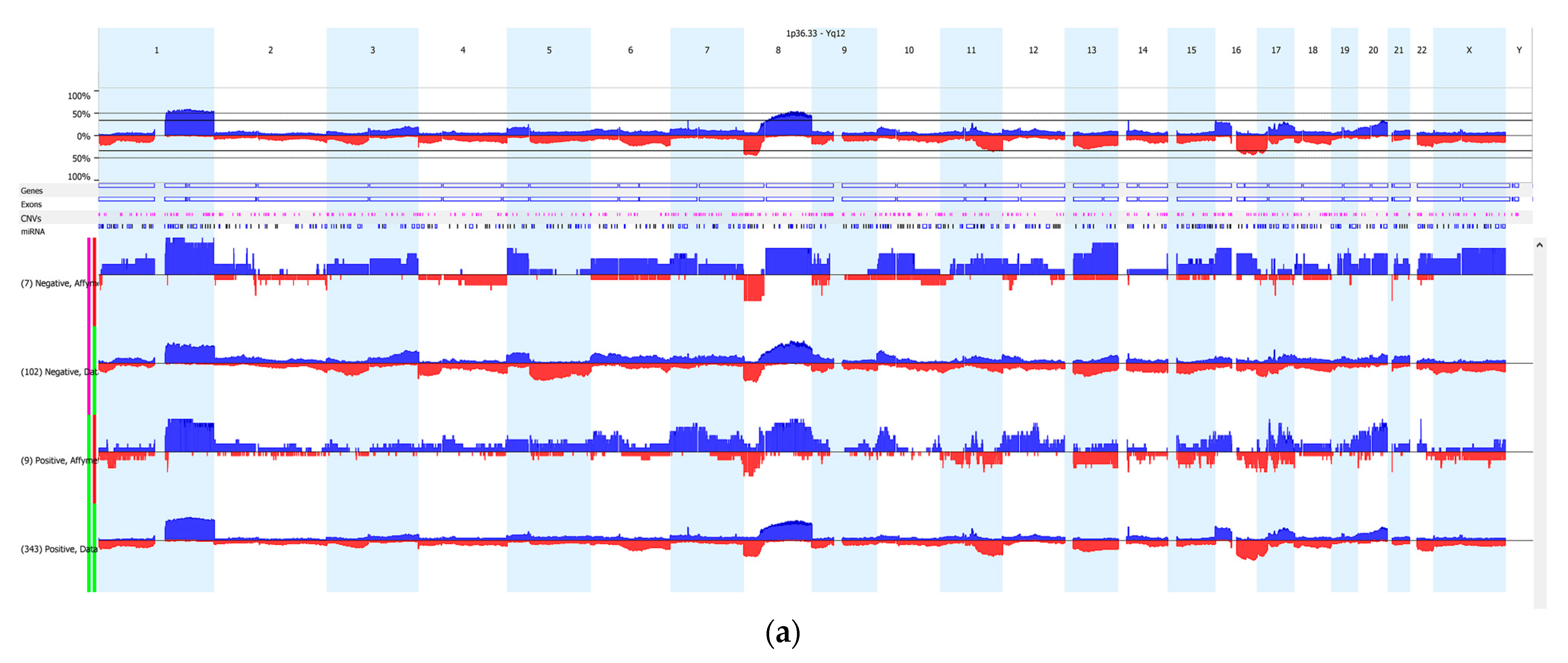
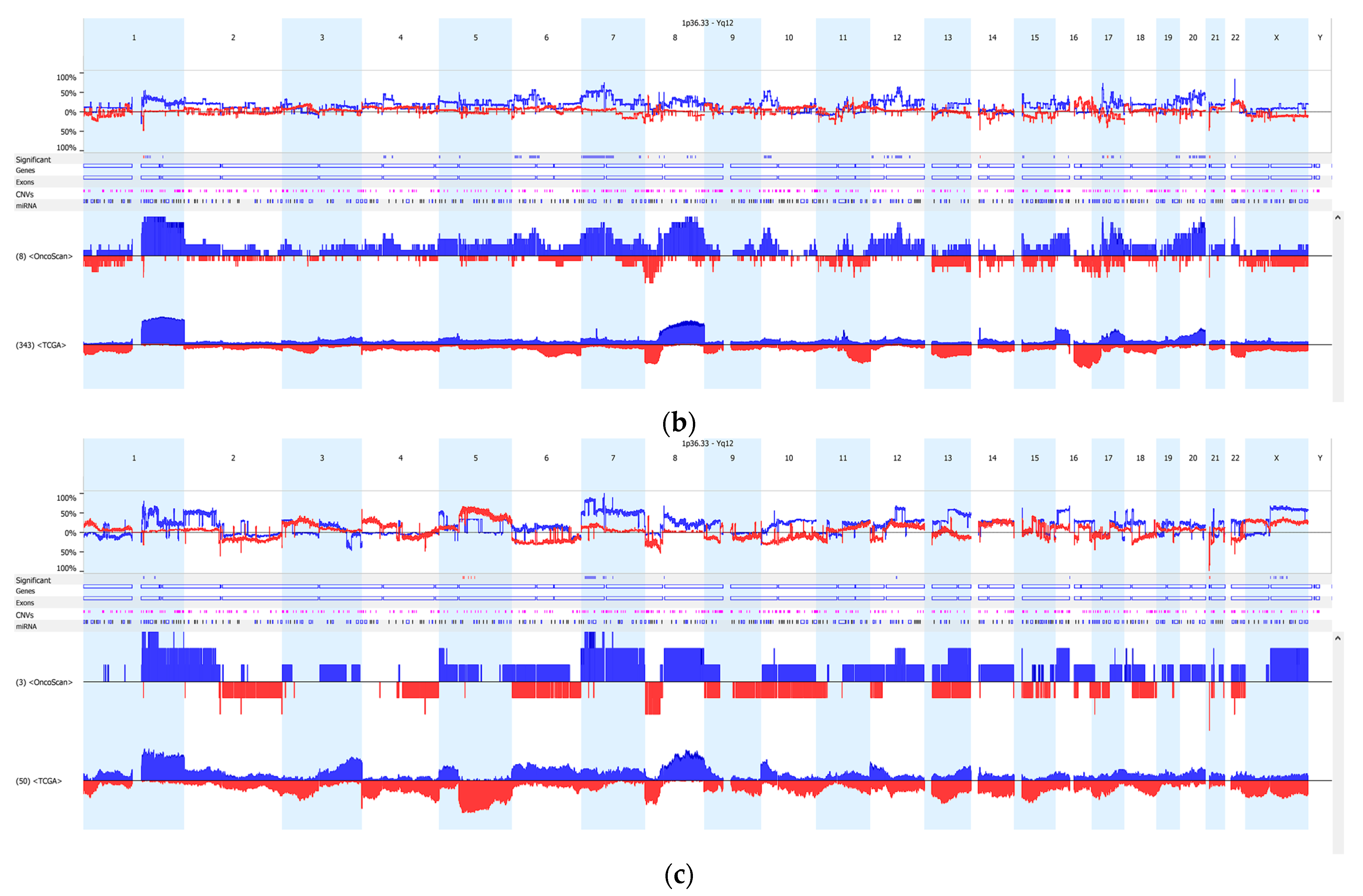
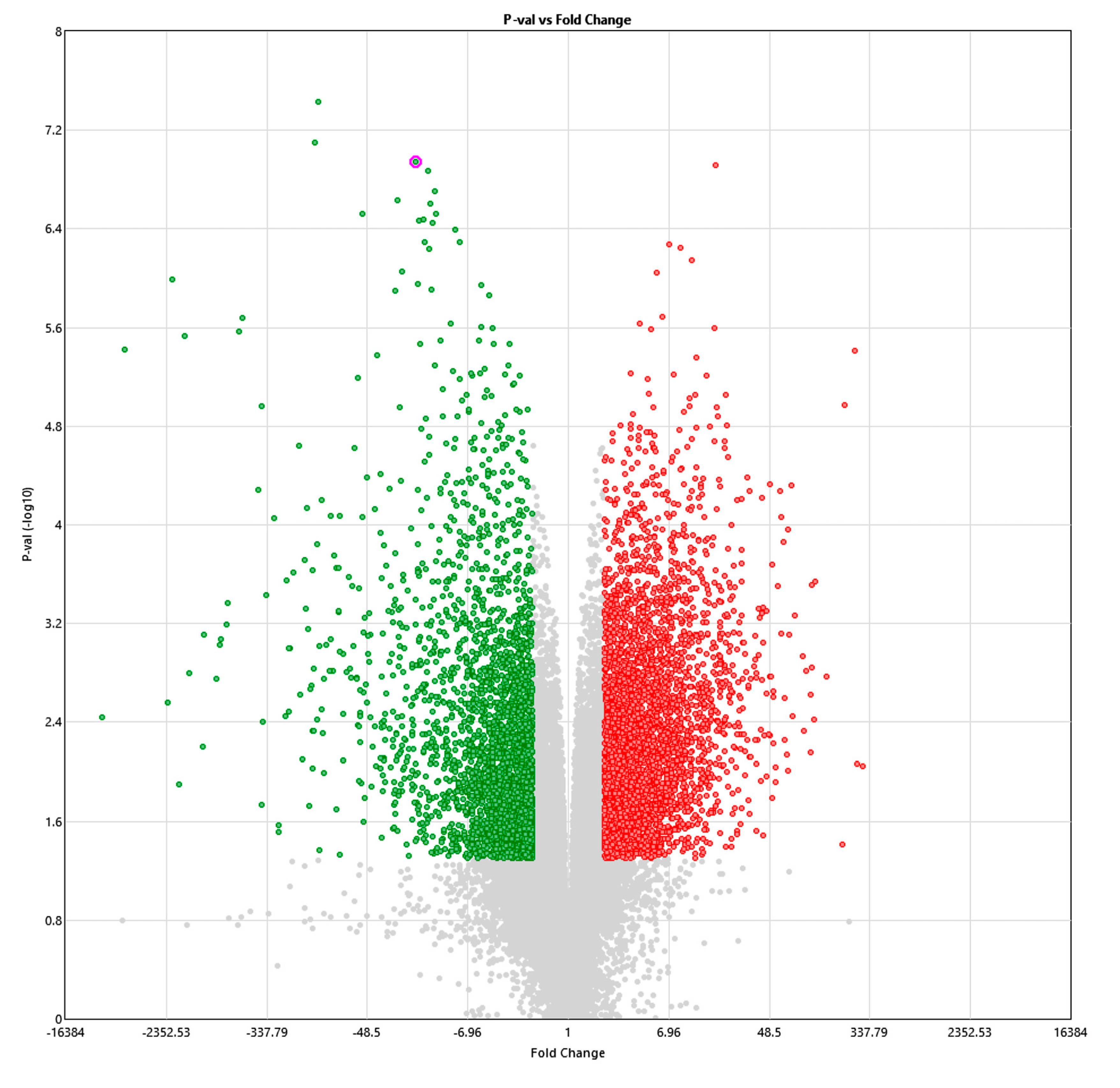
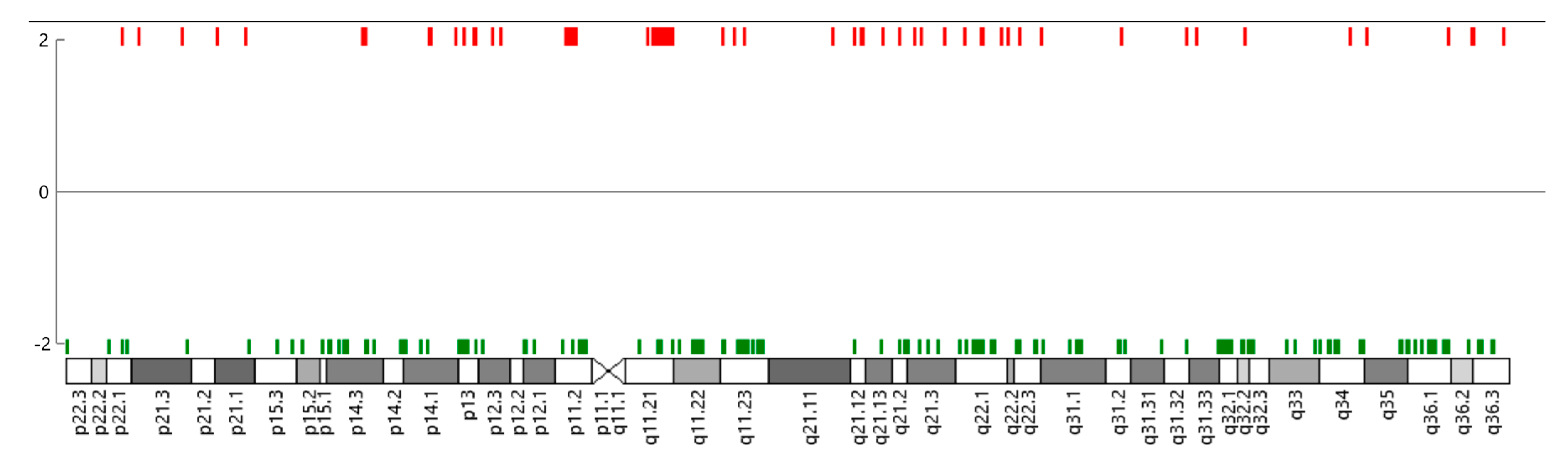
3.2. Copy Number Variation (CNV)
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hance, K.W.; Anderson, W.F.; Devesa, S.S.; Young, H.A.; Levine, P.H. Trends in Inflammatory Breast Carcinoma Incidence and Survival: The Surveillance, Epidemiology, and End Results Program at the National Cancer Institute. J. Natl. Cancer Inst. 2005, 97, 966–975. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Radunsky, G.S.; van Golen, K.L. The Current Understanding of the Molecular Determinants of Inflammatory Breast Cancer Metastasis. Clin. Exp. Metastasis 2005, 22, 615–620. [Google Scholar] [CrossRef] [PubMed]
- Robertson, F.M.; Bondy, M.; Yang, W.; Yamauchi, H.; Wiggins, S.; Kamrudin, S.; Krishnamurthy, S.; Le-Petross, H.; Bidaut, L.; Player, A.N.; et al. Inflammatory breast cancer: The Disease, the Biology, the Treatment. CA Cancer J. Clin. 2010, 60, 351–375. [Google Scholar] [CrossRef]
- Jagsi, R.; Mason, G.; Overmoyer, B.A.; Woodward, W.A.; Badve, S.; Schneider, R.J.; Lang, J.E.; Alpaugh, M.; Williams, K.P.; Vaught, D.; et al. Correction to: Inflammatory Breast Cancer Defined: Proposed Common Diagnostic Criteria to Guide Treatment and Research. Breast Cancer Res. Treat. 2022, 192, 245–247. [Google Scholar] [CrossRef]
- Fouad, T.M.; Kogawa, T.; Liu, D.D.; Shen, Y.; Masuda, H.; El-Zein, R.; Woodward, W.A.; Chavez-MacGregor, M.; Alvarez, R.H.; Arun, B.; et al. Erratum to: Overall Survival Differences between Patients with Inflammatory and Noninflammatory Breast Cancer Presenting with Distant Metastasis at Diagnosis. Breast Cancer Res. Treat. 2015, 152, 417. [Google Scholar] [CrossRef] [Green Version]
- Faldoni, F.L.C.; Villacis, R.A.R.; Canto, L.M.; Fonseca-Alves, C.E.; Cury, S.S.; Larsen, S.J.; Aagaard, M.M.; Souza, C.P.; Scapulatempo-Neto, C.; Osório, C.A.B.T.; et al. Inflammatory Breast Cancer: Clinical Implications of Genomic Alterations and Mutational Profiling. Cancers 2020, 12, 2816. [Google Scholar] [CrossRef] [PubMed]
- Bertucci, F.; Rypens, C.; Finetti, P.; Guille, A.; Adélaïde, J.; Monneur, A.; Carbuccia, N.; Garnier, S.; Dirix, P.; Gonçalves, A.; et al. NOTCH and DNA Repair Pathways Are More Frequently Targeted by Genomic Alterations in Inflammatory than in Non-Inflammatory Breast Cancers. Mol. Oncol. 2020, 14, 504–519. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Kumar, S.; Harmanci, A.; Li, S.; Kitchen, R.R.; Zhang, Y.; Wali, V.B.; Reddy, S.M.; Woodward, W.A.; Reuben, J.M.; et al. Whole-Genome Sequencing of Phenotypically Distinct Inflammatory Breast Cancers Reveals Similar Genomic Alterations to Non-Inflammatory Breast Cancers. Genome Med. 2021, 13, 70. [Google Scholar] [CrossRef]
- Woodward, W.; Krishnamurthy, S.; Yamauchi, H.; El-Zein, R.; Ogura, D.; Kitadai, E.; Niwa, S.-I.; Cristofanilli, M.; Vermeulen, P.; Dirix, L.; et al. Genomic and Expression Analysis of Microdissected Inflammatory Breast Cancer. Breast Cancer Res. Treat. 2013, 138, 761–772. [Google Scholar] [CrossRef]
- Rypens, C.; Marsan, M.; Van Berckelaer, C.; Billiet, C.; Melis, K.; Lopez, S.P.; van Dam, P.; Devi, G.R.; Finetti, P.; Ueno, N.T.; et al. Inflammatory Breast Cancer Cells Are Characterized by Abrogated Tgfbeta1-Dependent Cell Motility and SMAD3 Activity. Breast Cancer Res. Treat. 2020, 180, 385–395. [Google Scholar] [CrossRef]
- Van Laere, S.; Ueno, N.T.; Finetti, P.; Vermeulen, P.; Lucci, A.; Robertson, F.M.; Marsan, M.; Iwamoto, T.; Krishnamurthy, S.; Masuda, H.; et al. Uncovering the Molecular Secrets of Inflammatory Breast Cancer Biology: An Integrated Analysis of Three Distinct Affymetrix Gene Expression Datasets. Clin. Cancer Res. 2013, 19, 4685–4696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamm, C.A.; Moran, D.; Rao, K.; Trusk, P.B.; Pry, K.; Sausen, M.; Jones, S.; Velculesc, V.E.; Cristofanilli, M.; Bacus, S. Genomic and Immunological Tumor Profiling Identifies Targetable Pathways and Extensive CD8+/PDL1+ Immune Infiltration in Inflammatory Breast Cancer Tumors. Mol. Cancer Ther. 2016, 15, 1746–1756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Berckelaer, C.; Rypens, C.; Van Dam, P.; Pouillon, L.; Parizel, P.M.; Schats, K.A.; Kockx, M.; Tjalma, W.A.A.; Vermeulen, P.; Van Laere, S.; et al. Infiltrating Stromal Immune Cells in Inflammatory Breast Cancer Are Associated with an Improved Outcome and Increased PD-L1 Expression. Breast Cancer Res. 2019, 21, 28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arias-Pulido, H.; Cimino-Mathews, A.M.; Chaher, N.; Qualls, C.R.; Joste, N.; Colpaert, C.; Marotti, J.D.; Chamberlin, M.D.; Foisey, M.G.; Prossnitz, E.R.; et al. Differential Effects of CD20+ B Cells and PD-L1+ Immune Cells on Pathologic Complete Response and Outcome: Comparison between Inflammatory Breast Cancer and Locally Advanced Breast Cancer Patients. Breast Cancer Res. Treat. 2021, 190, 477–489. [Google Scholar] [CrossRef] [PubMed]
- Bekhouche, I.; Finetti, P.; Adelaïde, J.; Ferrari, A.; Tarpin, C.; Charafe-Jauffret, E.; Charpin, C.; Houvenaeghel, G.; Jacquemier, J.; Bidaut, G.; et al. High-Resolution Comparative Genomic Hybridization of Inflammatory Breast Cancer and Identification of Candidate Genes. PLoS ONE 2011, 6, e16950. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.H.; Lee, S.; Koo, J.S.; Jung, K.H.; Park, I.H.; Jeong, J.; Kim, S.I.; Park, S.; Park, H.S.; Park, B.-W.; et al. Anaplastic Lymphoma Kinase Gene Copy Number Gain in Inflammatory Breast Cancer (IBC): Prevalence, Clinicopathologic Features and Prognostic Implication. PLoS ONE 2015, 10, e0120320. [Google Scholar] [CrossRef] [Green Version]
- Bertucci, F.; Ueno, N.; Finetti, P.; Vermeulen, P.; Lucci, A.; Robertson, F.; Marsan, M.; Iwamoto, T.; Krishnamurthy, S.; Masuda, H.; et al. Gene Expression Profiles of Inflammatory Breast Cancer: Correlation with Response to Neoadjuvant Chemotherapy and Metastasis-Free Survival. Ann. Oncol. 2014, 25, 358–365. [Google Scholar] [CrossRef]
- Lim, B.; Woodward, W.A.; Wang, X.; Reuben, J.M.; Ueno, N.T. Author Correction: Inflammatory Breast Cancer Biology: The Tumour Microenvironment Is Key. Nat. Rev. Cancer 2018, 18, 526. [Google Scholar] [CrossRef]
- White, R.E.; Warren, L.; Nakhlis, F.; Rosenbluth, J.; Bellon, J.; Block, C.; Overmoyer, B. Characteristics Associated with Inflammatory Breast Cancer (IBC): An Epidemiologic Study from a Dedicated IBC Program. Breast J. 2020, 26, 1688–1694. [Google Scholar] [CrossRef]
- Thennavan, A.; Beca, F.; Xia, Y.; Garcia-Recio, S.; Allison, K.; Collins, L.C.; Tse, G.M.; Chen, Y.-Y.; Schnitt, S.J.; Hoadley, K.A.; et al. Molecular Analysis of Tcga Breast Cancer Histologic Types. Cell Genom. 2021, 1, 100067. [Google Scholar] [CrossRef]
- Mermel, C.H.; Schumacher, S.E.; Hill, B.; Meyerson, M.L.; Beroukhim, R.; Getz, G. GISTIC2.0 Facilitates Sensitive and Confident Localization of the Targets of Focal Somatic Copy-Number Alteration in Human Cancers. Genome Biol. 2011, 12, R41. [Google Scholar]
- Xu, W.; Seok, J.; Mindrinos, M.N.; Schweitzer, A.C.; Jiang, H.; Wilhelmy, J.; Clark, T.A.; Kapur, K.; Xing, Y.; Faham, M.; et al. Human Transcriptome Array for High-Throughput Clinical Studies. Proc. Natl. Acad. Sci. USA 2011, 108, 3707–3712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahomadegbe, J.C.; Tourpin, S.; Kaghad, M.; Zelek, L.; Vayssade, M.; Mathieu, M.C.; Rochard, F.; Spielmann, M.; Tursz, T.; Caput, D.; et al. Loss of Heterozygosity, Allele Silencing and Decreased Expression of P73 Gene in Breast Cancers: Prevalence of Alterations in Inflammatory Breast Cancers. Oncogene 2000, 19, 5413–5418. [Google Scholar] [CrossRef] [Green Version]
- Lerebours, F.; Bertheau, P.; Bieche, I.; Driouch, K.; de The, H.; Hacene, K.; Espie, M.; Marty, M.; Lidereau, R. Evidence of Chromosome Regions and Gene Involvement in Inflammatory Breast Cancer. Int. J. Cancer 2002, 102, 618–622. [Google Scholar] [CrossRef] [PubMed]
- Lerebours, F.; Bertheau, P.; Bieche, I.; Plassa, L.-F.; Champeme, M.-H.; Hacene, K.; Toulas, C.; Espie, M.; Marty, M.; Lidereau, R. Two Prognostic Groups of Inflammatory Breast Cancer Have Distinct Genotypes. Clin. Cancer Res. 2003, 9, 4184–4189. [Google Scholar]
- Luo, R.; Chong, W.; Wei, Q.; Zhang, Z.; Wang, C.; Ye, Z.; Abu-Khalaf, M.M.; Silver, D.P.; Stapp, R.T.; Jiang, W.; et al. Whole-Exome Sequencing Identifies Somatic Mutations and Intratumor Heterogeneity in Inflammatory Breast Cancer. NPJ Breast Cancer 2021, 7, 72. [Google Scholar] [CrossRef]
- Robertson, F.M.; Iii, E.F.P.; Van Laere, S.J.; Bertucci, F.; Chu, K.; Fernandez, S.V.; Mu, Z.; Alpaugh, K.; Pei, J.; Circo, R.; et al. Presence of Anaplastic Lymphoma Kinase in Inflammatory Breast Cancer. Springerplus 2013, 2, 497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ross, J.S.; Ali, S.M.; Wang, K.; Khaira, D.; Palma, N.A.; Chmielecki, J.; Palmer, G.A.; Morosini, D.; Elvin, J.A.; Fernandez, S.V.; et al. Comprehensive Genomic Profiling of Inflammatory Breast Cancer Cases Reveals a High Frequency of Clinically Relevant Genomic Alterations. Breast Cancer Res. Treat. 2015, 154, 155–162. [Google Scholar] [CrossRef]
- Tuma, R.S. ALK Gene Amplified in Most Inflammatory Breast Cancers. J. Natl. Cancer Inst. 2012, 104, 87–88. [Google Scholar] [CrossRef] [Green Version]
- Krishnamurthy, S.; Woodward, W.; Yang, W.; Reuben, J.M.; Tepperberg, J.; Ogura, D.; Niwa, S.-I.; Huo, L.; Gong, Y.; El-Zein, R.; et al. Status of the Anaplastic Lymphoma Kinase (ALK) Gene in Inflammatory Breast Carcinoma. Springerplus 2013, 2, 409. [Google Scholar] [CrossRef] [Green Version]
- Cabioglu, N.; Gong, Y.; Islam, R.; Broglio, K.R.; Sneige, N.; Sahin, A.; Gonzalez-Angulo, A.M.; Morandi, P.; Bucana, C.; Hortobagyi, G.N.; et al. Expression of Growth Factor and Chemokine Receptors: New Insights in the Biology of Inflammatory Breast Cancer. Ann. Oncol. 2007, 18, 1021–1029. [Google Scholar] [CrossRef]
- Zhang, D.; LaFortune, T.A.; Krishnamurthy, S.; Esteva, F.J.; Cristofanilli, M.; Liu, P.; Lucci, A.; Singh, B.; Hung, M.-C.; Hortobagyi, G.N.; et al. Epidermal Growth Factor Receptor Tyrosine Kinase Inhibitor Reverses Mesenchymal to Epithelial Phenotype and Inhibits Metastasis in Inflammatory Breast Cancer. Clin. Cancer Res. 2009, 15, 6639–6648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ali, S.M.; Alpaugh, R.K.; Downing, S.R.; Stephens, P.J.; Yu, J.Q.; Wu, H.; Buell, J.K.; Miller, V.A.; Lipson, D.; Palmer, G.A.; et al. Response of an ERBB2-Mutated Inflammatory Breast Carcinoma to Human Epidermal Growth Factor Receptor 2-Targeted Therapy. J. Clin. Oncol. 2014, 32, e88–e91. [Google Scholar] [CrossRef] [Green Version]
- Mueller, K.L.; Yang, Z.-Q.; Haddad, R.; Ethier, S.P.; Boerner, J.L. EGFR/Met Association Regulates EGFR TKI Resistance in Breast Cancer. J. Mol. Signal. 2010, 5, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masuda, H.; A Baggerly, K.; Wang, Y.; Iwamoto, T.; Brewer, T.; Pusztai, L.; Kai, K.; Kogawa, T.; Finetti, P.; Birnbaum, D.; et al. Comparison of Molecular Subtype Distribution in Triple-Negative Inflammatory and Non-Inflammatory Breast Cancers. Breast Cancer Res. 2013, 15, R112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masuda, H.; Zhang, D.; Bartholomeusz, C.; Doihara, H.; Hortobagyi, G.N.; Ueno, N.T. Role of Epidermal Growth Factor Receptor in Breast Cancer. Breast Cancer Res. Treat. 2012, 136, 331–345. [Google Scholar] [CrossRef] [Green Version]
- Matsuda, N.; Wang, X.; Lim, B.; Krishnamurthy, S.; Alvarez, R.H.; Willey, J.S.; Parker, C.A.; Song, J.; Shen, Y.; Hu, J.; et al. Safety and Efficacy of Panitumumab Plus Neoadjuvant Chemotherapy in Patients with Primary HER2-Negative Inflammatory Breast Cancer. JAMA Oncol. 2018, 4, 1207–1213. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Semba, T.; Manyam, G.C.; Wang, J.; Shao, S.; Bertucci, F.; Finetti, P.; Krishnamurthy, S.; Phi, L.T.H.; Pearson, T.; et al. EGFR Is a Master Switch between Immunosuppressive and Immunoactive Tumor Microenvironment in Inflammatory Breast Cancer. Sci. Adv. 2022, 8, eabn7983. [Google Scholar] [CrossRef]
- Selenz, C.; Compes, A.; Nill, M.; Borchmann, S.; Odenthal, M.; Florin, A.; Brägelmann, J.; Büttner, R.; Meder, L.; Ullrich, R.T. EGFR Inhibition Strongly Modulates the Tumour Immune Microenvironment in EGFR-Driven Non-Small-Cell Lung Cancer. Cancers 2022, 14, 3943. [Google Scholar] [CrossRef]
- Yang, L.; He, Y.-T.; Dong, S.; Wei, X.-W.; Chen, Z.-H.; Zhang, B.; Chen, W.-D.; Yang, X.-R.; Wang, F.; Shang, X.-M.; et al. Single-Cell Transcriptome Analysis Revealed a Suppressive Tumor Immune Microenvironment in EGFR Mutant Lung Adenocarcinoma. J. Immunother. Cancer 2022, 10, e003534. [Google Scholar] [CrossRef]
- Renault, A.-L.; Mebirouk, N.; Fuhrmann, L.; Bataillon, G.; Cavaciuti, E.; Le Gal, D.; Girard, E.; Popova, T.; La Rosa, P.; Beauvallet, J.; et al. Morphology and Genomic Hallmarks of Breast Tumours Developed by ATM Deleterious Variant Carriers. Breast Cancer Res. 2018, 20, 28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwon, M.J.; Kim, R.N.; Song, K.; Jeon, S.; Jeong, H.M.; Kim, J.S.; Han, J.; Hong, S.; Oh, E.; Choi, J.-S.; et al. Genes Co-Amplified with ERBB2 or MET as Novel Potential Cancer-Promoting Genes in Gastric Cancer. Oncotarget 2017, 8, 92209–92226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quiroz-Zárate, A.; Harshfield, B.J.; Hu, R.; Knoblauch, N.; Beck, A.H.; Hankinson, S.E.; Carey, V.; Tamimi, R.M.; Hunter, D.J.; Quackenbush, J.; et al. Expression Quantitative Trait Loci (Qtl) in Tumor Adjacent Normal Breast Tissue and Breast Tumor Tissue. PLoS ONE 2017, 12, e0170181. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ER + IBC (N = 10) | ER − IBC (N = 4) | |
|---|---|---|
| N (%) | N (%) | |
| Age at diagnosis, Mean years (SD) | 53.3 (6.7) | 53.5 (4.0) |
| Grade | ||
| II | 3 (30) | 1 (25) |
| III | 7 (70) | 3 (75) |
| % Dermal Lymphatic Invasion | 6 (60) | 1 (25) |
| % Family History of Breast Cancer | 6 (60) | 0 (0) |
| Body Mass Index, Mean kg/m2 (SD) | 29.1 (11.2) | 26.5 (3.4) |
| Region | Cytoband Location | Event | Region Length ** | Frequency in IBC (%) * | Frequency in non-IBC (%)* | Difference | p-Value |
|---|---|---|---|---|---|---|---|
| ER+ (n=8 IBC, 343 non-IBC) | |||||||
| chr7:58,019,983-58,025,423 | p11.1 | CN Gain | 5440 | 75 | 0 | 75 | 1.13 × 10−11 |
| chr7:57,950,944-58,019,983 | p11.2–p11.1 | CN Gain | 69039 | 75 | 0.58 | 74.42 | 3.12 × 10−10 |
| chr21:9,648,315-9,764,385 | p11.2 | CN Loss | 116070 | 50 | 0 | 50 | 1.13 × 10−07 |
| chr1:149,248,784-149,293,460 | q21.2 | CN Loss | 44676 | 50 | 0.29 | 49.71 | 5.58 × 10−07 |
| TN (n=3 IBC, 50 non-IBC) | |||||||
| chr7:57,950,944-58,025,423 | p11.2–p11.1 | CN Gain | 74479 | 100 | 0 | 100 | 4.27 × 10−05 |
| chr21:9,648,315-9,764,385 | p11.2 | CN Loss | 116070 | 100 | 0 | 100 | 4.27 × 10−05 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hazra, A.; O’Hara, A.; Polyak, K.; Nakhlis, F.; Harrison, B.T.; Giordano, A.; Overmoyer, B.; Lynce, F. Copy Number Variation in Inflammatory Breast Cancer. Cells 2023, 12, 1086. https://doi.org/10.3390/cells12071086
Hazra A, O’Hara A, Polyak K, Nakhlis F, Harrison BT, Giordano A, Overmoyer B, Lynce F. Copy Number Variation in Inflammatory Breast Cancer. Cells. 2023; 12(7):1086. https://doi.org/10.3390/cells12071086
Chicago/Turabian StyleHazra, Aditi, Andrea O’Hara, Kornelia Polyak, Faina Nakhlis, Beth T. Harrison, Antonio Giordano, Beth Overmoyer, and Filipa Lynce. 2023. "Copy Number Variation in Inflammatory Breast Cancer" Cells 12, no. 7: 1086. https://doi.org/10.3390/cells12071086