
Lyophilized Extracellular Vesicles from Adipose-Derived Stem Cells Increase Muscle Reperfusion but Degrade Muscle Structural Proteins in a Mouse Model of Hindlimb Ischemia-Reperfusion Injury
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Exosome Isolation and Preparation
2.2. Exosome Protein Extraction and Digestion for LC-MS Analysis
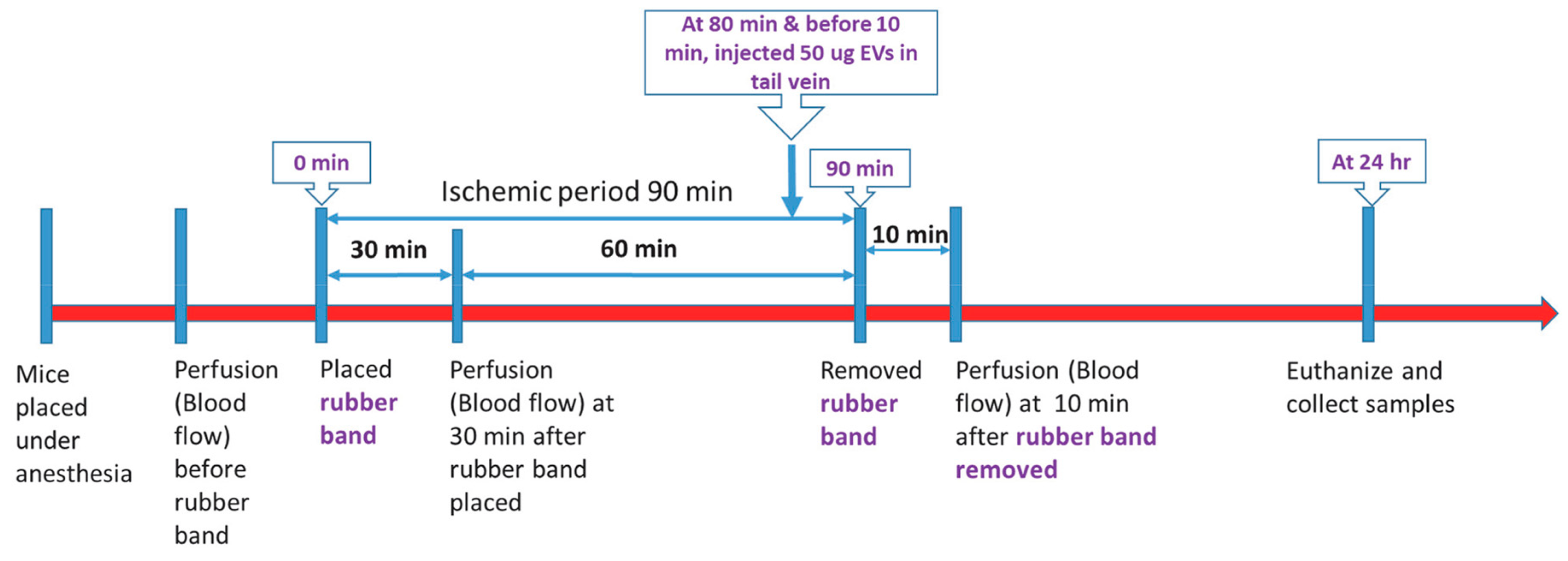
2.3. Ischemia-Reperfusion Experiments
2.4. Sample Collection and Tissue Analysis
2.5. Analysis of Inflammatory Cytokines
2.6. Proteomic Analysis of Skeletal Muscle
2.7. Statistical Analysis
3. Results
3.1. Proteomic Characterization of EV Cargo
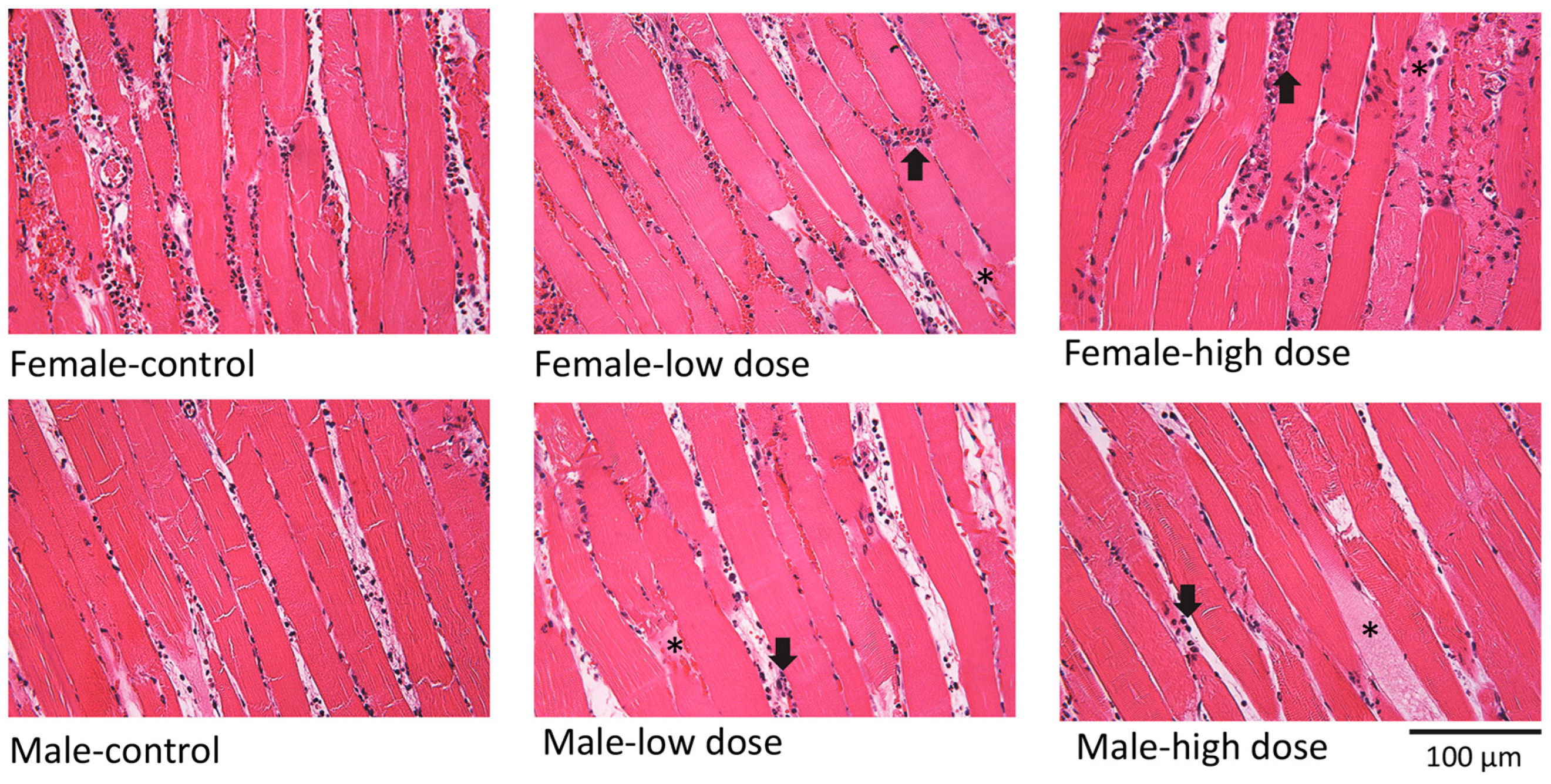
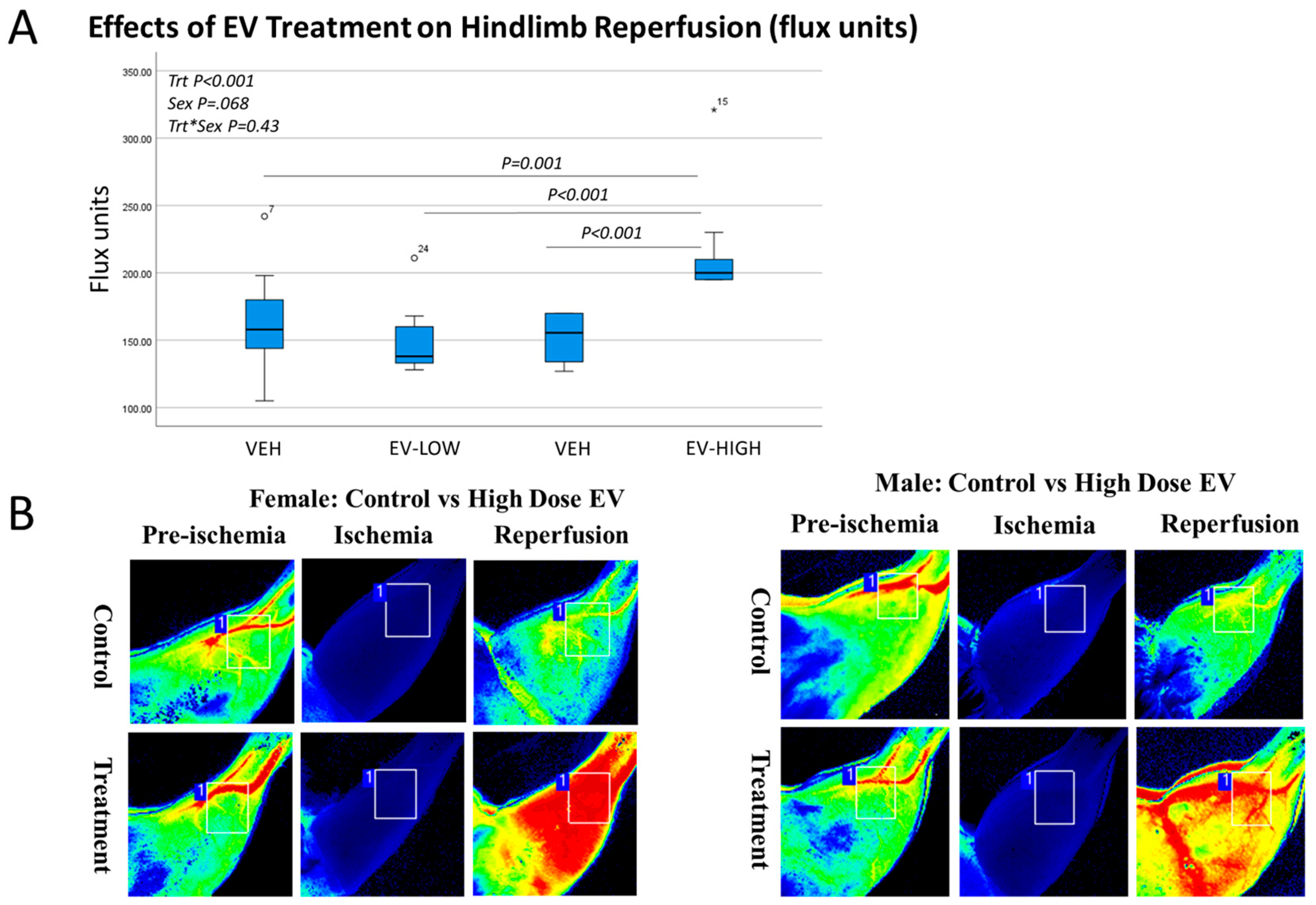
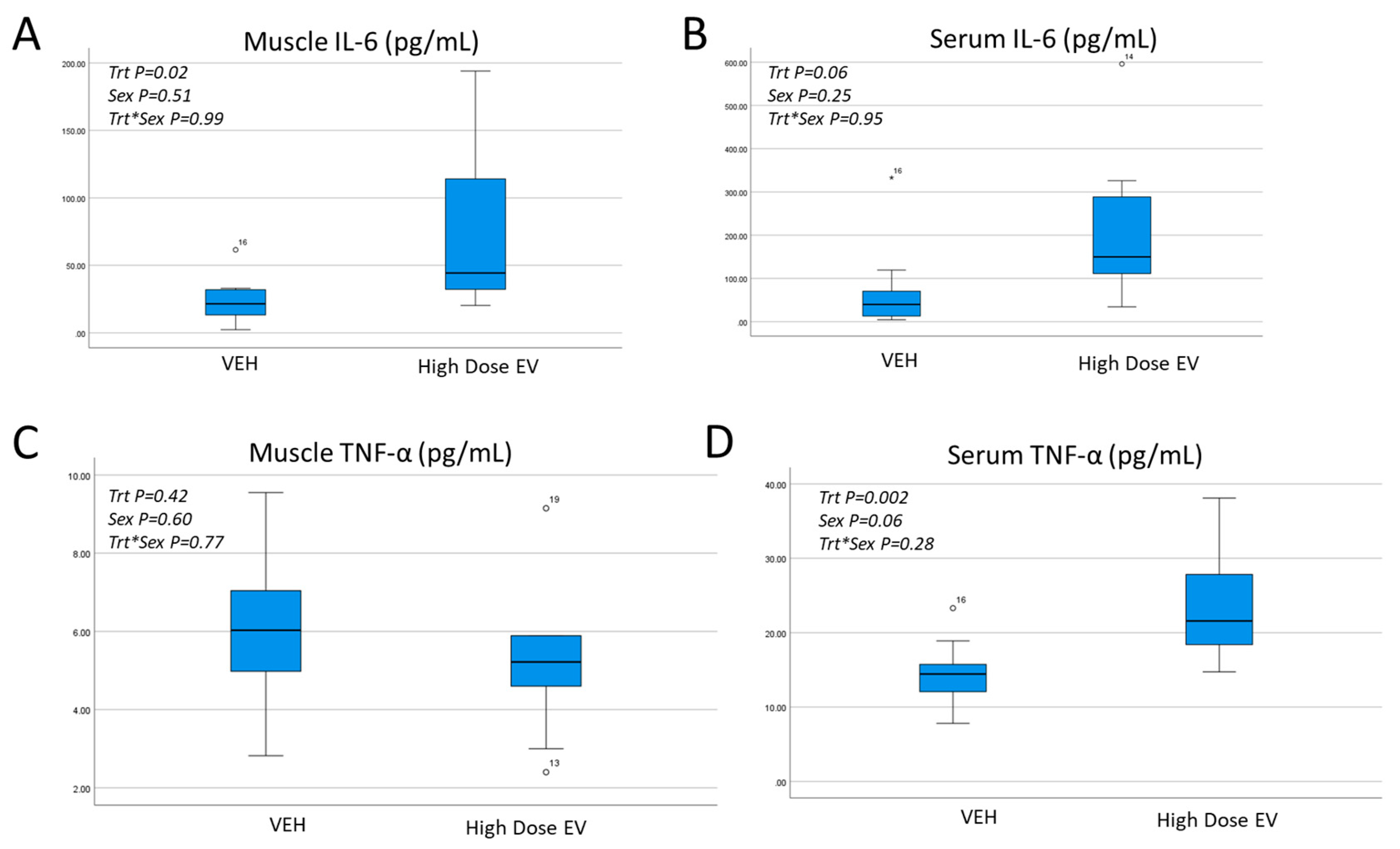
3.2. Effects of EV Treatment on Muscle Histology, Reperfusion, and Inflammatory Cytokines
3.3. Effects of High Dose EV Treatment on Muscle Proteins following Ischemia and Reperfusion
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lejay, A.; Meyer, A.; Schlagowski, A.I.; Charles, A.L.; Singh, F.; Bouitbir, J.; Pottecher, J.; Chakfé, N.; Zoll, J.; Geny, B. Mitochondria: Mitochondrial participation in ischemia-reperfusion injury in skeletal muscle. Int. J. Biochem. Cell Biol. 2014, 50, 101–105. [Google Scholar] [CrossRef] [PubMed]
- Pizzimenti, M.; Riou, M.; Charles, A.-L.; Talha, S.; Meyer, A.; Andres, E.; Chakfé, N.; Lejay, A.; Geny, B. The Rise of Mitochondria in Peripheral Arterial Disease Physiopathology: Experimental and Clinical Data. J. Clin. Med. 2019, 8, 2125. [Google Scholar] [CrossRef] [PubMed]
- Mittal, M.; Siddiqui, M.R.; Tran, K.; Reddy, S.P.; Malik, A.B. Reactive Oxygen Species in Inflammation and Tissue Injury. Antioxid. Redox Signal. 2014, 20, 1126–1167. [Google Scholar] [CrossRef] [PubMed]
- Garbaisz, D.; Turoczi, Z.; Aranyi, P.; Fulop, A.; Rosero, O.; Hermesz, E.; Ferencz, A.; Lotz, G.; Harsanyi, L.; Szijarto, A. Attenuation of Skeletal Muscle and Renal Injury to the Lower Limb following Ischemia-Reperfusion Using mPTP Inhibitor NIM-811. PLoS ONE 2014, 9, e101067. [Google Scholar] [CrossRef]
- El Baradie, K.B.Y.; Khan, M.B.; Mendhe, B.; Waller, J.; O’Brien, F., 3rd; Hamrick, M.W. The cyclophilin inhibitor NIM-811 increases muscle cell survival with hypoxia in vitro and improves gait performance following ischemia-reperfusion in vivo. Sci Rep. 2021, 11, 6152. [Google Scholar] [CrossRef]
- Arslan, F.; Lai, R.C.; Smeets, M.B.; Akeroyd, L.; Choo, A.; Aguor, E.N.; Timmers, L.; van Rijen, H.V.; Doevendans, P.A.; Pasterkamp, G. Mesenchymal stem cell-derived exosomes increase ATP levels, decrease oxidative stress and activate Pi3K/Akt pathway to enhance myocardial viability and prevent adverse remodeling after myocardial ischemia/reperfusion injury. Stem Cell Res. 2013, 10, 301–312. [Google Scholar]
- Potz, B.A.; Scrimgeour, L.A.; Pavlov, V.I.; Sodha, N.R.; Abid, M.R.; Sellke, F.W. Extracellular Vesicle Injection Improves Myocardial Function and Increases Angiogenesis in a Swine Model of Chronic Ischemia. J. Am. Hear. Assoc. 2018, 7, e008344. [Google Scholar] [CrossRef]
- Johnson, T.K.; Zhao, L.; Zhu, D.; Wang, Y.; Xiao, Y.; Oguljahan, B.; Zhao, X.; Kirlin, W.G.; Yin, L.; Chilian, W.M.; et al. Exosomes derived from induced vascular progenitor cells promote angiogenesis in vitro and in an in vivo rat hindlimb ischemia model. Am. J. Physiol. Circ. Physiol. 2019, 317, H765–H776. [Google Scholar] [CrossRef]
- Zhang, H.; Liu, X.-B.; Huang, S.; Bi, X.-Y.; Wang, H.-X.; Xie, L.-X.; Wang, Y.-Q.; Cao, X.-F.; Lv, J.; Xiao, F.-J.; et al. Microvesicles Derived from Human Umbilical Cord Mesenchymal Stem Cells Stimulated by Hypoxia Promote Angiogenesis Both In Vitro and In Vivo. Stem Cells Dev. 2012, 21, 3289–3297. [Google Scholar] [CrossRef]
- El Baradie, K.B.Y.; Nouh, M.; O’Brien Iii, F.; Liu, Y.; Fulzele, S.; Eroglu, A.; Hamrick, M.W. Freeze-Dried Extracellular Vesicles from Adipose-Derived Stem Cells Prevent Hypoxia-Induced Muscle Cell Injury. Front. Cell Dev. Biol. 2020, 8, 181. [Google Scholar] [CrossRef]
- Bari, E.; Perteghella, S.; Di Silvestre, D.; Sorlini, M.; Catenacci, L.; Sorrenti, M.; Marrubini, G.; Rossi, R.; Tripodo, G.; Mauri, P.; et al. Pilot Production of Mesenchymal Stem/Stromal Freeze-Dried Secretome for Cell-Free Regenerative Nanomedicine: A Validated GMP-Compliant Process. Cells 2018, 7, 190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ge, Q.; Zhou, Y.; Lu, J.; Bai, Y.; Xie, X.; Lu, Z. miRNA in Plasma Exosome is Stable under Different Storage Conditions. Molecules 2014, 19, 1568–1575. [Google Scholar] [CrossRef] [PubMed]
- Crawford, R.S.; Hashmi, F.F.; Jones, J.E.; Albadawi, H.; McCormack, M.; Eberlin, K.; Entabi, F.; Atkins, M.D.; Conrad, M.F.; Austen, W.G.; et al. A novel model of acute murine hindlimb ischemia. Am. J. Physiol. Circ. Physiol. 2007, 292, H830–H837. [Google Scholar] [CrossRef]
- Drysch, M.; Wallner, C.; Schmidt, S.V.; Reinkemeier, F.; Wagner, J.M.; Lehnhardt, M.; Behr, B. An optimized low-pressure tourniquet murine hind limb ischemia reperfusion model: Inducing acute ischemia reperfusion injury in C57BL/6 wild type mice. PLoS ONE 2019, 14, e0210961. [Google Scholar] [CrossRef]
- Mathivanan, S.; Ji, H.; Simpson, R.J. Exosomes: Extracellular organelles important in intercellular communication. J. Proteom. 2010, 73, 1907–1920. [Google Scholar] [CrossRef] [PubMed]
- Stamer, W.; Hoffman, E.; Luther, J.; Hachey, D.; Schey, K. Protein profile of exosomes from trabecular meshwork cells. J. Proteom. 2011, 74, 796–804. [Google Scholar] [CrossRef]
- Xing, X.; Han, S.; Cheng, G.; Ni, Y.; Li, Z.; Li, Z. Proteomic Analysis of Exosomes from Adipose-Derived Mesenchymal Stem Cells: A Novel Therapeutic Strategy for Tissue Injury. BioMed Res. Int. 2020, 2020, 6094562. [Google Scholar] [CrossRef] [PubMed]
- Kupatt, C.; Dessy, C.; Hinkel, R.; Raake, P.; Daneau, G.; Bouzin, C.; Boekstegers, P.; Feron, O. Heath shock protein 90 transfection reduces ischemia-reperfusion-induced myocardial dysfunction via reciprocal endothelial NO synthase serine 1177 phosphorylation and threonine 495 dephosphorylation. Arterioscler. Thromb. Vasc. Biol. 2004, 24, 1435–1441. [Google Scholar]
- Presley, T.; Vedam, K.; Velayutham, M.; Zweier, J.L.; Ilangovan, G. Activation of Hsp90-eNOS and increased NO generation attenuate respiration of hypoxia-treated endothelial cells. Am. J. Physiol. Cell Physiol. 2008, 295, C1281–C1291. [Google Scholar] [PubMed]
- Purvis, G.; Solito, E.; Thiemermann, C. Annexin-A1: Therapeutic potential in microvascular disease. Front. Immunol. 2019, 10, 3389. [Google Scholar] [CrossRef]
- Armstrong, S.C.; Latham, C.A.; Shivell, C.L.; Ganote, C.E. Ischemic loss of sarcolemmal dystrophin and spectrin: Correlation with myocardial injury. J. Mol. Cell Cardiol. 2001, 33, 1165–1179. [Google Scholar] [PubMed]
- Rodríguez, M.; Cai, W.-J.; Kostin, S.; Lucchesi, B.R.; Schaper, J. Ischemia depletes dystrophin and inhibits protein synthesis in the canine heart: Mechanisms of myocardial ischemic injury. J. Mol. Cell. Cardiol. 2005, 38, 723–733. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Geist, J.; Grogan, A.; Hu, L.R.; Kontrogianni-Konstantopoulos, A. Thick Filament Protein Network, Functions, and Disease Association. Compr. Physiol. 2018, 8, 631–709. [Google Scholar] [PubMed]
- Zrelski, M.M.; Kustermann, M.; Winter, L. Muscle-Related Plectinopathies. Cells 2021, 10, 2480. [Google Scholar] [CrossRef] [PubMed]
- Paulin, D.; Hovhannisyan, Y.; Kasakyan, S.; Agbulut, O.; Li, Z.; Xue, Z. Synemin-related skeletal and cardiac myopathies: An overview of pathogenic variants. Am. J. Physiol. Physiol. 2020, 318, C709–C718. [Google Scholar] [CrossRef]
- Naito, H.; Nojima, T.; Fujisaki, N.; Tsukahara, K.; Yamamoto, H.; Yamada, T.; Aokage, T.; Yumoto, T.; Osako, T.; Nakao, A. Therapeutic strategies for ischemia reperfusion injury in emergency medicine. Acute Med. Surg. 2020, 7, e501. [Google Scholar] [CrossRef]
- Fischesser, D.M.; Bo, B.; Benton, R.P.; Su, H.; Jahanpanah, N.; Haworth, K.J. Controlling Reperfusion Injury with Controlled Reperfusion: Historical Perspectives and New Paradigms. J. Cardiovasc. Pharmacol. Ther. 2021, 26, 504–523. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Symbol | UniProt Id | Sequence Name |
|---|---|---|
| Fn1 * | P11276 | Fibronectin-1 |
| Col12a1 | Q60847 | Collagen-1 alpha XII |
| Col1a2 | Q01149 | Collagen 1A2 |
| Tnc | Q80YX1 | Tenascin C |
| Lgals3bp | Q07797 | Galectin 3 binding protein |
| Postn | Q62009 | Periostin |
| Thbs2 * | Q03350 | Thrombospondin-2 |
| Myh9 * | Q8VDD5 | Myosin-9 |
| Lum | P51885 | Lumican |
| Lrp1 | Q91ZX7 | Prolow-density lipoprotein receptor-related protein 1 |
| Bgn | P28653 | Biglycan |
| Gsn * | P13020 | Gelsolin |
| Msn * | P26041 | Moesin |
| Hsp90ab1 | P11499 | Heat shock protein HSP 90-beta |
| Vcan | Q62059 | Versican |
| Hsp90b1 | P11499 | Endoplasmin |
| Hsp90aa1 * | P07901 | Heat shock protein HSP 90-alpha |
| Eno1 * | P17182 | Alpha-enolase |
| Pros1 | Q08761 | Vitamin K-dependent protein S (Pros1) |
| App | P12023 | Amyloid-beta A4 protein |
| Lbp | Q61805 | Lipopolysaccharide-binding protein |
| Calr | P14211 | Calreticulin |
| Cltc * | Q68FD5 | Clathrin heavy chain 1 |
| Nid1 | P10493 | Nidogen 1 |
| Fbln2 | P37889 | Fibulin 2 |
| Col1a1 | P11087 | Collagen alpha-1 (I) |
| Variable | Treatment | Sex | Treatment*Sex |
|---|---|---|---|
| Reperfusion (flux units) | p = 0.43 | 0.89 | 0.224 |
| Muscle TNF alpha | p = 0.19 | <0.001 | 0.09 |
| Muscle IL1 alpha | p = 0.09 | 0.68 | 0.39 |
| Muscle IL6 | p = 0.17 | 0.01 | 0.63 |
| Serum TNF alpha | p = 0.87 | 0.35 | 0.16 |
| Serum IL1 alpha | p = 0.58 | 0.72 | 0.22 |
| Serum IL6 | p = 0.40 | 0.23 | 0.72 |
| Acute Inflammatory Response | Wound Healing | ||
|---|---|---|---|
| Gene symbol | Gene name | Gene symbol | Gene name |
| Fga (FC = 2.74 ***) | fibrinogen alpha chain | Fga (FC = 2.74 ***) | fibrinogen alpha chain |
| Apcs (FC = 2.54 **) | amyloid P component, serum | Fgb (FC = 2.9 ***) | fibrinogen beta chain |
| Orm1 (FC = 3.3 ***) | orosomucoid 1 | Apcs (FC = 2.54 **) | amyloid P component, serum |
| Serpinc1 (FC = 2.48 **) | serpin family C member 1 | S100a9 (FC = 2.9 **) | S100 calcium-binding protein A9 |
| C3 (FC = 2.37 **) | complement C3 | Serpinc1 (FC = 2.48 **) | serpin family C member 1 |
| Fn1 (FC = 5.0 **) | fibronectin 1 | C3 (FC = 2.37 **) | complement C3 |
| Serpinf2 (FC = 2.6 ***) | serpin family F member 2 | Fgg (FC = 4.1 ***) | fibrinogen gamma chain |
| Pzp (FC = 4.6 **) | PZP alpha-2-macroglobulin like | Plg (FC = 3.5 **) | plasminogen |
| Hp (FC = 3.4 **) | haptoglobin | Fn1 Fn1 (FC = 5.0 **) | fibronectin 1 |
| Itih4 (FC = 2.8 **) | inter-alpha-trypsin inhibitor heavy chain 4 | Serpinf2 (FC = 2.6 ***) | serpin family F member 2 |
| Anxa1 (FC = 6.7 ***) | annexin A1 | ||
| Gsn (FC = 4.7 **) | gelsolin | ||
| Muscle Structural Proteins | |
|---|---|
| Gene symbol | Gene name |
| Tnni2 (FC = 0.69 **) | troponin I, fast skeletal muscle |
| Dmd (FC = 0.48 **) | dystrophin |
| Plec (FC = 0.44 ***) | plectin |
| Obscn (FC = 0.42 **) | obscurin |
| Synm (FC = 0.43 ***) | synemin |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mendhe, B.; Khan, M.B.; Dunwody, D.; El Baradie, K.B.Y.; Smith, K.; Zhi, W.; Sharma, A.; Lee, T.J.; Hamrick, M.W. Lyophilized Extracellular Vesicles from Adipose-Derived Stem Cells Increase Muscle Reperfusion but Degrade Muscle Structural Proteins in a Mouse Model of Hindlimb Ischemia-Reperfusion Injury. Cells 2023, 12, 557. https://doi.org/10.3390/cells12040557
Mendhe B, Khan MB, Dunwody D, El Baradie KBY, Smith K, Zhi W, Sharma A, Lee TJ, Hamrick MW. Lyophilized Extracellular Vesicles from Adipose-Derived Stem Cells Increase Muscle Reperfusion but Degrade Muscle Structural Proteins in a Mouse Model of Hindlimb Ischemia-Reperfusion Injury. Cells. 2023; 12(4):557. https://doi.org/10.3390/cells12040557
Chicago/Turabian StyleMendhe, Bharati, Mohammad B. Khan, Damon Dunwody, Khairat Bahgat Youssef El Baradie, Kathryn Smith, Wenbo Zhi, Ashok Sharma, Tae Jin Lee, and Mark W. Hamrick. 2023. "Lyophilized Extracellular Vesicles from Adipose-Derived Stem Cells Increase Muscle Reperfusion but Degrade Muscle Structural Proteins in a Mouse Model of Hindlimb Ischemia-Reperfusion Injury" Cells 12, no. 4: 557. https://doi.org/10.3390/cells12040557