
Cellular and Molecular Mechanisms Associating Obesity to Bone Loss

Abstract
:
1. Introduction
2. Materials and Methods
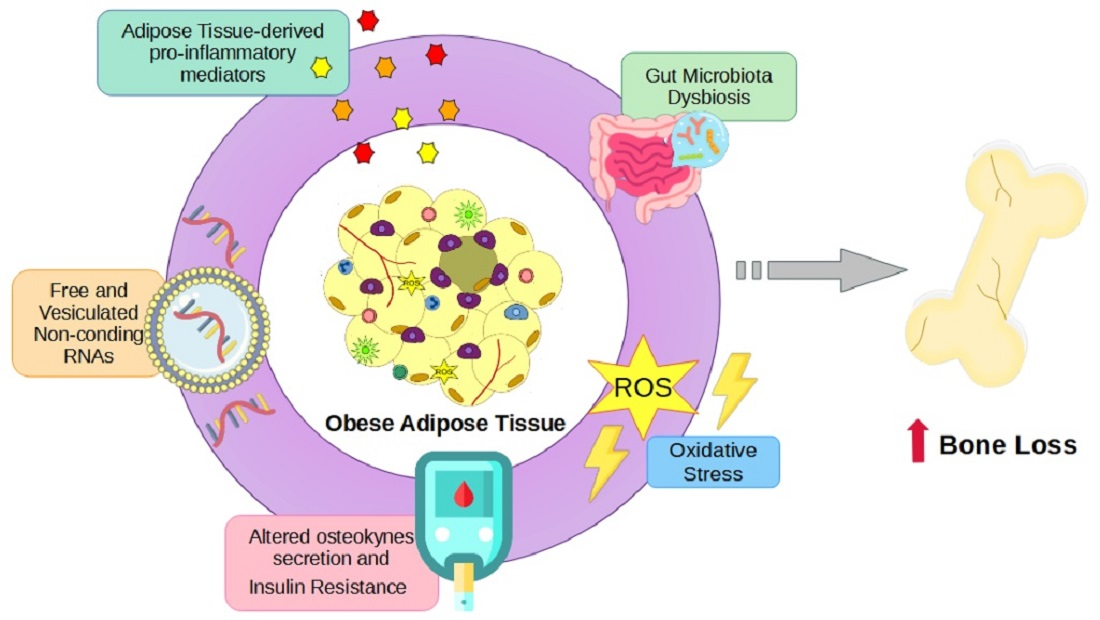
3. Results
3.1. Proinflammatory Cytokines in Bone Metabolism
3.2. Adipokines in Bone Remodeling
3.3. Muscle-Fat-Bone Crosstalk during Obesity
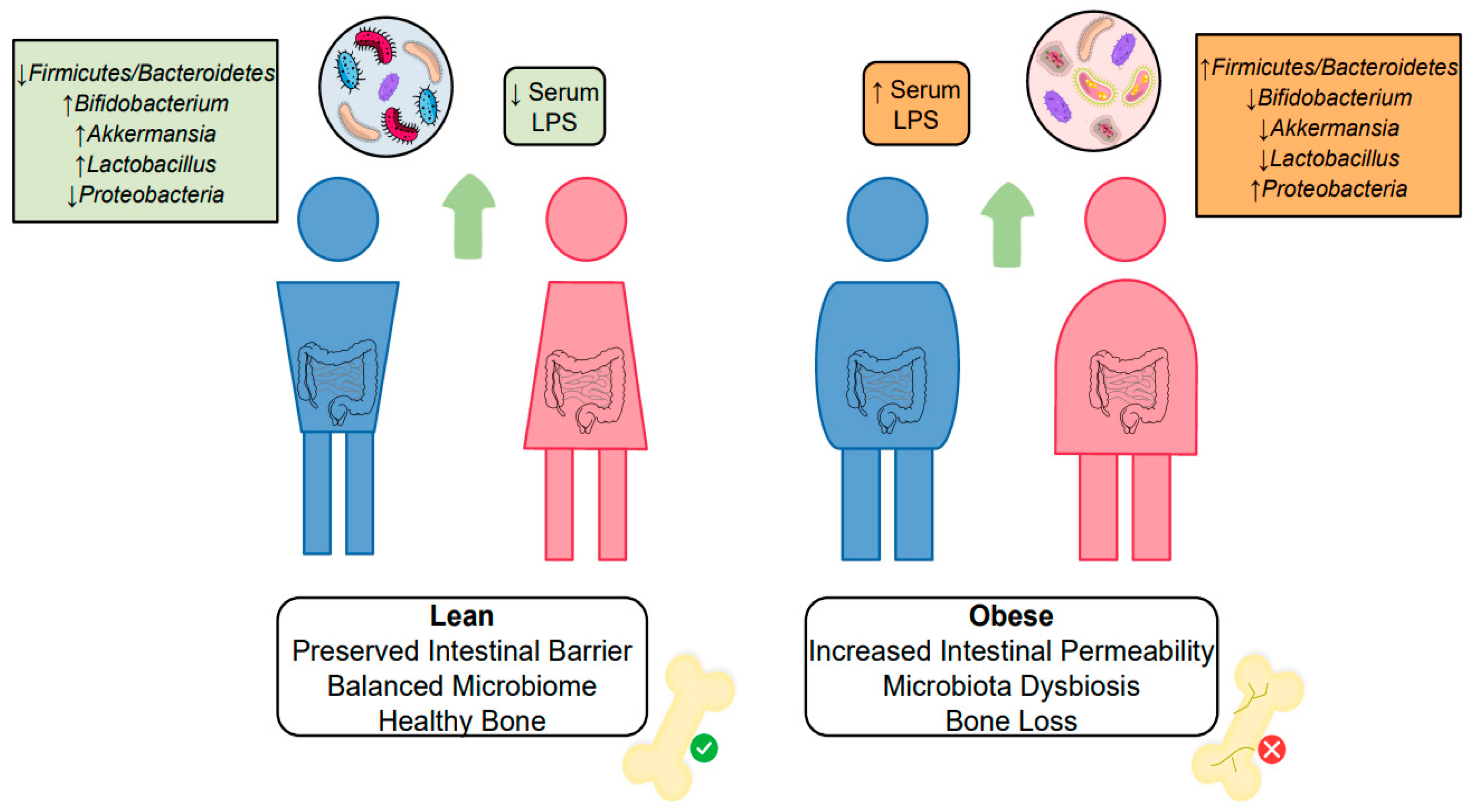
3.4. Gut and Microbiota Alterations Influencing Bone Metabolism
3.5. Osteocalcin, FGF23, and Lipocalin-2 Regulations upon Glucose Metabolism
3.6. Oxidative Stress and Bone Remodeling
3.7. Free and Vesiculated Noncoding RNAs

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Noncoding RNA | Target | Effect on Bone Cells | Obesity Status | Free or Vesiculated |
|---|---|---|---|---|
| MiR-34a-5p | WNT1 [233] | Inhibition of Osteogenesis [233] Decreased Bone Mineral Density [234] | Upregulated [231,232] | Free [231] Obese Adipocyte-derived Exosomes [232] |
| MiR-142-5p | VCAM-1 [245] PTEN [244] | Inhibits Bone Marrow MSCs Migration Leading to Osteoporosis [245] Favors Osteoclast Differentiation [244] | Upregulated [243] | Free |
| MiR-155 | SMAD5 [235] SOCS1 [236] LepR [246] IFN-β [247] | Inhibit Osteogenesis [235,236] Favors Osteoclast Differentiation [246] Inhibits Osteoclast Differentiation [247] | Upregulated [237,238] | Obese Adipose Tissue Macrophages-Derived Exosomes [237] Obese Adipocyte-Derived Microvesicles [238] |
| MiR-503 | BMPR1a [240] SMURF1 [239] | Inhibit Adipogenesis [240] Favors Osteogenesis [239] | Downregulated [241] | Free |
| Lnc-H19 | Mir-467/ HOX10 [248] | Favors Osteogenesis [248] Inhibits Adipogenesis [248] | Downregulated [248] | Mesenchymal Stem Cell-Derived Exosomes |
| MiR-149-3p | Obesity-Associated Gene FTO [242] | Favors Osteogenesis [242] Inhibit Adipogenesis [242] | Unknown Overexpression of FTO Gene is Associated with Obesity Development [249] | Unknown |
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- The World Health Organization (WHO). Heath Topics Obesity. Available online: https://www.who.int/health-topics/obesity#tab=tab_1 (accessed on 12 February 2022).
- World Obesity Atlas 2022. Available online: https://www.worldobesity.org/resources/resource-library/world-obesity-atlas-2022 (accessed on 12 February 2022).
- Rogero, M.M.; Calder, P.C. Obesity, Inflammation, Toll-Like Receptor 4 and Fatty Acids. Nutrients 2018, 10, 432. [Google Scholar] [CrossRef] [PubMed]
- Piché, M.E.; Tchernof, A.; Després, J.P. Obesity Phenotypes, Diabetes, and Cardiovascular Diseases. Circ. Res. 2020, 126, 1477–1500. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; He, C. Pro-inflammatory cytokines: The link between obesity and osteoarthritis. Cytokine Growth Factor Rev. 2018, 44, 38–50. [Google Scholar] [CrossRef]
- Avgerinos, K.I.; Spyrou, N.; Mantzoros, S.C.; Dalamaga, M. Obesity and cancer risk: Emerging biological mechanisms and perspective. Metab. Clin. Exp. 2019, 92, 121–135. [Google Scholar] [CrossRef] [PubMed]
- The World Health Organization (WHO) Obesity 2021. Available online: https://www.who.int/news-room/facts-in-pictures/detail/6-facts-on-obesity (accessed on 12 February 2022).
- Vecchié, A.; Dallegri, F.; Carbone, F.; Bonaventura, A.; Liberale, L.; Portincasa, P.; Frühbeck, G.; Montecucco, F. Obesity phenotypes and their paradoxical association with cardiovascular diseases. Eur. J. Intern. Med. 2017, 48, 6–17. [Google Scholar] [CrossRef] [PubMed]
- Unamuno, X.; Gómez-Ambrosi, J.; Rodríguez, A.; Becerril, S.; Frühbeck, G.; Catalán, V. Adipokine dysregulation and adipose tissue inflammation in human obesity. Eur. J. Clin. Investig. 2018, 48, e12997. [Google Scholar] [CrossRef]
- Sun, K.; Kusminski, C.M.; Scherer, E.P. Adipose tissue remodeling and obesity. J. Clin. Investig. 2011, 121, 2094–2101. [Google Scholar] [CrossRef]
- Deng, T.; Lyon, C.J.; Bergin, S.; Caligiuri, M.A.; 4 Hsueh, W.A. Obesity, Inflammation, and Cancer. Annu. Rev. Pathol. Mech. Dis. 2016, 11, 421–449. [Google Scholar] [CrossRef]
- Trayhurn, P. Hypoxia and Adipose Tissue Function and Dysfunction in Obesity. Physiol. Rev. 2013, 93, 1–21. [Google Scholar] [CrossRef]
- Stolarczyk, E. Adipose tissue inflammation in obesity: A metabolic or immune response? Curr. Opin. Pharmacol. 2017, 37, 35–40. [Google Scholar] [CrossRef]
- Thaler, J.P.; Yi, C.; Schur, E.A.; Guyenet, S.J.; Hwang, B.H.; Dietrich, M.O.; Zhao, X.; Sarruf, D.A.; Izgur, V.; Maravilla, K.R.; et al. Obesity is associated with hypothalamic injury in rodents and humans. J. Clin. Investig. 2012, 122, 153–162. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Ballantyne, C.M. Skeletal muscle inflammation and insulin resistance in obesity. J. Clin. Investig. 2017, 127, 43–54. [Google Scholar] [CrossRef] [PubMed]
- Stern, J.H.; Smith, G.I.; Chen, S.; Unger, R.H.; Klein, S.; Scherer, P.E. Obesity dysregulates fasting-induced changes in glucagon secretion. J. Endocrinol. 2019, 243, 149–160. [Google Scholar] [CrossRef]
- Van der Heijden, R.A.; Sheedfar, F.; Morrison, M.C.; Hommelberg, P.P.H.; Kor, D.; Kloosterhuis, N.J.; Gruben, N.; Youssef, S.A.; de Bruin, A.; Hofker, M.H.; et al. High-fat diet induced obesity primes inflammation in adipose tissue prior to liver in C57BL/6j mice. Aging 2015, 7, 256–268. [Google Scholar] [CrossRef] [PubMed]
- López-Gómez, J.J.; Castrillón, J.L.P.; de Luis Román, D.A. Influencia de la obesidad sobre el metabolismo óseo. Endocrinol. Y Nutr. 2016, 63, 551–559. [Google Scholar] [CrossRef]
- Kim, A.; Shah, A.S.; Nakamura, T. Extracellular Vesicles: A Potential Novel Regulator of Obesity and Its Associated Complications. Children 2018, 5, 152. [Google Scholar] [CrossRef]
- Abels, E.R.; Breakefield, X.O. Introduction to Extracellular Vesicles: Biogenesis, RNA Cargo Selection, Content, Release, and Uptake. Cell. Mol. Neurobiol. 2016, 36, 301–312. [Google Scholar] [CrossRef]
- Mathieu, M.; Martin-Jaular, L.; Lavieu, G.; Théry, C. Specificities of secretion and uptake of exosomes and other extracellular vesicles for cell-to-cell communication. Nat. Cell Biol. 2019, 21, 9–17. [Google Scholar] [CrossRef]
- Renovato-Martins, M.; Matheus, M.E.; de Andrade, I.R.; Moraes, J.A.; da Silva, S.V.; dos Reis, M.C.; de Souza, A.A.P.; da Silva, C.C.; Bouskela, E.; Barja-Fidalgo, C. Microparticles derived from obese adipose tissue elicit a pro-inflammatory phenotype of CD16 +, CCR5 + and TLR8 + monocytes. Biochim. Biophys. Acta (BBA) Mol. Basis Dis. 2017, 1863, 139–151. [Google Scholar] [CrossRef]
- Liu, M.; Sun, Y.; Zhang, Q. Emerging Role of Extracellular Vesicles in Bone Remodeling. J. Dent. Res. 2018, 97, 859–868. [Google Scholar] [CrossRef]
- Yáñez-Mó, M.; Siljander, P.R.-M.; Andreu, Z.; Bedina Zavec, A.; Borràs, F.E.; Buzas, E.I.; Buzas, K.; Casal, E.; Cappello, F.; Carvalho, J.; et al. Biological properties of extracellular vesicles and their physiological functions. J. Extracell. Vesicles 2015, 4, 27066. [Google Scholar] [CrossRef] [PubMed]
- Ramos-Andrade, I.; Moraes, J.; Brandão-Costa, R.M.; Da Silva, S.V.; De Souza, A.; Da Silva, C.; Renovato-Martins, M.; Barja-Fidalgo, C. Obese adipose tissue extracellular vesicles raise breast cancer cell malignancy. Endocr.-Relat. Cancer 2020, 27, 571–582. [Google Scholar] [CrossRef]
- Ji, C.; Guo, X. The clinical potential of circulating microRNAs in obesity. Nat. Rev. Endocrinol. 2019, 15, 731–743. [Google Scholar] [CrossRef]
- de Sousa, M.C.; Gjorgjieva, M.; Dolicka, D.; Sobolewski, C.; Foti, M. Deciphering miRNAs’ Action through miRNA Editing. Int. J. Mol. Sci. 2019, 20, 6249. [Google Scholar] [CrossRef] [PubMed]
- Florencio-Silva, R.; da Silva Sasso, G.R.; Sasso-Cerri, E.; Simões, M.J.; Cerri, P.S. Biology of Bone Tissue: Structure, Function, and Factors That Influence Bone Cells. BioMed. Res. Int. 2015, 2015, 421746. [Google Scholar] [CrossRef]
- Czekanska, E.; Stoddart, M.; Richards, R.; Hayes, J. In search of an osteoblast cell model for in vitro research. Eur. Cells Mater. 2012, 24, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Guntur, A.R.; Rosen, C.J. Bone As An Endocrine Organ. Endocr. Pr. 2012, 18, 758–762. [Google Scholar] [CrossRef]
- Xiao, W.; Wang, Y.; Pacios, S.; Li, S.; Graves, D.T. Cellular and Molecular Aspects of Bone Remodeling. Front. Oral. Biol. 2016, 18, 9–16. [Google Scholar] [CrossRef]
- Buck, D.W.; Dumanian, G.A. Bone Biology and Physiology. Plast. Reconstr. Surg. 2012, 129, 1314–1320. [Google Scholar] [CrossRef]
- Roeder, E.; Matthews, B.G.; Kalajzic, I. Visual reporters for study of the osteoblast lineage. Bone 2016, 92, 189–195. [Google Scholar] [CrossRef] [Green Version]
- Hou, Z.; Wang, Z.; Tao, Y.; Bai, J.; Yu, B.; Shen, J.; Sun, H.; Xiao, L.; Xu, Y.; Zhou, J.; et al. KLF2 regulates osteoblast differentiation by targeting of Runx2. Lab. Investig. 2019, 99, 271–280. [Google Scholar] [CrossRef]
- Gu, J.; Tong, X.-S.; Chen, G.-H.; Wang, D.; Chen, Y.; Yuan, Y.; Liu, X.-Z.; Bian, J.-C.; Liu, Z.-P. Effects of 1α,25-(OH)2D3 on the formation and activity of osteoclasts in RAW264.7 cells. J. Steroid Biochem. Mol. Biol. 2015, 152, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Chiu, Y.-H.; Ritchlin, C.T. DC-STAMP: A Key Regulator in Osteoclast Differentiation. J. Cell. Physiol. 2016, 231, 2402–2407. [Google Scholar] [CrossRef]
- Chen, X.; Wang, Z.; Duan, N.; Zhu, G.; Schwarz, E.M.; Xie, C. Osteoblast–osteoclast interactions. Connect. Tissue Res. 2017, 59, 99–107. [Google Scholar] [CrossRef] [PubMed]
- Hemmatian, H.; Bakker, A.D.; Klein-Nulend, J.; Van Lenthe, G.H. Aging, Osteocytes, and Mechanotransduction. Curr. Osteoporos. Rep. 2017, 15, 401–411. [Google Scholar] [CrossRef] [PubMed]
- Tresguerres, F.; Torres, J.; López-Quiles, J.; Hernández, G.; Vega, J.; Tresguerres, I. The osteocyte: A multifunctional cell within the bone. Ann. Anat.-Anat. Anz. 2019, 227, 151422. [Google Scholar] [CrossRef]
- Shapses, S.A.; Pop, L.C.; Wang, Y. Obesity is a concern for bone health with aging. Nutr. Res. 2017, 39, 1–13. [Google Scholar] [CrossRef]
- Cao, J.J. Effects of obesity on bone metabolism. J. Orthop. Surg. Res. 2011, 6, 30. [Google Scholar] [CrossRef]
- Fintini, D.; Cianfarani, S.; Cofini, M.; Andreoletti, A.; Ubertini, G.M.; Cappa, M.; Manco, M. The Bones of Children with Obesity. Front. Endocrinol. 2020, 11, 200. [Google Scholar] [CrossRef]
- Marahleh, A.; Kitaura, H.; Ohori, F.; Kishikawa, A.; Ogawa, S.; Shen, W.-R.; Qi, J.; Noguchi, T.; Nara, Y.; Mizoguchi, I. TNF-α Directly Enhances Osteocyte RANKL Expression and Promotes Osteoclast Formation. Front. Immunol. 2019, 10, 2925. [Google Scholar] [CrossRef] [Green Version]
- Shinohara, H.; Teramachi, J.; Okamura, H.; Yang, D.; Nagata, T.; Haneji, T. Double Stranded RNA-Dependent Protein Kinase is Necessary for TNF-α-Induced Osteoclast Formation In Vitro and In Vivo. J. Cell. Biochem. 2015, 116, 1957–1967. [Google Scholar] [CrossRef] [PubMed]
- Luo, G.; Li, F.; Li, X.; Wang, Z.-G.; Zhang, B. TNF-α and RANKL promote osteoclastogenesis by upregulating RANK via the NF-κB pathway. Mol. Med. Rep. 2018, 17, 6605–6611. [Google Scholar] [CrossRef] [PubMed]
- Ohori, F.; Kitaura, H.; Ogawa, S.; Shen, W.-R.; Qi, J.; Noguchi, T.; Marahleh, A.; Nara, Y.; Pramusita, A.; Mizoguchi, I. IL-33 Inhibits TNF-α-Induced Osteoclastogenesis and Bone Resorption. Int. J. Mol. Sci. 2020, 21, 1130. [Google Scholar] [CrossRef] [PubMed]
- Shu, L.; Beier, E.; Sheu, T.; Zhang, H.; Zuscik, M.J.; Puzas, E.J.; Boyce, B.F.; Mooney, R.A.; Xing, L. High-Fat Diet Causes Bone Loss in Young Mice by Promoting Osteoclastogenesis Through Alteration of the Bone Marrow Environment. Calcif. Tissue Int. 2015, 96, 313–323. [Google Scholar] [CrossRef]
- Noguchi, T.; Kitaura, H.; Ogawa, S.; Qi, J.; Shen, W.-R.; Ohori, F.; Marahleh, A.; Nara, Y.; Pramusita, A.; Mizoguchi, I. TNF-α stimulates the expression of RANK during orthodontic tooth movement. Arch. Oral Biol. 2020, 117, 104796. [Google Scholar] [CrossRef]
- Min, H.-Y.; Son, H.-E.; Jang, W.-G. Alpha-pinene promotes osteoblast differentiation and attenuates TNFα-induced inhibition of differentiation in MC3T3-E1 pre-osteoblasts. Clin. Exp. Pharmacol. Physiol. 2019, 47, 831–837. [Google Scholar] [CrossRef]
- Abuna, R.P.; De Oliveira, F.S.; Santos, T.D.S.; Guerra, T.R.; Rosa, A.L.; Beloti, M.M. Participation of TNF-α in Inhibitory Effects of Adipocytes on Osteoblast Differentiation. J. Cell. Physiol. 2015, 231, 204–214. [Google Scholar] [CrossRef]
- Jeong, B.-C. ATF3 mediates the inhibitory action of TNF-α on osteoblast differentiation through the JNK signaling pathway. Biochem. Biophys. Res. Commun. 2018, 499, 696–701. [Google Scholar] [CrossRef]
- Wang, N.; Zhou, Z.; Wu, T.; Liu, W.; Yin, P.; Pan, C.; Yu, X. TNF-α-induced NF-κB activation upregulates microRNA-150-3p and inhibits osteogenesis of mesenchymal stem cells by targeting β-catenin. Open Biol. 2016, 6, 150258. [Google Scholar] [CrossRef]
- Ye, X.; Huang, H.; Zhao, N.; Zhang, J.; Yang, P. Inhibition of Runx2 signaling by TNF-α in ST2 murine bone marrow stromal cells undergoing osteogenic differentiation. Vitr. Cell. Dev. Biol.-Anim. 2016, 52, 1026–1033. [Google Scholar] [CrossRef]
- Bin, G.; Cuifang, W.; Bo, Z.; Jing, W.; Jin, J.; Xiaoyi, T.; Cong, C.; Yonggang, C.; Liping, A.; Jinglin, M.; et al. Fluid shear stress inhibits TNF-α-induced osteoblast apoptosis via ERK5 signaling pathway. Biochem. Biophys. Res. Commun. 2015, 466, 117–123. [Google Scholar] [CrossRef]
- Tsai, C.-L.; Chen, W.-C.; Hsieh, H.-L.; Chi, P.-L.; Hsiao, L.-D.; Yang, C.-M. TNF-α induces matrix metalloproteinase-9-dependent soluble intercellular adhesion molecule-1 release via TRAF2-mediated MAPKs and NF-κB activation in osteoblast-like MC3T3-E1 cells. J. Biomed. Sci. 2014, 21, 12. [Google Scholar] [CrossRef]
- Constanze, B.; Popper, B.; Aggarwal, B.B.; Shakibaei, M. Evidence that TNF-β suppresses osteoblast differentiation of mesenchymal stem cells and resveratrol reverses it through modulation of NF-κB, Sirt1 and Runx2. Cell Tissue Res. 2020, 381, 83–98. [Google Scholar] [CrossRef]
- Muluke, M.; Gold, T.; Kiefhaber, K.; Al-Sahli, A.; Celenti, R.; Jiang, H.; Cremers, S.; Van Dyke, T.; Schulze-Späte, U. Diet-Induced Obesity and Its Differential Impact on Periodontal Bone Loss. J. Dent. Res. 2015, 95, 223–229. [Google Scholar] [CrossRef]
- Zhang, K.; Wang, C.; Chen, Y.; Ji, X.; Chen, X.; Tian, L.; Yu, X. Preservation of high-fat diet-induced femoral trabecular bone loss through genetic target of TNF-α. Endocrine 2015, 50, 239–249. [Google Scholar] [CrossRef]
- Galozzi, P.; Bindoli, S.; Doria, A.; Sfriso, P. The revisited role of interleukin-1 alpha and beta in autoimmune and inflammatory disorders and in comorbidities. Autoimmun. Rev. 2021, 20, 102785. [Google Scholar] [CrossRef]
- Lee, J.; Park, C.; Kim, H.J.; Lee, Y.D.; Lee, Z.H.; Song, Y.W.; Kim, H.-H. Stimulation of osteoclast migration and bone resorption by C–C chemokine ligands 19 and 21. Exp. Mol. Med. 2017, 49, e358. [Google Scholar] [CrossRef]
- Cao, Y.; Jansen, I.D.C.; Sprangers, S.; Stap, J.; Leenen, P.J.; Everts, V.; de Vries, T.J. IL-1β differently stimulates proliferation and multinucleation of distinct mouse bone marrow osteoclast precursor subsets. J. Leukoc. Biol. 2016, 100, 513–523. [Google Scholar] [CrossRef]
- Son, H.S.; Lee, J.; Lee, H.I.; Kim, N.; Jo, Y.-J.; Lee, G.-R.; Hong, S.-E.; Kwon, M.; Kim, N.Y.; Kim, H.J.; et al. Benzydamine inhibits osteoclast differentiation and bone resorption via down-regulation of interleukin-1 expression. Acta Pharm. Sin. B 2019, 10, 462–474. [Google Scholar] [CrossRef]
- Wang, Y.; Galli, M.; Silver, A.S.; Lee, W.; Song, Y.; Mei, Y.M.; Bachus, C.; Glogauer, M.; McCulloch, C.A. IL1β and TNFα promote RANKL-dependent adseverin expression and osteoclastogenesis. J. Cell Sci. 2018, 131, jcs213967. [Google Scholar] [CrossRef] [Green Version]
- Lee, W.-S.; Lee, E.-G.; Sung, M.-S.; Choi, Y.-J.; Yoo, W.-H. Atorvastatin inhibits osteoclast differentiation by suppressing NF-κB and MAPK signaling during IL-1β-induced osteoclastogenesis. Korean J. Intern. Med. 2018, 33, 397–406. [Google Scholar] [CrossRef]
- Moon, S.-J.; Ahn, I.E.; Jung, H.; Yi, H.; Kim, J.; Kim, Y.; Kwok, S.-K.; Park, K.-S.; Min, J.-K.; Park, S.-H.; et al. Temporal differential effects of proinflammatory cytokines on osteoclastogenesis. Int. J. Mol. Med. 2013, 31, 769–777. [Google Scholar] [CrossRef]
- Qu, X.; Mei, J.; Yu, Z.; Zhai, Z.; Qiao, H.; Dai, K. Lenalidomide regulates osteocytes fate and related osteoclastogenesis via IL-1β/NF-κB/RANKL signaling. Biochem. Biophys. Res. Commun. 2018, 501, 547–555. [Google Scholar] [CrossRef]
- Ozeki, N.; Kawai, R.; Yamaguchi, H.; Hiyama, T.; Kinoshita, K.; Hase, N.; Nakata, K.; Kondo, A.; Mogi, M.; Nakamura, H. RETRACTED: IL-1β-induced matrix metalloproteinase-13 is activated by a disintegrin and metalloprotease-28-regulated proliferation of human osteoblast-like cells. Exp. Cell Res. 2014, 323, 165–177. [Google Scholar] [CrossRef]
- Hengartner, N.-E.; Fiedler, J.; Ignatius, A.; Brenner, R.E. IL-1β Inhibits Human Osteoblast Migration. Mol. Med. 2013, 19, 36–42. [Google Scholar] [CrossRef]
- Lazzerini, P.E.; Capperucci, C.; Spreafico, A.; Capecchi, P.L.; Niccolini, S.; Ferrata, P.; Frediani, B.; Galeazzi, M.; Laghi-Pasini, F. Rosuvastatin inhibits spontaneous and IL-1β-induced interleukin-6 production from human cultured osteoblastic cells. Jt. Bone Spine 2013, 80, 195–200. [Google Scholar] [CrossRef] [PubMed]
- Hah, Y.-S.; Kang, H.-G.; Cho, H.-Y.; Shin, S.-H.; Kim, U.-K.; Park, B.-W.; Lee, S.-I.; Rho, G.-J.; Kim, J.-R.; Byun, J.-H. JNK signaling plays an important role in the effects of TNF-α and IL-1β on in vitro osteoblastic differentiation of cultured human periosteal-derived cells. Mol. Biol. Rep. 2013, 40, 4869–4881. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Liu, Q.; Ahn, J.H.; Kim, S.B.; Kim, Y.C.; Sung, S.H.; Hwang, B.Y.; Lee, M.K. Luteolin downregulates IL-1β-induced MMP-9 and -13 expressions in osteoblasts via inhibition of ERK signalling pathway. J. Enzym. Inhib. Med. Chem. 2011, 27, 261–266. [Google Scholar] [CrossRef] [PubMed]
- McKnight, Q.; Jenkins, S.; Li, X.; Nelson, T.; Marlier, A.; Cantley, L.G.; Finberg, K.E.; Fretz, J.A. IL-1β Drives Production of FGF-23 at the Onset of Chronic Kidney Disease in Mice. J. Bone Miner. Res. 2020, 35, 1352–1362. [Google Scholar] [CrossRef]
- Yamazaki, M.; Kawai, M.; Miyagawa, K.; Ohata, Y.; Tachikawa, K.; Kinoshita, S.; Nishino, J.; Ozono, K.; Michigami, T. Interleukin-1-induced acute bone resorption facilitates the secretion of fibroblast growth factor 23 into the circulation. J. Bone Miner. Metab. 2014, 33, 342–354. [Google Scholar] [CrossRef]
- Rupp, T.; Butscheidt, S.; Vettorazzi, E.; Oheim, R.; Barvencik, F.; Amling, M.; Rolvien, T. High FGF23 levels are associated with impaired trabecular bone microarchitecture in patients with osteoporosis. Osteoporos. Int. 2019, 30, 1655–1662. [Google Scholar] [CrossRef]
- He, Z.; Sun, Y.; Wu, J.; Xiong, Z.; Zhang, S.; Liu, J.; Liu, Y.; Li, H.; Jin, T.; Yang, Y.; et al. Evaluation of genetic variants in IL-1B and its interaction with the predisposition of osteoporosis in the northwestern Chinese Han population. J. Gene Med. 2020, 22, e3214. [Google Scholar] [CrossRef]
- Halade, G.V.; El Jamali, A.; Williams, P.J.; Fajardo, R.J.; Fernandes, G. Obesity-mediated inflammatory microenvironment stimulates osteoclastogenesis and bone loss in mice. Exp. Gerontol. 2011, 46, 43–52. [Google Scholar] [CrossRef]
- Hirano, T. IL-6 in inflammation, autoimmunity and cancer. Int. Immunol. 2020, 33, 127–148. [Google Scholar] [CrossRef]
- Wu, Q.; Zhou, X.; Huang, D.; Ji, Y.; Kang, F. IL-6 Enhances Osteocyte-Mediated Osteoclastogenesis by Promoting JAK2 and RANKL Activity In Vitro. Cell. Physiol. Biochem. 2017, 41, 1360–1369. [Google Scholar] [CrossRef]
- Bakker, A.; Kulkarni, R.; Klein-Nulend, J.; Lems, W. IL-6 Alters Osteocyte Signaling toward Osteoblasts but Not Osteoclasts. J. Dent. Res. 2014, 93, 394–399. [Google Scholar] [CrossRef]
- McGregor, N.E.; Murat, M.; Elango, J.; Poulton, I.J.; Walker, E.C.; Crimeen-Irwin, B.; Ho, P.W.M.; Gooi, J.H.; Martin, T.J.; Sims, N.A. IL-6 exhibits both cis- and trans-signaling in osteocytes and osteoblasts, but only trans-signaling promotes bone formation and osteoclastogenesis. J. Biol. Chem. 2019, 294, 7850–7863. [Google Scholar] [CrossRef]
- He, B.; Yin, X.; Hao, D.; Zhang, X.; Zhang, Z.; Zhang, K.; Yang, X. Blockade of IL-6 alleviates bone loss induced by modeled microgravity in mice. Can. J. Physiol. Pharmacol. 2020, 98, 678–683. [Google Scholar] [CrossRef]
- Wang, C.; Tian, L.; Zhang, K.; Chen, Y.; Chen, X.; Xie, Y.; Zhao, Q.; Yu, X. Interleukin-6 gene knockout antagonizes high-fat-induced trabecular bone loss. J. Mol. Endocrinol. 2016, 57, 161–170. [Google Scholar] [CrossRef]
- Pathak, J.L.; Bakker, A.D.; Luyten, F.P.; Verschueren, P.; Lems, W.F.; Klein-Nulend, J.; Bravenboer, N. Systemic Inflammation Affects Human Osteocyte-Specific Protein and Cytokine Expression. Calcif. Tissue Int. 2016, 98, 596–608. [Google Scholar] [CrossRef]
- Yokota, K.; Sato, K.; Miyazaki, T.; Kitaura, H.; Kayama, H.; Miyoshi, F.; Araki, Y.; Akiyama, Y.; Takeda, K.; Mimura, T. Combination of Tumor Necrosis Factor α and Interleukin-6 Induces Mouse Osteoclast-like Cells with Bone Resorption Activity Both In Vitro and In Vivo. Arthritis Rheumatol. 2013, 66, 121–129. [Google Scholar] [CrossRef] [PubMed]
- Ricci, R.; Bevilacqua, F. The potential role of leptin and adiponectin in obesity: A comparative review. Veter. J. 2012, 191, 292–298. [Google Scholar] [CrossRef]
- Zhang, Y.; Chua, S., Jr. Leptin Function and Regulation. Compr. Physiol. 2017, 8, 351–369. [Google Scholar] [CrossRef] [PubMed]
- Obradovic, M.; Sudar-Milovanovic, E.; Soskic, S.; Essack, M.; Arya, S.; Stewart, A.J.; Gojobori, T.; Isenovic, E.R. Leptin and obesity: Role and clinical implication. Front. Endocrinol. 2021, 12, 585887. [Google Scholar] [CrossRef]
- Wang, Z.V.; Scherer, P.E. Adiponectin, the past two decades. J. Mol. Cell Biol. 2016, 8, 93–100. [Google Scholar] [CrossRef] [PubMed]
- Achari, A.E.; Jain, S.K. Adiponectin, a Therapeutic Target for Obesity, Diabetes, and Endothelial Dysfunction. Int. J. Mol. Sci. 2017, 18, 1321. [Google Scholar] [CrossRef]
- Upadhyay, J.; Farr, O.M.; Mantzoros, C.S. The role of leptin in regulating bone metabolism. Metabolism 2014, 64, 105–113. [Google Scholar] [CrossRef]
- China, S.P.; Sanyal, S.; Chattopadhyay, N. Adiponectin signaling and its role in bone metabolism. Cytokine 2018, 112, 116–131. [Google Scholar] [CrossRef]
- Guimarães, M.F.B.D.R.; de Andrade, M.V.M.; Machado, C.J.; Vieira, L.M.; Pinto, M.R.D.C.; Júnior, A.L.T.; Kakehasi, A.M. Leptin as an obesity marker in rheumatoid arthritis. Rheumatol. Int. 2018, 38, 1671–1677. [Google Scholar] [CrossRef]
- Fasshauer, M.; Blüher, M. Adipokines in health and disease. Trends Pharmacol. Sci. 2015, 36, 461–470. [Google Scholar] [CrossRef]
- Xie, C.; Chen, Q. Adipokines: New Therapeutic Target for Osteoarthritis? Curr. Rheumatol. Rep. 2019, 21, 71. [Google Scholar] [CrossRef]
- Yang, J.; Park, O.-J.; Kim, J.; Han, S.; Yang, Y.; Yun, C.-H.; Han, S.H. Adiponectin Deficiency Triggers Bone Loss by Up-Regulation of Osteoclastogenesis and Down-Regulation of Osteoblastogenesis. Front. Endocrinol. 2019, 10, 815. [Google Scholar] [CrossRef]
- Wang, F.; Wang, P.-X.; Wu, X.-L.; Dang, S.-Y.; Chen, Y.; Ni, Y.-Y.; Gao, L.-H.; Lu, S.-Y.; Kuang, Y.; Huang, L.; et al. Deficiency of Adiponectin Protects against Ovariectomy-Induced Osteoporosis in Mice. PLoS ONE 2013, 8, e68497. [Google Scholar] [CrossRef] [PubMed]
- Tu, Q.; Zhang, J.; Dong, L.Q.; Saunders, E.; Luo, E.; Tang, J.; Chen, J. Adiponectin Inhibits Osteoclastogenesis and Bone Resorption via APPL1-mediated Suppression of Akt1. J. Biol. Chem. 2011, 286, 12542–12553. [Google Scholar] [CrossRef]
- Abbott, M.J.; Roth, T.M.; Ho, L.; Wang, L.; O’Carroll, D.; Nissenson, R.A. Negative Skeletal Effects of Locally Produced Adiponectin. PLoS ONE 2015, 10, e0134290. [Google Scholar] [CrossRef]
- Liu, Z.; Liu, H.; Li, Y.; Wang, Y.; Xing, R.; Mi, F.; Xiang, C.; Fu, R. Adiponectin inhibits the differentiation and maturation of osteoclasts via the mTOR pathway in multiple myeloma. Int. J. Mol. Med. 2020, 45, 1112–1120. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Huang, L.; Wu, X.; Liu, X.; Xu, Q.; Li, F.; Dai, M.; Zhang, B. Adiponectin inhibits osteoclastogenesis by suppressing NF-κB and p38 signaling pathways. Biochem. Biophys. Res. Commun. 2018, 503, 2075–2082. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Huang, L.; Liu, J. Effects of adiponectin on osteoclastogenesis from mouse bone marrow-derived monocytes. Exp. Ther. Med. 2018, 17, 1228–1233. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Liu, S.; Ji, H.; Zhao, Q.; Liu, Y.; Hu, P.; Luo, E. An adiponectin receptor agonist promote osteogenesis via regulating bone-fat balance. Cell Prolif. 2021, 54, e13035. [Google Scholar] [CrossRef] [PubMed]
- China, S.P.; Pal, S.; Chattopadhyay, S.; Porwal, K.; Kushwaha, S.; Bhattacharyya, S.; Mittal, M.; Gurjar, A.A.; Barbhuyan, T.; Singh, A.K.; et al. Globular adiponectin reverses osteo-sarcopenia and altered body composition in ovariectomized rats. Bone 2017, 105, 75–86. [Google Scholar] [CrossRef]
- Haugen, S.; He, J.; Sundaresan, A.; Stunes, A.K.; Aasarød, K.M.; Tiainen, H.; Syversen, U.; Skallerud, B.; Reseland, J.E. Adiponectin Reduces Bone Stiffness: Verified in a Three-Dimensional Artificial Human Bone Model In Vitro. Front. Endocrinol. 2018, 9, 236. [Google Scholar] [CrossRef] [PubMed]
- Pacheco-Pantoja, E.L.; Fraser, W.D.; Wilson, P.J.M.; Gallagher, J.A. Differential effects of adiponectin in osteoblast-like cells. J. Recept. Signal Transduct. 2014, 34, 351–360. [Google Scholar] [CrossRef] [PubMed]
- Yin, G.; Liu, H.; Li, J.; Liu, Y.; Liu, X.; Luo, E. Adenoviral delivery of adiponectin ameliorates osteogenesis around implants in ovariectomized rats. J. Gene Med. 2019, 21, e3069. [Google Scholar] [CrossRef] [PubMed]
- Dragojevič, J.; Zupan, J.; Haring, G.; Herman, S.; Komadina, R.; Marc, J. Triglyceride metabolism in bone tissue is associated with osteoblast and osteoclast differentiation: A gene expression study. J. Bone Miner. Metab. 2013, 31, 512–519. [Google Scholar] [CrossRef]
- Yue, R.; Zhou, B.O.; Shimada, I.S.; Zhao, Z.; Morrison, S.J. Leptin Receptor Promotes Adipogenesis and Reduces Osteogenesis by Regulating Mesenchymal Stromal Cells in Adult Bone Marrow. Cell Stem Cell 2016, 18, 782–796. [Google Scholar] [CrossRef]
- Li, W.; Xu, P.; Wang, C.; Ha, X.; Gu, Y.; Wang, Y.; Zhang, J.; Xie, J. The effects of fat-induced obesity on bone metabolism in rats. Obes. Res. Clin. Pr. 2017, 11, 454–463. [Google Scholar] [CrossRef]
- Carnovali, M.; Luzi, L.; Terruzzi, I.; Banfi, G.; Mariotti, M. Metabolic and bone effects of high-fat diet in adult zebrafish. Endocrine 2017, 61, 317–326. [Google Scholar] [CrossRef]
- Feresin, R.G.; Johnson, S.A.; Elam, M.L.; Jo, E.; Arjmandi, B.H.; Hakkak, R. Effects of Obesity on Bone Mass and Quality in Ovariectomized Female Zucker Rats. J. Obes. 2014, 2014, 690123. [Google Scholar] [CrossRef]
- Philbrick, K.A.; Wong, C.P.; Branscum, A.J.; Turner, R.T.; Iwaniec, U.T. Leptin stimulates bone formation in ob/ob mice at doses having minimal impact on energy metabolism. J. Endocrinol. 2017, 232, 461–474. [Google Scholar] [CrossRef]
- Wu, Z.; Shao, P.; Dass, C.R.; Wei, Y. Systemic leptin administration alters callus VEGF levels and enhances bone fracture healing in wildtype and ob/ob mice. Injury 2018, 49, 1739–1745. [Google Scholar] [CrossRef]
- Tencerova, M.; Frost, M.; Figeac, F.; Nielsen, T.K.; Ali, D.; Lauterlein, J.-J.L.; Andersen, T.L.; Haakonsson, A.K.; Rauch, A.; Madsen, J.S.; et al. Obesity-Associated Hypermetabolism and Accelerated Senescence of Bone Marrow Stromal Stem Cells Suggest a Potential Mechanism for Bone Fragility. Cell Rep. 2019, 27, 2050–2062.e6. [Google Scholar] [CrossRef]
- Li, J.; Gao, Y.; Yu, T.; Lange, J.K.; LeBoff, M.S.; Gorska, A.; Luu, S.; Zhou, S.; Glowacki, J. Obesity and leptin influence vitamin D metabolism and action in human marrow stromal cells. J. Steroid Biochem. Mol. Biol. 2019, 198, 105564. [Google Scholar] [CrossRef]
- Maggio, A.B.; Belli, D.C.; Puigdefabregas, J.W.B.; Rizzoli, R.; Farpour-Lambert, N.J.; Beghetti, M.; McLin, V.A. High Bone Density in Adolescents with Obesity Is Related to Fat Mass and Serum Leptin Concentrations. J. Craniofacial Surg. 2014, 58, 723–728. [Google Scholar] [CrossRef]
- Nakamura, Y.; Nakano, M.; Suzuki, T.; Sato, J.; Kato, H.; Takahashi, J.; Shiraki, M. Two adipocytokines, leptin and adiponectin, independently predict osteoporotic fracture risk at different bone sites in postmenopausal women. Bone 2020, 137, 115404. [Google Scholar] [CrossRef] [PubMed]
- Dimitri, P.; Jacques, R.; Paggiosi, M.; King, D.; Walsh, J.; Taylor, Z.; Frangi, A.; Bishop, N.; Eastell, R. Leptin May Play a Role in Bone Microstructural Alterations in Obese Children. J. Clin. Endocrinol. Metab. 2015, 100, 594–602. [Google Scholar] [CrossRef]
- Reid, I.R.; A Baldock, P.; Cornish, J. Effects of Leptin on the Skeleton. Endocr. Rev. 2018, 39, 938–959. [Google Scholar] [CrossRef] [PubMed]
- Wong, I.P.; Nguyen, A.D.; Khor, E.C.; Enriquez, R.F.; A Eisman, J.; Sainsbury, A.; Herzog, H.; A Baldock, P. Neuropeptide Y is a critical modulator of Leptin’s regulation of cortical bone. J. Bone Miner. Res. 2012, 28, 886–898. [Google Scholar] [CrossRef]
- Wee, N.K.Y.; Enriquez, R.F.; Nguyen, A.; Horsnell, H.; Kulkarni, R.; Khor, E.C.; Herzog, H.; Baldock, P.A. Diet-induced obesity suppresses cortical bone accrual by a neuropeptide Y-dependent mechanism. Int. J. Obes. 2018, 42, 1925–1938. [Google Scholar] [CrossRef]
- Srivastava, G.; Apovian, C. Future Pharmacotherapy for Obesity: New Anti-obesity Drugs on the Horizon. Curr. Obes. Rep. 2018, 7, 147–161. [Google Scholar] [CrossRef]
- Erener, T.; Ceritoğlu, K.U.; Aktekin, C.N.; Dalgic, A.D.; Keskin, D.; Geneci, F.; Ocak, M.; Bilecenoğlu, B.; Hücümenoğlu, S.; Çaydere, M.; et al. Investigation of the effect of ghrelin on bone fracture healing in rats. Clin. Exp. Pharmacol. Physiol. 2021, 48, 1382–1390. [Google Scholar] [CrossRef] [PubMed]
- Xiao, L.; Zhang, H.; Wang, Y.; Li, J.; Yang, G.; Wang, L.; Liang, Z. Dysregulation of the ghrelin/RANKL/OPG pathway in bone mass is related to AIS osteopenia. Bone 2020, 134, 115291. [Google Scholar] [CrossRef] [PubMed]
- Müller, T.; Finan, B.; Bloom, S.; D’Alessio, D.; Drucker, D.; Flatt, P.; Fritsche, A.; Gribble, F.; Grill, H.; Habener, J.; et al. Glucagon-like peptide 1 (GLP-1). Mol. Metab. 2019, 30, 72–130. [Google Scholar] [CrossRef] [PubMed]
- Meng, J.; Ma, X.; Wang, N.; Jia, M.; Bi, L.; Wang, Y.; Li, M.; Zhang, H.; Xue, X.; Hou, Z.; et al. Activation of GLP-1 Receptor Promotes Bone Marrow Stromal Cell Osteogenic Differentiation through β-Catenin. Stem Cell Rep. 2016, 6, 579–591. [Google Scholar] [CrossRef] [PubMed]
- Mansur, S.A.; Mieczkowska, A.; Flatt, P.R.; Chappard, D.; Irwin, N.; Mabilleau, G. The GLP-1 Receptor Agonist Exenatide Ameliorates Bone Composition and Tissue Material Properties in High Fat Fed Diabetic Mice. Front. Endocrinol. 2019, 10, 51. [Google Scholar] [CrossRef]
- Kirk, B.; Feehan, J.; Lombardi, G.; Duque, G. Muscle, Bone, and Fat Crosstalk: The Biological Role of Myokines, Osteokines, and Adipokines. Curr. Osteoporos. Rep. 2020, 18, 388–400. [Google Scholar] [CrossRef]
- Ilich, J.Z.; Gilman, J.C.; Cvijetic, S.; Boschiero, D. Chronic Stress Contributes to Osteosarcopenic Adiposity via Inflammation and Immune Modulation: The Case for More Precise Nutritional Investigation. Nutrients 2020, 12, 989. [Google Scholar] [CrossRef]
- Al Saedi, A.; Hassan, E.B.; Duque, G. The diagnostic role of fat in osteosarcopenia. J. Lab. Precis. Med. 2019, 4, 7. [Google Scholar] [CrossRef]
- Li, H.; Wang, F.; Yang, M.; Sun, J.; Zhao, Y.; Tang, D. The Effect of Irisin as a Metabolic Regulator and Its Therapeutic Potential for Obesity. Int. J. Endocrinol. 2021, 2021, 6572342. [Google Scholar] [CrossRef]
- Xue, Y.; Hu, S.; Chen, C.; He, J.; Sun, J.; Jin, Y.; Zhang, Y.; Zhu, G.; Shi, Q.; Rui, Y. Myokine Irisin promotes osteogenesis by activating BMP/SMAD signaling via αV integrin and regulates bone mass in mice. Int. J. Biol. Sci. 2022, 18, 572–584. [Google Scholar] [CrossRef]
- He, X.; Hua, Y.; Li, Q.; Zhu, W.; Pan, Y.; Yang, Y.; Li, X.; Wu, M.; Wang, J.; Gan, X. FNDC5/irisin facilitates muscle−adipose−bone connectivity through ubiquitination-dependent activation of runt-related transcriptional factors RUNX1/2. J. Biol. Chem. 2022, 298, 101679. [Google Scholar] [CrossRef]
- Ozgor, B.Y.; Demiral, I.; Zeybek, U.; Celik, F.; Buyru, F.; Yeh, J.; Bastu, E. Effects of Irisin Compared with Exercise on Specific Metabolic and Obesity Parameters in Female Mice with Obesity. Metab. Syndr. Relat. Disord. 2020, 18, 141–145. [Google Scholar] [CrossRef]
- Kang, Y.S.; Kim, J.C.; Kim, J.S.; Kim, S.H. Effects of Swimming Exercise on Serum Irisin and Bone FNDC5 in Rat Models of High-Fat Diet-Induced Osteoporosis. J. Sport. Sci. Med. 2019, 18, 596–603. [Google Scholar]
- Bäckhed, F.; Ding, H.; Wang, T.; Hooper, L.V.; Koh, G.Y.; Nagy, A.; Semenkovich, C.F.; Gordon, J.I. The Gut Microbiota as an Environmental Factor That Regulates Fat Storage. Proc. Natl. Acad. Sci. USA 2004, 101, 15718–15723. [Google Scholar] [CrossRef] [PubMed]
- Turnbaugh, P.J.; Ley, R.E.; Mahowald, M.A.; Magrini, V.; Mardis, E.R.; Gordon, J.I. An Obesity-Associated Gut Microbiome with Increased Capacity for Energy Harvest. Nature 2006, 444, 1027–1031. [Google Scholar] [CrossRef]
- Chelakkot, C.; Ghim, J.; Ryu, S.H. Mechanisms regulating intestinal barrier integrity and its pathological implications. Exp. Mol. Med. 2018, 50, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Wang, P.; Cao, M.; Li, M.; Wang, F. Protective Role of Oligomycin Against Intestinal Epithelial Barrier Dysfunction Caused by IFN-? and TNF-a. Cell. Physiol. Biochem. 2012, 29, 799–808. [Google Scholar] [CrossRef] [PubMed]
- Saad, M.J.A.; Santos, A.; Prada, P.O. Linking Gut Microbiota and Inflammation to Obesity and Insulin Resistance. Physiology 2016, 31, 283–293. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Chen, G.-L.; Hannemann, N.; Ipseiz, N.; Krönke, G.; Bäuerle, T.; Munos, L.; Wirtz, S.; Schett, G.; Bozec, A. Microbiota from Obese Mice Regulate Hematopoietic Stem Cell Differentiation by Altering the Bone Niche. Cell Metab. 2015, 22, 886–894. [Google Scholar] [CrossRef]
- Sato, K.; Yamazaki, K.; Kato, T.; Nakanishi, Y.; Tsuzuno, T.; Yokoji-Takeuchi, M.; Yamada-Hara, M.; Miura, N.; Okuda, S.; Ohno, H.; et al. Obesity-Related Gut Microbiota Aggravates Alveolar Bone Destruction in Experimental Periodontitis through Elevation of Uric Acid. Mbio 2021, 12, e0077121. [Google Scholar] [CrossRef]
- Gomes, A.C.; Hoffmann, C.; Mota, J.F. The human gut microbiota: Metabolism and perspective in obesity. Gut Microbes 2018, 9, 308–325. [Google Scholar] [CrossRef]
- Stojanov, S.; Berlec, A.; Štrukelj, B. The Influence of Probiotics on the Firmicutes/Bacteroidetes Ratio in the Treatment of Obesity and Inflammatory Bowel disease. Microorganisms 2020, 8, 1715. [Google Scholar] [CrossRef] [PubMed]
- McCabe, L.R.; Irwin, R.; Tekalur, A.; Evans, C.; Schepper, J.D.; Parameswaran, N.; Ciancio, M. Exercise prevents high fat diet-induced bone loss, marrow adiposity and dysbiosis in male mice. Bone 2018, 118, 20–31. [Google Scholar] [CrossRef] [PubMed]
- Xie, W.; Han, Y.; Li, F.; Gu, X.; Su, D.; Yu, W.; Li, Z.; Xiao, J. Neuropeptide Y1 Receptor Antagonist Alters Gut Microbiota and Alleviates the Ovariectomy-Induced Osteoporosis in Rats. Calcif. Tissue Int. 2019, 106, 444–454. [Google Scholar] [CrossRef] [PubMed]
- Méndez-Salazar, E.O.; Ortiz-López, M.G.; Granados-Silvestre, M.D.L.; Palacios-González, B.; Menjivar, M. Altered Gut Microbiota and Compositional Changes in Firmicutes and Proteobacteria in Mexican Undernourished and Obese Children. Front. Microbiol. 2018, 9, 2494. [Google Scholar] [CrossRef] [PubMed]
- Palacios-González, B.; Ramírez-Salazar, E.G.; Rivera-Paredez, B.; Quiterio, M.; Flores, Y.N.; Macias-Kauffer, L.; Moran-Ramos, S.; Denova-Gutiérrez, E.; Ibarra-González, I.; Vela-Amieva, M.; et al. A Multi-Omic Analysis for Low Bone Mineral Density in Postmenopausal Women Suggests a Relationship between Diet, Metabolites, and Microbiota. Microorganisms 2020, 8, 1630. [Google Scholar] [CrossRef] [PubMed]
- Amabebe, E.; Robert, F.O.; Agbalalah, T.; Orubu, E.S.F. Microbial dysbiosis-induced obesity: Role of gut microbiota in homoeostasis of energy metabolism. Br. J. Nutr. 2020, 123, 1127–1137. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Chen, C.; Liu, Z.; Luo, Z.; Rao, S.; Jin, L.; Wan, T.; Yue, T.; Tan, Y.; Yin, H.; et al. Extracellular Vesicles from Child Gut Microbiota Enter into Bone to Preserve Bone Mass and Strength. Adv. Sci. 2021, 8, 2004831. [Google Scholar] [CrossRef]
- Huck, O.; Mulhall, H.; Rubin, G.; Kizelnik, Z.; Iyer, R.; Perpich, J.D.; Haque, N.; Cani, P.D.; de Vos, W.M.; Amar, S. Akkermansia muciniphila reduces Porphyromonas gingivalis -induced inflammation and periodontal bone destruction. J. Clin. Periodontol. 2019, 47, 202–212. [Google Scholar] [CrossRef] [PubMed]
- Raftar, S.K.A.; Tavassol, Z.H.; Amiri, M.; Ejtahed, H.-S.; Zangeneh, M.; Sadeghi, S.; Ashrafian, F.; Kariman, A.; Khatami, S.; Siadat, S.D. Assessment of fecal Akkermansia muciniphila in patients with osteoporosis and osteopenia: A pilot study. J. Diabetes Metab. Disord. 2021, 20, 279–284. [Google Scholar] [CrossRef]
- Fernández-Murga, M.L.; Olivares, M.; Sanz, Y. Bifidobacterium pseudocatenulatum CECT 7765 reverses the adverse effects of diet-induced obesity through the gut-bone axis. Bone 2020, 141, 115580. [Google Scholar] [CrossRef]
- Behera, J.; Ison, J.; Voor, M.J.; Tyagi, N. Probiotics Stimulate Bone Formation in Obese Mice via Histone Methylations. Theranostics 2021, 11, 8605–8623. [Google Scholar] [CrossRef] [PubMed]
- Eaimworawuthikul, S.; Tunapong, W.; Chunchai, T.; Yasom, S.; Wanchai, K.; Suntornsaratoon, P.; Charoenphandhu, N.; Thiennimitr, P.; Chattipakorn, N.; Chattipakorn, S.C. Effects of probiotics, prebiotics or synbiotics on jawbone in obese-insulin resistant rats. Eur. J. Nutr. 2018, 58, 2801–2810. [Google Scholar] [CrossRef] [PubMed]
- Eaimworawuthikul, S.; Tunapong, W.; Chunchai, T.; Suntornsaratoon, P.; Charoenphandhu, N.; Thiennimitr, P.; Chattipakorn, N.; Chattipakorn, S.C. Altered gut microbiota ameliorates bone pathology in the mandible of obese–insulin-resistant rats. Eur. J. Nutr. 2019, 59, 1453–1462. [Google Scholar] [CrossRef] [PubMed]
- Tyagi, A.M.; Yu, M.; Darby, T.M.; Vaccaro, C.; Li, J.-Y.; Owens, J.A.; Hsu, E.; Adams, J.; Weitzmann, M.N.; Jones, R.M.; et al. The Microbial Metabolite Butyrate Stimulates Bone Formation via T Regulatory Cell-Mediated Regulation of WNT10B Expression. Immunity 2018, 49, 1116–1131.e7. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Zhao, H.; Liu, Z.; Sun, X.; Zhang, D.; Wang, S.; Xu, Y.; Zhang, G.; Wang, D. Modulation of Gut Microbiota by Fucoxanthin During Alleviation of Obesity in High-Fat Diet-Fed Mice. J. Agric. Food Chem. 2020, 68, 5118–5128. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Lin, T.; Meng, Y.; Hu, M.; Shu, L.; Jiang, H.; Gao, R.; Ma, J.; Wang, C.; Zhou, X. FOS/GOS attenuates high-fat diet induced bone loss via reversing microbiota dysbiosis, high intestinal permeability and systemic inflammation in mice. Metabolism 2021, 119, 154767. [Google Scholar] [CrossRef] [PubMed]
- Rios, J.L.; Bomhof, M.R.; Reimer, R.A.; Hart, D.A.; Collins, K.H.; Herzog, W. Protective effect of prebiotic and exercise intervention on knee health in a rat model of diet-induced obesity. Sci. Rep. 2019, 9, 3893. [Google Scholar] [CrossRef]
- Kim, K.-J.; Lee, J.; Wang, W.; Lee, Y.; Oh, E.; Park, K.-H.; Park, C.; Woo, G.-E.; Son, Y.-J.; Kang, H. Austalide K from the Fungus Penicillium rudallense Prevents LPS-Induced Bone Loss in Mice by Inhibiting Osteoclast Differentiation and Promoting Osteoblast Differentiation. Int. J. Mol. Sci. 2021, 22, 5493. [Google Scholar] [CrossRef]
- Goel, P.N.; Egol, A.J.; Moharrer, Y.; Brandfield-Harvey, B.; Ahn, J.; Ashley, J.W. Notch signaling inhibition protects against LPS mediated osteolysis. Biochem. Biophys. Res. Commun. 2019, 515, 538–543. [Google Scholar] [CrossRef]
- Kishikawa, A.; Kitaura, H.; Kimura, K.; Ogawa, S.; Qi, J.; Shen, W.-R.; Ohori, F.; Noguchi, T.; Marahleh, A.; Nara, Y.; et al. Docosahexaenoic Acid Inhibits Inflammation-Induced Osteoclast Formation and Bone Resorption in vivo Through GPR120 by Inhibiting TNF-α Production in Macrophages and Directly Inhibiting Osteoclast Formation. Front. Endocrinol. 2019, 10, 157. [Google Scholar] [CrossRef]
- Jin, J.; Machado, E.R.; Yu, H.; Zhang, X.; Lu, Z.; Li, Y.; Lopes-Virella, M.F.; Kirkwood, K.; Huang, Y. Simvastatin inhibits LPS-induced alveolar bone loss during metabolic syndrome. J. Dent. Res. 2013, 93, 294–299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, C.; Yuan, L.; Wang, J.-G.; Wang, F.; Yang, X.-K.; Zhang, F.-H.; Song, J.-L.; Ma, X.-Y.; Cheng, Q.; Song, G.-H. Lipopolysaccharide (LPS) Induces the Apoptosis and Inhibits Osteoblast Differentiation Through JNK Pathway in MC3T3-E1 Cells. Inflammation 2013, 37, 621–631. [Google Scholar] [CrossRef] [PubMed]
- Gao, A.; Wang, X.; Yu, H.; Li, N.; Hou, Y.; Yu, W. Effect of Porphyromonas gingivalis lipopolysaccharide (Pg-LPS) on the expression of EphA2 in osteoblasts and osteoclasts. Vitr. Cell. Dev. Biol.-Anim. 2015, 52, 228–234. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.; Hu, C.-C.; Wu, Y.-Y.; Ueng, S.; Chang, C.-H.; Chen, M.-F. Ibudilast Mitigates Delayed Bone Healing Caused by Lipopolysaccharide by Altering Osteoblast and Osteoclast Activity. Int. J. Mol. Sci. 2021, 22, 1169. [Google Scholar] [CrossRef] [PubMed]
- Irie, N.; Takada, Y.; Watanabe, Y.; Matsuzaki, Y.; Naruse, C.; Asano, M.; Iwakura, Y.; Suda, T.; Matsuo, K. Bidirectional Signaling through EphrinA2-EphA2 Enhances Osteoclastogenesis and Suppresses Osteoblastogenesis. J. Biol. Chem. 2009, 284, 14637–14644. [Google Scholar] [CrossRef]
- Xing, Q.; de Vos, P.; Faas, M.; Ye, Q.; Ren, Y. LPS Promotes Pre-osteoclast Activity by Up-regulating CXCR4 via TLR-4. J. Dent. Res. 2011, 90, 157–162. [Google Scholar] [CrossRef]
- Han, Y.; You, X.; Xing, W.; Zhang, Z.; Zou, W. Paracrine and endocrine actions of bone—the functions of secretory proteins from osteoblasts, osteocytes, and osteoclasts. Bone Res. 2018, 6, 16. [Google Scholar] [CrossRef]
- Ho, B.B.; Bergwitz, C. FGF23 signalling and physiology. J. Mol. Endocrinol. 2021, 66, R23–R32. [Google Scholar] [CrossRef]
- Prié, D. FGF23 and Cardiovascular Risk. Ann. d’Endocrinologie 2020, 82, 141–143. [Google Scholar] [CrossRef]
- Mizokami, A.; Kawakubo-Yasukochi, T.; Hirata, M. Osteocalcin and its endocrine functions. Biochem. Pharmacol. 2017, 132, 1–8. [Google Scholar] [CrossRef]
- Mosialou, I.; Shikhel, S.; Liu, J.-M.; Maurizi, A.; Luo, N.; He, Z.; Huang, Y.; Zong, H.; Friedman, R.A.; Barasch, J.; et al. MC4R-dependent suppression of appetite by bone-derived lipocalin 2. Nature 2017, 543, 385–390. [Google Scholar] [CrossRef] [PubMed]
- Yaylali, Y.T.; Fidan-Yaylali, G.; Dedeoglu, O.; Senol, H. Osteocalcin and epicardial adipose tissue in obesity: New hints for epicardial adipose tissue–bone crosstalk. Scand. Cardiovasc. J. 2019, 53, 296–298. [Google Scholar] [CrossRef] [PubMed]
- Geserick, M.; Vogel, M.; Eckelt, F.; Schlingmann, M.; Hiemisch, A.; Baber, R.; Thiery, J.; Körner, A.; Kiess, W.; Kratzsch, J. Children and adolescents with obesity have reduced serum bone turnover markers and 25-hydroxyvitamin D but increased parathyroid hormone concentrations—Results derived from new pediatric reference ranges. Bone 2019, 132, 115124. [Google Scholar] [CrossRef] [PubMed]
- Kim, G.S.; Jekal, Y.; Kim, H.S.; Im, J.-A.; Park, J.Y.; Chu, S.H. Reduced serum total osteocalcin is associated with central obesity in Korean children. Obes. Res. Clin. Pr. 2014, 8, e230–e237. [Google Scholar] [CrossRef] [PubMed]
- Razny, U.; Goralska, J.; Zdzienicka, A.; Fedak, D.; Masania, J.; Rabbani, N.; Thornalley, P.; Pawlica-Gosiewska, D.; Gawlik, K.; Dembinska-Kiec, A.; et al. Relation of the protein glycation, oxidation and nitration to the osteocalcin level in obese subjects. Acta Biochim. Pol. 2017, 64, 415–422. [Google Scholar] [CrossRef] [PubMed]
- Vigevano, F.; Gregori, G.; Colleluori, G.; Chen, R.; Autemrongsawat, V.; Napoli, N.; Qualls, C.; Villareal, D.T.; Armamento-Villareal, R. In Men With Obesity, T2DM Is Associated with Poor Trabecular Microarchitecture and Bone Strength and Low Bone Turnover. J. Clin. Endocrinol. Metab. 2021, 106, 1362–1376. [Google Scholar] [CrossRef]
- Guedes, J.; Esteves, J.; Morais, M.; Zorn, T.; Furuya, D. Osteocalcin improves insulin resistance and inflammation in obese mice: Participation of white adipose tissue and bone. Bone 2018, 115, 68–82. [Google Scholar] [CrossRef]
- Zhou, B.; Li, H.; Liu, J.; Xu, L.; Guo, Q.; Zang, W.; Sun, H.; Wu, S. Autophagic dysfunction is improved by intermittent administration of osteocalcin in obese mice. Int. J. Obes. 2016, 40, 833–843. [Google Scholar] [CrossRef]
- Ferron, M.; McKee, M.D.; Levine, R.L.; Ducy, P.; Karsenty, G. Intermittent injections of osteocalcin improve glucose metabolism and prevent type 2 diabetes in mice. Bone 2012, 50, 568–575. [Google Scholar] [CrossRef]
- Hanks, L.J.; Casazza, K.; Judd, S.E.; Jenny, N.S.; Gutiérrez, O.M. Associations of Fibroblast Growth Factor-23 with Markers of Inflammation, Insulin Resistance and Obesity in Adults. PLoS ONE 2015, 10, e0122885. [Google Scholar] [CrossRef]
- Zaheer, S.; De Boer, I.H.; Allison, M.; Brown, J.M.; Psaty, B.M.; Robinson-Cohen, C.; Michos, E.D.; Ix, J.H.; Kestenbaum, B.; Siscovick, D.; et al. Fibroblast Growth Factor 23, Mineral Metabolism, and Adiposity in Normal Kidney Function. J. Clin. Endocrinol. Metab. 2017, 102, 1387–1395. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Ma, X.; Luo, Y.; Xu, Y.; Xiong, Q.; Pan, X.; Xiao, Y.; Bao, Y.; Jia, W. Associations of serum fibroblast growth factor 23 levels with obesity and visceral fat accumulation. Clin. Nutr. 2016, 37, 223–228. [Google Scholar] [CrossRef] [PubMed]
- Grethen, E.; Hill, K.M.; Jones, R.; Cacucci, B.M.; Gupta, C.E.; Acton, A.; Considine, R.V.; Peacock, M. Serum Leptin, Parathyroid Hormone, 1,25-Dihydroxyvitamin D, Fibroblast Growth Factor 23, Bone Alkaline Phosphatase, and Sclerostin Relationships in Obesity. J. Clin. Endocrinol. Metab. 2012, 97, 1655–1662. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Zhang, L.; Zhang, H.; Yang, Z.; Qi, L.; Wang, Y.; Ren, S. The participation of fibroblast growth factor 23 (FGF23) in the progression of osteoporosis via JAK/STAT pathway. J. Cell. Biochem. 2017, 119, 3819–3828. [Google Scholar] [CrossRef]
- Garland, J.S.; Holden, R.M.; Ross, R.; Adams, M.A.; Nolan, R.; Hopman, W.M.; Morton, A.R. Insulin resistance is associated with Fibroblast Growth Factor-23 in stage 3–5 chronic kidney disease patients. J. Diabetes Its Complicat. 2013, 28, 61–65. [Google Scholar] [CrossRef]
- Saito, M.; Marumo, K. Bone Quality in Diabetes. Front. Endocrinol. 2013, 4, 72. [Google Scholar] [CrossRef]
- Liu, C.; Jiang, D. High glucose-induced LIF suppresses osteoblast differentiation via regulating STAT3/SOCS3 signaling. Cytokine 2017, 91, 132–139. [Google Scholar] [CrossRef]
- Yang, L.; Liu, J.; Shan, Q.; Geng, G.; Shao, P. High glucose inhibits proliferation and differentiation of osteoblast in alveolar bone by inducing pyroptosis. Biochem. Biophys. Res. Commun. 2019, 522, 471–478. [Google Scholar] [CrossRef]
- Bornstein, S.; Moschetta, M.; Kawano, Y.; Sacco, A.; Huynh, D.; Brooks, D.; Manier, S.; Fairfield, H.; Falank, C.; Roccaro, A.M.; et al. Metformin Affects Cortical Bone Mass and Marrow Adiposity in Diet-Induced Obesity in Male Mice. Endocrinology 2017, 158, 3369–3385. [Google Scholar] [CrossRef]
- Qu, B.; Gong, K.; Yang, H.; Li, Y.; Jiang, T.; Zeng, Z.; Cao, Z.; Pan, X. SIRT1 suppresses high glucose and palmitate-induced osteoclast differentiation via deacetylating p66Shc. Mol. Cell. Endocrinol. 2018, 474, 97–104. [Google Scholar] [CrossRef]
- Xu, F.; Ye, Y.-P.; Dong, Y.-H.; Guo, F.-J.; Chen, A.-M.; Huang, S.-L. Inhibitory effects of high glucose/insulin environment on osteoclast formation and resorption in vitro. J. Huazhong Univ. Sci. Technol. 2013, 33, 244–249. [Google Scholar] [CrossRef] [PubMed]
- Mosialou, I.; Shikhel, S.; Luo, N.; Petropoulou, P.I.; Panitsas, K.; Bisikirska, B.; Rothman, N.J.; Tenta, R.; Cariou, B.; Wargny, M.; et al. Lipocalin-2 counteracts metabolic dysregulation in obesity and diabetes. J. Exp. Med. 2020, 217, e20191261. [Google Scholar] [CrossRef] [PubMed]
- Wen, X.; Zhu, B.; Zhang, Y.; Mei, F.; Cheng, X.; Qian, C.; Yang, P.; Lu, L.; Zhou, D.; Gao, J.; et al. Alterations in Fat Mass and Bone Mineral Density Are Associated with Decreased Lipocalin-2 After Laparoscopic Sleeve Gastrectomy in Obese Chinese Women. Obes. Surg. 2019, 29, 2862–2868. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Ma, X.; Pan, X.; He, X.; Xiao, Y.; Bao, Y. Correlations between serum concentration of three bone-derived factors and obesity and visceral fat accumulation in a cohort of middle aged men and women. Cardiovasc. Diabetol. 2018, 17, 143. [Google Scholar] [CrossRef] [PubMed]
- Lim, W.H.; Wong, G.; Lim, E.M.; Byrnes, E.; Zhu, K.; Devine, A.; Pavlos, N.J.; Prince, R.L.; Lewis, J.R. Circulating Lipocalin 2 Levels Predict Fracture-Related Hospitalizations in Elderly Women: A Prospective Cohort Study. J. Bone Miner. Res. 2015, 30, 2078–2085. [Google Scholar] [CrossRef]
- Kim, H.-J.; Ohk, B.; Kang, W.Y.; Seong, S.J.; Suk, K.; Lim, M.-S.; Kim, S.-Y.; Yoon, Y.-R. Deficiency of Lipocalin-2 Promotes Proliferation and Differentiation of Osteoclast Precursors via Regulation of c-Fms Expression and Nuclear Factor-kappa B Activation. J. Bone Metab. 2016, 23, 8–15. [Google Scholar] [CrossRef]
- Villalvilla, A.; García-Martín, A.; Largo, R.; Gualillo, O.; Herrero-Beaumont, G.; Gómez, R. The adipokine lipocalin-2 in the context of the osteoarthritic osteochondral junction. Sci. Rep. 2016, 6, 29243. [Google Scholar] [CrossRef]
- Costa, D.; Lazzarini, E.; Canciani, B.; Giuliani, A.; Spanò, R.; Marozzi, K.; Manescu, A.; Cancedda, R.; Tavella, S. Altered bone development and turnover in transgenic mice over-expressing Lipocalin-2 in bone. J. Cell. Physiol. 2013, 228, 2210–2221. [Google Scholar] [CrossRef]
- Rucci, N.; Capulli, M.; Piperni, S.G.; Cappariello, A.; Lau, P.; Frings-Meuthen, P.; Heer, M.; Teti, A. Lipocalin 2: A New Mechanoresponding Gene Regulating Bone Homeostasis. J. Bone Miner. Res. 2015, 30, 357–368. [Google Scholar] [CrossRef]
- Pisoschi, A.M.; Pop, A. The role of antioxidants in the chemistry of oxidative stress: A review. Eur. J. Med. Chem. 2015, 97, 55–74. [Google Scholar] [CrossRef]
- Murdolo, G.; Piroddi, M.; Luchetti, F.; Tortoioli, C.; Canonico, B.; Zerbinati, C.; Galli, F.; Iuliano, L. Oxidative stress and lipid peroxidation by-products at the crossroad between adipose organ dysregulation and obesity-linked insulin resistance. Biochimie 2013, 95, 585–594. [Google Scholar] [CrossRef] [PubMed]
- Manna, P.; Jain, S.K. Obesity, Oxidative Stress, Adipose Tissue Dysfunction, and the Associated Health Risks: Causes and Therapeutic Strategies. Metab. Syndr. Relat. Disord. 2015, 13, 423–444. [Google Scholar] [CrossRef] [PubMed]
- Engin, A. The pathogenesis of obesity-associated adipose tissue inflammation. In Obesity and Lipotoxicity; Springer: Berlin/Heidelberg, Germany, 2017; pp. 221–245. [Google Scholar]
- Lee, Y.S.; Kim, A.Y.; Choi, J.W.; Kim, M.; Yasue, S.; Son, H.J.; Masuzaki, H.; Park, K.S.; Kim, J.B. Dysregulation of Adipose Glutathione Peroxidase 3 in Obesity Contributes to Local and Systemic Oxidative Stress. Mol. Endocrinol. 2008, 22, 2176–2189. [Google Scholar] [CrossRef] [PubMed]
- Chattopadhyay, M.; Khemka, V.K.; Chatterjee, G.; Ganguly, A.; Mukhopadhyay, S.; Chakrabarti, S. Enhanced ROS production and oxidative damage in subcutaneous white adipose tissue mitochondria in obese and type 2 diabetes subjects. Mol. Cell. Biochem. 2014, 399, 95–103. [Google Scholar] [CrossRef]
- Yang, Y.; Sun, Y.; Mao, W.-W.; Zhang, H.; Ni, B.; Jiang, L. Oxidative stress induces downregulation of TP53INP2 and suppresses osteogenic differentiation of BMSCs during osteoporosis through the autophagy degradation pathway. Free. Radic. Biol. Med. 2021, 166, 226–237. [Google Scholar] [CrossRef]
- Kanda, Y.; Hinata, T.; Kang, S.W.; Watanabe, Y. Reactive oxygen species mediate adipocyte differentiation in mesenchymal stem cells. Life Sci. 2011, 89, 250–258. [Google Scholar] [CrossRef]
- Dong, X.; Bi, L.; He, S.; Meng, G.; Wei, B.; Jia, S.; Liu, J. FFAs-ROS-ERK/P38 pathway plays a key role in adipocyte lipotoxicity on osteoblasts in co-culture. Biochimie 2014, 101, 123–131. [Google Scholar] [CrossRef]
- Mukherjee, R.; Moreno-Fernandez, M.E.; Giles, D.A.; Cappelletti, M.; Stankiewicz, T.E.; Chan, C.C.; Divanovic, S. Nicotinamide adenine dinucleotide phosphate (reduced) oxidase 2 modulates inflammatory vigor during nonalcoholic fatty liver disease progression in mice. Hepatol. Commun. 2018, 2, 546–560. [Google Scholar] [CrossRef]
- Rahman, M.; El Jamali, A.; Halade, G.V.; Ouhtit, A.; Abou-Saleh, H.; Pintus, G. Nox2 Activity Is Required in Obesity-Mediated Alteration of Bone Remodeling. Oxidative Med. Cell. Longev. 2018, 2018, 1–10. [Google Scholar] [CrossRef]
- Kang, I.S.; Kim, C. NADPH oxidase gp91phox contributes to RANKL-induced osteoclast differentiation by upregulating NFATc1. Sci. Rep. 2016, 6, 38014. [Google Scholar] [CrossRef]
- Shen, C.-L.; Cao, J.J.; Dagda, R.Y.; Chanjaplammootil, S.; Lu, C.; Chyu, M.-C.; Gao, W.; Wang, J.-S.; Yeh, J.K. Green tea polyphenols benefits body composition and improves bone quality in long-term high-fat diet–induced obese rats. Nutr. Res. 2012, 32, 448–457. [Google Scholar] [CrossRef] [PubMed]
- Cao, J.J.; Picklo, M.J. N-Acetylcysteine Supplementation Decreases Osteoclast Differentiation and Increases Bone Mass in Mice Fed a High-Fat Diet. J. Nutr. 2013, 144, 289–296. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Li, B.; Wu, F.; Liu, X.; Liu, M.; Wang, C.; Shi, Y.; Ye, L. GPX7 Facilitates BMSCs Osteoblastogenesis via ER Stress and mTOR Pathway. J. Cell. Mol. Med. 2021, 25, 10454–10465. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.-C.; Yu, Y.-H.; Shew, J.-Y.; Lee, W.-J.; Hwang, J.-J.; Chen, Y.-H.; Chen, Y.-R.; Wei, P.-C.; Chuang, L.-M.; Lee, W.-H. Deficiency of NPGP x, an oxidative stress sensor, leads to obesity in mice and human. EMBO Mol. Med. 2013, 5, 1165–1179. [Google Scholar] [CrossRef]
- Matoušková, P.; Hanousková, B.; Skálová, L. MicroRNAs as Potential Regulators of Glutathione Peroxidases Expression and Their Role in Obesity and Related Pathologies. Int. J. Mol. Sci. 2018, 19, 1199. [Google Scholar] [CrossRef]
- He, H.; Zhang, Y.; Sun, Y.; Zhang, Y.; Xu, J.; Yang, Y.; Chen, J. Folic Acid Attenuates High-Fat Diet-Induced Osteoporosis Through the AMPK Signaling Pathway. Front. Cell Dev. Biol. 2022, 9. [Google Scholar] [CrossRef]
- Narimiya, T.; Kanzaki, H.; Yamaguchi, Y.; Wada, S.; Katsumata, Y.; Tanaka, K.; Tomonari, H. Nrf2 activation in osteoblasts suppresses osteoclastogenesis via inhibiting IL-6 expression. Bone Rep. 2019, 11, 100228. [Google Scholar] [CrossRef]
- Tang, X.; Ma, S.; Li, Y.; Sun, Y.; Zhang, K.; Zhou, Q.; Yu, R. Evaluating the Activity of Sodium Butyrate to Prevent Osteoporosis in Rats by Promoting Osteal GSK-3β/Nrf2 Signaling and Mitochondrial Function. J. Agric. Food Chem. 2020, 68, 6588–6603. [Google Scholar] [CrossRef]
- Chan, J.J.; Tay, Y. Noncoding RNA:RNA Regulatory Networks in Cancer. Int. J. Mol. Sci. 2018, 19, 1310. [Google Scholar] [CrossRef]
- Dykes, I.M.; Emanueli, C. Transcriptional and Post-transcriptional Gene Regulation by Long Non-coding RNA. Genom. Proteom. Bioinform. 2017, 15, 177–186. [Google Scholar] [CrossRef]
- Wang, L.; Cho, K.B.; Li, Y.; Tao, G.; Xie, Z.; Guo, B. Long Noncoding RNA (lncRNA)-Mediated Competing Endogenous RNA Networks Provide Novel Potential Biomarkers and Therapeutic Targets for Colorectal Cancer. Int. J. Mol. Sci. 2019, 20, 5758. [Google Scholar] [CrossRef] [Green Version]
- Wijesinghe, S.N.; Nicholson, T.; Tsintzas, K.; Jones, S.W. Involvements of long noncoding RNAs in obesity-associated inflammatory diseases. Obes. Rev. 2020, 22. [Google Scholar] [CrossRef] [PubMed]
- Mugoni, V.; Ciani, Y.; Nardella, C.; Demichelis, F. Circulating RNAs in prostate cancer patients. Cancer Lett. 2021, 524, 57–69. [Google Scholar] [CrossRef] [PubMed]
- Xu, F.; Jin, L.; Jin, Y.; Nie, Z.; Zheng, H. Long noncoding RNAs in autoimmune diseases. J. Biomed. Mater. Res. Part A 2018, 107, 468–475. [Google Scholar] [CrossRef] [PubMed]
- Gurha, P. Noncoding RNAs in cardiovascular diseases. Curr. Opin. Cardiol. 2019, 34, 241–245. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Xia, K.; Cen, X.; Huang, X.; Sun, W.; Zhao, Z.; Liu, J. DNA methylation of noncoding RNAs: New insights into osteogenesis and common bone diseases. Stem Cell Res. Ther. 2020, 11, 109. [Google Scholar] [CrossRef]
- Endisha, H.; Datta, P.; Sharma, A.; Nakamura, S.; Rossomacha, E.; Younan, C.; Ali, S.A.; Tavallaee, G.; Lively, S.; Potla, P.; et al. MicroRNA-34a-5p Promotes Joint Destruction During Osteoarthritis. Arthritis Rheumatol. 2020, 73, 426–439. [Google Scholar] [CrossRef]
- Pan, Y.; Hui, X.; Hoo, R.L.C.; Ye, D.; Chan, C.Y.C.; Feng, T.; Wang, Y.; Lam, K.S.L.; Xu, A. Adipocyte-secreted exosomal microRNA-34a inhibits M2 macrophage polarization to promote obesity-induced adipose inflammation. J. Clin. Investig. 2019, 129, 834–849. [Google Scholar] [CrossRef]
- Yuan, T.; Shi, C.; Xu, W.; Yang, H.-L.; Xia, B.; Tian, C. Extracellular vesicles derived from T-cell acute lymphoblastic leukemia inhibit osteogenic differentiation of bone marrow mesenchymal stem cells via miR-34a-5p. Endocr. J. 2021, 68, 1197–1208. [Google Scholar] [CrossRef]
- Nóbrega, O.T.; Morais-Junior, G.S.; Viana, N.I.; Reis, S.T.; Perez, D.I.V.; Freitas, W.M.; Sposito, A.C.; Leite, K.R.M.; Srougi, M. Circulating miR-34a and Bone Mineral Density of Brazilian Very-Old Adults. J. Aging Res. 2020, 2020, 3431828. [Google Scholar] [CrossRef]
- Gu, Y.; Ma, L.; Song, L.; Li, X.; Chen, D.; Bai, X. miR-155 Inhibits Mouse Osteoblast Differentiation by Suppressing SMAD5 Expression. BioMed Res. Int. 2017, 2017, 1893520. [Google Scholar] [CrossRef] [Green Version]
- Wu, T.; Xie, M.; Wang, X.; Jiang, X.; Li, J.; Huang, H. miR-155 modulates TNF-α-inhibited osteogenic differentiation by targeting SOCS1 expression. Bone 2012, 51, 498–505. [Google Scholar] [CrossRef] [PubMed]
- Ying, W.; Riopel, M.; Bandyopadhyay, G.; Dong, Y.; Birmingham, A.; Seo, J.B.; Ofrecio, J.M.; Wollam, J.; Hernandez-Carretero, A.; Fu, W.; et al. Adipose Tissue Macrophage-Derived Exosomal miRNAs Can Modulate In Vivo and In Vitro Insulin Sensitivity. Cell 2017, 171, 372–384.e12. [Google Scholar] [CrossRef]
- Zhang, Y.; Mei, H.; Chang, X.; Chen, F.; Zhu, Y.; Han, X. Adipocyte-derived microvesicles from obese mice induce M1 macrophage phenotype through secreted miR-155. J. Mol. Cell Biol. 2016, 8, 505–517. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Xu, J.; Xu, L.; Zhang, J.; Chan, K.; Pan, X.; Li, G. MiR-503 Promotes Bone Formation in Distraction Osteogenesis through Suppressing Smurf1 Expression. Sci. Rep. 2017, 7, 409. [Google Scholar] [CrossRef] [PubMed]
- Man, X.-F.; Tan, S.-W.; Tang, H.-N.; Guo, Y.; Tang, C.-Y.; Tang, J.; Zhou, C.-L.; Zhou, H.-D. MiR-503 inhibits adipogenesis by targeting bone morphogenetic protein receptor 1a. Am. J. Transl. Res. 2016, 8, 2727–2737. [Google Scholar]
- Yue, H.-Q.; Zhou, Y.-H.; Guo, Y.; Tang, C.-Y.; Wang, F.; Zhou, H.-D. Serum miR-503 is a Candidate Biomarker for Differentiating Metabolic Healthy Obesity from Metabolic Unhealthy Obesity. Diabetes Metab. Syndr. Obesity Targets Ther. 2020, 13, 2667–2676. [Google Scholar] [CrossRef]
- Li, Y.; Yang, F.; Gao, M.; Gong, R.; Jin, M.; Liu, T.; Sun, Y.; Fu, Y.; Huang, Q.; Zhang, W.; et al. miR-149-3p Regulates the Switch between Adipogenic and Osteogenic Differentiation of BMSCs by Targeting FTO. Mol. Ther.-Nucleic Acids 2019, 17, 590–600. [Google Scholar] [CrossRef]
- Chartoumpekis, D.V.; Zaravinos, A.; Ziros, P.G.; Iskrenova, R.P.; Psyrogiannis, A.I.; Kyriazopoulou, V.E.; Habeos, I.G. Differential Expression of MicroRNAs in Adipose Tissue after Long-Term High-Fat Diet-Induced Obesity in Mice. PLoS ONE 2012, 7, e34872. [Google Scholar] [CrossRef]
- Lou, Z.; Peng, Z.; Wang, B.; Li, X.; Li, X.; Zhang, X. miR-142-5p promotes the osteoclast differentiation of bone marrow-derived macrophages via PTEN/PI3K/AKT/FoxO1 pathway. J. Bone Miner. Metab. 2019, 37, 815–824. [Google Scholar] [CrossRef]
- Teng, Z.; Xie, X.; Zhu, Y.; Liu, J.; Hu, X.; Na, Q.; Zhang, X.; Wei, G.; Xu, S.; Liu, Y.; et al. miR-142-5p in Bone Marrow-Derived Mesenchymal Stem Cells Promotes Osteoporosis Involving Targeting Adhesion Molecule VCAM-1 and Inhibiting Cell Migration. BioMed Res. Int. 2018, 2018, 3274641. [Google Scholar] [CrossRef] [PubMed]
- Mao, Z.; Zhu, Y.; Hao, W.; Chu, C.; Su, H. MicroRNA-155 inhibition up-regulates LEPR to inhibit osteoclast activation and bone resorption via activation of AMPK in alendronate-treated osteoporotic mice. IUBMB Life 2019, 71, 1916–1928. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zhao, H.; Chen, J.; Xia, B.; Jin, Y.; Wei, W.; Shen, J.; Huang, Y. Interferon-β-induced miR-155 inhibits osteoclast differentiation by targeting SOCS1 and MITF. FEBS Lett. 2012, 586, 3255–3262. [Google Scholar] [CrossRef]
- Wang, Y.; Chen, W.; Zhao, L.; Li, Y.; Liu, Z.; Gao, H.; Bai, X.; Wang, B. Obesity regulates miR-467/HoxA10 axis on osteogenic differentiation and fracture healing by BMSC-derived exosome LncRNA H19. J. Cell. Mol. Med. 2021, 25, 1712–1724. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Yang, Y.; Sun, B.F.; Zhao, Y.L.; Yang, Y.G. FTO and obesity: Mechanisms of association. Curr. Diabetes Rep. 2014, 14, 486. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Forte, Y.S.; Renovato-Martins, M.; Barja-Fidalgo, C. Cellular and Molecular Mechanisms Associating Obesity to Bone Loss. Cells 2023, 12, 521. https://doi.org/10.3390/cells12040521
Forte YS, Renovato-Martins M, Barja-Fidalgo C. Cellular and Molecular Mechanisms Associating Obesity to Bone Loss. Cells. 2023; 12(4):521. https://doi.org/10.3390/cells12040521
Chicago/Turabian StyleForte, Yasmin Silva, Mariana Renovato-Martins, and Christina Barja-Fidalgo. 2023. "Cellular and Molecular Mechanisms Associating Obesity to Bone Loss" Cells 12, no. 4: 521. https://doi.org/10.3390/cells12040521