1. Introduction
Acute respiratory distress syndrome (ARDS), an acute lung inflammation responsible of severe hypoxemia, is a common and lethal condition in critically ill patients [
1]. Unbalanced lung inflammation following direct pulmonary aggression, such as bacterial pneumonia or COVID-19 infection, is a major contributor to ARDS. The major histological feature of ARDS, known as diffuse alveolar damage (DAD), may result from endothelial and epithelial injury, neutrophil-dependent lung invasion, and acute systemic pro-inflammatory states [
2]. The migration of leukocytes, and especially neutrophils, from the blood compartment to the alveolar space plays a key role in the development of DAD [
3]. Pre-emptive therapeutics to decrease leukocyte diapedesis could alleviate DAD and improve the prognosis of ARDS [
1].
Endocan (Esm-1) is a 50 kilodaltons (kDa) proteoglycan mainly expressed by lung endothelial cells and upregulated by pro-inflammatory cytokines [
4]. Previous laboratory data have shown that endocan could interfere with ICAM-1-dependent adhesion and the migration of human and mouse activated leukocytes, thus inhibiting their trans-endothelial migration. In addition, the protective effect of endocan on LPS-induced acute lung injury has been reported in Balb/c mice in a previous study [
5]. However, the conclusions to be drawn from these data were limited, since major interference with the endogenous murine endocan could not be ruled out. Indeed, mouse endocan seems to be characterized by the co-existence of two functionally opposite forms: one glycosylated, with likely anti-inflammatory effects, and another non-glycosylated, which is thought to act as an antagonist of glycosylated endocan [
6,
7]. Subsequently, further models allowing for the isolation of the effects of glycosylated endocan was deemed necessary.
Therefore, the aim of our study was to assess the hypothetical anti-inflammatory effect of the glycosylated form of endocan, which is the only type found in humans.
Several clinical studies conducted in trauma and septic patients support the hypothesis of an anti-inflammatory role of endocan, as higher blood concentrations of endocan at admission in intensive care units seem to reduce the risk of further progression to ARDS [
8,
9,
10]. Therefore, it has been proposed that a lack of secretion of endocan could be associated with a higher risk of respiratory failure and ARDS [
7].
The aim of this study was to assess the anti-inflammatory effect of endocan, while ruling out any possible interference of endogenous endocan in our explorations. Furthermore, the therapeutic effect of endocan on hypoxemia, reflecting the severity of respiratory failure, has never been explored. To these ends, we compared the features of lung inflammation as well as the severity of hypoxemia in an LPS-induced ALI model conducted in endocan knockout (Esm1−/−) and WT littermate mice. However, in contrast with human endocan, which is exclusively found in a fully glycosylated form, murine endocan is physiologically found in two forms: a 50 kDa glycosylated type, similar to human endocan, and a 20 kDa non-glycosylated type. Therefore, the results obtained when comparing Esm1−/− and WT mice may reflect either the effect of glycosylated or non-glycosylated endocan. We thus aimed to specifically explore, in a complementary model, the effects of a continuous infusion of glycosylated endocan in an LPS-induced ALI model in Esm1−/− mice.
2. Materials and Methods
2.1. Production and Purification of Murine Endocan
We produced and purified murine glycosylated endocan in our laboratory using stably transfected CHO DG44 cells, as previously described [
11]. Cells were cultured within CL1000 bioreactors (ref. Z688029, Integra Biosciences, Cergy, France) and in serum-free CHO media (Gibco, Waltham, MA, USA). The supernatant of the cells was enriched with recombinant murine glycosylated endocan. Then, the recombinant protein was purified on an ion exchange column (HiTrap Q FF (ref. 17-5156-01, GE Healthcare, Chicago, IL, USA)), followed by an affinity column coated with the anti-murine endocan MEP14 monoclonal antibody, as previously described [
11]. We concentrated the protein by centrifugation using Amicon
® Ultra Centrifugal Filters and finally, titrated the protein concentration using the endocan ELISA kit (ref. LIK-1101, DIYEK M1, Biothelis, France). Each production batch of the recombinant protein was controlled using a Western blot assay to confirm that the purified protein only consisted of glycosylated endocan. The preparations were shown to be LPS-free using the LAL test (ref. 50-650U, Lonza, Switzerland).
2.2. Mice and Ethical Aspects
We used C57Bl/6 Esm1−/− and WT littermate 7–11-week-old male mice, bred in our animal facility. The study protocol was approved by the local ethics committee (N° APAFIS#6059). All experimentations complied with standard ethical practices.
2.3. Generation of Esm1 Knockout Mice
Esm1−/− mice were generated as described elsewhere [
12]. Briefly, loxP recognition loci were inserted into flank exons 1 and 2 of the mouse
Esm1 gene (Ensembl ID ENSMUSG00000042379). In collaboration with the Institut Clinique de la Souris (Strasbourg, France), a targeting cDNA fragment harboring a neomycin resistance cassette flanked by loxP loci was transfected into mouse embryonic stem cells by electroporation, resulting in the replacement of
Esm1 exons 1 and 2 with the neomycin resistance cassette by homologous recombination. After transfection, embryonic stem cells for
Esm1 recombination were selected using G418-containing medium, and these clones were validated by Southern blotting and injected into blastocysts and then into pseudopregnant female mice. The progeny was then crossed to generate mice lacking exons 1 and 2 of
Esm1 on homologous chromosomes. These mice were then crossed with Cre deleterious mice to eliminate the neomycin-resistant cassette. For genotyping, we extracted genomic DNA from the mouse ears and performed PCR with the REDExtract-N-Amp™ Tissue PCR kit (ref. R4775, Sigma-Aldrich, St. Louis, MO, USA), following the manufacturer’s instructions, and analyzed the samples on agarose gel. We used primer pairs 1f (ATGTCTATGTCAGTCTCCTC) and 1r (CTCTTGCCAGCCTCTCTTGT) to generate a 310 bp fragment for the wild-type allele and primer pairs 1f and 2r (CTCCAAAATCCAGAAACCC) to generate a 442 bp fragment for the knockout allele.
2.4. Experimental Protocols
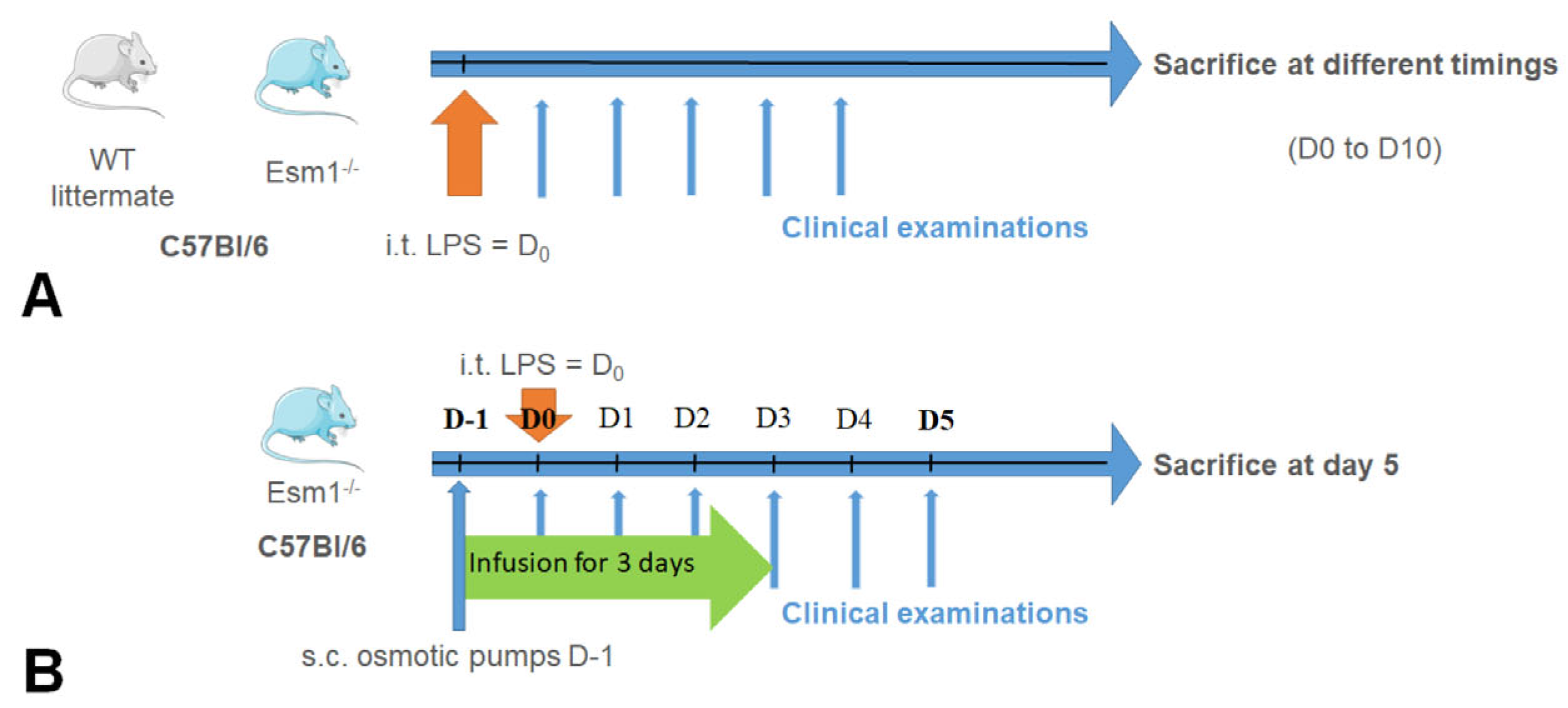
The two experimental protocols carried out in our study are described in
Figure 1. In the first protocol (
Figure 1A), we used C57Bl/6
Esm1−/− and WT littermate mice, as previously described. After intratracheal instillation of LPS, the clinical examination and evaluation of physiological pulmonary function were performed each day, as described. The mice were sacrificed at different time points in order to obtain the complete kinetics of each parameter explored from Day 0 to Day 10. In the second protocol, described in
Figure 1B, we exclusively used C57Bl/6
Esm1−/− mice. We performed subcutaneous implantation of osmotic pumps (ref. 1003D, Alzet, Cupertino, CA, USA) previously filled with PBS or murine endocan, as described, followed by the intratracheal instillation of LPS. A non-lethal blood sample was collected by retro orbital puncture just before the intratracheal instillation. We performed the same follow-up as in the first protocol, except that the mice were sacrificed on Day 5 by the intraperitoneal injection of pentobarbital (100 mg/kg).
2.5. Preparation of Osmotic Pumps and Subcutaneous Implantation
We used osmotic pumps to obtain the continuous delivery of endocan for 3 days at 1 µL/h. Each pump of 100 µL was filled with either 200 µg of murine glycosylated endocan (100 µL of endocan solution at a final concentration of 2 mg/mL) or with 100 µL of PBS. The filled pumps were placed in PBS for a maximal duration of 4 h before subcutaneous implantation. We then anesthetized the mice following the aforementioned protocol, and performed subcutaneous the implantation of the osmotic pumps after antiseptic preparation of the skin. General anesthesia was then initiated using intraperitoneal injection of 5 mg/kg of atipamezole.
2.6. Preparation and Administration of LPS for the ALI Model
LPS used in our protocol (E. Coli O111: B4, Sigma-Aldrich, St. Louis, MO, USA) was prepared in sterile conditions. We diluted 10 mg/kg of LPS in PBS for a total volume of 50 µL for each mouse. For the ALI protocol, all mice underwent general anesthesia using an intraperitoneal injection of 75 mg/kg of ketamine and 0.5 mg/kg of medetomidine, followed by the intratracheal instillation of LPS. General anesthesia was then initiated, as previously described.
2.7. Clinical Follow-Up and Noninvasive Oximetry
In both protocols, daily clinical examination was performed to assess mice viability and weight. Noninvasive recording of oximetry, heart rate, and respiratory rate were performed daily and averaged over 5 to 10 min on live mice, using the MouseOx® Plus device (Starr Life Sciences Corp, Oakmont, PA, USA).
2.8. Biological Sampling after Sacrifice and Conservation
Immediately after sacrifice, we collected blood samples, BAL, and the lungs from the mice. Blood samples were collected by obtaining a section of large thoracic vessels, which were then stored in aliquot at room temperature until coagulation. Then, samples were centrifugated at 1500× g for 5 min at room temperature. The serum and centrifugation pellets were separated, and the serum was stored at −20 °C. BAL were performed on the right lung by tracheal cannulation and after ligation of the left bronchus, with two successive washes of 500 μL of sterile PBS, and ice-stored in aliquots. BAL were centrifugated at 1500× g for 10 min at 4 °C. The supernatant and the centrifugation pellets were separated, and the supernatant was stored at −20 °C. The centrifugation pellets were then mixed with 1 mL of PBS, and the cells were manually counted using Thoma counting chambers. One hundred thousand cells were spotted using a Shandon Cytospin 4 device (Thermo electron corporation, Waltham, MA, USA), and then colored by May-Grünwald Giemsa. The left lung was placed in Antigenfix® (ref. 2545826, MM FRANCE, France) for 4 h, then rinsed in two successive PBS baths. The lungs were gradually dehydrated in successively increasing alcohol concentration baths, put in Diasolv® (ref. 2545839, MM FRANCE, France) for 30 min, and embedded in paraffin overnight.
2.9. Cell Counting and Histological Examination
For each BAL Cytospin cell identification, at least 200 cells were manually counted by an operator blinded to the mice groups. Each leukocyte visualized was categorized as a neutrophil, macrophage/monocyte, or lymphocyte. The respective proportions and absolute counts were subsequently calculated for each cell type. The paraffin-embedded lungs were prepared for histological examination with microtome to obtain sections of 5 µm thickness. Samples were then stained with hematoxylin and eosin and examined by a trained pathologist blinded to the mouse treatment to determine the main histological features of the mouse acute lung injury using the lung injury scoring system (LISS) At least 20 random fields were independently scored regarding several parameters, including neutrophils in the alveolar space, neutrophils in the interstitial space, hyaline membranes, proteinaceous debris filling the air spaces, and alveolar septal thickening. A lung injury score was then calculated by the addition of each of the five independent parameters previously mentioned, weighted according to their relevance, as described by Matute-Bello, et al. [
13].
2.10. Evaluation of Total Proteins, MPO, TNFα, and Endocan Concentrations in Sera and in BAL Samples
The sera and BAL samples were thawed and homogenized by soft shaking. The BAL protein concentrations were assessed with a Pierce™ BCA Protein Assay Kit (ref. 23227, Thermo Fisher Scientific, Waltham, MA, USA). The mouse myeloperoxidase (mMPO) concentrations in BAL were assessed with mMPO DuoSet ELISA kit (ref. DY3667, R&D Systems, Minneapolis, MN, USA). The mouse TNF-alpha (mTNF-α) concentrations in BAL were assessed with the mTNF DuoSet ELISA kit (ref. DY410, R&D Systems, Minneapolis, MN, USA). The murine endocan concentrations in the sera were assessed using DIYEK M1 ELISA kit (ref. LIK-1101, Biothelis, Lille, France).
2.11. Statistics
All statistical analyses were performed with SPSS® 26 software (IBM, Armonk, NY, USA) for Windows, and all graphs were created using GraphPad Prism 6.0 software. The normality of quantitative variables was not tested because of the small sample sizes. We used Mann–Whitney tests to compare the distributions of groups of unpaired variables. For the evaluation of the effect of the group in our first protocol, from D0 to D10 for inflammatory parameters and oximetry, as well as oximetry in our second protocol, we used a linear mixed model test with the Satterthwaite approximation using the variable “Group” as the fixed effect and the variable “Time” as the random effect. The distributions of values from a repeated variable were compared using the Kruskal–Wallis tests. All tests were performed with a 2-tailed alpha-risk set at 0.05.
4. Discussion
ARDS is a severe disease resulting from a dysregulated neutrophilic-dependent lung invasion [
2]. Endocan could play an anti-inflammatory role through its direct interaction with the leukocyte integrin LFA-1, subsequently leading to the alleviation of dysregulated leukocyte diapedesis [
14]. Data from the literature suggest that the anti-inflammatory action of human endocan relies on both the protein core and the presence of its glycanic chain. As a consequence, the suppression of the glycosylated portion of the endocan seems to be associated with the loss of this anti-inflammatory effect [
14]. In contrast with human endocan, which is exclusively found in a fully glycosylated form, murine endocan is physiologically found in two forms: the 50 kDa glycosylated type, similar to human endocan, and the 20 kDa non-glycosylated type. Such a non-glycosylated endocan has been found to be pro-inflammatory in vivo due to its antagonizing effect on the anti-inflammatory influence of glycosylated endocan [
15]. Therefore, the endogenous secretion of non-glycosylated endocan may result in significant bias in the interpretation of ALI models in mice [
6,
15], making it difficult to assess the exact effect of glycosylated endocan. Thus, the aim of this study was to specifically evaluate the physiological effect of glycosylated endocan in an LPS-induced model of ALI, using
Esm1−/− mice to rule out any potential interference of endogenous endocan.
In order to compare our results with previously published data, we assessed the inflammatory and histological features of lung inflammation according to the guidelines released by the American Thoracic Society for measurements of experimental acute lung injury in animals [
13]. To specifically address the effect of endocan, all C57BL/6 mice used in this study were similar in age and weight, as well as being littermates. The only difference between the groups was the presence or the absence of endocan. Finally, to study the effects of a continuous infusion of endocan, we used osmotic pumps in order to obtain a constant delivery of glycosylated murine endocan over 3 days.
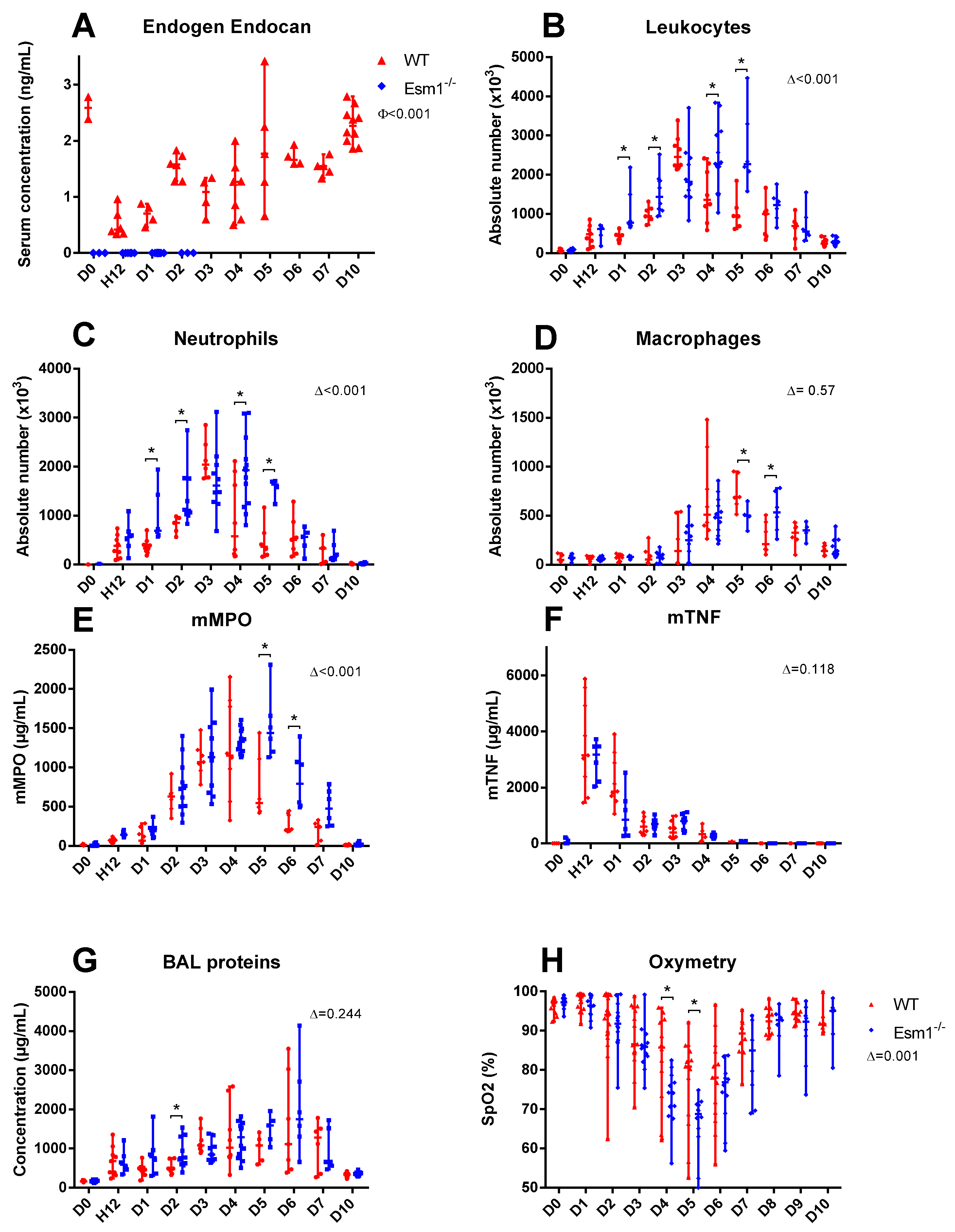
When comparing
Esm1−/− mice and WT littermates, we found that endogenous murine endocan may potentially decrease the severity of LPS-induced ALI and alleviate lung physiological dysfunction, as reflected by greater SpO2 values in the WT group (
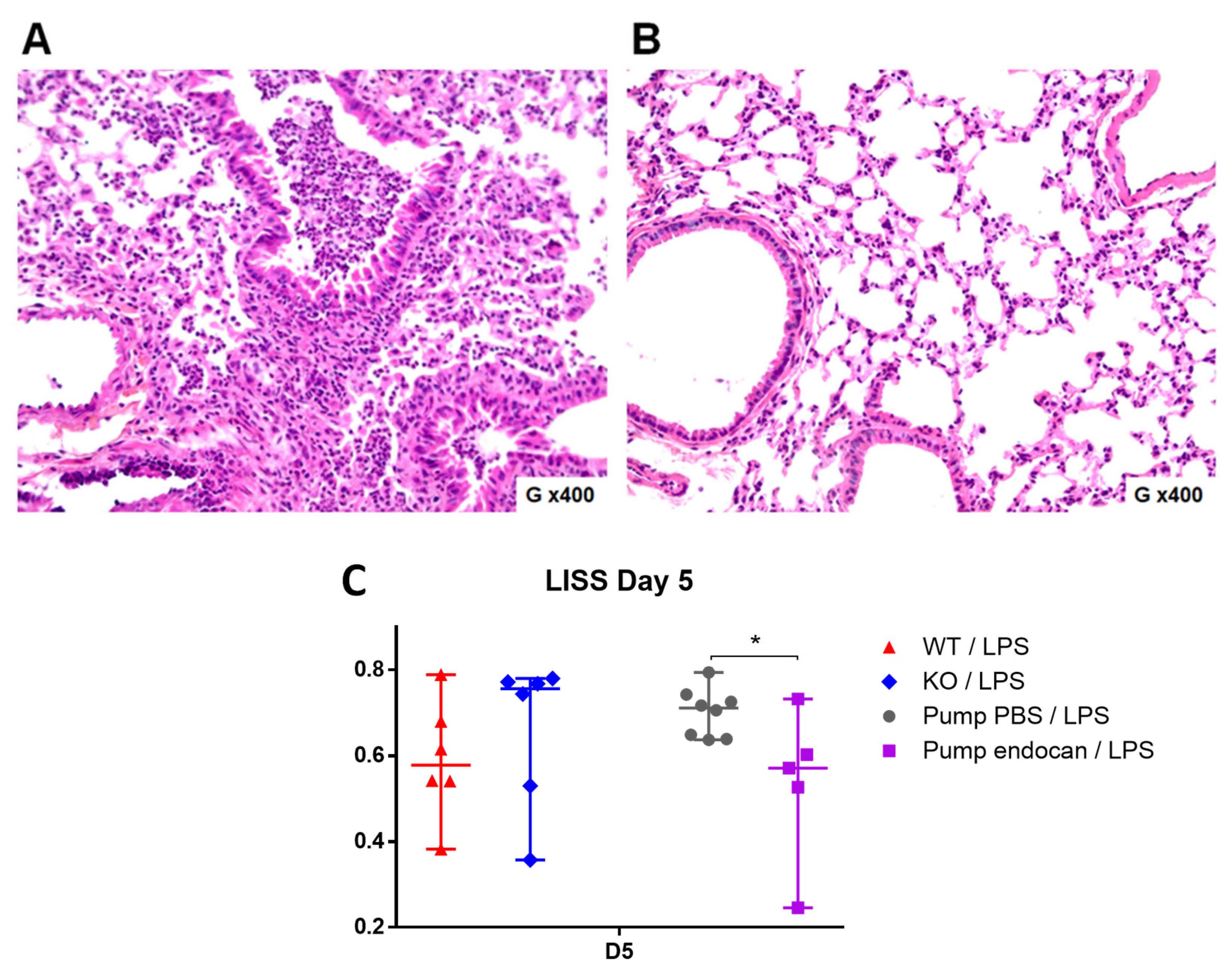
Figure 2). However, the LISS values were not different between the two groups (
Figure 4). In contrast, we found significant differences in BAL cellularity and SpO2 between the
Esm1−/− mice and their WT littermates. This discrepancy may be explained by a higher variability in LISS assessment, which does not strictly rely on a numerical result, but also on semi-quantitative evaluations for several parameters.
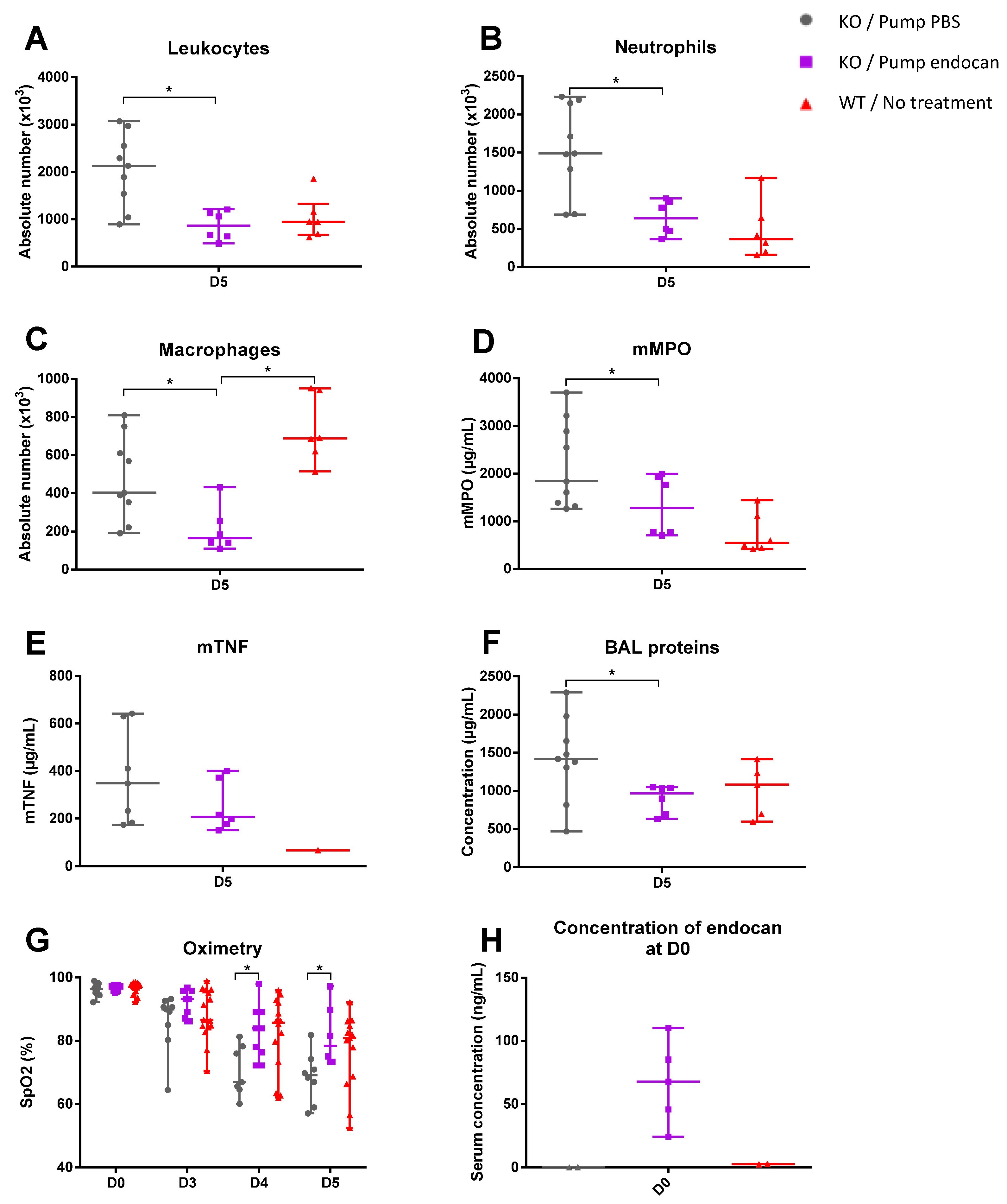
A model of endocan supplementation in Esm1−/− mice was therefore used to establish an in vivo model to assess clearly the physiological role of the glycosylated endocan, which is the unique form in humans. Indeed, the continuous infusion of murine glycosylated endocan in the Esm1−/− mice led to an alleviation of lung inflammation associated with an improvement in pulmonary physiological function when compared with the Esm1−/− mice treated with PBS.
Moreover, the continuous infusion of glycosylated murine endocan in the
Esm1−/− mice reduced the histological features of LPS-induced ALI, as reflected by the lower LISS values observed in the endocan-treated
Esm1−/− mice than those observed in the PBS-treated WT mice. Such low LISS values were not observed in the BALB/c mice [
5], confirming that the endogenous endocan can interfere with the exogenous type. Such interference may explain some conflicting results due to the use of a non-qualified glycosylation form of endocan in experimental models [
6].
Taken together, all these data reinforce the hypothesis that glycosylated endocan exerts anti-inflammatory effects, as it is likely to reduce the severity of lung inflammation and improve the pulmonary physiological function after LPS-induced ALI. These data are consistent with those in previously published studies, showing that the injection of human endocan in BALB/c mice alleviates the features of LPS-induced ALI and attenuates the respiratory physiological dysfunction [
5,
16].
Interestingly, when comparing Esm1−/− mice treated with endocan to WT mice, the only significant difference between them was regarding the macrophages, which showed greater counts in WT mice. Importantly, we found no significant difference between the WT mice and the endocan-treated Esm1−/− mice for all other variables explored, yet the conclusions to be drawn from these results remain limited, as the balance between glycosylated and non-glycosylated endocan in WT mice cannot be measured.
Our subcutaneous infusion model allowed for the achievement of sustainable high circulating concentrations of endocan, while previously published models failed to maintain such concentrations [
5]. However, several issues can be raised: (1) there was a variable diffusion (at equal loaded amounts, blood endocan levels vary from 25 to 116 ng/mL, far greater than values observed in WT mice); (2) we did not measure endocan blood levels beyond Day 0 in the endocan-treated
Esm1−/− mice, limiting the interpretability of our results, and (3) this remains a preventive model in which endocan starts to spread before the initiation of experimental ARDS. Therefore, much more work is required to refine the significance of endocan administration during the course of ALI.
Additional limitations to this study are: first, it would have been interesting to more precisely evaluate the changes in the M1 and M2 macrophage subsets in our models, yet this characterization was not performed in our study. Secondly, the accurate assessment of changes in vascular density and vascular leakage in endocan-treated mice would have been necessary to understand the changes in immune cell infiltration reported in our results. This was assessed through the measurement of BAL proteins, which remains an imperfect method for the estimation of this parameter. Thirdly, assessing the respective levels of non-glycosylated and glycosylated endocan in mice would have been informative for the interpretation of our results. However, there is no method currently available to perform the quantifications of non-glycosylated and glycosylated endocan. This latter point underlines the importance of our study, which is, to our knowledge, the first to describe an in vivo model able to clearly decipher the role of glycosylated endocan by exploring the action of the continuous infusion of endocan to achieve a supra-physiological concentration.

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}



