1. Introduction
Acute respiratory distress syndrome (ARDS) is characterized by enhanced pulmonary vascular permeability leading to a non-cardiac pulmonary edema. Excessive inflammation in the lungs is a major trigger for ARDS. The pathophysiology of ARDS includes inflammation; the uncontrolled activation of leukocytes, platelets, and coagulation systems; and increased permeability of the endothelial and alveolar epithelial barriers [
1]. Pulmonary vascular endothelial cells and lung epithelial cells generate pro-inflammatory signals during ARDS progression [
2]. The pathological phases of ARDS are classified into three stages: exudative, proliferative, and fibrotic. The exudative stage is characterized by increased pulmonary vascular permeability and increased neutrophils in the alveolar septum and airspaces, along with the death of epithelial and endothelial cells. The proliferative stage is characterized by fibroblast proliferation and type 2 pneumocyte hyperplasia [
3]. This phase is also important in terms of endothelial repair and regeneration, which are essential steps for recovering from ARDS [
4]. Although numerous studies have focused on the mechanisms and treatment strategies of ARDS, no effective drug therapies have been established.
CD26/dipeptidyl peptidase-4 (DPP4) is a transmembrane protein expressed in a variety of cells that also exists as a soluble protein in tissue and the circulation. CD26/DPP4 exerts its peptidase activity towards various proteins, such as incretin hormones, and CD26/DPP4 inhibitors have been developed as therapeutic agents for diabetes. In addition, CD26/DPP4 can participate in immune stimulation and the promotion of inflammation [
5,
6]. Previous studies showed that pharmacological CD26/DPP4 inhibition has cardiovascular protective and anti-inflammatory effects in the vessels [
7].
CD26/DPP4 is widely expressed in a variety of cell types in lung tissue, such as type I and II alveolar cells, alveolar macrophages, and vascular endothelia [
8,
9,
10], and has been recently suggested to be a therapeutic target in lung diseases [
11]. For example, previous reports have indicated that CD26/DPP4 inhibitors have a protective effect on lung ischemia-reperfusion injury through promoting the recruitment of endothelial progenitor cells by retaining SDF-1/CXCL12 activity [
12,
13]. Another study suggested that CD26/DPP4 inhibitors can be novel prophylactic drugs for chronic allograft dysfunction after clinical transplantation [
14]. Furthermore, recent studies have reported that CD26/DPP4 contributed to non-typeable H. influenzae-induced lung inflammation in COPD [
15], and can participate in the pathogenesis of pulmonary hypertension [
16]. We previously demonstrated that CD26/DPP4 inhibition by sitagliptin ameliorated lipopolysaccharide (LPS)-induced lung injury in mice through its anti-inflammatory effects on pulmonary endothelial cells, indicating a relationship between pulmonary endothelium in ARDS and CD26/DPP4 [
17]. However, the mechanistic roles of CD26/DPP4 in the pulmonary endothelium at each stage of ARDS remain unclear.
Studies of pulmonary vascular endothelial cells are essential for understanding the mechanism of ARDS because these cells exert multiple relevant effects, such as the generation of inflammatory mediators and the regulation of intracellular adhesion, which are related to the endothelial barrier [
2]. Pulmonary endothelial injury as a key disease mechanism and the contribution of progenitor cells in mediating endothelial repair have also been suggested as important processes in lung injury [
18]. We recently reported that CD26/DPP4 inhibition has anti-inflammatory effects on human lung microvascular endothelial cells (HLMVECs) [
17], indicating that CD26/DPP4 is involved in lung endothelial cell functions related to the pathophysiology of ARDS.
In this study, we examined the roles of CD26/DPP4 in ARDS pathology, focusing on pulmonary vascular endothelial cells. We performed an in vitro transcriptome analysis and functional experiments employing specific DPP4 knockdown using microRNA on HLMVECs, to evaluate the response of these cells to the pro-inflammatory stimulus LPS.
2. Materials and Methods
2.1. Human Lung Endothelial Cell Culture
HLMVECs were obtained from Lonza (Basel, Switzerland) and cultured in endothelial growth medium-2 supplemented with 10% fetal bovine serum. The cells were incubated at 37 °C in a 5% CO2 incubator and used at passages 6–8 for all experiments.
2.2. Reagents
Non-specific control siRNA (siCon) (Cat# 4390843: Silencer™ Select Negative Control No. 1 siRNA) and DPP4 siRNA (Cat# 4392421: siRNA ID s4255) were purchased from Thermo Fisher Scientific (Waltham, MA, USA). Escherichia coli LPS (O127:B8, L3137) and all other reagents were purchased from Merck (Darmstadt, Germany), unless otherwise specified.
2.3. Transfections with Silencing RNA
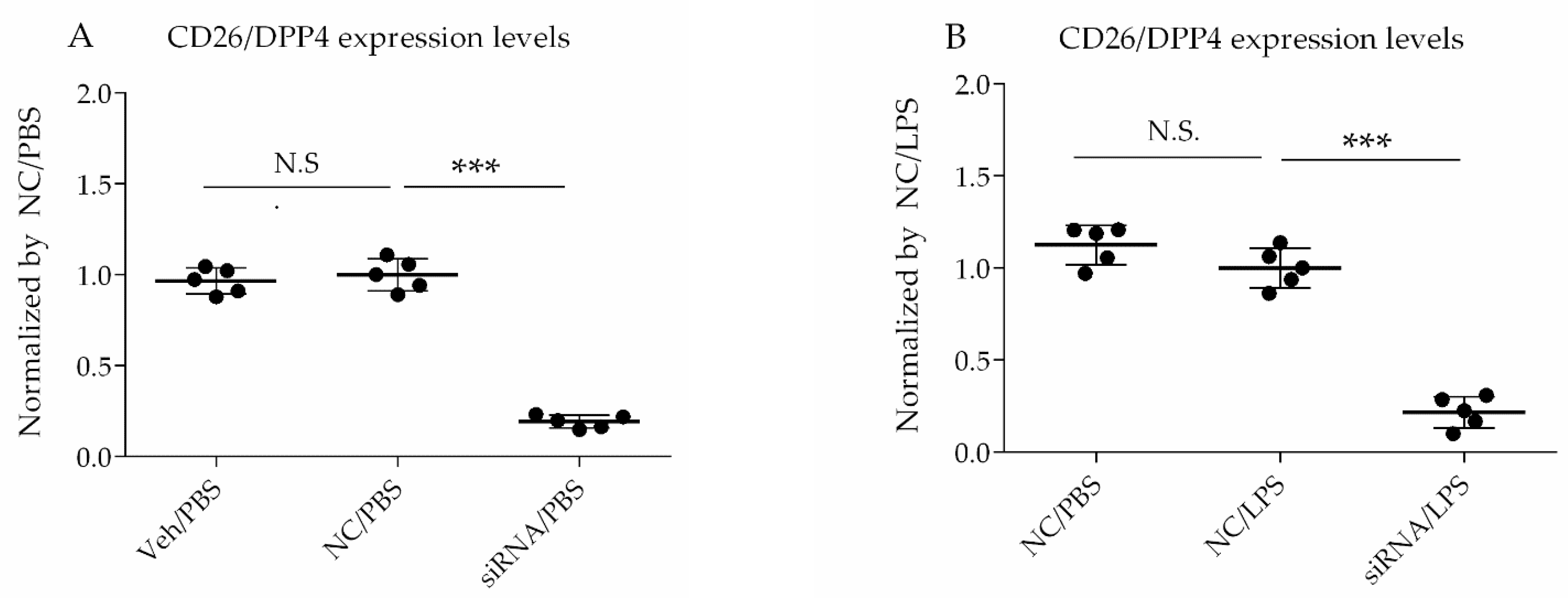
For siRNA transfection, the Lipofectamine™ RNAiMAX Transfection Reagent (Invitrogen, Carlsbad, CA, USA) was used. The cells were transfected with siRNA at a 60% confluence, according to the manufacturer’s protocol, and then challenged with LPS at 72 h after siRNA treatment. The selective silencing of CD26/DPP4 was confirmed using flow cytometry analysis and quantitative PCR.
2.4. RNA Sequencing
Four primary cell lines of HLMVECs were used; the donors included a 44-year-old Hispanic male, 38-year-old Caucasian male, 59-year-old Hispanic female, and 30-year-old Hispanic female, according to the information from Lonza. Total RNA was isolated from the HLMVECs and stored in Isogen (Nippon Gene, Tokyo, Japan). Two milliliters of this solution was vigorously vortexed and then centrifuged after adding 400 μL of chloroform. The supernatants were removed, and 40 μg of glycogen (Roche, Basel, Switzerland) was added. RNA was precipitated by adding 1000 μL of isopropyl alcohol. The solution was vortexed vigorously and centrifuged. The RNA pellets were washed with 75% ethanol and then dissolved in 10 μL RNase-free water. The concentration and quality of the RNA were verified using an Agilent 2100 Bioanalyzer (Agilent Technologies, Santa Clara, CA, USA).
Purified total RNA (500 ng) with an RIN value of >9 was used for RNA library preparation, according to the instructions of the QuantSeq 3′mRNA-Seq Library Prep Kit FWD for Illumina (Lexogen, Vienna, Austria). The libraries were amplified using 12 cycles of polymerase chain reaction (PCR). The RNA libraries were sequenced using an Illumina NextSeq 500 system (75 cycles; San Diego, CA, USA). FASTQ files were prepared with reads using bcl2fastq ver2.17 (Illumina). The quality of the FASTQ sequence data was assessed using FastQC v0.11.9 (Illumina). After removing the adapter sequences from the raw reads, trimmed reads were aligned using STAR v2.7.6a to the GRCh38 human reference genome. Reads per million values were calculated using samtools v1.11 and htseq-count v0.12.4.
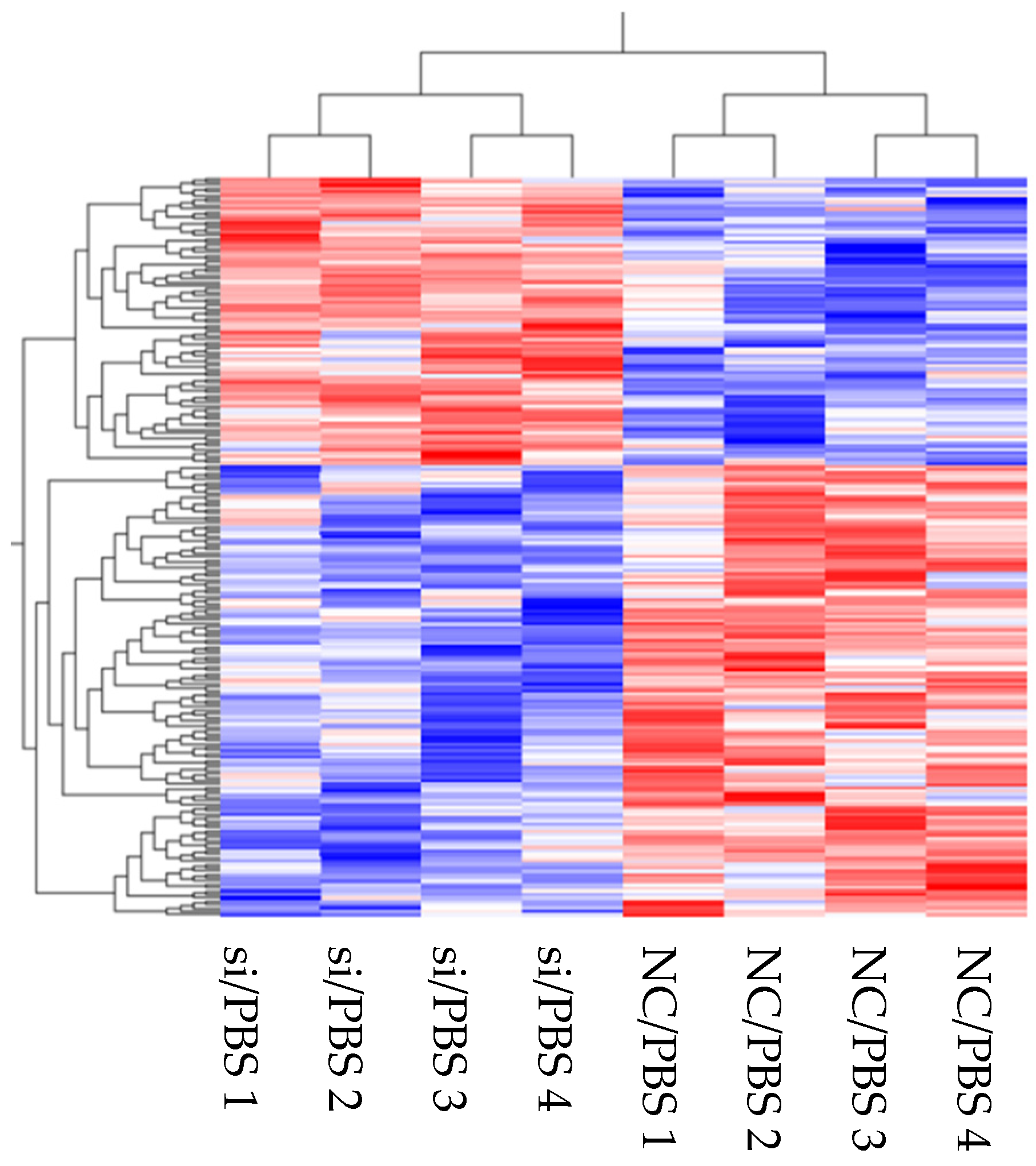
2.5. Differentially Expressed Genes (DEGs), Gene Ontology (GO), and the Kyoto Encyclopedia of Genes and Genomes (KEGG) Pathway Enrichment Analysis
The expression levels of the genes identified in the transcriptome were normalized and compared. Hierarchical clustering and heat maps were created using the Qlucore omics explorer software program (Qlucore AB, Lund, Sweden). Differentially expressed genes (DEGs) were detected for each gene between the two type samples. The fold-changes between each group were >1.5 (upregulated) or <0.66 (downregulated) (
p < 0.1). Significantly, over-represented functional categories were identified using Enricher [
12]. Genes significantly upregulated or downregulated in “siRNA for
DPP4 (siRNA) and PBS vs. negative control of siRNA (NC) and PBS”, “NC/PBS vs. NC/LPS”, or “NC/LPS vs. siRNA/LPS” were annotated. Subsequently, the gene ontology (GO) terms and the Kyoto Encyclopedia of Genes and Genomes (KEGG) pathways were identified. Selected GO terms and KEGG pathways were considered as significant at
p < 0.05.
2.6. Flow Cytometry Analysis
Cultured HLMVECs were detached from culture plates using STEMPRO ACCUTASE (Thermo Fisher Scientific), pretreated with Human TruStain FcX (BioLegend, San Diego, CA, USA) to block Fc receptors for 10 min, followed by incubation with anti-human CD26-phycoerythrin (BioLegend) or anti-human ICAM-1-phycoerythrin (BioLegend) in the dark at 4 °C for 20 min. Cell fluorescence was measured using a BD FACSCanto™ II (BD Biosciences, Franklin Lakes, NJ, USA), and the data were analyzed using FlowJo software (TreeStar, Ashland, OR, USA).
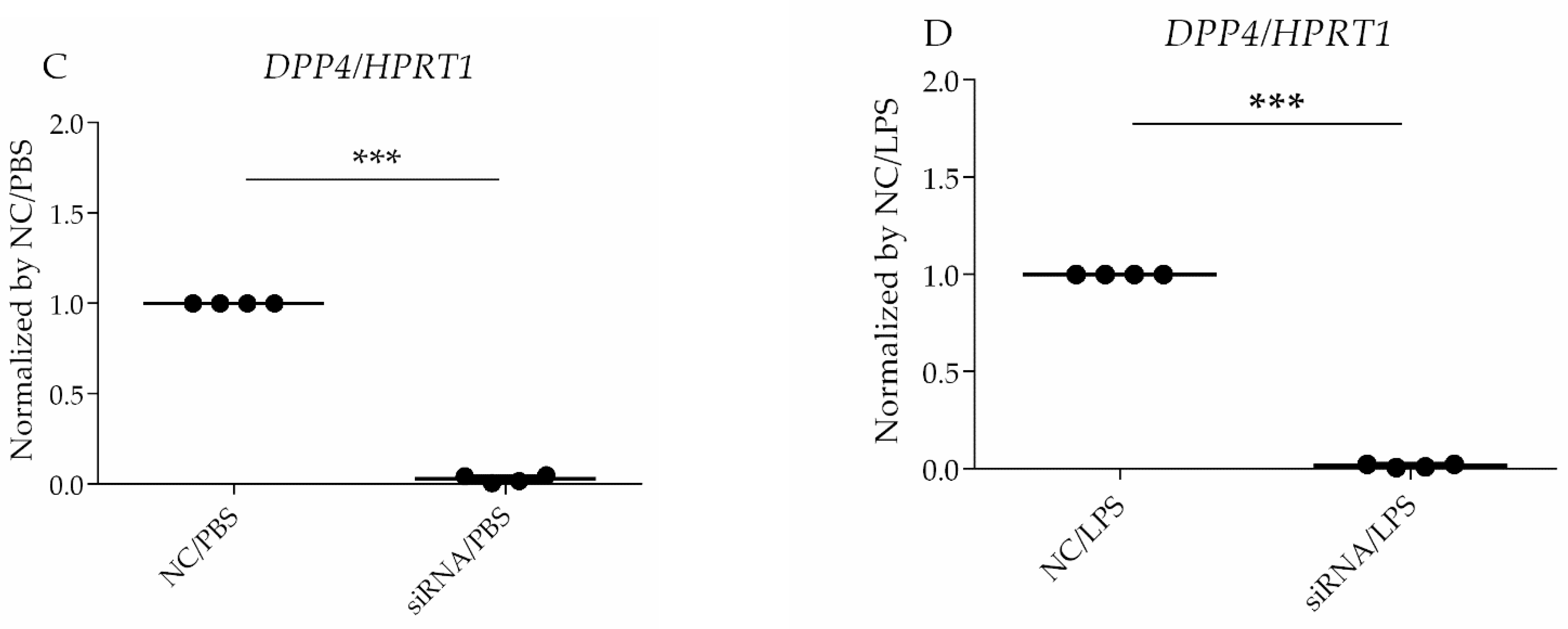
2.7. Real-Time Quantitative PCR
Total RNA from HLMVECs was extracted using TRIzol and a Direct-zol RNA MiniPrep Plus kit (Zymo Research Corporation, Irvine, CA, USA). The RNA was reverse-transcribed via PCR with SuperScript IV VILO Master Mix (Thermo Fisher Scientific) to synthesize single-stranded cDNA. The cDNA samples were amplified via quantitative PCR (qPCR) with Taqman Fast Universal PCR Master Mix (Thermo Fisher Scientific), using the GeneAmp PCR System (Thermo Fisher Scientific). Specific primers were designed using web software from the Universal ProbeLibrary Assay Design Center (Roche Applied Science, Waltham, MA, USA). The expression level of each target gene was normalized to the HPRT1 threshold cycle (CT) values and calculated using the 2−ΔΔCt method. ΔΔCT = (target gene CT of experimental group − reference gene CT of experimental group) − (target gene CT of control group − reference gene CT of control group).
2.8. Enzyme-Linked Immunosorbent Assay (ELISA) of Conditioned Medium
Cultured HLMVECs were treated with an siRNA or vehicle (water) for 72 h, followed by treatment with LPS (1 µg/mL) or vehicle (PBS) for 6 h. The harvested culture medium was centrifuged (500× g for 10 min at 4 °C), and the supernatant was collected for ELISA. Human interleukin (IL)-6 and IL-8 levels were measured using ELISA kits (BioLegend), according to the manufacturer’s protocols.
2.9. Cell Proliferation Assay
A cell proliferation assay was performed to assess the number of viable cells using Cell Counting Kit-8 (WST-8) (Dojindo Molecular Technologies, Kumamoto, Japan), according to the manufacturer’s protocol. HLMVECs (5000 cells/well) treated with siRNA or vehicle for 72 h were detached using ACCUTASE and precultured on a 96-well plate for 24 h, followed by treatment with LPS (1 µg/mL) or PBS (-) for 18 h. The cells were cultured with 10 μL WST-8 in each well at 37 °C for 2 h. Cell viability was measured as the absorbance (optical density (OD)) read at 450 nm using a microplate reader. The result was calculated using the following formula: cell viability = (treatment group OD − blank group OD)/(control group OD − blank group OD).
2.10. Wound Healing Assay
A wound was established by manually scraping the confluent cell monolayer using a 200 µL pipette tip. Initial images of the culture plates were acquired as the reference points. After incubating the plates for 7 h at 37 °C in a 5% CO2 incubator, a second image was acquired. The wounded region lacking cells was measured using ImageJ software (National Institutes of Health, Bethesda, MD, USA).
2.11. Tube Formation Assay
Matrigel (Corning, Inc., Corning, NY, USA) was thawed at 4 °C, and then 96-well plates were coated with 50 µL/well of Matrigel. The cells were detached from culture plates using ACCUTASE, and the volume was adjusted with a complete medium to obtain 2 × 10
4/200 µL, and the cell mixture (200 µL) was added to each Matrigel-coated well. The plate was incubated for 6 h at 37 °C in a 5% CO
2 incubator. Briefly, for the quantification of the tube formation, the number of branch nodes was counted and the lengths of all the tubes were measured per one field of a microscope image of each well, as shown in
Supplementary Figure S1 [
19].
2.12. In Vitro Vascular Permeability Assay
Endothelial cell monolayer permeability was assessed using an in vitro vascular permeability assay kit (Millipore, Billerica, MA, USA). The inserts were coated with collagen and included a membrane with 1 µm pores. HLMVECs were treated with siRNA for 3 days and then cultured in the inserts on a 24-well plate to form an endothelial cell monolayer over the membrane (3 × 105 cells per insert). The cells were incubated for another 2 days, followed by LPS challenge (1 µg/mL) for 4 h. Vascular permeability was quantified using fluorescein isothiocyanate (FITC)-dextran. FITC-dextran diluted by 1:40 with complete culture medium was added to the inserts (150 µL each), and the plate was incubated at room temperature for 20 min. The reaction was stopped, and the fluorescence signal was detected using a Qubit 4 fluorometer (Thermo Fisher Scientific) at 485 nm excitation.
2.13. Statistical Analysis
The results are expressed as the mean ± standard deviation (SD). One-way analysis of variance was used for multiple-group comparisons, followed by Tukey’s post hoc test. Student’s t-test was used to compare two groups. Statistical analyses were performed using GraphPad Prism 6 software (GraphPad, Inc., La Jolla, CA, USA). The statistical significance was set at p < 0.05.
4. Discussion
We previously reported that CD26/DPP4 inhibition by sitagliptin attenuates LPS-induced lung injury in mice and has anti-inflammatory effects on LPS-stimulated HLMVECs [
17]. In the present study, we substantially advance this prior work by performing in vitro experiments using
DPP4 siRNA to better understand the detailed mechanisms by which CD26/DPP4 expression is related to the functions of HLMVECs under baseline and inflammatory conditions. We employed transcriptome analysis to detect DEGs and identify the possible functions affected by
DPP4 knockdown, followed by confirmatory functional experiments in HLMVECs. These results suggest that CD26/DPP4 expression is related to multiple important functions in HLMVECs, including inflammation, barrier function, and regenerative processes, all of which are all known as hallmarks of ARDS pathophysiology and essential processes for recovery from lung injury.
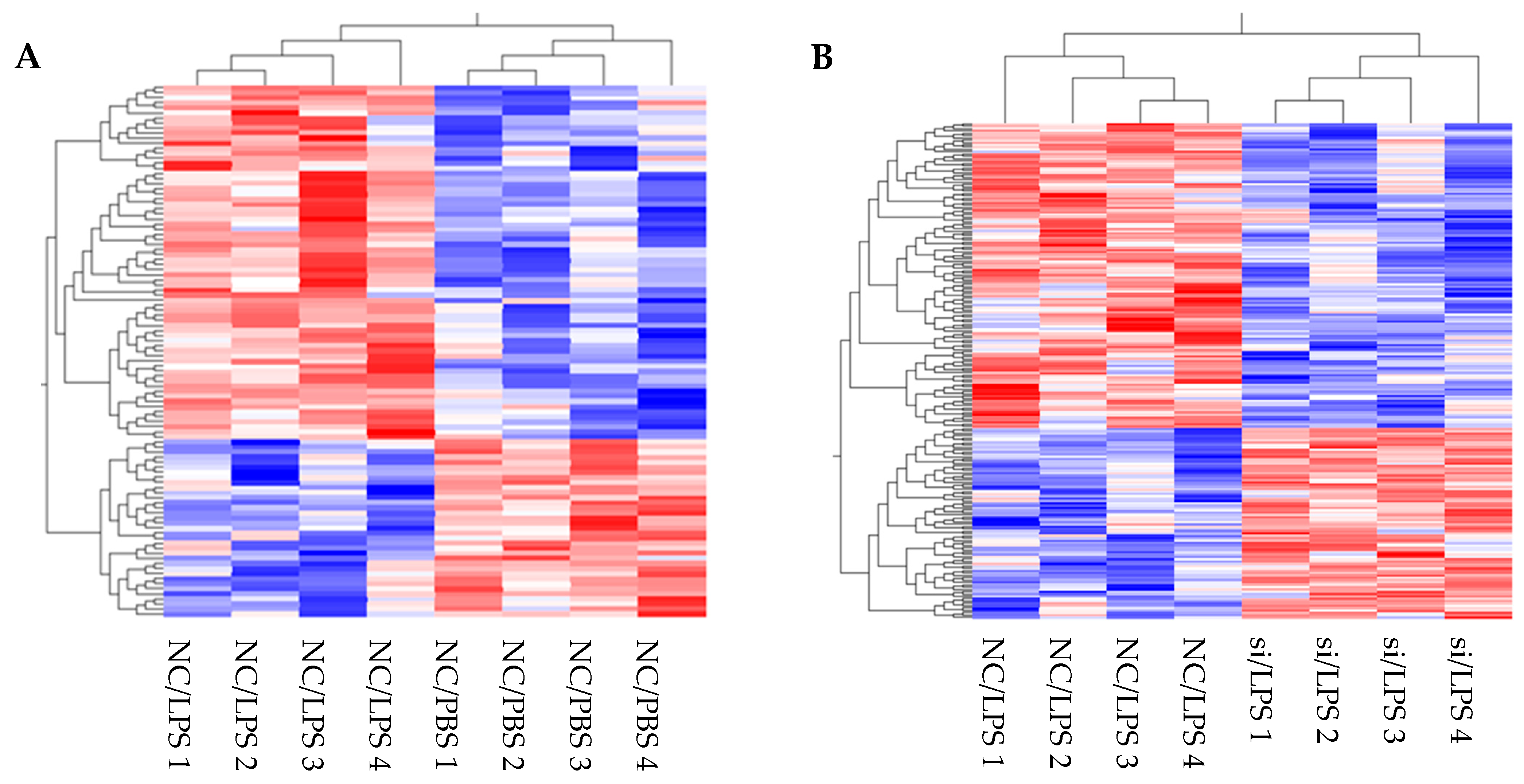
First, transcriptome analysis of enriched DEGs suggested changes in multiple cell functions and pathways following
DPP4 knockdown or LPS stimulation. Under baseline conditions without LPS stimulation, enrichment analysis (GO and KEGG pathways) suggested that CD26/DPP4 reduction affects inflammation, as indicated by terms such as MAPK or TNF signaling pathways (
Table 2B), as well as proliferation and angiogenesis, as indicated by terms such as positive regulation of endothelial cell proliferation (GO:0001938) and angiogenesis involved in wound healing (GO:0060055) (
Table 2A,B). Under the inflammatory state induced by LPS, enrichment analysis suggested that upregulation occurs in the TLR4 signaling pathway and other pro-inflammatory pathways, such as TNF signaling, NF-kappa B signaling, chemokine signaling, PI3K-Akt signaling, and MAPK signaling (
Table 3). LPS also downregulates tight junctions, which can lead to increased permeability (
Table 3). In addition, enrichment analysis suggested that a reduction of CD26/DPP4 expression via siRNA alters LPS-induced inflammation, as indicated by the terms TNF signaling pathway. Endothelial proliferation and angiogenesis can also be affected by the
DPP4 siRNA treatment, as indicated by terms such as positive regulation of endothelial cell proliferation (GO:0001938), regulation of endothelial cell proliferation (GO:0001936), endothelial cell proliferation (GO:0001935), and positive regulation of vasculature development (GO:1904018). Monolayer permeability can also be affected by
DPP4 siRNA treatment, as indicated by the terms actin filament bundle assembly (GO:0051017), actin filament bundle organization (GO:0061572), and focal adhesion (
Table 5A,B). Although previous studies demonstrated that LPS challenge provokes inflammation, the enhancement of monolayer permeability, and regenerative processes in pulmonary vascular endothelial cells [
2], transcriptome analysis of HLMVECs stimulated by LPS has not been performed under these conditions. Our results suggest that comprehensive mechanistic pathways are involved in mediating the effects of LPS. Therefore, these transcriptome analysis results are useful for further understanding and exploring the functional responses of HLMVECs during LPS-induced inflammation.
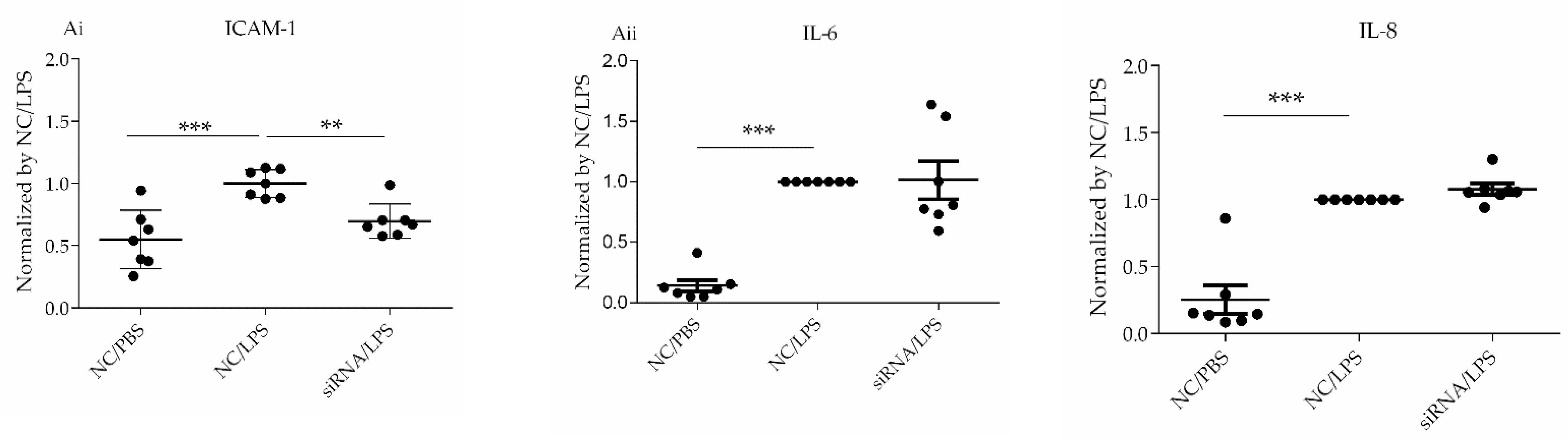
Based on the results of the transcriptome data indicating a relationship between inflammation and CD26/DPP4, we characterized the possible differences in the expression of TNFα, IL-6, IL-8, and ICAM-1 at gene and protein levels, which are related to neutrophil inflammation, a hallmark of acute lung injury. The raw data for transcriptome analysis are summarized in
Table 1,
Table 3 and
Table 5. We observed wide variations in gene expression levels, which may have occurred in part because we employed four lots of HLMVECs to make the results more universal. Heterogeneity, in the responses of these primary cells, is expected and likely contributed to the wide variability in gene expression.
IL6,
CXCL8, and
ICAM1 were upregulated by LPS, whereas
TNF remained unchanged after LPS stimulation (
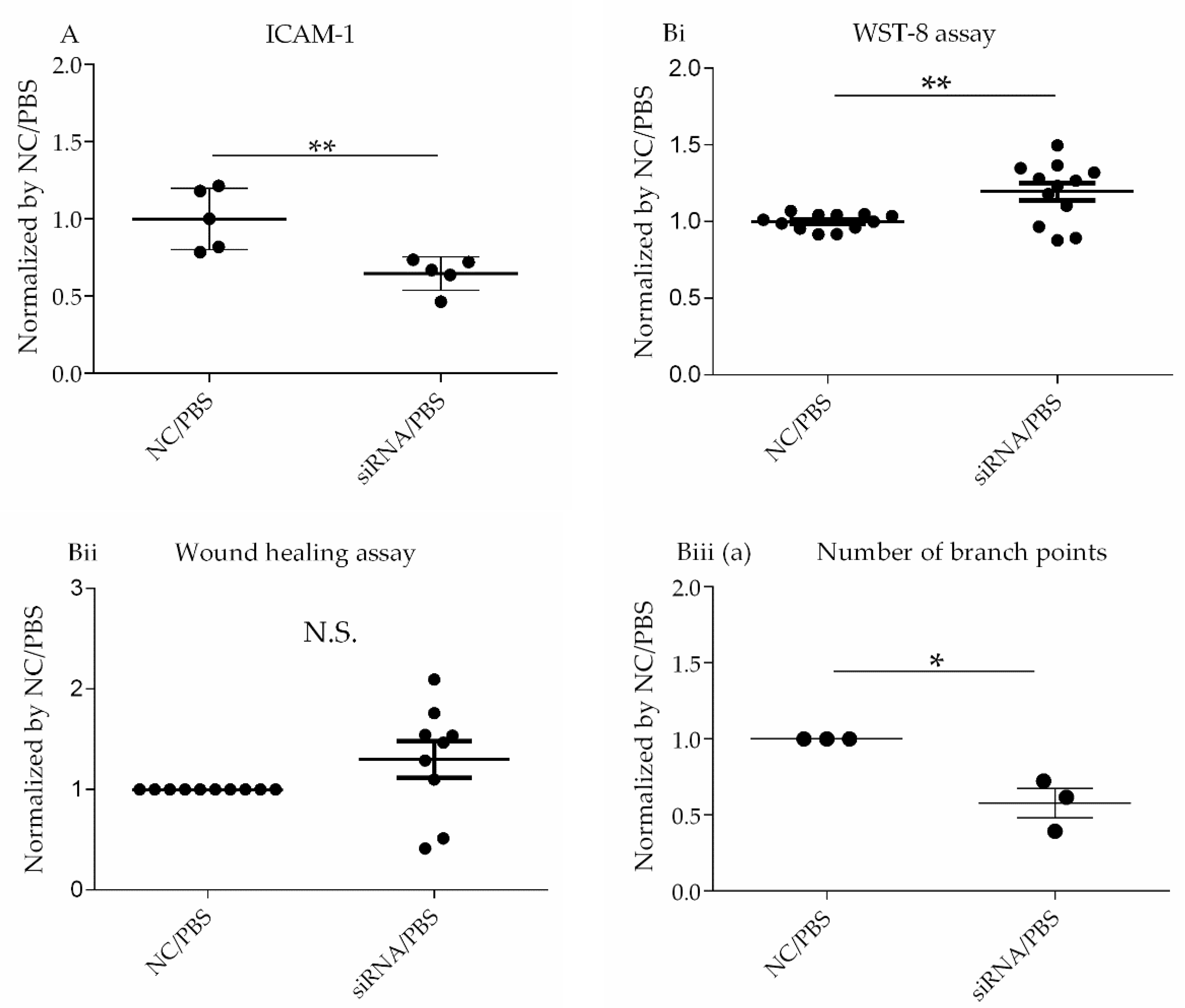
Table 4). ICAM1 trended toward downregulation at both the gene and protein levels regardless of LPS stimulation (
Table 1 and
Table 6,
Figure 3 and
Figure 5Ai), whereas IL-6 and IL-8 expression at gene and protein levels were unchanged (
Table 1 and
Table 6,
Figure 3 and
Figure 5Aii). We previously demonstrated that pharmacological CD26/DPP4 inhibition in cultured HLMVECs reduced ICAM1 and IL-6 levels [
17]. Together, CD26/DPP4 inhibition can exert an anti-inflammatory effect by reducing ICAM levels in HLMVECs, whereas other mechanisms related to pharmacological CD26/DPP4 inhibition, but not reduced CD26/DPP4 expression, can exist to reduce IL-6 release.
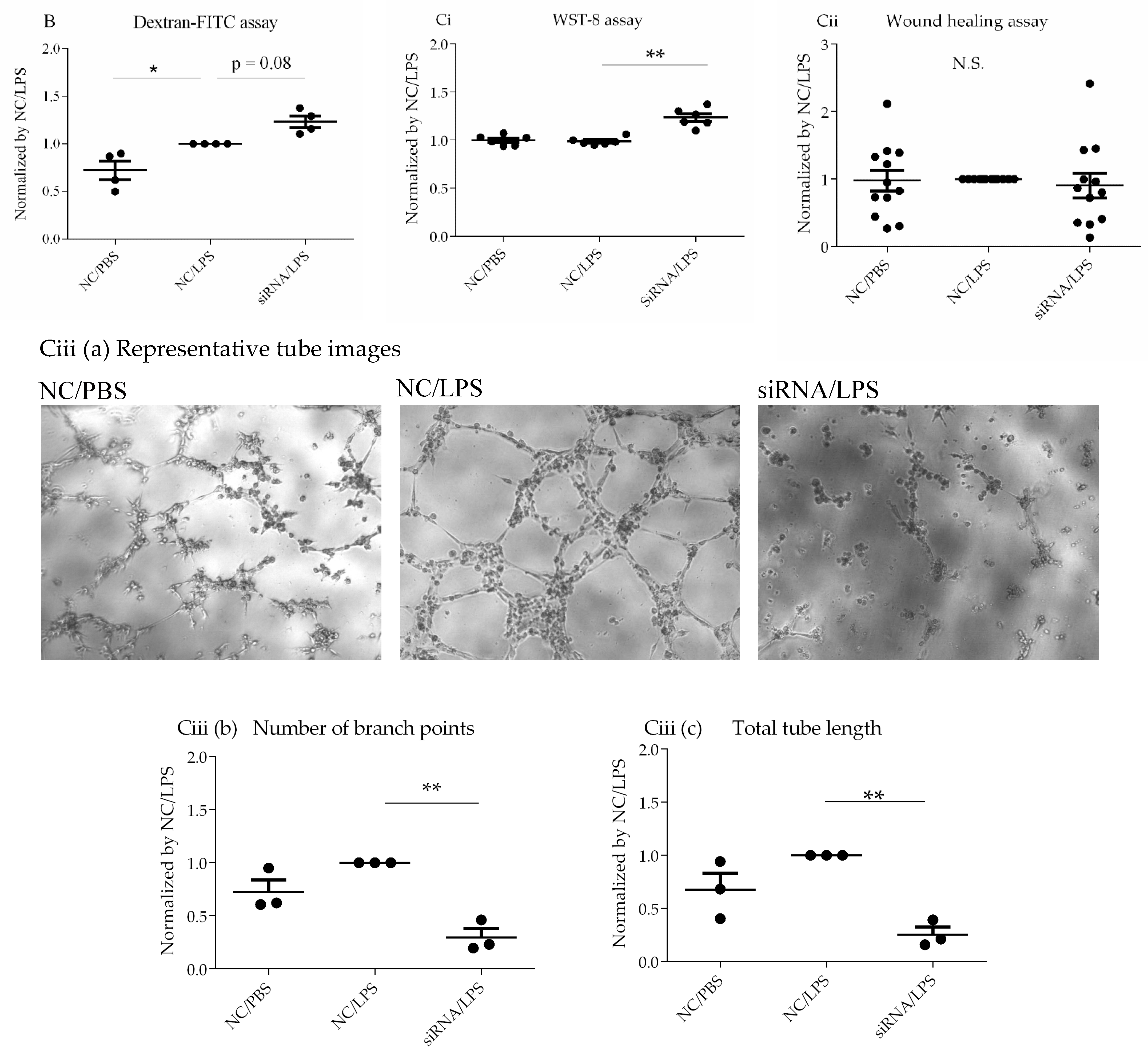
Based on the results of transcriptome analysis, suggesting a potential role for CD26/DPP4 in the barrier function of the pulmonary endothelium, we evaluated the effects of CD26/DPP4 suppression on the monolayer permeability of HLMVECs after LPS. Under the baseline conditions, without LPS stimulation, no terms related to barrier function were identified after
DPP4 knockdown via siRNA. However, after LPS stimulation, several terms related to monolayer permeability were enriched, and the term “tight junction” was identified after CD26/DPP4 suppression using siRNA. Tight junctions, intercellular adherent junctions, and gap junctions in endothelial monolayers are major determinants of barrier function between the blood and interstitial spaces in the lung [
2] [
30] [
31]. Therefore, we evaluated the effect of CD26/DPP4 suppression on monolayer permeability. Although our data are limited, the results revealed a trend toward enhanced permeability after LPS challenge, which was further augmented by
DPP4 knockdown, suggesting that CD26/DPP4 can be involved in regulating monolayer barrier permeability in HLMVECs. Regarding the role of CD26/DPP4 in the permeability of endothelial monolayers, we previously reported that high concentrations of the pharmacological CD26/DPP4 inhibitor sitagliptin enhanced the permeability of the HLMVEC monolayer. In fact, clinical case reports have described the development of angioedema in some patients receiving CD26/DPP4 inhibitors (anagliptin and sitagliptin) [
32,
33]. Our results can provide a mechanistic explanation for the development of angioedema in these patients. Therefore, caution may be required when considering the use of CD26/DPP4 inhibitors in patients who are at an increased risk of vascular permeability. In contrast, it has been reported that pharmacological CD26/DPP4 inhibition by diprotin A helps maintain the VE-cadherin and adherens junctions of HUVECs under hypoxia by suppressing β-catenin cleavage [
34]. Taken together, the role of CD26/DPP4 in regulating endothelial barrier function can be affected by the environment and/or types of endothelial cells and/or CD26/DPP4 inhibitors; however, further research is warranted to explore these details.
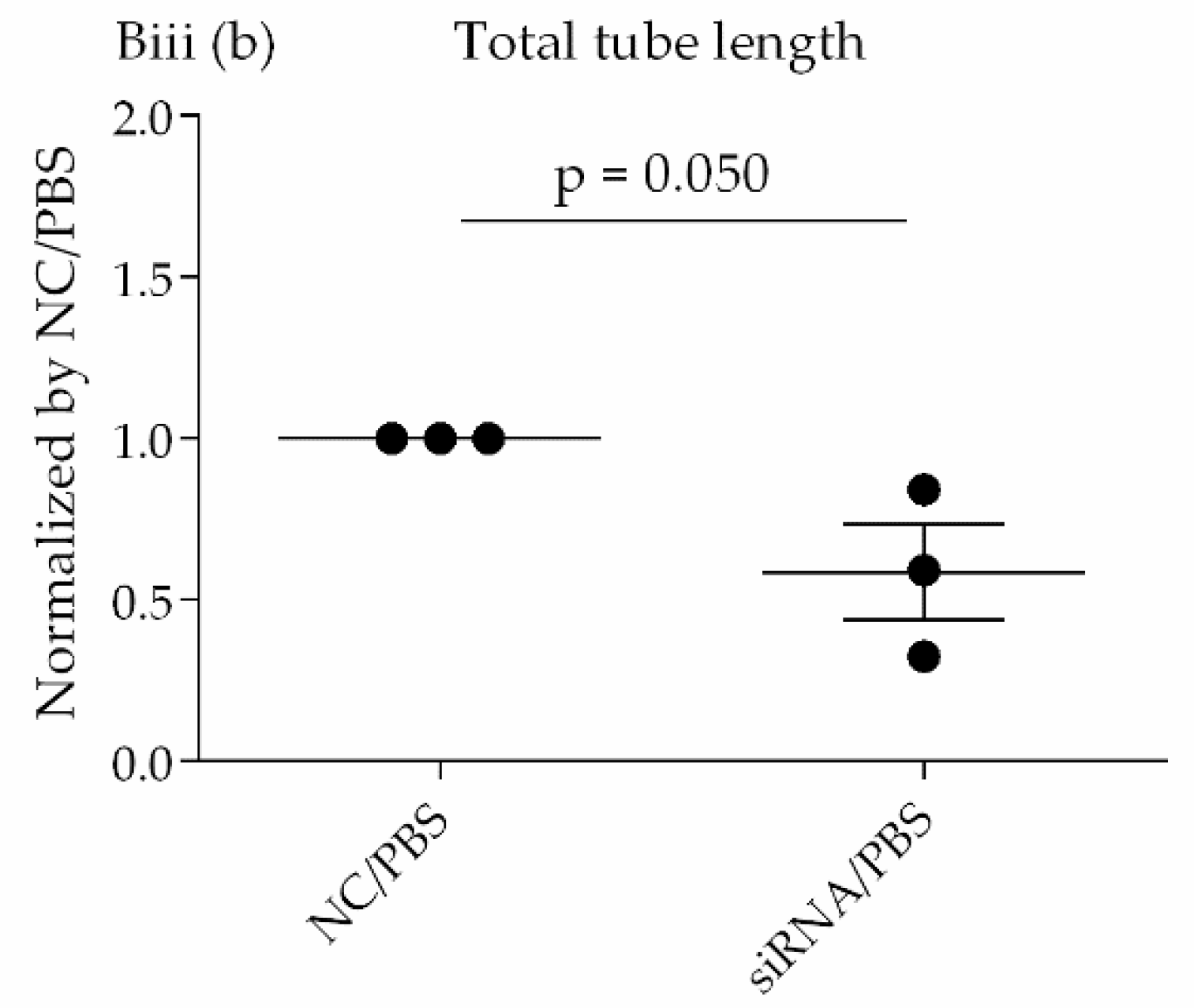
Transcriptome analysis suggested that CD26/DPP4 expression can be involved in the regenerative processes of the pulmonary endothelium. Based on these results, we evaluated the possible differences in related cell functions, such as proliferation, migration, and lumen formation. The suppression of CD26/DPP4 by siRNA promoted the cell proliferative capacity, but no changes in the migratory ability or suppression of tube formation were observed. Previous studies suggested that endothelial regeneration is required for the recovery from ARDS [
22]; therefore, CD26/DPP4 suppression can promote endothelial repair, leading to the recovery from ARDS. However, how the migration or angiogenesis functions are related to the recovery from the pathophysiology of ARDS is unclear and requires further analysis.
Our study had several limitations. First, the details of localization and characterization of CD26/DPP4 in HLMVECs remain to be clarified, although CD26/DPP4 is known to be expressed on the surface of various cell types in the lung, and its expression levels on the cell surface of HLMVECs were suppressed via siRNA using flow cytometry analysis, in the present study (
Figure 1A,B). Second, the effects of CD26/DPP4 overexpression in HLMVECs were not evaluated. Third, whether CD26/DPP4 suppression affects HLMVECs stimulated by LPS at different time points should be examined. Fourth, additional assays of monolayer permeability can be useful to better characterize the effects of CD26/DPP4 expression on the HLMVEC barrier function. Fifth, our results were obtained in vitro, and additional in vivo studies are needed to evaluate the effects of
DPP4 knockdown in an animal model of lung injury. Finally, and importantly, the effectiveness of CD26/DPP4 inhibition in patients with ARDS must be determined. Further research is ongoing to better understand these issues.



 ,
, 








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}