
Hepatoprotective Effects of Hyaluronic Acid-Preconditioned Bone Marrow Mesenchymal Stem Cells against Liver Toxicity via the Inhibition of Apoptosis and the Wnt/β-Catenin Signaling Pathway
 , , , , , ,
, , , , , ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. In Vitro Study
BMSC Isolation, Expansion, Characterization, and HA Treatment
2.2. Assessment of Cell Proliferation and Viability
2.3. Experimental Animals
2.4. Study Design and Animal Groups
2.5. Biochemical Assay
2.6. Evaluation of Oxidant/Antioxidant Parameters
2.7. Real-Time Polymerase Chain Reaction
2.8. Histopathological Studies
2.9. Immunohistochemistry
2.10. Statistical Studies
3. Results
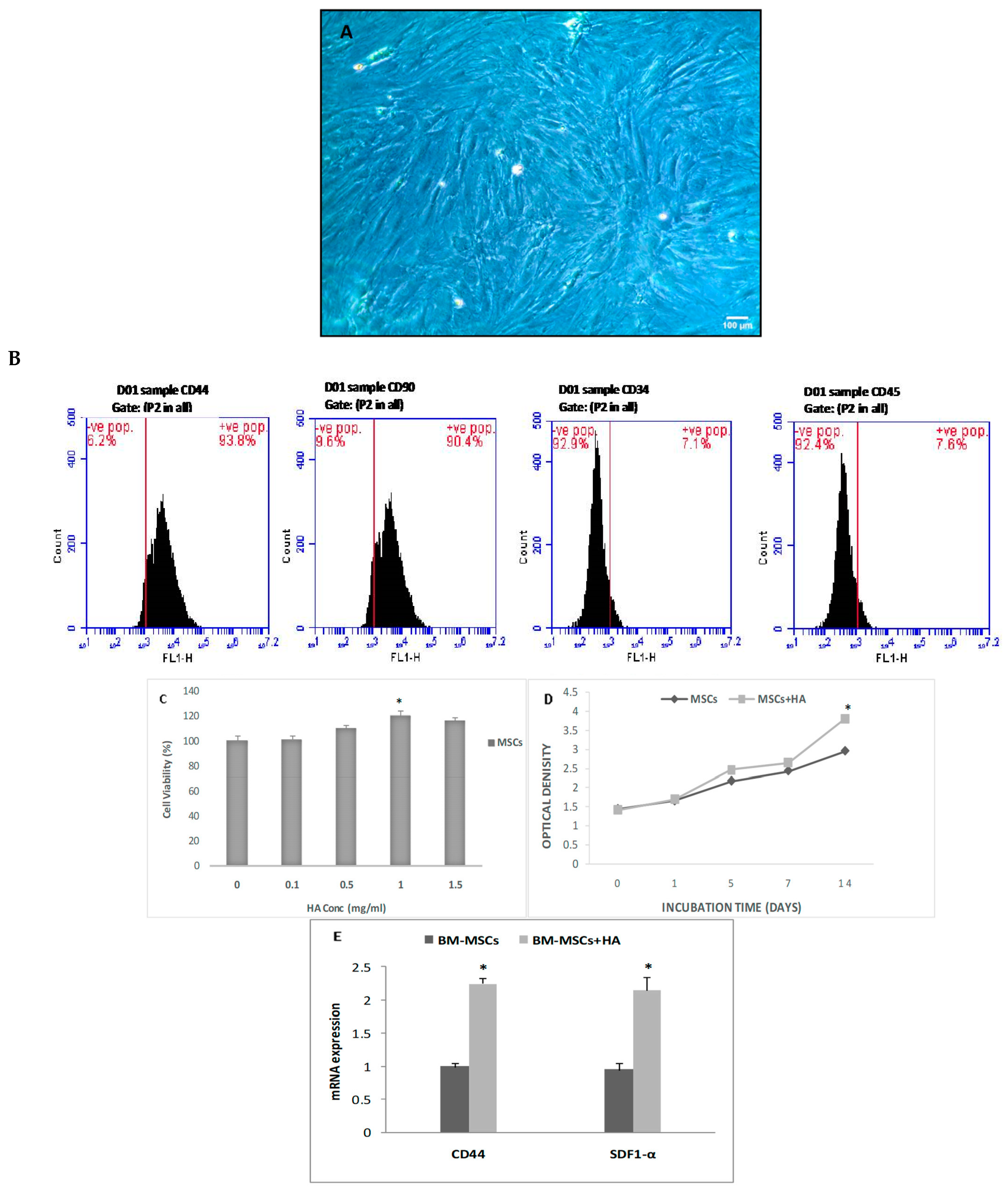
3.1. In Vitro Treatment of BMSCs with HA
Characteristics of BMSCs, Cell Viability, and Proliferation
3.2. Impact of HA-Treated BMSCs on Liver Function
3.3. Effect of BMSCs on Oxidative Stress in Liver Tissues
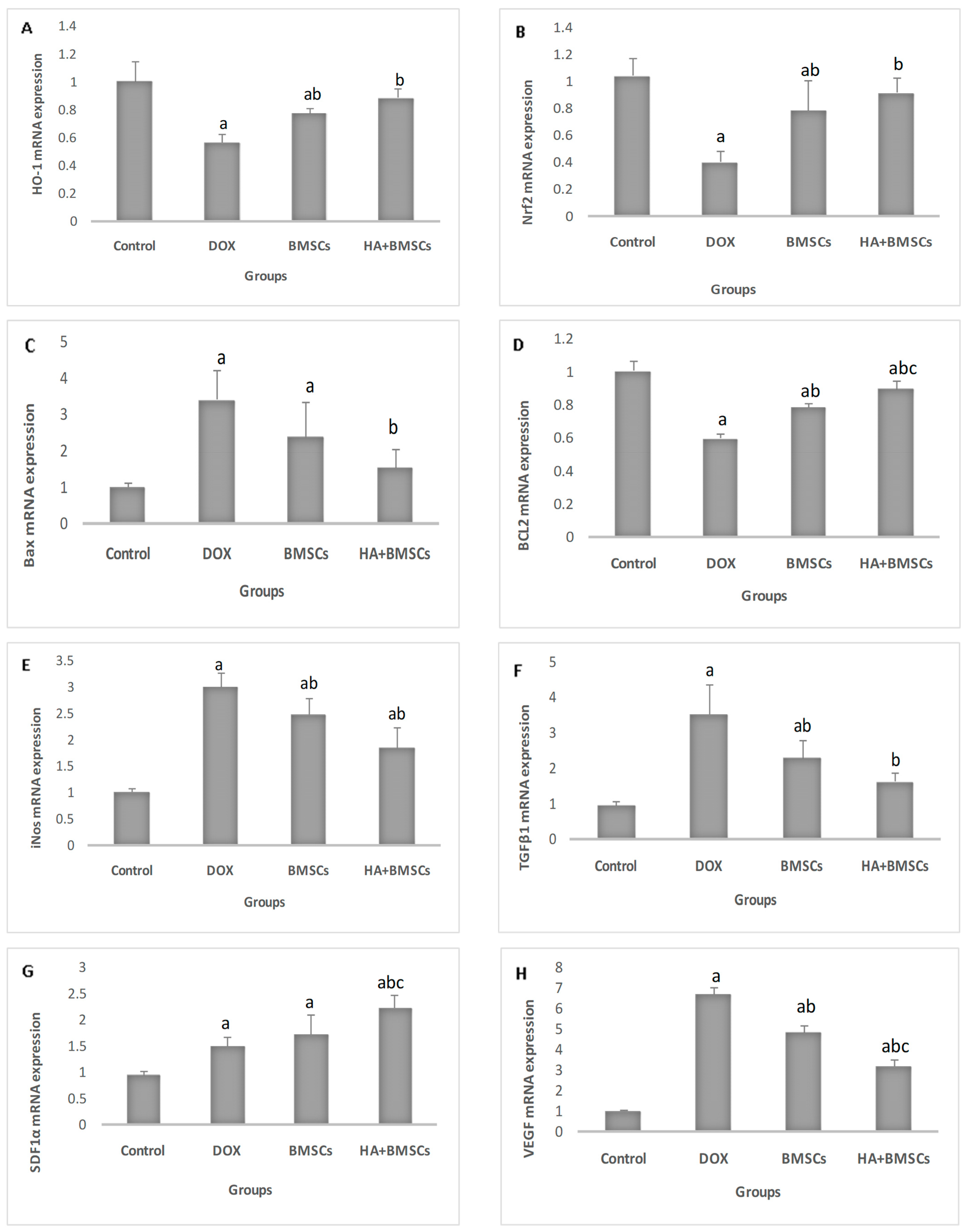
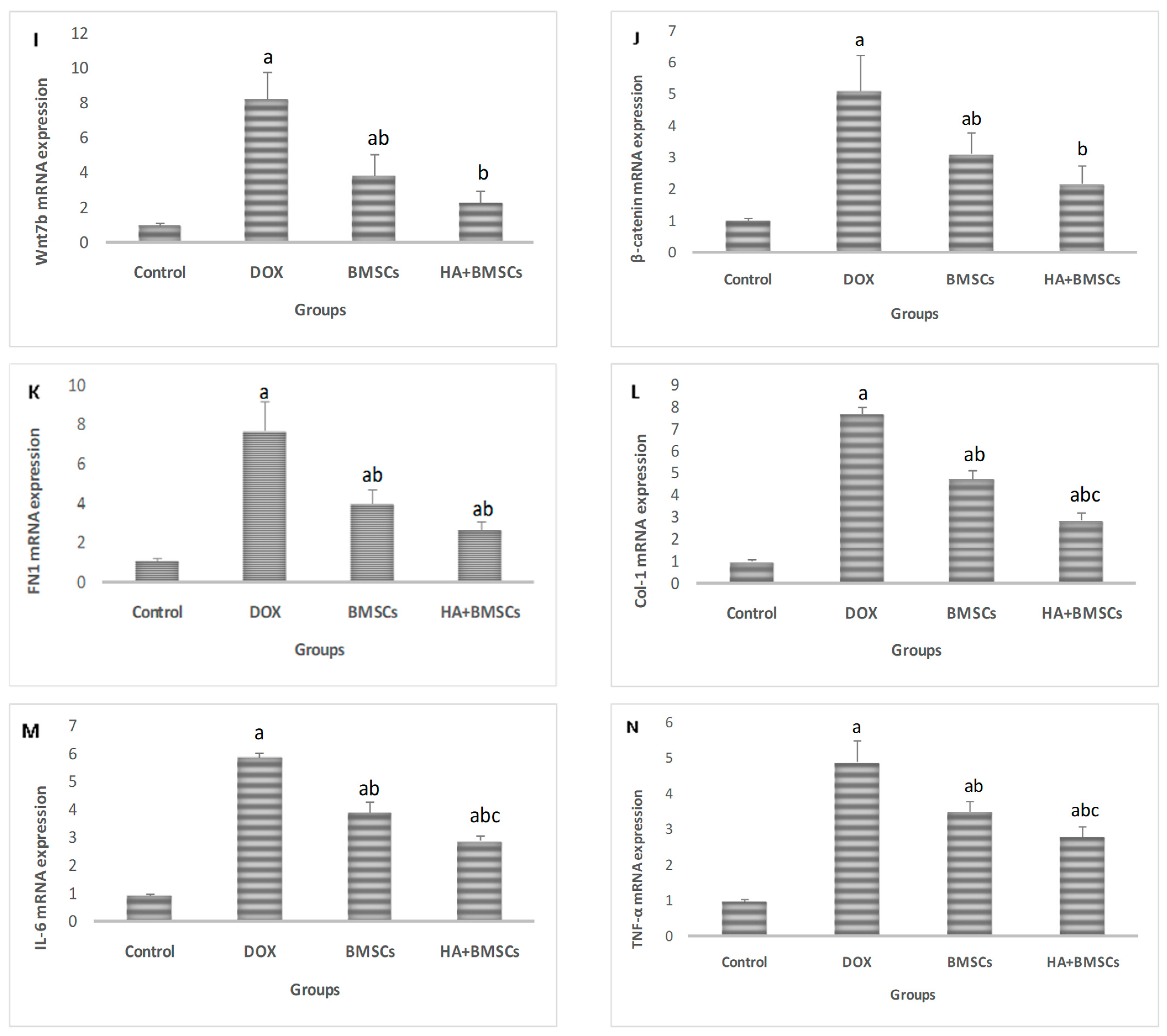
3.4. Effect of HA-Treated BMSCs on the Expression of Various Genes
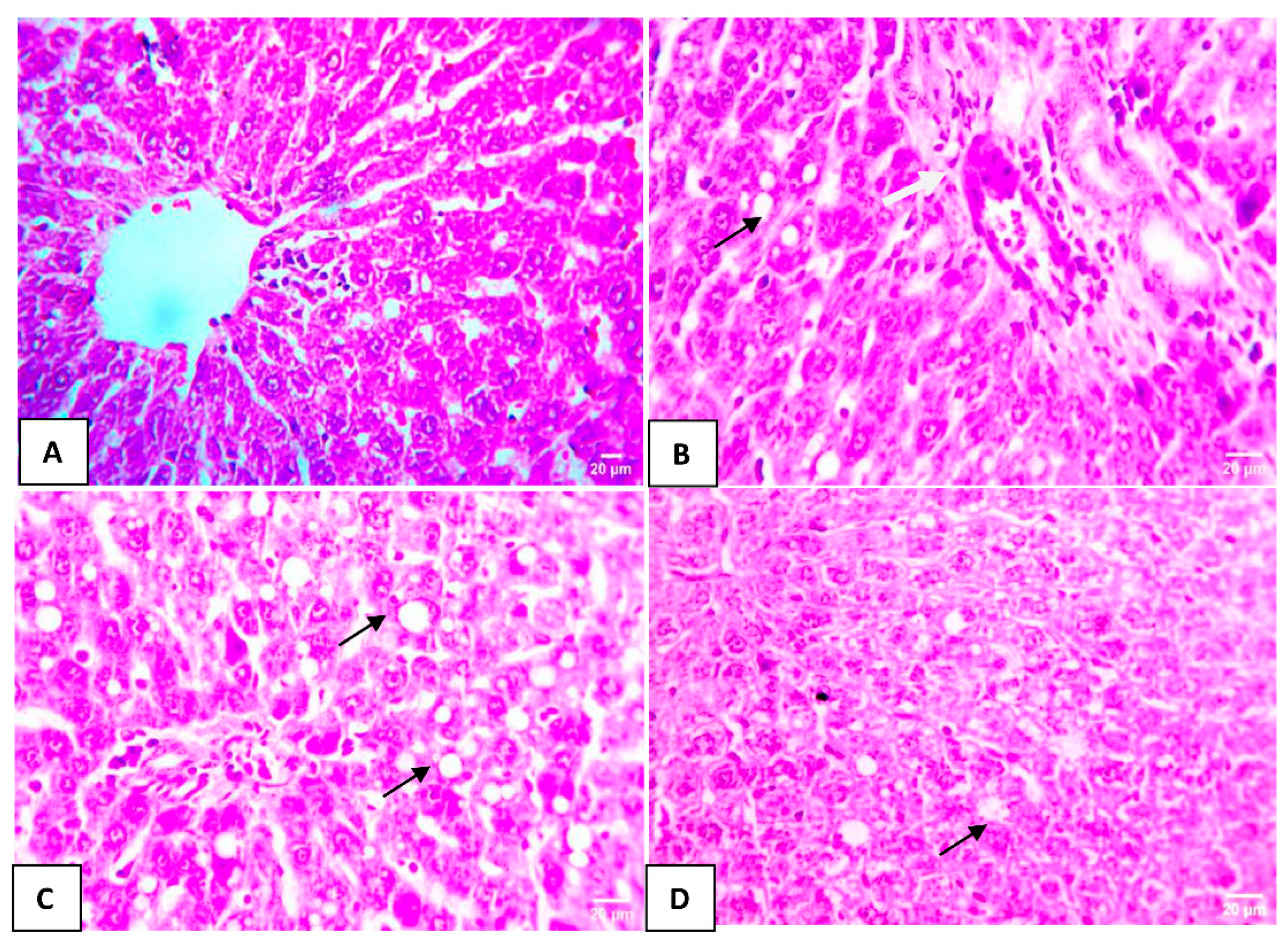
3.5. Effect of HA-Treated BMSCs on Histopathological Liver Changes
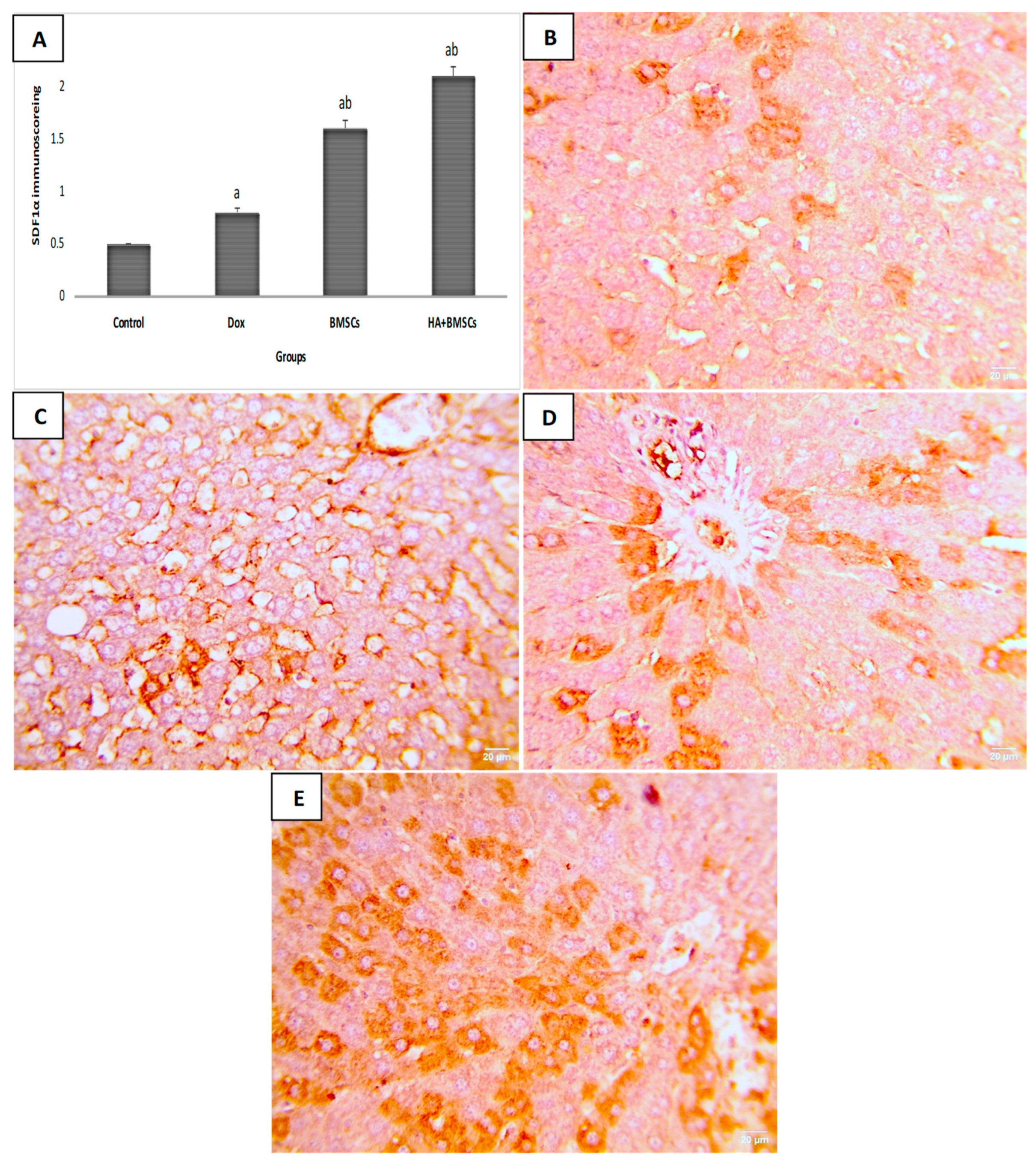
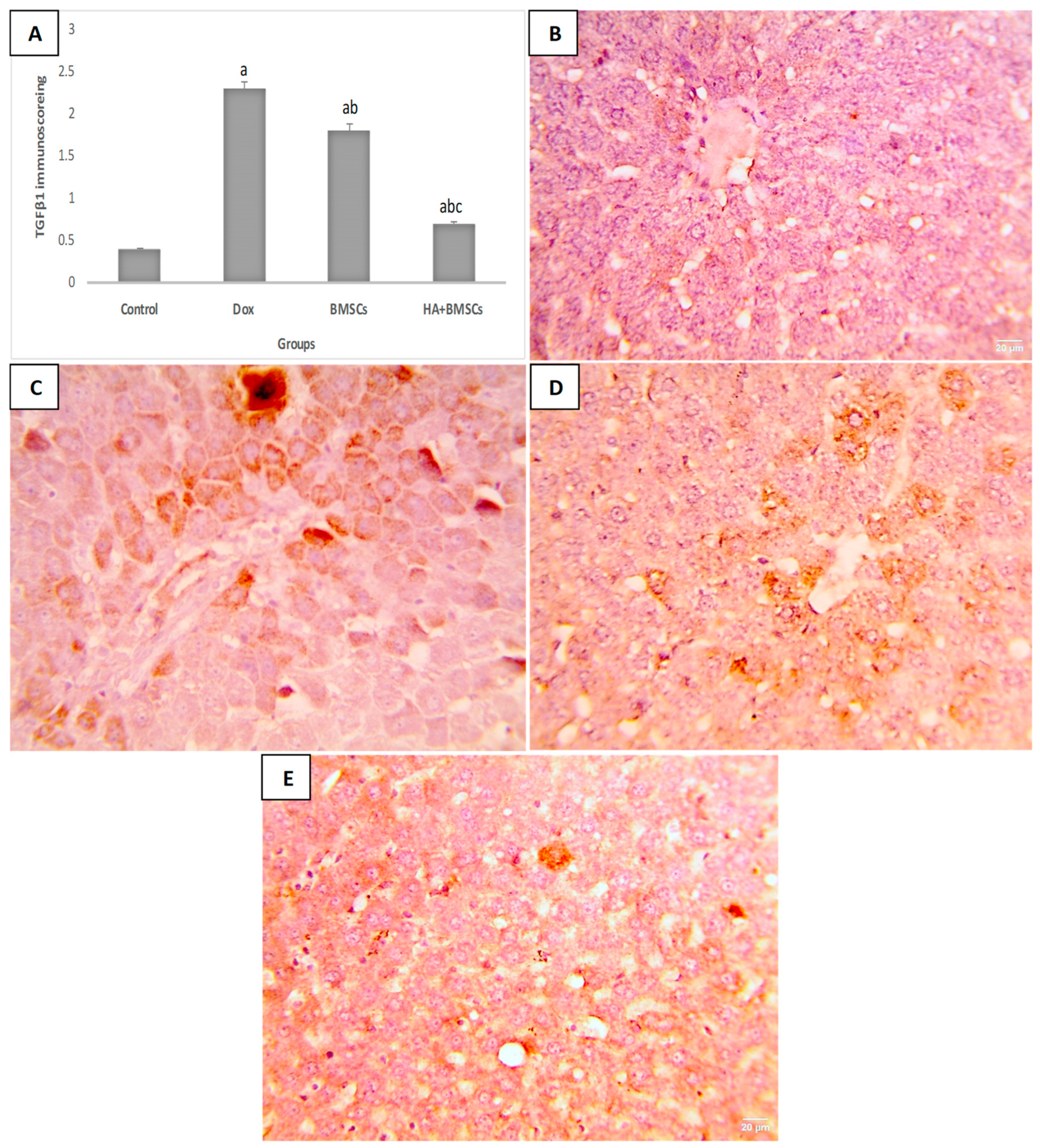
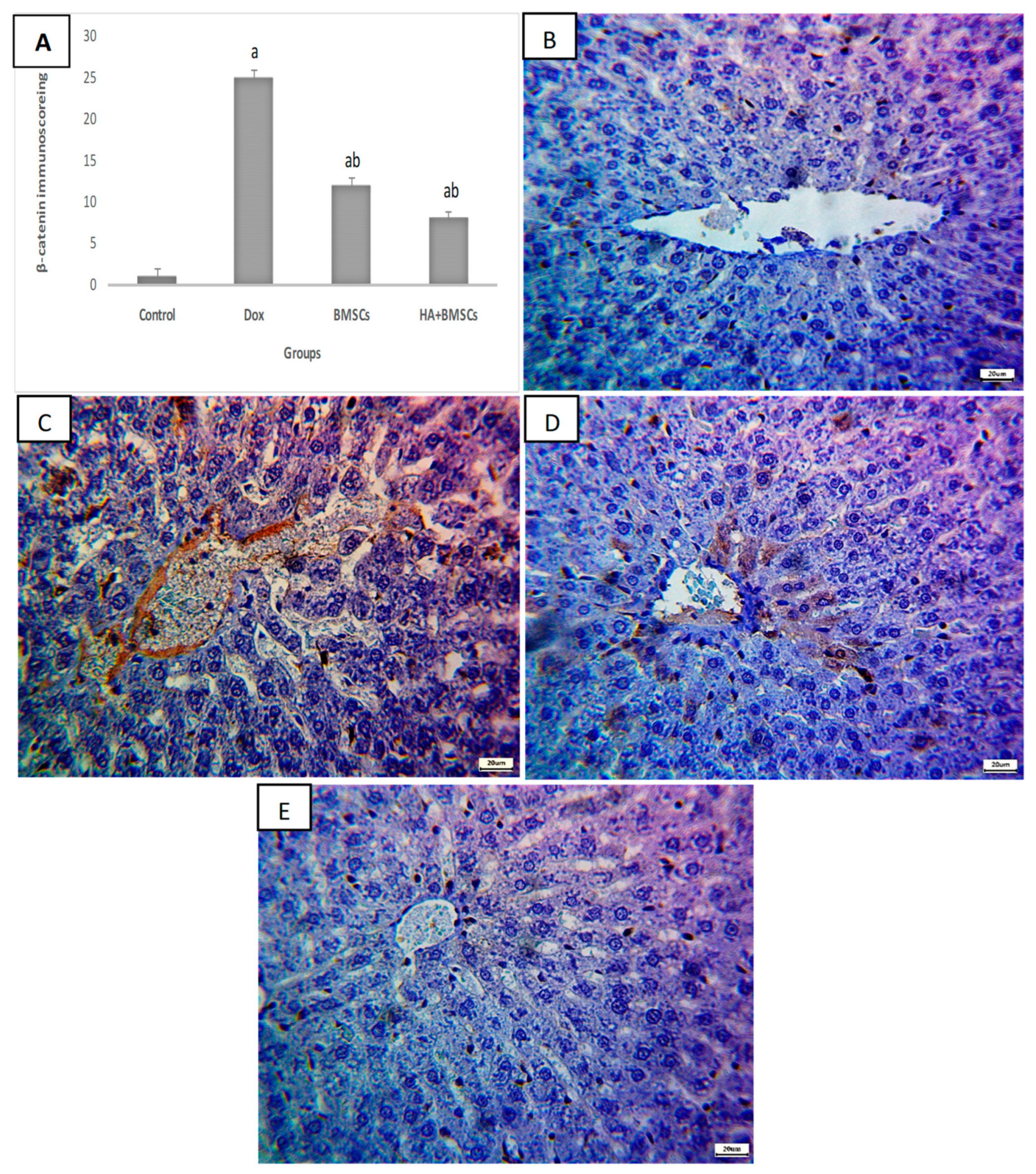
3.6. Effect of HA-Treated BMSCs on the Expression of TGF-β1, SDF1α, and β-Catenin
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA A Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Alqahtani, A.; Khan, Z.; Alloghbi, A.; Said Ahmed, T.S.; Ashraf, M.; Hammouda, D.M. Hepatocellular carcinoma: Molecular mechanisms and targeted therapies. Medicina 2019, 55, 526. [Google Scholar] [CrossRef] [PubMed]
- Mahmoudi, F.; Elyasi, S. None Herbal Agents as Preventive and Therapeutic Measures for Doxorubicin Induced Hepatotoxicity: A Comprehensive Review. J. Pharm. Care 2022, 10, 151–166. [Google Scholar] [CrossRef]
- Sohail, M.; Sun, Z.; Li, Y.; Gu, X.; Xu, H. Research progress in strategies to improve the efficacy and safety of doxorubicin for cancer chemotherapy. Expert Rev. Anticancer. Ther. 2021, 21, 1385–1398. [Google Scholar] [CrossRef]
- Yang, F.; Teves, S.S.; Kemp, C.J.; Henikoff, S. Doxorubicin, DNA torsion, and chromatin dynamics. Biochim. Biophys. Acta (BBA)-Rev. Cancer 2014, 1845, 84–89. [Google Scholar] [CrossRef]
- Shivakumar, P.; Rani, M.U.; Reddy, A.G.; Anjaneyulu, Y. A study on the toxic effects of doxorubicin on the histology of certain organs. Toxicol. Int. 2012, 19, 241. [Google Scholar] [PubMed]
- dos Santos Arruda, F.; Tomé, F.D.; Miguel, M.P.; de Menezes, L.B.; Nagib, P.R.; Campos, E.C.; Soave, D.F.; Celes, M.R.N. Doxorubicin-induced cardiotoxicity and cardioprotective agents: Classic and new players in the game. Curr. Pharm. Des. 2019, 25, 109–118. [Google Scholar] [CrossRef]
- Aljobaily, N.; Viereckl, M.J.; Hydock, D.S.; Aljobaily, H.; Wu, T.-Y.; Busekrus, R.; Jones, B.; Alberson, J.; Han, Y. Creatine alleviates doxorubicin-induced liver damage by inhibiting liver fibrosis, inflammation, oxidative stress, and cellular senescence. Nutrients 2020, 13, 41. [Google Scholar] [CrossRef]
- Mohajeri, M.; Sahebkar, A. Protective effects of curcumin against doxorubicin-induced toxicity and resistance: A review. Crit. Rev. Oncol./Hematol. 2018, 122, 30–51. [Google Scholar] [CrossRef] [PubMed]
- Perugorria, M.J.; Olaizola, P.; Labiano, I.; Esparza-Baquer, A.; Marzioni, M.; Marin, J.J.G.; Bujanda, L.; Banales, J.M. Wnt–β-catenin signalling in liver development, health and disease. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 121–136. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Xing, Y.; Jia, L.; Ji, Y.; Zhao, B.; Wen, Y.; Xu, X. An In Vitro Comparative Study of Multisource Derived Human Mesenchymal Stem Cells for Bone Tissue Engineering. Stem. Cells Dev. 2018, 27, 1634–1645. [Google Scholar] [CrossRef]
- Celik Samanci, T.; Gökcimen, A.; Kilic Eren, M.; Gürses, K.M.; Pilevneli, H.; Kuyucu, Y. Effects of bone marrow-derived mesenchymal stem cells on doxorubicin-induced liver injury in rats. J. Biochem. Mol. Toxicol. 2022, 36, e22985. [Google Scholar] [PubMed]
- Solis, M.A.; Chen, Y.H.; Wong, T.Y.; Bittencourt, V.Z.; Lin, Y.C.; Huang, L.L. Hyaluronan regulates cell behavior: A potential niche matrix for stem cells. Biochem. Res. Int. 2012, 2012, 346972. [Google Scholar] [CrossRef]
- Bian, X.H.; Zhou, G.Y.; Wang, L.N.; Ma, J.F.; Fan, Q.L.; Liu, N.; Bai, Y.; Guo, W.; Wang, Y.Q.; Sun, G.P.; et al. The role of CD44-hyaluronic acid interaction in exogenous mesenchymal stem cells homing to rat remnant kidney. Kidney Blood Press Res. 2013, 38, 11–20. [Google Scholar] [CrossRef]
- Sarhan, M.; El Serougy, H.; Hussein, A.M.; El-Dosoky, M.; Sobh, M.A.; Fouad, S.A.; Sobh, M.; Elhusseini, F. Impact of bone-marrow-derived mesenchymal stem cells on adriamycin-induced chronic nephropathy. Can. J. Physiol. Pharmacol. 2014, 92, 733–743. [Google Scholar] [CrossRef]
- Azzam, M.M.; Hussein, A.M.; Marghani, B.H.; Barakat, N.M.; Khedr, M.M.; Heakel, N.A. Retinoic Acid Potentiates the Therapeutic Efficiency of Bone Marrow-Derived Mesenchymal Stem Cells (BM-MSCs) against Cisplatin-Induced Hepatotoxicity in Rats. Sci. Pharm. 2022, 90, 58. [Google Scholar] [CrossRef]
- La Manna, G.; Bianchi, F.; Cappuccilli, M.; Cenacchi, G.; Tarantino, L.; Pasquinelli, G.; Valente, S.; Della Bella, E.; Cantoni, S.; Claudia, C.; et al. Mesenchymal stem cells in renal function recovery after acute kidney injury: Use of a differentiating agent in a rat model. Cell Transpl. 2011, 20, 1193–1208. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar]
- Feldman, A.T.; Wolfe, D. Tissue processing and hematoxylin and eosin staining. Histopathol. Methods Protoc. 2014, 1180, 31–43. [Google Scholar]
- Sharp, P.; La Regina, M.; Suckow, M. The Laboratory Rat; Boca Raton (Collection: The Laboratory Animal Pocket Reference Series); CRC Press: Boca Raton, FL, USA, 1998; 214p. [Google Scholar]
- Chen, X.; Zhang, Y.; Zhu, Z.; Liu, H.; Guo, H.; Xiong, C.; Xie, K.; Zhang, X.; Su, S. Protective effect of berberine on doxorubicin-induced acute hepatorenal toxicity in rats. Mol. Med. Rep. 2016, 13, 3953–3960. [Google Scholar] [CrossRef]
- AlAsmari, A.F.; Alharbi, M.; Alqahtani, F.; Alasmari, F.; AlSwayyed, M.; Alzarea, S.I.; Al-Alallah, I.A.; Alghamdi, A.; Hakami, H.M.; Alyousef, M.K.; et al. Diosmin Alleviates Doxorubicin-Induced Liver Injury via Modulation of Oxidative Stress-Mediated Hepatic Inflammation and Apoptosis via NfkB and MAPK Pathway: A Preclinical Study. Antioxidants 2021, 10, 1998. [Google Scholar] [CrossRef] [PubMed]
- Fu, Z.-J.; Li, X.-D.; Wei, D.-W.; Ding, X.-L. Clinical application potential of umbilical cord mesenchymal stem cells in chemotherapeutic ovarian failure. Reprod. Dev. Med. 2021, 5, 55–62. [Google Scholar] [CrossRef]
- Rühle, A.; Huber, P.E.; Saffrich, R.; Lopez Perez, R.; Nicolay, N.H. The current understanding of mesenchymal stem cells as potential attenuators of chemotherapy-induced toxicity. Int. J. Cancer 2018, 143, 2628–2639. [Google Scholar] [CrossRef] [PubMed]
- Rühle, A.; Lopez Perez, R.; Zou, B.; Grosu, A.-L.; Huber, P.E.; Nicolay, N.H. The therapeutic potential of mesenchymal stromal cells in the treatment of chemotherapy-induced tissue damage. Stem Cell Rev. Rep. 2019, 15, 356–373. [Google Scholar] [CrossRef]
- Abdelaziz, M.H.; Salah EL-Din, E.Y.; El-Dakdoky, M.H.; Ahmed, T.A. The impact of mesenchymal stem cells on doxorubicin-induced testicular toxicity and progeny outcome of male prepubertal rats. Birth Defects Res. 2019, 111, 906–919. [Google Scholar] [CrossRef] [PubMed]
- Hmadcha, A.; Martin-Montalvo, A.; Gauthier, B.R.; Soria, B.; Capilla-Gonzalez, V. Therapeutic potential of mesenchymal stem cells for cancer therapy. Front. Bioeng. Biotechnol. 2020, 8, 43. [Google Scholar] [CrossRef]
- Chu, D.-T.; Nguyen, T.T.; Tien, N.L.B.; Tran, D.-K.; Jeong, J.-H.; Anh, P.G.; Thanh, V.V.; Truong, D.T.; Dinh, T.C. Recent progress of stem cell therapy in cancer treatment: Molecular mechanisms and potential applications. Cells 2020, 9, 563. [Google Scholar] [CrossRef]
- Awadalla, A.; Hussein, A.M.; Ali, M.; Barakat, N.; Hamam, E.T.; Magar, R.W.; Shokeir, A.A. Possible mechanisms for the renoprotective action of adipose-derived mesenchymal stem cells with CD44-targeted hyaluronic acid against renal ischemia. Life Sci. 2021, 272, 119221. [Google Scholar] [CrossRef]
- Chanthick, C.; Thongboonkerd, V. Effects of Hyaluronic Acid on Calcium Oxalate Crystallization, Growth, Aggregation, Adhesion on Renal Tubular Cells, and Invasion Through Extracellular Matrix. Curr. Dev. Nutr. 2020, 4, 13. [Google Scholar] [CrossRef]
- Dzudzor, B.; Hammond, H.; Tachi, K.; Alisi, A.; Vento, S.; Gyasi, R.K.; Aheto, J.M.K. Serum 25-hydroxyvitamin D and hyaluronic acid levels as markers of fibrosis in patients with chronic liver disease at the main tertiary referral hospital in Ghana: A case-control study design. Health Sci. Rep. 2023, 6, e1101. [Google Scholar] [CrossRef]
- Luo, Y.; Liang, F.; Wan, X.; Liu, S.; Fu, L.; Mo, J.; Meng, X.; Mo, Z. Hyaluronic Acid Facilitates Angiogenesis of Endothelial Colony Forming Cell Combining With Mesenchymal Stem Cell via CD44/MicroRNA-139-5p Pathway. Front Bioeng. Biotechnol. 2022, 10, 794037. [Google Scholar] [CrossRef] [PubMed]
- Moreno, A.; Martínez, A.; Olmedillas, S.; Bello, S.; de Miguel, F. Hyaluronic acid effect on adipose-derived stem cells. Biological in vitro evaluation. Rev. Esp. Cir. Ortop. Traumatol. 2015, 59, 215–221. [Google Scholar] [CrossRef] [PubMed]
- Wali, A.F.; Rashid, S.; Rashid, S.M.; Ansari, M.A.; Khan, M.R.; Haq, N.; Alhareth, D.Y.; Ahmad, A.; Rehman, M.U. Naringenin Regulates Doxorubicin-Induced Liver Dysfunction: Impact on Oxidative Stress and Inflammation. Plants 2020, 9, 550. [Google Scholar] [CrossRef]
- Afsar, T.; Razak, S.; Almajwal, A. Effect of Acacia hydaspica R. Parker extract on lipid peroxidation, antioxidant status, liver function test and histopathology in doxorubicin treated rats. Lipids Health Dis. 2019, 18, 126. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Zhang, J.; Tong, N.; Chen, Y.; Luo, Y. Protective effects of berberine on doxorubicin-induced hepatotoxicity in mice. Biol. Pharm. Bull 2012, 35, 796–800. [Google Scholar] [CrossRef] [PubMed]
- Henninger, C.; Huelsenbeck, J.; Huelsenbeck, S.; Grösch, S.; Schad, A.; Lackner, K.J.; Kaina, B.; Fritz, G. The lipid lowering drug lovastatin protects against doxorubicin-induced hepatotoxicity. Toxicol. Appl. Pharmacol. 2012, 261, 66–73. [Google Scholar] [CrossRef]
- Shokeir, A.A.; Hussein, A.M.; Barakat, N.; Abdelaziz, A.; Elgarba, M.; Awadalla, A. Activation of nuclear factor erythroid 2-related factor 2 (Nrf2) and Nrf-2-dependent genes by ischaemic pre-conditioning and post-conditioning: New adaptive endogenous protective responses against renal ischaemia/reperfusion injury. Acta Physiol. 2014, 210, 342–353. [Google Scholar] [CrossRef]
- Hussein, A.M.; Barakat, N.; Awadalla, A.; Gabr, M.M.; Khater, S.; Harraz, A.M.; Shokeir, A.A. Modulation of renal ischemia/reperfusion in rats by a combination of ischemic preconditioning and adipose-derived mesenchymal stem cells (ADMSCs). Can. J. Physiol. Pharmacol. 2016, 94, 936–946. [Google Scholar] [CrossRef]
- Zahran, M.H.; Hussein, A.M.; Barakat, N.; Awadalla, A.; Khater, S.; Harraz, A.; Shokeir, A.A. Sildenafil activates antioxidant and antiapoptotic genes and inhibits proinflammatory cytokine genes in a rat model of renal ischemia/reperfusion injury. Int. Urol. Nephrol. 2015, 47, 1907–1915. [Google Scholar] [CrossRef]
- Dekel, B.; Shezen, E.; Even-Tov-Friedman, S.; Katchman, H.; Margalit, R.; Nagler, A.; Reisner, Y. Transplantation of human hematopoietic stem cells into ischemic and growing kidneys suggests a role in vasculogenesis but not tubulogenesis. Stem. Cells 2006, 24, 1185–1193. [Google Scholar] [CrossRef]
- da Silva, L.B.L.; Palma, P.V.B.; Cury, P.M.; Bueno, V. Evaluation of stem cell administration in a model of kidney ischemia-reperfusion injury. Int. Immunopharmacol. 2007, 7, 1609–1616. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Cohen, A.; Hudson, T.E.; Motlagh, D.; Amrani, D.L.; Duffield, J.S. Mobilized human hematopoietic stem/progenitor cells promote kidney repair following ischemia reperfusion injury. Circulation 2010, 121, 2211. [Google Scholar] [CrossRef]
- Du, T.; Cheng, J.; Zhong, L.; Zhao, X.-F.; Zhu, J.; Zhu, Y.-J.; Liu, G.-H. The alleviation of acute and chronic kidney injury by human Wharton’s jelly-derived mesenchymal stromal cells triggered by ischemia-reperfusion injury via an endocrine mechanism. Cytotherapy 2012, 14, 1215–1227. [Google Scholar] [CrossRef]
- Du, T.; Ju, G.; Wu, S.; Cheng, Z.; Cheng, J.; Zou, X.; Zhang, G.; Miao, S.; Liu, G.; Zhu, Y. Microvesicles derived from human Wharton’s jelly mesenchymal stem cells promote human renal cancer cell growth and aggressiveness through induction of hepatocyte growth factor. PLoS ONE 2014, 9, e96836. [Google Scholar] [CrossRef] [PubMed]
- Du, T.; Zou, X.; Cheng, J.; Wu, S.; Zhong, L.; Ju, G.; Zhu, J.; Liu, G.; Zhu, Y.; Xia, S. Human Wharton’s jelly-derived mesenchymal stromal cells reduce renal fibrosis through induction of native and foreign hepatocyte growth factor synthesis in injured tubular epithelial cells. Stem Cell Res. Ther. 2013, 4, 59. [Google Scholar] [CrossRef] [PubMed]
- Cóndor, J.M.; Rodrigues, C.E.; de Sousa Moreira, R.; Canale, D.; Volpini, R.A.; Shimizu, M.H.; Camara, N.O.; Noronha, I.d.L.; Andrade, L. Treatment with human Wharton’s jelly-derived mesenchymal stem cells attenuates sepsis-induced kidney injury, liver injury, and endothelial dysfunction. Stem Cells Transl. Med. 2016, 5, 1048–1057. [Google Scholar] [CrossRef] [PubMed]
- Ventura, C.; Cantoni, S.; Bianchi, F.; Lionetti, V.; Cavallini, C.; Scarlata, I.; Foroni, L.; Maioli, M.; Bonsi, L.; Alviano, F. Hyaluronan mixed esters of butyric and retinoic acid drive cardiac and endothelial fate in term placenta human mesenchymal stem cells and enhance cardiac repair in infarcted rat hearts. J. Biol. Chem. 2007, 282, 14243–14252. [Google Scholar] [CrossRef]
- Belmonte, F.; Das, S.; Sysa-Shah, P.; Sivakumaran, V.; Stanley, B.; Guo, X.; Paolocci, N.; Aon, M.A.; Nagane, M.; Kuppusamy, P.; et al. ErbB2 overexpression upregulates antioxidant enzymes, reduces basal levels of reactive oxygen species, and protects against doxorubicin cardiotoxicity. Am. J. Physiol. Heart Circ. Physiol. 2015, 309, H1271–H1280. [Google Scholar] [CrossRef]
- Jiao, W.; Mi, X.; Yang, Y.; Liu, R.; Liu, Q.; Yan, T.; Chen, Z.-J.; Qin, Y.; Zhao, S. Mesenchymal stem cells combined with autocrosslinked hyaluronic acid improve mouse ovarian function by activating the PI3K-AKT pathway in a paracrine manner. Stem Cell Res. Ther. 2022, 13, 1–17. [Google Scholar] [CrossRef]
- Serag, W.M.; Barakat, N.; Elshehabi, M.E.; Hafez, H.S.; Zahran, F. Renoprotective effect of bone marrow mesenchymal stem cells with hyaluronic acid against adriamycin- induced kidney fibrosis via inhibition of Wnt/β-catenin pathway. International J. Biol. Macromol. 2022, 207, 741–749. [Google Scholar] [CrossRef]
- Bahrehbar, K.; Rezazadeh Valojerdi, M.; Esfandiari, F.; Fathi, R.; Hassani, S.N.; Baharvand, H. Human embryonic stem cell-derived mesenchymal stem cells improved premature ovarian failure. World J. Stem Cells 2020, 12, 857–878. [Google Scholar] [CrossRef]
- Wu, H.; Liu, S.; Gong, J.; Liu, J.; Zhang, Q.; Leng, X.; Zhang, N.; Li, Y. VCPA, a novel synthetic derivative of α-tocopheryl succinate, sensitizes human gastric cancer to doxorubicin-induced apoptosis via ROS-dependent mitochondrial dysfunction. Cancer Lett. 2017, 393, 22–32. [Google Scholar] [CrossRef]
- Fabregat, I.; Moreno-Càceres, J.; Sánchez, A.; Dooley, S.; Dewidar, B.; Giannelli, G.; Ten Dijke, P.; Consortium, I.L. TGF-β signalling and liver disease. FEBS J. 2016, 283, 2219–2232. [Google Scholar] [CrossRef] [PubMed]
- Haque, N.; Fareez, I.M.; Fong, L.F.; Mandal, C.; Kasim, N.H.A.; Kacharaju, K.R.; Soesilawati, P. Role of the CXCR4-SDF1-HMGB1 pathway in the directional migration of cells and regeneration of affected organs. World J. Stem Cells 2020, 12, 938. [Google Scholar] [CrossRef] [PubMed]
- El Nashar, E.M.; Alghamdi, M.A.; Alasmari, W.A.; Hussein, M.M.; Hamza, E.; Taha, R.I.; Ahmed, M.M.; Al-Khater, K.M.; Abdelfattah-Hassan, A. Autophagy promotes the survival of adipose mesenchymal stem/stromal cells and enhances their therapeutic effects in cisplatin-induced liver injury via modulating TGF-β1/Smad and PI3K/AKT signaling pathways. Cells 2021, 10, 2475. [Google Scholar] [CrossRef]
- Akcora, B.Ö.; Storm, G.; Bansal, R. Inhibition of canonical WNT signaling pathway by β-catenin/CBP inhibitor ICG-001 ameliorates liver fibrosis in vivo through suppression of stromal CXCL12. Biochim. Et Biophys. Acta (BBA)-Mol. Basis Dis. 2018, 1864, 804–818. [Google Scholar] [CrossRef]
- Du, Z.; Wei, C.; Yan, J.; Han, B.; Zhang, M.; Peng, C.; Liu, Y. Mesenchymal stem cells overexpressing C-X-C chemokine receptor type 4 improve early liver regeneration of small-for-size liver grafts. Liver Transplant. 2013, 19, 215–225. [Google Scholar] [CrossRef] [PubMed]
- Peifer, M.; Polakis, P. Wnt signaling in oncogenesis and embryogenesis—A look outside the nucleus. Science 2000, 287, 1606–1609. [Google Scholar] [CrossRef]
- Lancaster, M.A.; Louie, C.M.; Silhavy, J.L.; Sintasath, L.; DeCambre, M.; Nigam, S.K.; Willert, K.; Gleeson, J.G. Impaired Wnt–β-catenin signaling disrupts adult renal homeostasis and leads to cystic kidney ciliopathy. Nat. Med. 2009, 15, 1046–1054. [Google Scholar] [CrossRef]
- Angers, S.; Moon, R.T. Proximal events in Wnt signal transduction. Nat. Rev. Mol. Cell Biol. 2009, 10, 468–477. [Google Scholar] [CrossRef]
- Rong, X.; Liu, J.; Yao, X.; Jiang, T.; Wang, Y.; Xie, F. Human bone marrow mesenchymal stem cells-derived exosomes alleviate liver fibrosis through the Wnt/β-catenin pathway. Stem Cell Res. Ther. 2019, 10, 98. [Google Scholar] [CrossRef] [PubMed]
- Anavi, S.; Eisenberg-Bord, M.; Hahn-Obercyger, M.; Genin, O.; Pines, M.; Tirosh, O. The role of iNOS in cholesterol-induced liver fibrosis. Lab. Investig. 2015, 95, 914–924. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.X.; Zeng, Z.C.; Sun, J.; Zeng, H.Y.; Huang, Y.; Zhang, Z.Y. Mesenchymal stem cell-conditioned medium prevents radiation-induced liver injury by inhibiting inflammation and protecting sinusoidal endothelial cells. J. Radiat. Res. 2015, 56, 700–708. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.E.; Lee, J.Y.; Shim, K.S.; Lee, S.; Min, K.; Bae, J.H.; Kim, H.J.; Park, K.; Song, H.R. Attenuation of inflammation and cartilage degradation by sulfasalazine-containing hyaluronic acid on osteoarthritis rat model. Int. J. Biol. Macromol. 2018, 114, 341–348. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Common Name | Sequence (5′-3′) | Accession No. |
|---|---|---|
| Wnt7b | F: 5′-CCATCATCGTGATCGGGGAG-3′ R: 5′-TCCAGTTTCATGCGGTCCTC-3′ | NM_001009695.1 |
| iNOS | F: 5′-AGTCAACTACAAGCCCCACG-3′ R- 5′-GCAGCTTGTCCAGGGATTCT-3′ | NM_012611.3 |
| BCL2 | F: 5′-GGTGAACTGGGGGAGGATTG-3′ R: 5′-GCATGCTGGGGCCATATAGT-3′ | NM_016993.1 |
| VEGF | F: 5′-ACGAAAGCGCAAGAAATCCC-3′ R: 5′-TTAACTCAAGCTGCCTCGCC-3′ | NM_031836.3 |
| Bax | F: 5′-GGCGATGAACTGGACAACAA-3′ R: 5′-CAAAGTAGAAAAGGGCAACC-3′ | NM_017059.2 |
| SDF1a | F: 5′-GAGCCATGTCGCCAGAGCCAAC R: 5′-CACACCTCTCACATCTTGAGCCTCT | NM_001033882.1 |
| Collagen-1 | F: 5′-ACGTCCTGGTGAAGTTGGTC-3′ R: 5′-CAGGGAAGCCTCTTTCTCCT-3′ | NM_ 29,393 |
| TGF-β1 | F: 5′-CACTCCCGTGGCTTCTAGTG-3′ R: 5′-GGAC TGGCGAGCCTTAGTTT-3′ | NM_021578.2 |
| β-catenin | F: 5′-ACAGCACCTTCAGCACTCT-3′ R: 5′-AAGTTCTTGGCTATTACGACA-3′ | NM_053357.2 |
| Nrf2 | F: 5′-ATTGCTGTCCATCTCTGTCAG-3′ R: 5′-GCTATTTTCCATTCCCGAGTTAC-3′ | NM_001399173.1 |
| HO-1 | F: 5′-TGCTTGTTTCGCTCTATCTCC-3′ R: 5′-CTTTCAGAAGGGTCAGGTGTC-3′ | NM_012580.2 |
| Fibronectin | F: 5′-GTGGCTGCCTTCAACTTCTC-3′ R: 5′-GTGGGTTGCAAACCTTCAAT-3 | NM_U82612.1 |
| CD44 | F: 5′-TGGCACAGCAGCAGATC-3′ R: 5′-CTGCACAGATAGCGTTGG-3′ | NM_012924.3 |
| IL_6 | F: 5′-CGAGCCCACCAGGAACGAAAGTC-3′ R: 5′-CTGGCTGGAAGTCTCTTGCGGAG-3′ | NM_012589.2 |
| TNF-α | F: 5′-TTC GGA ACT CAC TGG ATC CC-3′ R: 5′-CGGA ACA GTC TGG GAA GCT CT-3′ | NM_012675.3 |
| GAPDH | F: 5′-AGACAGCCGCATCTTCTTGT-3′ R: 5′-TTCCCATTCTCAGCCTTGAC-3′ | NM_017008.4 |
| Control | DOX | BMSCs | BMSCs + HA | |
|---|---|---|---|---|
| ALT (U/L) | 49 ± 3.4 | 137.83 ± 6.43 a | 98.67 ± 10.24 ab | 77.5 ± 15.50 abc |
| AST (U/L) | 104.16 ± 15.7 | 261.16 ± 52.02 a | 149.66 ± 49.2 b | 117.33 ± 20.64 b |
| Alb (g/dL) | 3.51 ± 0.55 | 1.98 ± 0.25 a | 2.48 ± 0.68 a | 3.12 ± 0.41 b |
| CAT (U/gm) | 8.1 ± 1.96 | 1.6 ± 0.42 a | 5.25 ± 0.82 ab | 6.4 ± 1.4 b |
| GSH (U/gm) | 10.21 ± 3.28 | 3.16 ± 0.95 a | 7.5 ± 1.4 b | 9.2 ± 1.59 b |
| SOD (U/gm) | 289.41 ± 37.1 | 115.26 ± 32.53 a | 220.27 ± 41.9 ab | 273.48 ± 23.05 b |
| MDA (nmol/gm) | 77.38 ± 10.89 | 168.44 ± 15.77 a | 113.37 ± 14.43 ab | 88.83 ± 14.47 bc |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Awadalla, A.; Hamam, E.T.; Mostafa, S.A.; Mahmoud, S.A.; Elazab, K.M.; El Nakib, A.M.; Eldesoqui, M.; El-Sherbiny, M.; Ammar, O.A.; Al-Serwi, R.H.; et al. Hepatoprotective Effects of Hyaluronic Acid-Preconditioned Bone Marrow Mesenchymal Stem Cells against Liver Toxicity via the Inhibition of Apoptosis and the Wnt/β-Catenin Signaling Pathway. Cells 2023, 12, 1526. https://doi.org/10.3390/cells12111526
Awadalla A, Hamam ET, Mostafa SA, Mahmoud SA, Elazab KM, El Nakib AM, Eldesoqui M, El-Sherbiny M, Ammar OA, Al-Serwi RH, et al. Hepatoprotective Effects of Hyaluronic Acid-Preconditioned Bone Marrow Mesenchymal Stem Cells against Liver Toxicity via the Inhibition of Apoptosis and the Wnt/β-Catenin Signaling Pathway. Cells. 2023; 12(11):1526. https://doi.org/10.3390/cells12111526
Chicago/Turabian StyleAwadalla, Amira, Eman T. Hamam, Sally Abdallah Mostafa, Seham Ahmed Mahmoud, Khalid Mohamed Elazab, Ahmed Mohamed El Nakib, Mamdouh Eldesoqui, Mohamed El-Sherbiny, Omar A. Ammar, Rasha Hamed Al-Serwi, and et al. 2023. "Hepatoprotective Effects of Hyaluronic Acid-Preconditioned Bone Marrow Mesenchymal Stem Cells against Liver Toxicity via the Inhibition of Apoptosis and the Wnt/β-Catenin Signaling Pathway" Cells 12, no. 11: 1526. https://doi.org/10.3390/cells12111526