
The Human Placental Amniotic Membrane Mesenchymal-Stromal-Cell-Derived Conditioned Medium Inhibits Growth and Promotes Apoptosis of Human Cholangiocarcinoma Cells In Vitro and In Vivo by Suppressing IL-6/JAK2/STAT3 Signaling
 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
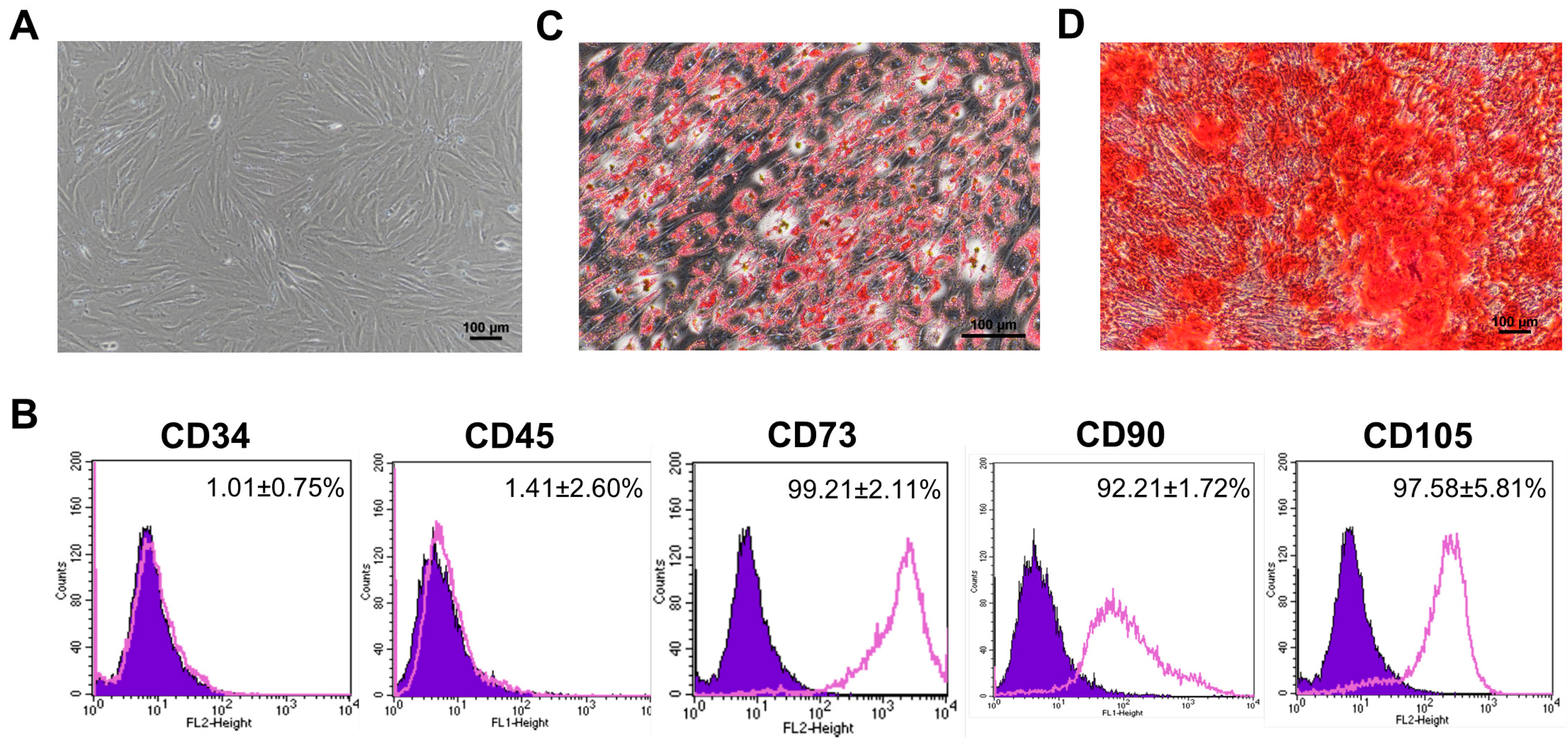
2.1. Isolation and Characterization of hPAMSCs
2.2. Preparation of the Conditioned Medium
2.3. Cell Lines and Culture
2.4. MTT Assay
2.5. Colony Formation Assay
2.6. Flow Cytometry Analysis of Apoptosis
2.7. Mitochondrial Membrane Potential (∆Ψm) Assay
2.8. Caspase 3 Activity Assay
2.9. Western Blot Analysis
2.10. In Vivo Assay
2.11. Immunohistochemical Staining (IHC)
2.12. Statistical Analysis
3. Results
3.1. Characterization of Human Placental Amniotic Membrane Mesenchymal Stromal Cells (hPAMSCs)
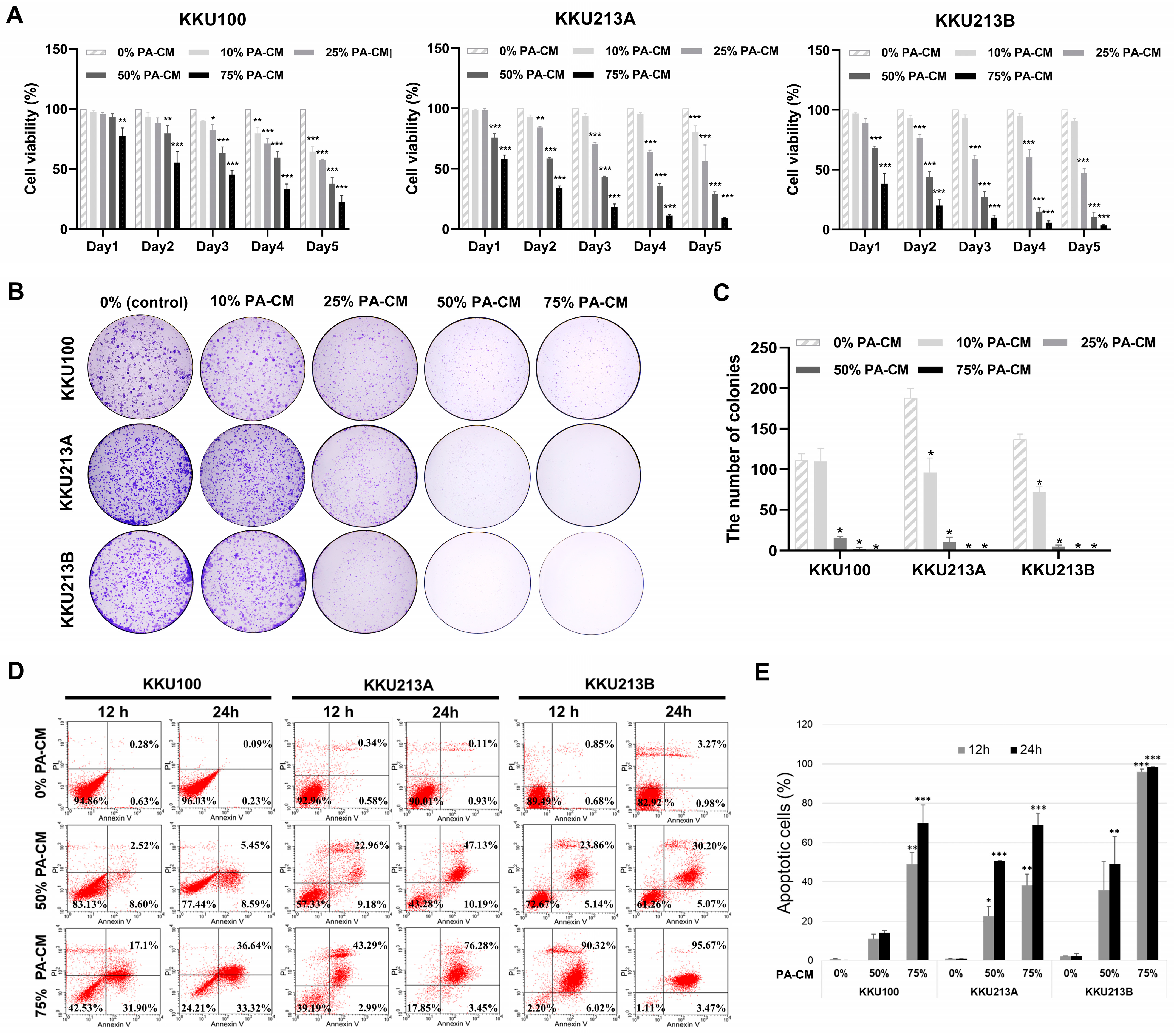
3.2. PA-CM Suppresses CCA Cell Growth
3.3. PA-CM Promotes CCA Cell Apoptosis
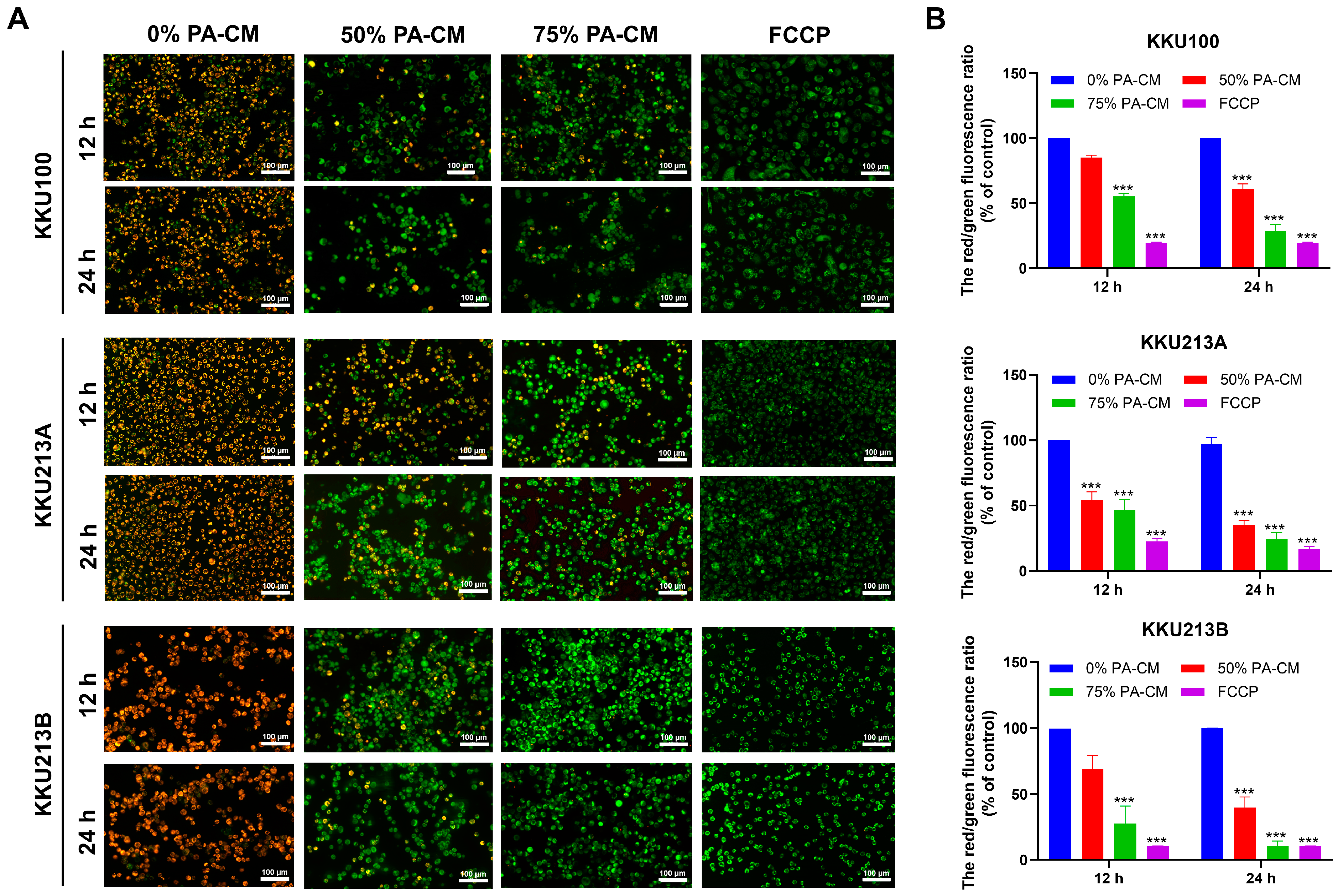
3.4. PA-CM Induces the Loss of Mitochondrial Membrane Potential in CCA Cells
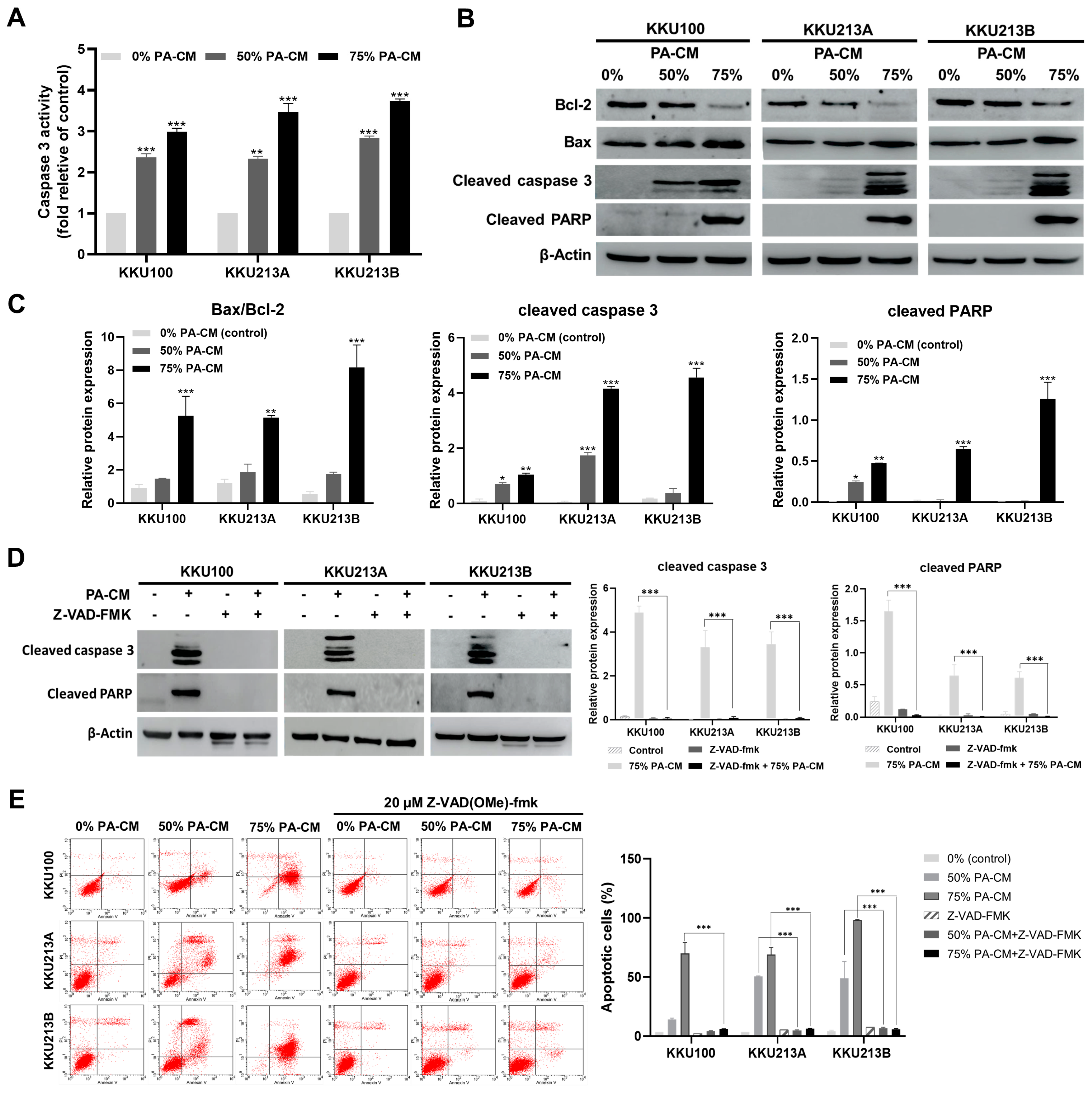
3.5. PA-CM Induces CCA Cell Apoptosis through the Mitochondrial-Mediated Caspase Pathway
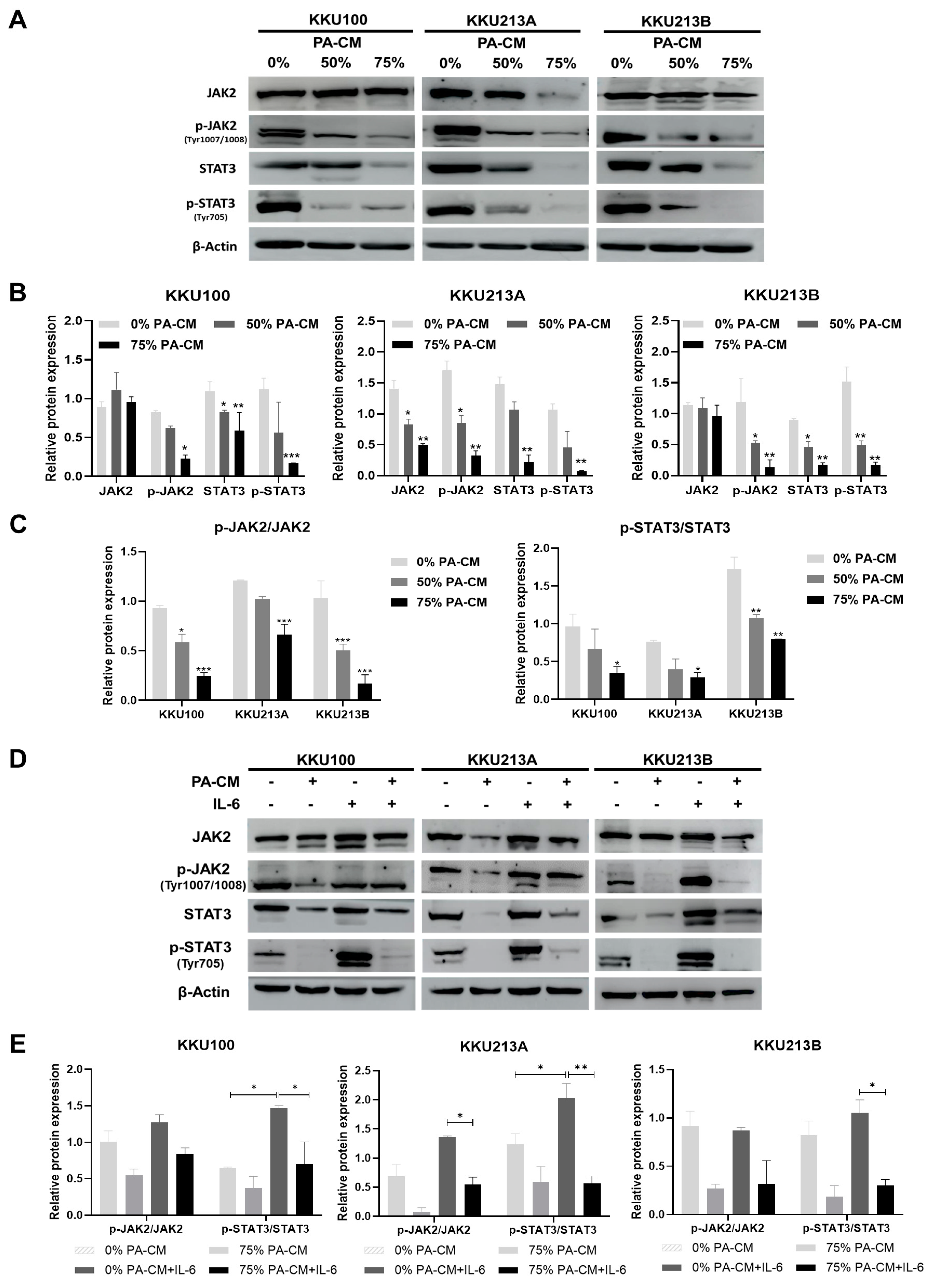
3.6. PA-CM Suppresses the IL-6-Mediated JAK2/STAT3 Signaling Pathway in CCA Cells
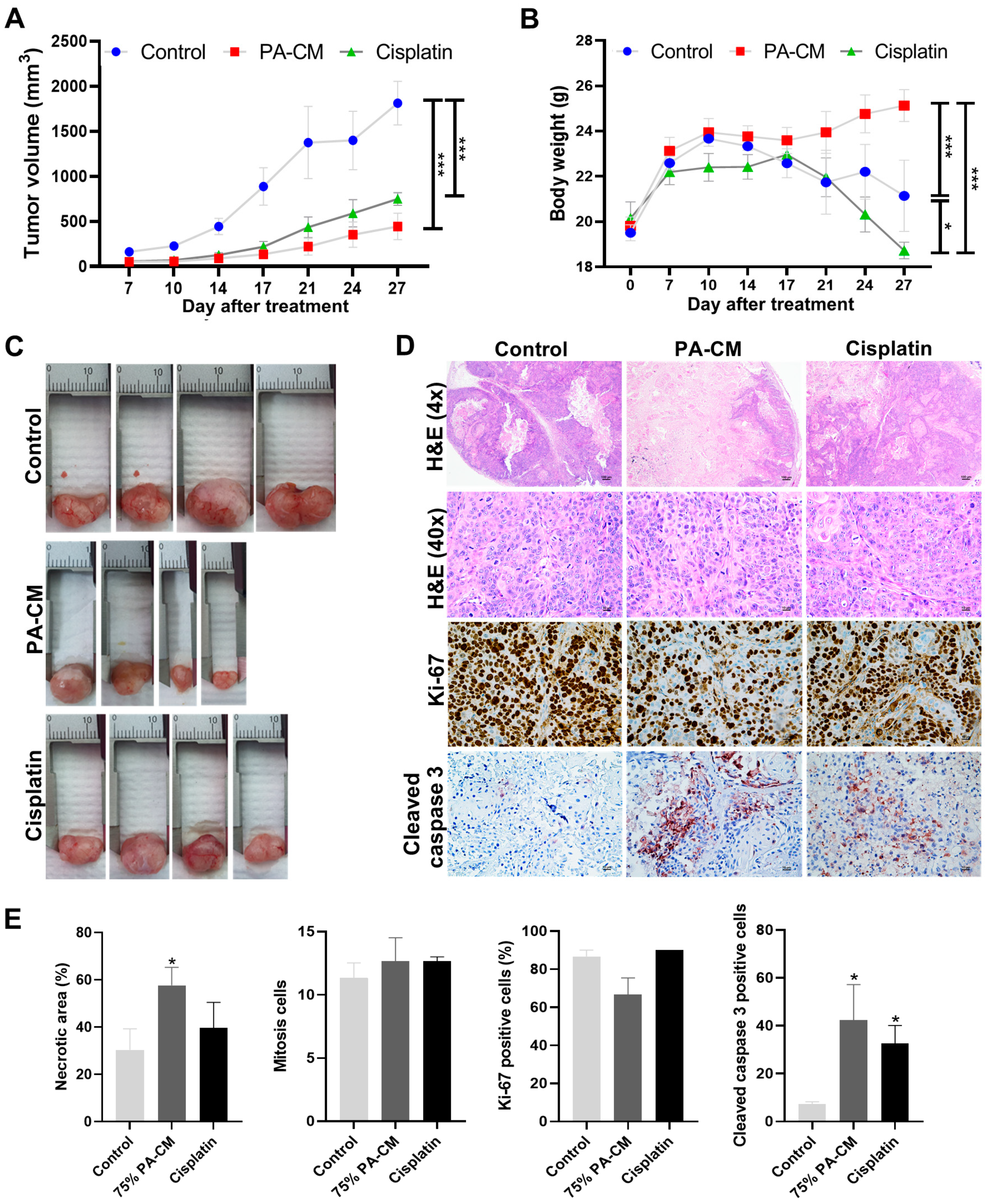
3.7. PA-CM Suppresses CCA Cell Growth In Vivo by Inducing Apoptosis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nakanuma, Y.; Sato, Y.; Harada, K.; Sasaki, M.; Xu, J.; Ikeda, H. Pathological classification of intrahepatic cholangiocarcinoma based on a new concept. World J. Hepatol. 2010, 2, 419–427. [Google Scholar] [CrossRef] [PubMed]
- Florio, A.A.; Ferlay, J.; Znaor, A.; Ruggieri, D.; Alvarez, C.S.; Laversanne, M.; Bray, F.; McGlynn, K.A.; Petrick, J.L. Global trends in intrahepatic and extrahepatic cholangiocarcinoma incidence from 1993 to 2012. Cancer 2020, 126, 2666–2678. [Google Scholar] [CrossRef] [PubMed]
- Forner, A.; Vidili, G.; Rengo, M.; Bujanda, L.; Ponz-Sarvisé, M.; Lamarca, A. Clinical presentation, diagnosis and staging of cholangiocarcinoma. Liver Int. 2019, 39, 98–107. [Google Scholar] [CrossRef] [PubMed]
- Rizvi, S.; Khan, S.A.; Hallemeier, C.L.; Kelley, R.K.; Gores, G.J. Cholangiocarcinoma—Evolving concepts and therapeutic strategies. Nat. Rev. Clin. Oncol. 2018, 15, 95. [Google Scholar] [CrossRef] [PubMed]
- DeOliveira, M.L.; Cunningham, S.C.; Cameron, J.L.; Kamangar, F.; Winter, J.M.; Lillemoe, K.D.; Choti, M.A.; Yeo, C.J.; Schulick, R.D. Cholangiocarcinoma: Thirty-one-year experience with 564 patients at a single institution. Ann. Surg. 2007, 245, 755. [Google Scholar] [CrossRef] [PubMed]
- Reger, R.L.; Tucker, A.H.; Wolfe, M.R. Differentiation and characterization of human MSCs. Methods Mol. Biol. 2008, 449, 93–107. [Google Scholar] [PubMed]
- Whiteside, T.L. Exosome and mesenchymal stem cell cross-talk in the tumor microenvironment. Semin. Immunol. 2018, 35, 69–79. [Google Scholar] [CrossRef]
- Hsu, H.-S.; Lin, J.-H.; Hsu, T.-W.; Su, K.; Wang, C.-W.; Yang, K.-Y.; Chiou, S.-H.; Hung, S.-C. Mesenchymal stem cells enhance lung cancer initiation through activation of IL-6/JAK2/STAT3 pathway. Lung Cancer 2012, 75, 167–177. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Hu, F.; Li, G.; Li, G.; Yang, X.; Liu, L.; Zhang, R.; Zhang, B.; Feng, Y. Human colorectal cancer-derived mesenchymal stem cells promote colorectal cancer progression through IL-6/JAK2/STAT3 signaling. Cell Death Dis. 2018, 9, 25. [Google Scholar] [CrossRef]
- Wang, M.; Zhang, Y.; Liu, M.; Jia, Y.; He, J.; Xu, X.; Shi, H.; Zhang, Y.; Zhang, J.; Liu, Y. Inhibition of STAT3 signaling as critical molecular event in HUC-MSCs suppressed Glioblastoma Cells. J. Cancer 2023, 14, 611. [Google Scholar] [CrossRef]
- Szyposzynska, A.; Bielawska-Pohl, A.; Krawczenko, A.; Doszyn, O.; Paprocka, M.; Klimczak, A. Suppression of Ovarian Cancer Cell Growth by AT-MSC Microvesicles. Int. J. Mol. Sci. 2020, 21, 9143. [Google Scholar] [CrossRef]
- Abd-Allah, S.H.; Shalaby, S.M.; El-Shal, A.S.; Elkader, E.A.; Hussein, S.; Emam, E.; Mazen, N.F.; El Kateb, M.; Atfy, M. Effect of bone marrow-derived mesenchymal stromal cells on hepatoma. Cytotherapy 2014, 16, 1197–1206. [Google Scholar] [CrossRef] [PubMed]
- He, N.; Kong, Y.; Lei, X.; Liu, Y.; Wang, J.; Xu, C.; Wang, Y.; Du, L.; Ji, K.; Wang, Q. MSCs inhibit tumor progression and enhance radiosensitivity of breast cancer cells by down-regulating Stat3 signaling pathway. Cell Death Dis. 2018, 9, 1026. [Google Scholar] [CrossRef] [PubMed]
- Zuk, P.A.; Zhu, M.; Ashjian, P.; De Ugarte, D.A.; Huang, J.I.; Mizuno, H.; Alfonso, Z.C.; Fraser, J.K.; Benhaim, P.; Hedrick, M.H. Human adipose tissue is a source of multipotent stem cells. Mol. Biol. Cell. 2002, 13, 4279–4295. [Google Scholar] [CrossRef] [PubMed]
- Maghen, L.; Shlush, E.; Gat, I.; Filice, M.; Barretto, T.; Jarvi, K.; Lo, K.; Gauthier-Fisher, A.S.; Librach, C.L. Human umbilical perivascular cells: A novel source of MSCs to support testicular niche regeneration. Reproduction 2017, 153, 85–95. [Google Scholar] [CrossRef] [PubMed]
- Bernardo, M.; Emons, J.; Karperien, M.; Nauta, A.; Willemze, R.; Roelofs, H.; Romeo, S.; Marchini, A.; Rappold, G.; Vukicevic, S. Human mesenchymal stem cells derived from bone marrow display a better chondrogenic differentiation compared with other sources. Connect. Tissue Res. 2007, 48, 132–140. [Google Scholar] [CrossRef] [PubMed]
- Klingemann, H.; Matzilevich, D.; Marchand, J. Mesenchymal stem cells–sources and clinical applications. Transfus. Med. Hemother. 2008, 35, 272–277. [Google Scholar] [CrossRef] [PubMed]
- Silini, A.R.; Di Pietro, R.; Lang-Olip, I.; Alviano, F.; Banerjee, A.; Basile, M.; Borutinskaite, V.; Eissner, G.; Gellhaus, A.; Giebel, B. Perinatal derivatives: Where do we stand? A roadmap of the human placenta and consensus for tissue and cell nomenclature. Front. Bioeng. Biotechnol. 2020, 8, 1438. [Google Scholar] [CrossRef]
- Parolini, O.; Alviano, F.; Bagnara, G.P.; Bilic, G.; Bühring, H.-J.; Evangelista, M.; Hennerbichler, S.; Liu, B.; Magatti, M.; Mao, N. Concise review: Isolation and characterization of cells from human term placenta: Outcome of the first international Workshop on Placenta Derived Stem Cells. Stem Cells 2008, 26, 300–311. [Google Scholar] [CrossRef]
- Sripa, B.; Leungwattanawanit, S.; Nitta, T.; Wongkham, C.; Bhudhisawasdi, V.; Puapairoj, A.; Sripa, C.; Miwa, M. Establishment and characterization of an opisthorchiasis-associated cholangiocarcinoma cell line (KKU-100). World J. Gastroenterol. 2005, 11, 3392. [Google Scholar] [CrossRef]
- Sripa, B.; Seubwai, W.; Vaeteewoottacharn, K.; Sawanyawisuth, K.; Silsirivanit, A.; Kaewkong, W.; Muisuk, K.; Dana, P.; Phoomak, C.; Lert-Itthiporn, W. Functional and genetic characterization of three cell lines derived from a single tumor of an Opisthorchis viverrini-associated cholangiocarcinoma patient. Hum. Cell 2020, 33, 695–708. [Google Scholar] [CrossRef]
- Thanee, M.; Padthaisong, S.; Suksawat, M.; Dokduang, H.; Phetcharaburanin, J.; Klanrit, P.; Titapun, A.; Namwat, N.; Wangwiwatsin, A.; Sa-Ngiamwibool, P. Sulfasalazine modifies metabolic profiles and enhances cisplatin chemosensitivity on cholangiocarcinoma cells in in vitro and in vivo models. Cancer Metab. 2021, 9, 11. [Google Scholar] [CrossRef]
- Wang, W.; Zhan, M.; Li, Q.; Chen, W.; Chu, H.; Huang, Q.; Hou, Z.; Man, M.; Wang, J. FXR agonists enhance the sensitivity of biliary tract cancer cells to cisplatin via SHP dependent inhibition of Bcl-xL expression. Oncotarget 2016, 7, 34617. [Google Scholar] [CrossRef] [PubMed]
- Polewski, M.D.; Nielsen, G.B.; Gu, Y.; Weaver, A.T.; Gegg, G.; Tabuena-Frolli, S.; Cajaiba, M.; Hanks, D.; Method, M.; Press, M.F. A Standardized Investigational Ki-67 Immunohistochemistry Assay Used to Assess High-Risk Early Breast Cancer Patients in the monarchE Phase 3 Clinical Study Identifies a Population With Greater Risk of Disease Recurrence When Treated With Endocrine Therapy Alone. Appl. Immunohistochem. Mol. Morphol. 2022, 30, 237. [Google Scholar] [PubMed]
- Liu, C.; Feng, X.; Wang, B.; Wang, X.; Wang, C.; Yu, M.; Cao, G.; Wang, H. Bone marrow mesenchymal stem cells promote head and neck cancer progression through Periostin-mediated phosphoinositide 3-kinase/Akt/mammalian target of rapamycin. Cancer Sci. 2018, 109, 688–698. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Lee, Y.W.; Rui, Y.F.; Cheng, T.Y.; Jiang, X.H.; Li, G. Bone marrow-derived mesenchymal stem cells promote growth and angiogenesis of breast and prostate tumors. Stem Cell Res. Ther. 2013, 4, 70. [Google Scholar] [CrossRef]
- Wu, Y.L.; Li, H.Y.; Zhao, X.P.; Jiao, J.Y.; Tang, D.X.; Yan, L.J.; Wan, Q.; Pan, C.B. Mesenchymal stem cell-derived CCN 2 promotes the proliferation, migration and invasion of human tongue squamous cell carcinoma cells. Cancer Sci. 2017, 108, 897–909. [Google Scholar] [CrossRef]
- Kalamegam, G.; Sait, K.H.W.; Anfinan, N.; Kadam, R.; Ahmed, F.; Rasool, M.; Naseer, M.I.; Pushparaj, P.N.; Al-Qahtani, M. Cytokines secreted by human Wharton’s jelly stem cells inhibit the proliferation of ovarian cancer (OVCAR3) cells in vitro. Oncol. Lett. 2019, 17, 4521–4531. [Google Scholar] [CrossRef]
- Hendijani, F.; Javanmard, S.H.; Sadeghi-aliabadi, H. Human Wharton’s jelly mesenchymal stem cell secretome display antiproliferative effect on leukemia cell line and produce additive cytotoxic effect in combination with doxorubicin. Tissue Cell 2015, 47, 229–234. [Google Scholar] [CrossRef]
- Sagaradze, G.; Grigorieva, O.; Nimiritsky, P.; Basalova, N.; Kalinina, N.; Akopyan, Z.; Efimenko, A. Conditioned medium from human mesenchymal stromal cells: Towards the clinical translation. Int. J. Mol. Sci. 2019, 20, 1656. [Google Scholar] [CrossRef]
- Zhao, J.; Zhang, Z.; Cui, Q.; Zhao, L.; Hu, Y.; Zhao, S. Human adipose-derived mesenchymal stem cells inhibit proliferation and induce apoptosis of human gastric cancer HGC-27 cells. 3 Biotech 2020, 10, 129. [Google Scholar] [CrossRef]
- Clarke, M.R.; Imhoff, F.M.; Baird, S.K. Mesenchymal stem cells inhibit breast cancer cell migration and invasion through secretion of tissue inhibitor of metalloproteinase-1 and-2. Mol. Carcinog. 2015, 54, 1214–1219. [Google Scholar] [CrossRef]
- Lee, J.-K.; Park, S.-R.; Jung, B.-K.; Jeon, Y.-K.; Lee, Y.-S.; Kim, M.-K.; Kim, Y.-G.; Jang, J.-Y.; Kim, C.-W. Exosomes derived from mesenchymal stem cells suppress angiogenesis by down-regulating VEGF expression in breast cancer cells. PLoS ONE 2013, 8, e84256. [Google Scholar] [CrossRef] [PubMed]
- Katakowski, M.; Buller, B.; Zheng, X.; Lu, Y.; Rogers, T.; Osobamiro, O.; Shu, W.; Jiang, F.; Chopp, M. Exosomes from marrow stromal cells expressing miR-146b inhibit glioma growth. Cancer Lett. 2013, 335, 201–204. [Google Scholar] [CrossRef] [PubMed]
- Wong, R.S. Apoptosis in cancer: From pathogenesis to treatment. J. Exp. Clin. Cancer Res. 2011, 30, 87. [Google Scholar] [CrossRef] [PubMed]
- Siu, W.P.; Pun, P.B.L.; Latchoumycandane, C.; Boelsterli, U.A. Bax-mediated mitochondrial outer membrane permeabilization (MOMP), distinct from the mitochondrial permeability transition, is a key mechanism in diclofenac-induced hepatocyte injury: Multiple protective roles of cyclosporin A. Toxicol. Appl. Pharmacol. 2008, 227, 451–461. [Google Scholar] [CrossRef]
- Tsujimoto, Y. Role of Bcl-2 family proteins in apoptosis: Apoptosomes or mitochondria? Genes Cells. 1998, 3, 697–707. [Google Scholar] [CrossRef] [PubMed]
- Fan, T.-J.; Han, L.-H.; Cong, R.-S.; Liang, J. Caspase family proteases and apoptosis. Acta Biochim. Biophys. Sin. 2005, 37, 719–727. [Google Scholar] [CrossRef]
- Fiandalo, M.; Kyprianou, N. Caspase control: Protagonists of cancer cell apoptosis. Exp. Oncol. 2012, 34, 165. [Google Scholar]
- Bu, S.; Wang, Q.; Zhang, Q.; Sun, J.; He, B.; Xiang, C.; Liu, Z.; Lai, D. Human endometrial mesenchymal stem cells exhibit intrinsic anti-tumor properties on human epithelial ovarian cancer cells. Sci. Rep. 2016, 6, 37019. [Google Scholar] [CrossRef]
- Yoshikawa, T.; Miyamoto, M.; Aoyama, T.; Soyama, H.; Goto, T.; Hirata, J.; Suzuki, A.; Nagaoka, I.; Tsuda, H.; Furuya, K. JAK2/STAT3 pathway as a therapeutic target in ovarian cancers. Oncol. Lett. 2018, 15, 5772–5780. [Google Scholar] [CrossRef]
- Judd, L.M.; Menheniott, T.R.; Ling, H.; Jackson, C.B.; Howlett, M.; Kalantzis, A.; Priebe, W.; Giraud, A.S. Inhibition of the JAK2/STAT3 pathway reduces gastric cancer growth in vitro and in vivo. PLoS ONE 2014, 9, e95993. [Google Scholar] [CrossRef]
- Jin, W. Role of JAK/STAT3 signaling in the regulation of metastasis, the transition of cancer stem cells, and chemoresistance of cancer by epithelial–mesenchymal transition. Cells 2020, 9, 217. [Google Scholar] [CrossRef] [PubMed]
- Yang, N.; Han, F.; Cui, H.; Huang, J.; Wang, T.; Zhou, Y.; Zhou, J. Matrine suppresses proliferation and induces apoptosis in human cholangiocarcinoma cells through suppression of JAK2/STAT3 signaling. Pharmacol. Rep. 2015, 67, 388–393. [Google Scholar] [CrossRef] [PubMed]
- Dokduang, H.; Techasen, A.; Namwat, N.; Khuntikeo, N.; Pairojkul, C.; Murakami, Y.; Loilome, W.; Yongvanit, P. STAT s profiling reveals predominantly-activated STAT3 in cholangiocarcinoma genesis and progression. J. Hepatobiliary Pancreat. Sci. 2014, 21, 767–776. [Google Scholar] [CrossRef] [PubMed]
- Laudisi, F.; Cherubini, F.; Monteleone, G.; Stolfi, C. STAT3 interactors as potential therapeutic targets for cancer treatment. Int. J. Mol. Sci. 2018, 19, 1787. [Google Scholar] [CrossRef] [PubMed]
- Johnson, D.E.; O’Keefe, R.A.; Grandis, J.R. Targeting the IL-6/JAK/STAT3 signalling axis in cancer. Nat. Rev. Clin. Oncol. 2018, 15, 234. [Google Scholar] [CrossRef]
- Xiong, A.; Yang, Z.; Shen, Y.; Zhou, J.; Shen, Q. Transcription factor STAT3 as a novel molecular target for cancer prevention. Cancers 2014, 6, 926–957. [Google Scholar] [CrossRef]
- Jantalika, T.; Manochantr, S.; Kheolamai, P.; Tantikanlayaporn, D.; Saijuntha, W.; Pinlaor, S.; Chairoungdua, A.; Paraoan, L.; Tantrawatpan, C. Human chorion-derived mesenchymal stem cells suppress JAK2/STAT3 signaling and induce apoptosis of cholangiocarcinoma cell lines. Sci. Rep. 2022, 12, 11341. [Google Scholar] [CrossRef]
- Liu, Q.W.; Li, J.Y.; Zhang, X.C.; Liu, Y.; Liu, Q.Y.; Xiao, L.; Zhang, W.J.; Wu, H.Y.; Deng, K.Y.; Xin, H.B. Human amniotic mesenchymal stem cells inhibit hepatocellular carcinoma in tumour-bearing mice. J. Cell. Mol. Med. 2020, 24, 10525–10541. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jantalika, T.; Manochantr, S.; Kheolamai, P.; Tantikanlayaporn, D.; Thongsepee, N.; Warnnissorn, N.; Saijuntha, W.; Pinlaor, S.; Tantrawatpan, C. The Human Placental Amniotic Membrane Mesenchymal-Stromal-Cell-Derived Conditioned Medium Inhibits Growth and Promotes Apoptosis of Human Cholangiocarcinoma Cells In Vitro and In Vivo by Suppressing IL-6/JAK2/STAT3 Signaling. Cells 2023, 12, 2788. https://doi.org/10.3390/cells12242788
Jantalika T, Manochantr S, Kheolamai P, Tantikanlayaporn D, Thongsepee N, Warnnissorn N, Saijuntha W, Pinlaor S, Tantrawatpan C. The Human Placental Amniotic Membrane Mesenchymal-Stromal-Cell-Derived Conditioned Medium Inhibits Growth and Promotes Apoptosis of Human Cholangiocarcinoma Cells In Vitro and In Vivo by Suppressing IL-6/JAK2/STAT3 Signaling. Cells. 2023; 12(24):2788. https://doi.org/10.3390/cells12242788
Chicago/Turabian StyleJantalika, Tanachapa, Sirikul Manochantr, Pakpoom Kheolamai, Duangrat Tantikanlayaporn, Nattaya Thongsepee, Naree Warnnissorn, Weerachai Saijuntha, Somchai Pinlaor, and Chairat Tantrawatpan. 2023. "The Human Placental Amniotic Membrane Mesenchymal-Stromal-Cell-Derived Conditioned Medium Inhibits Growth and Promotes Apoptosis of Human Cholangiocarcinoma Cells In Vitro and In Vivo by Suppressing IL-6/JAK2/STAT3 Signaling" Cells 12, no. 24: 2788. https://doi.org/10.3390/cells12242788