

Enterotoxin Gene Cluster and selX Are Associated with Atopic Dermatitis Severity—A Cross-Sectional Molecular Study of Staphylococcus aureus Superantigens
 , , ,
, , ,
Abstract
:1. Introduction
2. Methods
2.1. Staphylococcus aureus Isolation
2.2. Genomic DNA Extraction
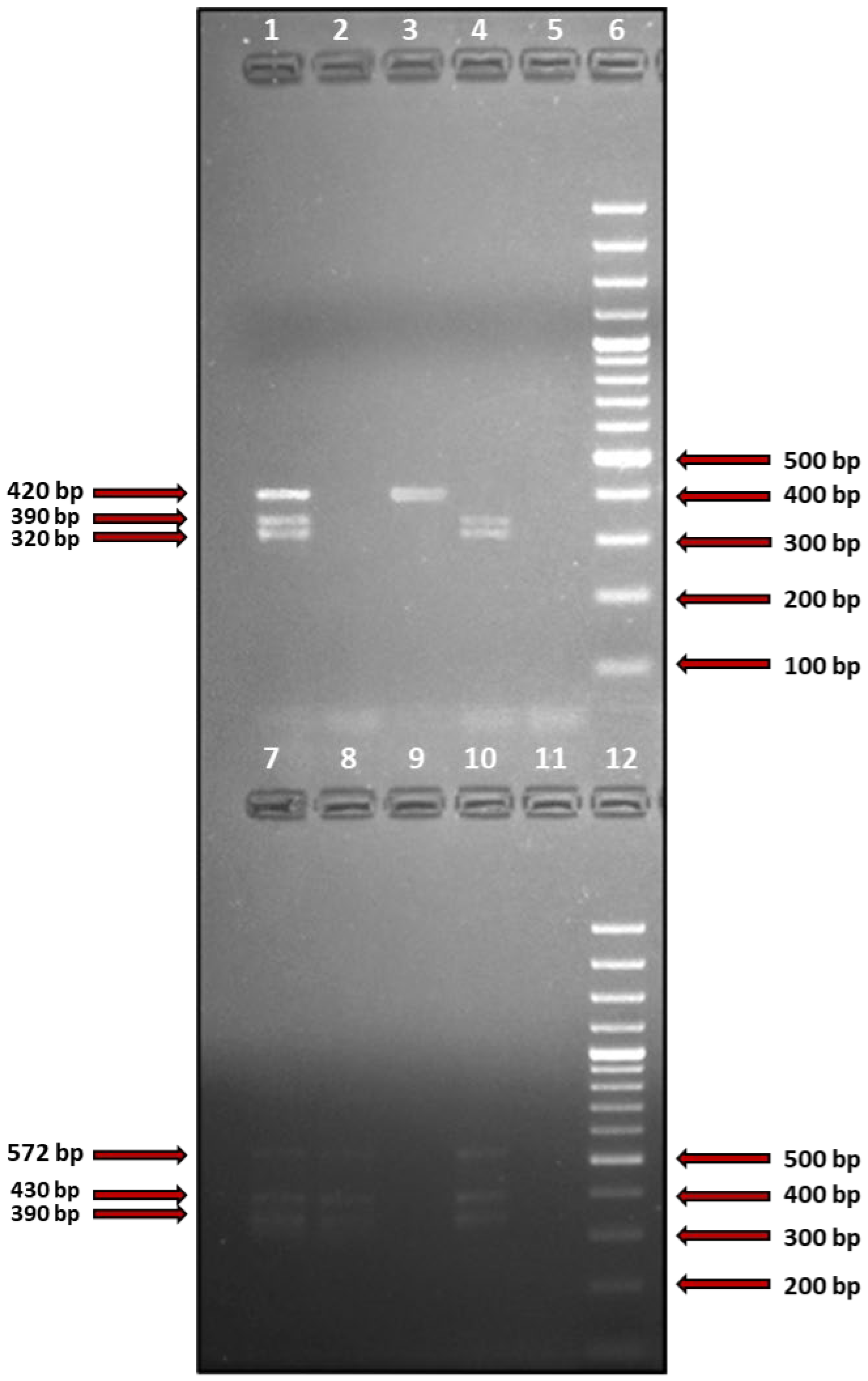
2.3. Detection of S. aureus Superantigen-Coding Genes
2.4. Identification of the Obtained PCR Products
2.5. Statistical Analysis
3. Results
- on the lesional skin: selX (23/43 strains, 53.5%), selN (9/43 strains, 20.9%), seg, selI, selM, selO, selU (all identified in 7/43 strains, 16.3%). see was not identified in any of the isolates;
- on the nonlesional skin: selX (22/33 strains, 66.7%), selN (10/33 strains, 30.3%), seg, selI, selM, selO, selU (all identified in 9/33 strains, 27,6%). seb, sed, and see were not identified in any of the isolates;
- in the anterior nares: selX (15/37 strains, 40.5%), selN (9/37 strains, 24.3%), selM, selO (both identified in 5/37 strains, 13.5%). seb and see were not identified in any of the isolates.
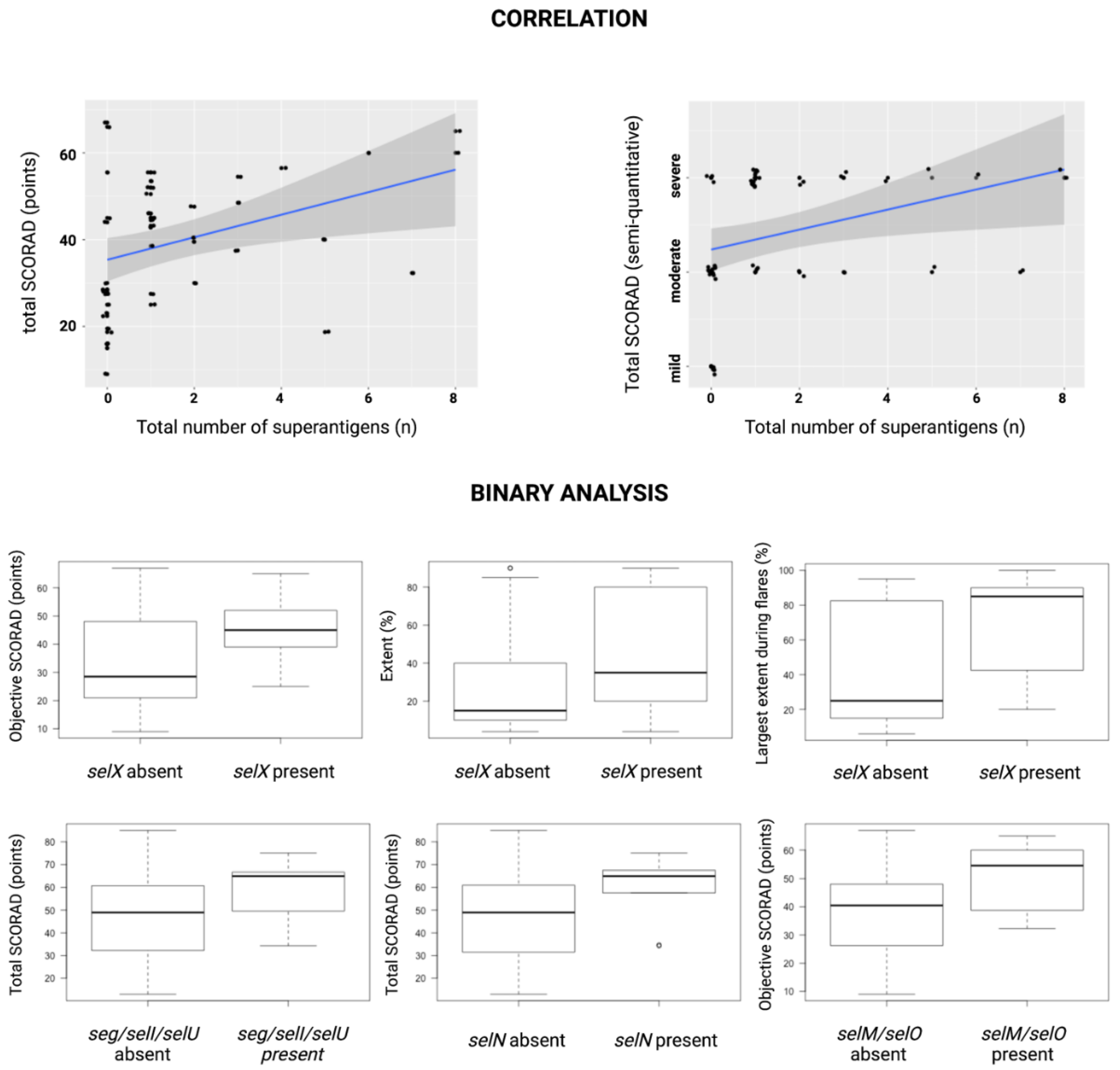
- selN—higher mean values of total SCORAD (58.73 ± 14.6 vs. 47.04 ± 17.14, p < 0.03), intensity (11.33 ± 3.24 vs. 8.59 ± 3, p < 0.04), redness (2.22 ± 0.97 vs. 1.49 ± 0.68, p < 0.03), and lichenification (2.44 ± 0.53 vs. 1.80 ± 0.78, p < 0.03);
- seg, selI, and selU—higher mean values of total SCORAD (58.14 ± 16.53 vs. 47.68 ± 17.02, p < 0.05) and lichenification (2.57 ± 0.53 vs. 1.81 ± 0.76, p < 0.02);
- selM and selO—higher mean values of objective SCORAD (49.9 ± 13 vs. 37.58 ± 14.84, p < 0.05), intensity (11 ± 1.83 vs. 8.77 ± 3.27, p < 0.03), and dryness (2.29 ± 0.49 vs. 1.40 ± 0.93, p < 0.02)
4. Discussion
4.1. Therapeutic Implications
4.2. Study Limitations
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Weidinger, S.; Beck, L.A.; Bieber, T.; Kabashima, K.; Irvine, A.D. Atopic Dermatitis. Nat. Rev. Dis. Primer 2018, 4, 1. [Google Scholar] [CrossRef] [PubMed]
- Langan, S.M.; Irvine, A.D.; Weidinger, S. Atopic Dermatitis. Lancet Lond. Engl. 2020, 396, 345–360. [Google Scholar] [CrossRef] [PubMed]
- Sroka-Tomaszewska, J.; Trzeciak, M. Molecular Mechanisms of Atopic Dermatitis Pathogenesis. Int. J. Mol. Sci. 2021, 22, 4130. [Google Scholar] [CrossRef] [PubMed]
- Blicharz, L.; Rudnicka, L.; Czuwara, J.; Waśkiel-Burnat, A.; Goldust, M.; Olszewska, M.; Samochocki, Z. The Influence of Microbiome Dysbiosis and Bacterial Biofilms on Epidermal Barrier Function in Atopic Dermatitis—An Update. Int. J. Mol. Sci. 2021, 22, 8403. [Google Scholar] [CrossRef] [PubMed]
- Koh, L.F.; Ong, R.Y.; Common, J.E. Skin Microbiome of Atopic Dermatitis. Allergol. Int. 2022, 71, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Paller, A.S.; Kong, H.H.; Seed, P.; Naik, S.; Scharschmidt, T.C.; Gallo, R.L.; Luger, T.; Irvine, A.D. The Microbiome in Patients with Atopic Dermatitis. J. Allergy Clin. Immunol. 2019, 143, 26–35. [Google Scholar] [CrossRef] [Green Version]
- Tam, K.; Torres, V.J. Staphylococcus Aureus Secreted Toxins and Extracellular Enzymes. Microbiol. Spectr. 2019, 7, 16. [Google Scholar] [CrossRef]
- Blicharz, L.; Rudnicka, L.; Samochocki, Z. Staphylococcus Aureus: An Underestimated Factor in the Pathogenesis of Atopic Dermatitis? Postepy Dermatol. Alergol. 2019, 36, 11–17. [Google Scholar] [CrossRef]
- Oogai, Y.; Matsuo, M.; Hashimoto, M.; Kato, F.; Sugai, M.; Komatsuzawa, H. Expression of Virulence Factors by Staphylococcus Aureus Grown in Serum. Appl. Environ. Microbiol. 2011, 77, 8097–8105. [Google Scholar] [CrossRef] [Green Version]
- Novick, R.P. Pathogenicity Islands and Their Role in Staphylococcal Biology. Microbiol. Spectr. 2019, 7, 21. [Google Scholar] [CrossRef]
- Schwendimann, L.; Merda, D.; Berger, T.; Denayer, S.; Feraudet-Tarisse, C.; Kläui, A.J.; Messio, S.; Mistou, M.Y.; Nia, Y.; Hennekinne, J.A.; et al. Staphylococcal Enterotoxin Gene Cluster: Prediction of Enterotoxin (SEG and SEI) Production and of the Source of Food Poisoning on the Basis of v Saβ Typing. Appl. Environ. Microbiol. 2021, 87, e02662-20. [Google Scholar] [CrossRef] [PubMed]
- Aung, M.S.; Urushibara, N.; Kawaguchiya, M.; Ito, M.; Habadera, S.; Kobayashi, N. Prevalence and Genetic Diversity of Staphylococcal Enterotoxin (-Like) Genes Sey, Selw, Selx, Selz, Sel26 and Sel27 in Community-Acquired Methicillin-Resistant Staphylococcus Aureus. Toxins 2020, 12, 347. [Google Scholar] [CrossRef] [PubMed]
- Langley, R.J.; Ting, Y.T.; Clow, F.; Young, P.G.; Radcliff, F.J.; Choi, J.M.; Sequeira, R.P.; Holtfreter, S.; Baker, H.; Fraser, J.D. Staphylococcal Enterotoxin-like X (SElX) Is a Unique Superantigen with Functional Features of Two Major Families of Staphylococcal Virulence Factors. PLoS Pathog. 2017, 13, e1006549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanifin, J.M.; Rajka, G. Diagnostic Features of Atopic Dermatitis. Acta Derm. Venereol. 1980, 60, 44–47. [Google Scholar]
- Wallace, A.B. The Exposure Treatment of Burns. Lancet Lond. Engl. 1951, 1, 501–504. [Google Scholar] [CrossRef]
- Salgado-Pabón, W.; Case-Cook, L.C.; Schlievert, P.M. Molecular Analysis of Staphylococcal Superantigens. Methods Mol. Biol. Clifton NJ 2014, 1085, 169–185. [Google Scholar] [CrossRef]
- Vu, B.G.; Stach, C.S.; Salgado-Pabón, W.; Diekema, D.J.; Gardner, S.E.; Schlievert, P.M. Superantigens of Staphylococcus Aureus from Patients with Diabetic Foot Ulcers. J. Infect. Dis. 2014, 210, 1920–1927. [Google Scholar] [CrossRef] [Green Version]
- Nada, H.A.; Gomaa, N.I.M.; Elakhras, A.; Wasfy, R.; Baker, R.A. Skin Colonization by Superantigen-Producing Staphylococcus Aureus in Egyptian Patients with Atopic Dermatitis and Its Relation to Disease Severity and Serum Interleukin-4 Level. Int. J. Infect. Dis. 2012, 16, e29–e33. [Google Scholar] [CrossRef] [Green Version]
- Van, T.C.; Tat, T.N.; Lan, A.T.; Van, T.N.; Gandolfi, M.; Satolli, F.; Feliciani, C.; Tirant, M.; Vojvodic, A.; Lotti, T. Superantigens of Staphylococcus Aureus Colonization in Atopic Dermatitis and Treatment Efficacy of Oral Cefuroxim in Vietnamese Patients. Open Access Maced. J. Med. Sci. 2019, 7, 243–246. [Google Scholar] [CrossRef]
- Schlievert, P.M.; Case, L.C.; Strandberg, K.L.; Abrams, B.B.; Leung, D.Y.M. Superantigen Profile of Staphylococcus Aureus Isolates from Patients with Steroid-Resistant Atopic Dermatitis. Clin. Infect. Dis. 2008, 46, 1562–1567. [Google Scholar] [CrossRef] [Green Version]
- Fukushima, H.; Hirano, T.; Shibayama, N.; Miwa, K.; Ito, T.; Saito, M.; Sumida, H.; Oyake, S.; Tsuboi, R.; Oka, K. The Role of Immune Response to Staphylococcus Aureus Superantigens and Disease Severity in Relation to the Sensitivity to Tacrolimus in Atopic Dermatitis. Int. Arch. Allergy Immunol. 2006, 141, 281–289. [Google Scholar] [CrossRef] [PubMed]
- Sonkoly, E.; Muller, A.; Lauerma, A.I.; Pivarcsi, A.; Soto, H.; Kemeny, L.; Alenius, H.; Dieu-Nosjean, M.-C.; Meller, S.; Rieker, J.; et al. IL-31: A New Link between T Cells and Pruritus in Atopic Skin Inflammation. J. Allergy Clin. Immunol. 2006, 117, 411–417. [Google Scholar] [CrossRef] [PubMed]
- de Wit, J.; Totté, J.E.E.; van Buchem, F.J.M.; Pasmans, S.G.M.A. The Prevalence of Antibody Responses against Staphylococcus Aureus Antigens in Patients with Atopic Dermatitis: A Systematic Review and Meta-Analysis. Br. J. Dermatol. 2018, 178, e438. [Google Scholar] [CrossRef] [Green Version]
- Merriman, J.A.; Mueller, E.A.; Cahill, M.P.; Beck, L.A.; Paller, A.S.; Hanifin, J.M.; Ong, P.Y.; Schneider, L.; Babineau, D.C.; David, G.; et al. Temporal and Racial Differences Associated with Atopic Dermatitis Staphylococcus Aureus and Encoded Virulence Factors. mSphere 2016, 1, e00295-16. [Google Scholar] [CrossRef] [Green Version]
- Fisher, E.L.; Otto, M.; Cheung, G.Y.C. Basis of Virulence in Enterotoxin-Mediated Staphylococcal Food Poisoning. Front. Microbiol. 2018, 9, 436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakr, A.; Brégeon, F.; Mège, J.-L.; Rolain, J.-M.; Blin, O. Staphylococcus Aureus Nasal Colonization: An Update on Mechanisms, Epidemiology, Risk Factors, and Subsequent Infections. Front. Microbiol. 2018, 9, 2419. [Google Scholar] [CrossRef]
- Cho, S.-H.; Strickland, I.; Boguniewicz, M.; Leung, D.Y.M. Fibronectin and Fibrinogen Contribute to the Enhanced Binding of Staphylococcus Aureus to Atopic Skin. J. Allergy Clin. Immunol. 2001, 108, 269–274. [Google Scholar] [CrossRef]
- Mempel, M.; Lina, G.; Hojka, M.; Schnopp, C.; Seidl, H.-P.; Schäfer, T.; Ring, J.; Vandenesch, F.; Abeck, D. High Prevalence of Superantigens Associated with the Egc Locus in Staphylococcus Aureus Isolates from Patients with Atopic Eczema. Eur. J. Clin. Microbiol. Infect. Dis. 2003, 22, 306–309. [Google Scholar] [CrossRef]
- Mekhloufi, O.A.; Chieffi, D.; Hammoudi, A.; Bensefia, S.A.; Fanelli, F.; Fusco, V. Prevalence, Enterotoxigenic Potential and Antimicrobial Resistance of Staphylococcus Aureus and Methicillin-Resistant Staphylococcus Aureus (MRSA) Isolated from Algerian Ready to Eat Foods. Toxins 2021, 13, 835. [Google Scholar] [CrossRef]
- Moran, M.C.; Cahill, M.P.; Brewer, M.G.; Yoshida, T.; Knowlden, S.; Perez-Nazario, N.; Schlievert, P.M.; Beck, L.A. Staphylococcal Virulence Factors on the Skin of Atopic Dermatitis Patients. mSphere 2019, 4, e00616-19. [Google Scholar] [CrossRef] [Green Version]
- Roetzer, A.; Haller, G.; Beyerly, J.; Geier, C.B.; Wolf, H.M.; Gruener, C.S.; Model, N.; Eibl, M.M. Genotypic and Phenotypic Analysis of Clinical Isolates of Staphylococcus Aureus Revealed Production Patterns and Hemolytic Potentials Unlinked to Gene Profiles and Source. BMC Microbiol. 2016, 16, 13. [Google Scholar] [CrossRef] [PubMed]
- Wilson, G.J.; Seo, K.S.; Cartwright, R.A.; Connelley, T.; Chuang-Smith, O.N.; Merriman, J.A.; Guinane, C.M.; Park, J.Y.; Bohach, G.A.; Schlievert, P.M.; et al. A Novel Core Genome-Encoded Superantigen Contributes to Lethality of Community-Associated MRSA Necrotizing Pneumonia. PLoS Pathog. 2011, 7, e1002271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parker, D.; Ryan, C.L.; Alonzo, F.; Torres, V.J.; Planet, P.J.; Prince, A.S. CD4+ T Cells Promote the Pathogenesis of Staphylococcus Aureus Pneumonia. J. Infect. Dis. 2015, 211, 835–845. [Google Scholar] [CrossRef] [Green Version]
- Rasmussen, G.; Monecke, S.; Ehricht, R.; Söderquist, B. Prevalence of Clonal Complexes and Virulence Genes among Commensal and Invasive Staphylococcus Aureus Isolates in Sweden. PLoS ONE 2013, 8, e77477. [Google Scholar] [CrossRef] [PubMed]
- Tuffs, S.W.; James, D.B.A.; Bestebroer, J.; Richards, A.C.; Goncheva, M.I.; O’Shea, M.; Wee, B.A.; Seo, K.S.; Schlievert, P.M.; Lengeling, A.; et al. The Staphylococcus Aureus Superantigen SElX Is a Bifunctional Toxin That Inhibits Neutrophil Function. PLOS Pathog. 2017, 13, e1006461. [Google Scholar] [CrossRef] [Green Version]
- Fevre, C.; Bestebroer, J.; Mebius, M.M.; de Haas, C.J.C.; van Strijp, J.A.G.; Fitzgerald, J.R.; Haas, P.-J.A. Staphylococcus Aureus Proteins SSL6 and SElX Interact with Neutrophil Receptors as Identified Using Secretome Phage Display. Cell. Microbiol. 2014, 16, 1646–1665. [Google Scholar] [CrossRef] [PubMed]
- Soares, J.; Lopes, C.; Tavaria, F.; Delgado, L.; Pintado, M. A Diversity Profile from the Staphylococcal Community on Atopic Dermatitis Skin: A Molecular Approach. J. Appl. Microbiol. 2013, 115, 1411–1419. [Google Scholar] [CrossRef] [PubMed]
- Prignano, F.; Donetti, E. Looking at Interleukin-22 from a New Dermatological Perspective: From Epidermal Homeostasis to Its Role in Chronic Skin Diseases. Dermatol. Basel Switz. 2022, 238, 829–836. [Google Scholar] [CrossRef]
- Zhao, N.; Cheng, D.; Yang, Z.; Liu, Y.; Wang, Y.; Jian, Y.; Wang, H.; Li, M.; Bae, T.; Liu, Q. Virulence Adaption to Environment Promotes the Age-Dependent Nasal Colonization of Staphylococcus Aureus. Emerg. Microbes Infect. 2022, 11, 1402–1415. [Google Scholar] [CrossRef]
- Orfali, R.L.; da Silva Oliveira, L.M.; de Lima, J.F.; de Carvalho, G.C.; Ramos, Y.A.L.; Pereira, N.Z.; Pereira, N.V.; Zaniboni, M.C.; Sotto, M.N.; da Silva Duarte, A.J.; et al. Staphylococcus Aureus Enterotoxins Modulate IL-22-Secreting Cells in Adults with Atopic Dermatitis. Sci. Rep. 2018, 8, 6665. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niebuhr, M.; Mamerow, D.; Heratizadeh, A.; Satzger, I.; Werfel, T. Staphylococcal α-Toxin Induces a Higher T Cell Proliferation and Interleukin-31 in Atopic Dermatitis. Int. Arch. Allergy Immunol. 2011, 156, 412–415. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.-T.; Wang, C.-T.; Chao, P.-S.; Lee, J.-H.; Wang, L.-C.; Yu, H.-H.; Yang, Y.-H.; Chiang, B.-L. Skin-Homing CD4+ Foxp3+ T Cells Exert Th2-like Function after Staphylococcal Superantigen Stimulation in Atopic Dermatitis Patients: Skin-Homing SsAg Stimulated Foxp3+ T Cells in AD. Clin. Exp. Allergy 2011, 41, 516–525. [Google Scholar] [CrossRef] [PubMed]
- Friedman, M.; Rasooly, R.; Do, P.M.; Henika, P.R. The Olive Compound 4-Hydroxytyrosol Inactivates Staphylococcus Aureus Bacteria and Staphylococcal Enterotoxin A (SEA). J. Food Sci. 2011, 76, M558–M563. [Google Scholar] [CrossRef] [PubMed]
- Hisano, M.; Yamaguchi, K.; Inoue, Y.; Ikeda, Y.; Iijima, M.; Adachi, M.; Shimamura, T. Inhibitory Effect of Catechin against the Superantigen Staphylococcal Enterotoxin B (SEB). Arch. Dermatol. Res. 2003, 295, 183–189. [Google Scholar] [CrossRef] [PubMed]
- Rasooly, R.; Do, P.M.; Friedman, M. Inhibition of Biological Activity of Staphylococcal Enterotoxin A (SEA) by Apple Juice and Apple Polyphenols. J. Agric. Food Chem. 2010, 58, 5421–5426. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Name of the Gene | Name of the Primer | Nucleotide Sequence of the Primer (5′→3′) | PCR Product Size (bp) | Reference |
|---|---|---|---|---|
| sea | SEA-F | GATTCACAAAGGATATTGTTGATAAATAT | 400 | Salgado-Pabon W, et al. |
| SEA-R | GTCCTTGAGCACCAAATAAATC | |||
| seb | SEB-F | GTATGATGATAATCATGTATCAGCAA | 625 | Salgado-Pabon W, et al. |
| SEB-R | CGTAAGATAAACTTCAATCTTCACAT | |||
| sec | SEC-F | GAGTCAACCAGACCCTATGCC | 650 | Salgado-Pabon W, et al. |
| SEC-R | CGCCTGGTGCAGGCATC | |||
| sed | SED-F | GCATTACTCTTTTTTACTAGTTTGGTA | 530 | Salgado-Pabon W, et al. |
| SED-R | CCTTGCTTGTGCATCTAATTC | |||
| see | SEE-F | CTGAATTACAAAGAAATGCTTTAAGC | 420 | Salgado-Pabon W, et al. |
| SEE-R | GCCTTGCCTGAAGATCTA | |||
| seg | SEG-F | GGGAACTATGGGTAATGTAATGAATC | 430 | Vu BG, et al. |
| SEG-R | TGAGCCAGTGTCTTGCTTTG | |||
| selI | SEL-I-F | GCTCAAGGTGATATTGGTGTAGG | 572 | Vu BG, et al. |
| SEL-I-R | CTTACAGGCAGTCCATCTCC | |||
| selM | SEL-M-F | CGGTGGAGTTACATTAGCAGGT | 320 | Vu BG, et al. |
| SEL-M-R | TTTCAGCTTGTCCTGTTCCA | |||
| selN | SEL-N-F | GCTTATACGGAGGAGTTACG | 298 | Vu BG, et al. |
| SEL-N-R | GCTCCCACTGAACCTTTTACG | |||
| selO | SEL-O-F | GGAATTTAGCTCATCAGCGATT | 390 | Vu BG, et al. |
| SEL-O-R | TGCTCCGAATGAGAATGAAA | |||
| selU | SEL-U-F | GCAGCTTACTATTTATGTTAAATGGC | 390 | Vu BG, et al. |
| SEL-U-R | CTATTTGATTTCCATCATGCTCGG | |||
| selX | SEL-X-F | TCTATGGGGGAACATTTGGA | 420 | Salgado-Pabon W, et al. |
| SEL-X-R | CCGCCATCTTTTGTATTTATGA | |||
| tstH | TSST-1-F | GAAATTTTTCATCGTAAGCCCTTTGTTG | 655 | Salgado-Pabon W, et al. |
| TSST-1-R | TTCATCAATATTTATAGGTGGTTTTTCA |
| Females/males | 17/33 (34%/66%) |
| Age | range 18–70, mean 32.7 ± 12.9 |
| SCORAD index (points) | range 13–85, mean 49.1 ± 17.2 |
| <25 points (mild AD) | 5/50 (10%) |
| 25–50 points (moderate AD) | 19/50 (38%) |
| >50 points (severe) | 26/50 (52%) |
| Extent—% of involved body area | range 4–90, mean 37.6 ± 32 |
| Intensity (points): | |
| Redness | range 0–3, mean 1.62 ± 0.78 |
| Swelling | range 0–3, mean 1.4 ± 0.67 |
| Oozing/crust | range 0–3, mean 1.08 ± 0.99 |
| Scratch marks | range 0–3, mean 1.54 ± 0.84 |
| Lichenification | range 0–3, mean 1.92 ± 0.78 |
| Dryness | range 0–3, mean 1.52 ± 0.93 |
| Subjective symptoms (points): | |
| Itch | range 0–10, mean 6.0 ± 2.77 |
| Sleeplessness | range 0–10, mean 3.84 ± 3.59 |
| Objective SCORAD (points) | range 9–67, mean 39.3 ± 15.1 |
| Largest extent of skin lesions during flares in the year preceding the examination (%) | range 6–100, mean 56.2 ± 34.6 |
| Extent of skin lesions during stable periods of the disease (%) | range 0–15, mean 4.68 ± 3.68 |
| Total IgE (UI/mL) | range 5.4–2500, mean 1354 ± 1081 |
| Gene | Lesional Skin, n (%) | Nonlesional Skin, n (%) | Anterior Nares, n (%) |
|---|---|---|---|
| sea | 3 (7.0%) | 2 (6.1%) | 3 (8.1%) |
| seb | 1 (2.3%) | - | - |
| sec | 3 (7.0%) | 1 (3.0%) | 2 (5.4%) |
| sed | 1 (2.3%) | - | 2 (5.4%) |
| see | - | - | - |
| seg | 7 (16.3%) | 9 (27.3%) | 3 (8.1%) |
| selI | 7 (16.3%) | 9 (27.3%) | 3 (8.1%) |
| selM | 7 (16.3%) | 9 (27.3%) | 5 (13.5%) |
| selO | 7 (16.3%) | 9 (27.3%) | 5 (13.5%) |
| selN | 9 (20.9%) | 10 (30.3%) | 9 (24.3%) |
| selU | 7 (16.3%) | 9 (27.3%) | 3 (8.1%) |
| selX | 23 (53.5%) | 22 (66.7%) | 15 (40.5%) |
| tstH | 1 (2.3%) | 2 (6.1%) | 2 (5.4%) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Blicharz, L.; Żochowski, M.; Szymanek-Majchrzak, K.; Czuwara, J.; Goldust, M.; Skowroński, K.; Młynarczyk, G.; Olszewska, M.; Samochocki, Z.; Rudnicka, L. Enterotoxin Gene Cluster and selX Are Associated with Atopic Dermatitis Severity—A Cross-Sectional Molecular Study of Staphylococcus aureus Superantigens. Cells 2022, 11, 3921. https://doi.org/10.3390/cells11233921
Blicharz L, Żochowski M, Szymanek-Majchrzak K, Czuwara J, Goldust M, Skowroński K, Młynarczyk G, Olszewska M, Samochocki Z, Rudnicka L. Enterotoxin Gene Cluster and selX Are Associated with Atopic Dermatitis Severity—A Cross-Sectional Molecular Study of Staphylococcus aureus Superantigens. Cells. 2022; 11(23):3921. https://doi.org/10.3390/cells11233921
Chicago/Turabian StyleBlicharz, Leszek, Maciej Żochowski, Ksenia Szymanek-Majchrzak, Joanna Czuwara, Mohamad Goldust, Krzysztof Skowroński, Grażyna Młynarczyk, Małgorzata Olszewska, Zbigniew Samochocki, and Lidia Rudnicka. 2022. "Enterotoxin Gene Cluster and selX Are Associated with Atopic Dermatitis Severity—A Cross-Sectional Molecular Study of Staphylococcus aureus Superantigens" Cells 11, no. 23: 3921. https://doi.org/10.3390/cells11233921