
Insulin-like Growth Factor Binding Protein 3 Increases Mouse Preimplantation Embryo Cleavage Rate by Activation of IGF1R and EGFR Independent of IGF1 Signalling


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Media and Chemical Preparation for Embryo Collection and Culture
2.2. Embryo Collection and Culture
2.3. Western Blotting
2.4. Immunofluorescent Staining of Oocytes and Embryos
2.5. Statistical Analysis
3. Results
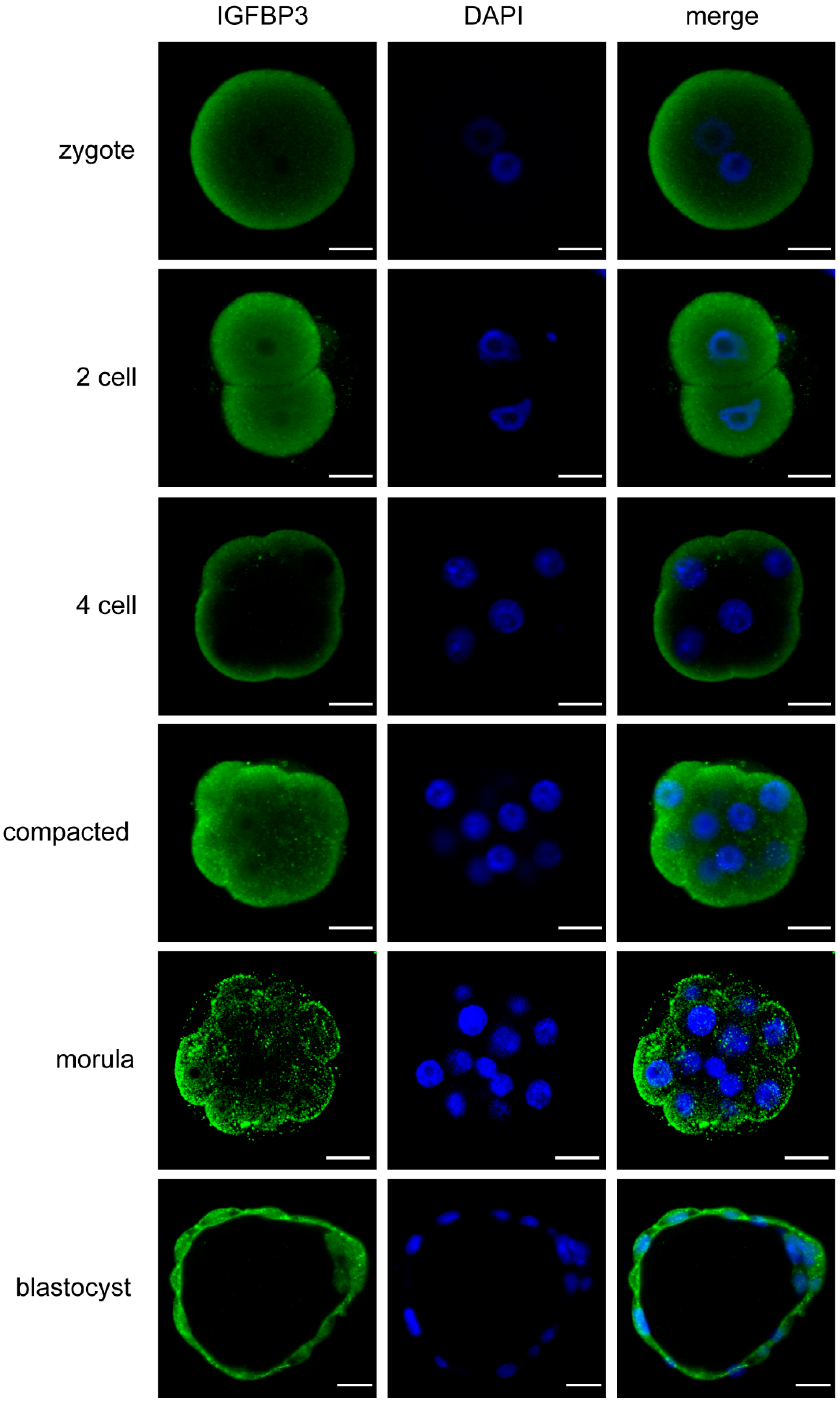
3.1. IGFBP3 Is Expressed across All Stages of Mouse Preimplantation Embryo Development
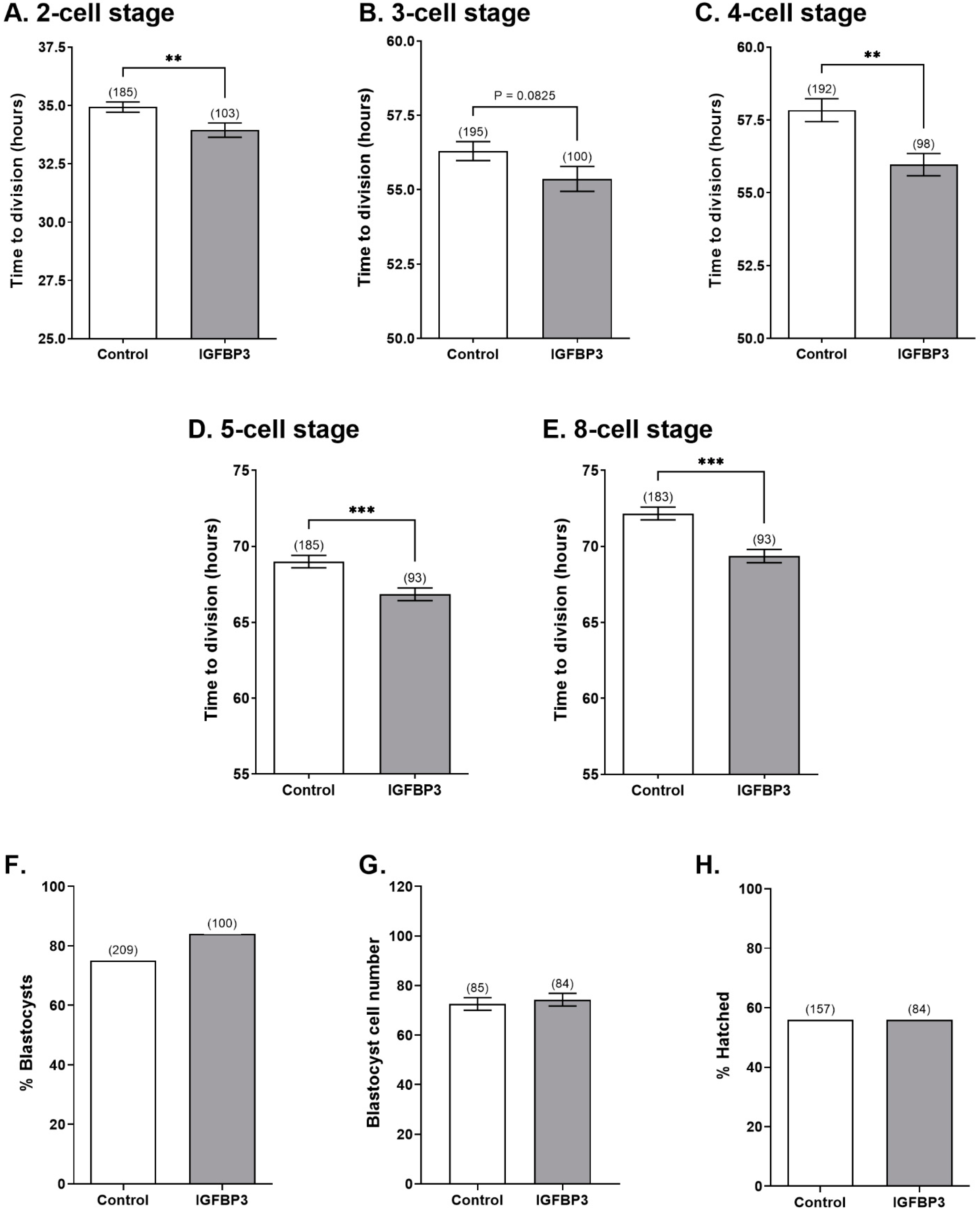
3.2. IGFBP3 Increases the Developmental Rate of Preimplantation Embryos In Vitro
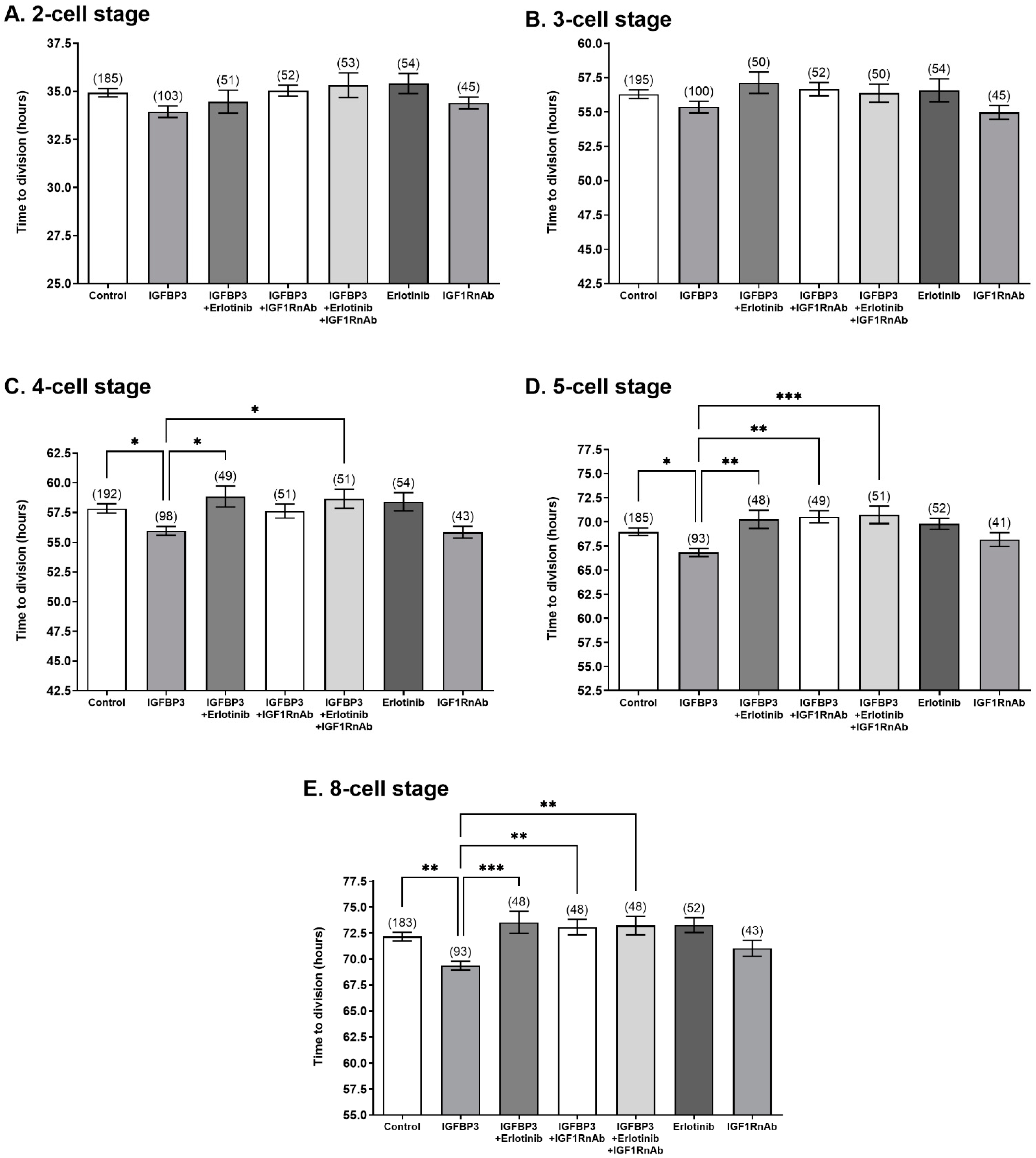
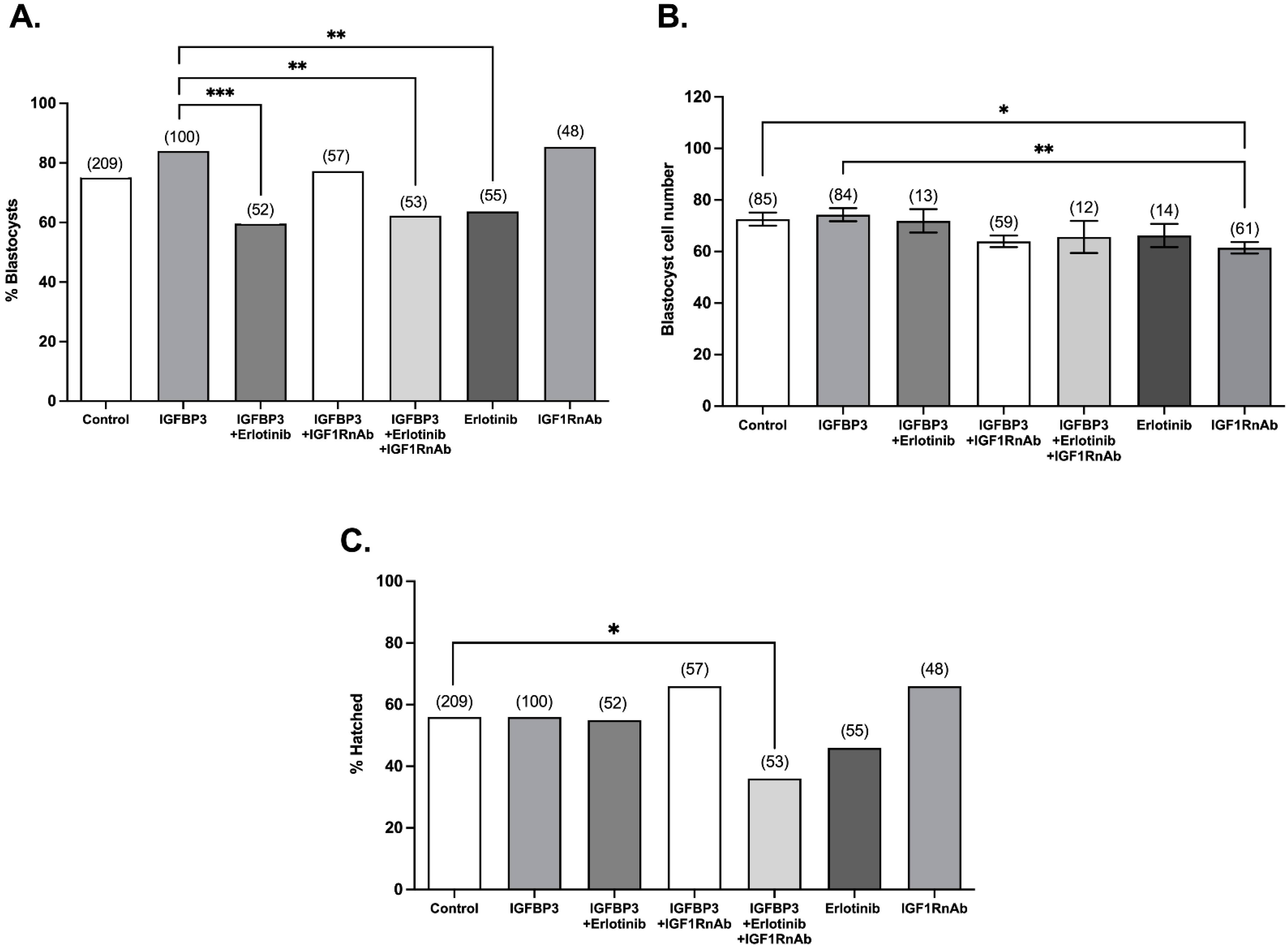
3.3. The IGF1 and EGF Receptors Are Required for Stimulatory IGFBP3 Signalling in the Preimplantation Embryo
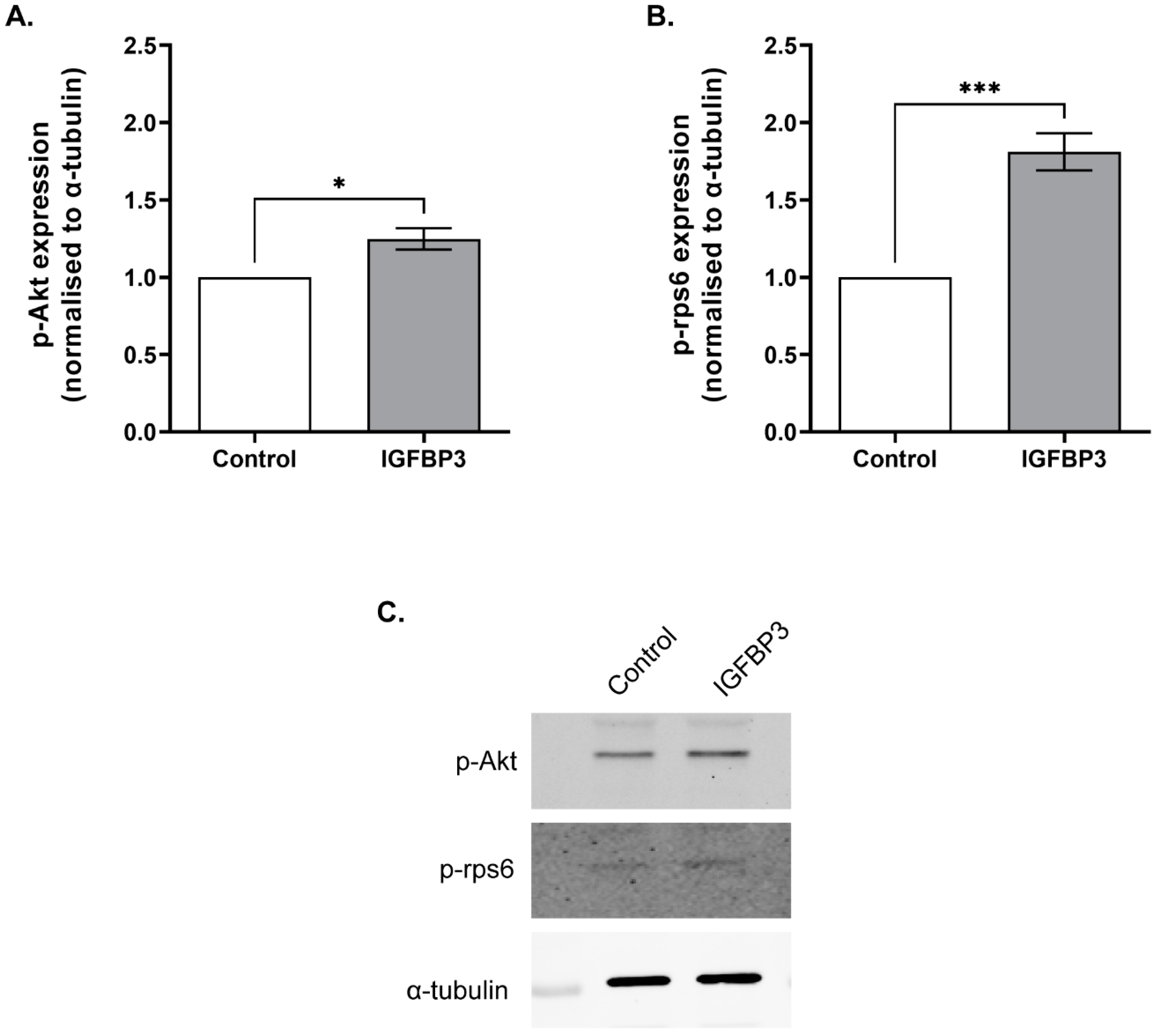
3.4. Short Term IGFBP3 Treatment Increases Phosphorylation of Akt and rps6
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Green, C.J.; Fraser, S.T.; Day, M.L. Insulin-like Growth Factor 1 Increases Apical Fibronectin in Blastocysts to Increase Blastocyst Attachment to Endometrial Epithelial Cells In Vitro. Hum. Reprod. 2014, 30, 284–298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Firth, S.M.; Baxter, R.C. Cellular Actions of the Insulin-like Growth Factor Binding Proteins. Endocr. Rev. 2002, 23, 824–854. [Google Scholar] [CrossRef]
- Mohan, S.; Baylink, D.J. IGF-Binding Proteins Are Multifunctional and Act via IGF-Dependent and -Independent Mechanisms. J. Endocrinol. 2002, 175, 19–31. [Google Scholar] [CrossRef] [Green Version]
- Hahnel, A.; Schultz, G.A. Insulin-like Growth Factor Binding Proteins Are Transcribed by Preimplantation Mouse Embryos. Endocrinology 1994, 134, 1956–1959. [Google Scholar] [CrossRef] [Green Version]
- Lai, Y.M.; Wang, H.S.; Lee, C.L.; Lee, J.D.; Huang, H.Y.; Chang, F.H.; Lee, J.F.; Soong, Y.K. Insulin-like Growth Factor-Binding Proteins Produced by Vero Cells, Human Oviductal Cells and Human Endometrial Cells, and the Role of Insulin-like Growth Factor-Binding Protein-3 in Mouse Embryo Co-Culture Systems. Hum. Reprod. 1996, 11, 1281–1286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, H.C.; He, Z.Y.; Tang, Y.X.; Mele, C.A.; Veeck, L.L.; Davis, O.; Rosenwaks, Z. Simultaneous Detection of Multiple Gene Expression in Mouse and Human Individual Preimplantation Embryos. Fertil. Steril. 1997, 67, 733–741. [Google Scholar] [CrossRef] [PubMed]
- Sawai, K.; Kageyama, S.; Moriyasu, S.; Hirayama, H.; Minamihashi, A.; Onoe, S. Analysis of MRNA Transcripts for Insulin-like Growth Factor Receptors and Binding Proteins in Bovine Embryos Derived from Somatic Cell Nuclear Transfer. Cloning Stem Cells 2005, 7, 189–198. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.C.; Mele, C.A.; Noyes, N.; Rosenwaks, Z. Endometrial Secretory Proteins Enhance Early Embryo Development. J. Assist. Reprod. Genet. 1994, 11, 217–224. [Google Scholar] [CrossRef]
- Liu, H.C.; Mele, C.; Catz, D.; Noyes, N.; Rosenwaks, Z. Production of Insulin-like Growth Factor Binding Proteins (IGFBPs) by Human Endometrial Stromal Cell Is Stimulated by the Presence of Embryos. J. Assist. Reprod. Genet. 1995, 12, 78–87. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.H.; Chang, C.L.; Wu, H.M.; Chiu, Y.M.; Chen, C.K.; Wang, H.S. Insulin-like Growth Factor-II (IGF-II), IGF-Binding Protein-3 (IGFBP-3), and IGFBP-4 in Follicular Fluid Are Associated with Oocyte Maturation and Embryo Development. Fertil. Steril. 2006, 86, 1392–1401. [Google Scholar] [CrossRef] [PubMed]
- Cerro, J.A.; Pintar, J.E. Insulin-like Growth Factor Binding Protein Gene Expression in the Pregnant Rat Uterus and Placenta. Dev. Biol. 1997, 184, 278–295. [Google Scholar] [CrossRef] [Green Version]
- Luo, J.; Zhu, H.; Chang, H.; Lin, Y.; Yang, J.; Leung, P.C.K. The Regulation of IGFBP3 by BMP2 Has a Role in Human Endometrial Remodeling. FASEB J. 2020, 34, 15462–15479. [Google Scholar] [CrossRef]
- Zhao, H.; Klausen, C.; Zhu, H.; Chang, H.; Li, Y.; Leung, P.C.K. Bone Morphogenetic Protein 2 Promotes Human Trophoblast Cell Invasion and Endothelial-like Tube Formation through ID1-mediated Upregulation of IGF Binding Protein-3. FASEB J. 2020, 34, 3151–3164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baxter, R.C. Signalling Pathways Involved in Antiproliferative Effects of IGFBP-3: A Review. Mol. Pathol. 2001, 54, 145–148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fanayan, S.; Firth, S.M.; Baxter, R.C. Signaling through the Smad Pathway by Insulin-like Growth Factor-Binding Protein-3 in Breast Cancer Cells. Relationship to Transforming Growth Factor-Beta 1 Signaling. J. Biol. Chem. 2002, 277, 7255–7261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fanayan, S.; Firth, S.M.; Butt, A.J.; Baxter, R.C. Growth Inhibition by Insulin-like Growth Factor-Binding Protein-3 in T47D Breast Cancer Cells Requires Transforming Growth Factor-β (TGF-β) and the Type II TGF-β Receptor. J. Biol. Chem. 2000, 275, 39146–39151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, K.W.; Ma, L.; Yan, X.; Liu, B.; Zhang, X.K.; Cohen, P. Rapid Apoptosis Induction by IGFBP-3 Involves an Insulin-like Growth Factor-Independent Nucleomitochondrial Translocation of RXRalpha/Nur77. J. Biol. Chem. 2005, 280, 16942–16948. [Google Scholar] [CrossRef] [Green Version]
- Liu, B.; Lee, H.Y.; Weinzimer, S.A.; Powell, D.R.; Clifford, J.L.; Kurie, J.M.; Cohen, P. Direct Functional Interactions between Insulin-like Growth Factor-Binding Protein-3 and Retinoid X Receptor-Alpha Regulate Transcriptional Signaling and Apoptosis. J. Biol. Chem. 2000, 275, 33607–33613. [Google Scholar] [CrossRef] [Green Version]
- Schedlich, L.J.; O’Han, M.K.; Leong, G.M.; Baxter, R.C. Insulin-like Growth Factor Binding Protein-3 Prevents Retinoid Receptor Heterodimerization: Implications for Retinoic Acid-Sensitivity in Human Breast Cancer Cells. Biochem. Biophys. Res. Commun. 2004, 314, 83–88. [Google Scholar] [CrossRef]
- Schedlich, L.J.; Page, S.L.L.; Firth, S.M.; Briggs, L.J.; Jans, D.A.; Baxter, R.C. Nuclear Import of Insulin-like Growth Factor-Binding Protein-3 and -5 Is Mediated by the Importin Beta Subunit. J. Biol. Chem. 2000, 275, 23462–23470. [Google Scholar] [CrossRef]
- Schedlich, L.J.; Graham, L.D.; O’Han, M.K.; Muthukaruppan, A.; Yan, X.; Firth, S.M.; Baxter, R.C. Molecular Basis of the Interaction between IGFBP-3 and Retinoid X Receptor: Role in Modulation of RAR-Signaling. Arch. Biochem. Biophys. 2007, 465, 359–369. [Google Scholar] [CrossRef]
- Agostini-Dreyer, A.; Jetzt, A.E.; Stires, H.; Cohick, W.S. Endogenous IGFBP-3 Mediates Intrinsic Apoptosis Through Modulation of Nur77 Phosphorylation and Nuclear Export. Endocrinology 2015, 156, 4141–4151. [Google Scholar] [CrossRef] [Green Version]
- Krishnan, A.V.; Peehl, D.M.; Feldman, D. Inhibition of Prostate Cancer Growth by Vitamin D: Regulation of Target Gene Expression. J. Cell. Biochem. 2003, 88, 363–371. [Google Scholar] [CrossRef]
- Jia, Q.; Xiao-li, M.; Xin, W.; Hong, C.; Bing-ren, H. Insulin-like Growth Factor Binding Protein-3 Interacts with the Thyroid Hormone Receptor A1 and Modulates Transcription of Thyroid Hormone Responsive Gene. Zhongguo Yi Xue Ke Xue Yuan Xue Bao Acta Acad Med. Sin. 2011, 33, 156–161. [Google Scholar] [CrossRef]
- Grkovic, S.; O’Reilly, V.C.; Han, S.; Hong, M.; Baxter, R.C.; Firth, S.M. IGFBP-3 Binds GRP78, Stimulates Autophagy and Promotes the Survival of Breast Cancer Cells Exposed to Adverse Microenvironments. Oncogene 2013, 32, 2412–2420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, M.Z.; Marzec, K.A.; Martin, J.L.; Baxter, R.C. The Role of Insulin-like Growth Factor Binding Protein-3 in the Breast Cancer Cell Response to DNA-Damaging Agents. Oncogene 2014, 33, 85–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, J.L.; de Silva, H.C.; Lin, M.Z.; Scott, C.D.; Baxter, R.C. Inhibition of Insulin-like Growth Factor-Binding Protein-3 Signaling through Sphingosine Kinase-1 Sensitizes Triple-Negative Breast Cancer Cells to EGF Receptor Blockade. Mol. Cancer Ther. 2014, 13, 316–328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morris, M.B.; Ozsoy, S.; Zada, M.; Zada, M.; Zamfirescu, R.C.; Todorova, M.G.; Day, M.L. Selected Amino Acids Promote Mouse Pre-Implantation Embryo Development in a Growth Factor-Like Manner. Front. Physiol. 2020, 11, 140. [Google Scholar] [CrossRef] [PubMed]
- Martin, J.L.; Lin, M.Z.; McGowan, E.M.; Baxter, R.C. Potentiation of Growth Factor Signaling by Insulin-like Growth Factor-Binding Protein-3 in Breast Epithelial Cells Requires Sphingosine Kinase Activity. J. Biol. Chem. 2009, 284, 25542–25552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arcaro, A. Targeting the Insulin-like Growth Factor-1 Receptor in Human Cancer. Front. Pharmacol. 2013, 4, 30. [Google Scholar] [CrossRef] [PubMed]
- Green, C.J.; Day, M.L. Insulin-like Growth Factor 1 Acts as an Autocrine Factor to Improve Early Embryogenesis in Vitro. Int. J. Dev. Biol. 2013, 57, 837–844. [Google Scholar] [CrossRef] [Green Version]
- Grill, C.J.; Sivaprasad, U.; Cohick, W.S. Constitutive Expression of IGF-Binding Protein-3 by Mammary Epithelial Cells Alters Signaling through Akt and P70S6 Kinase. J. Mol. Endocrinol. 2002, 29, 153–162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Summers, M.C.; Biggers, J.D. Chemically Defined Media and the Culture of Mammalian Preimplantation Embryos: Historical Perspective and Current Issues. Hum. Reprod. Update 2003, 9, 557–582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bowman, P.; McLaren, A. Cleavage Rate of Mouse Embryos In Vivo and In Vitro. J. Embryol. Exp. Morphol. 1970, 24, 203–207. [Google Scholar] [CrossRef] [PubMed]
- Harlow, G.M.; Quinn, P. Development of Preimplantation Mouse Embryos in Vivo and in Vitro. Aust. J. Biol. Sci. 1982, 35, 187–193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Titone, R.; Zhu, M.; Robertson, D.M. Mutual Regulation between IGF-1R and IGFBP-3 in Human Corneal Epithelial Cells. J. Cell. Physiol. 2019, 234, 1426–1441. [Google Scholar] [CrossRef] [PubMed]
- Shrivastav, S.V.; Bhardwaj, A.; Pathak, K.A.; Shrivastav, A. Insulin-Like Growth Factor Binding Protein-3 (IGFBP-3): Unraveling the Role in Mediating IGF-Independent Effects within the Cell. Front. Cell Dev. Biol. 2020, 8, 286. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Zhang, X.; Gao, J.; Palombo, M.; Gao, D.; Chen, P.; Sinko, P. Core Functional Sequence of C-Terminal GAG-Binding Domain Directs Cellular Uptake of IGFBP-3-Derived Peptides. Protein Pept. Lett. 2013, 21, 124–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fowlkes, J.L.; Serra, D.M. Characterization of Glycosaminoglycan-Binding Domains Present in Insulin-like Growth Factor-Binding Protein-3. J. Biol. Chem. 1996, 271, 14676–14679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goda, N.; Tenno, T.; Inomata, K.; Shirakawa, M.; Tanaka, T.; Hiroaki, H. Intracellular Protein Delivery Activity of Peptides Derived from Insulin-like Growth Factor Binding Proteins 3 and 5. Exp. Cell Res. 2008, 314, 2352–2361. [Google Scholar] [CrossRef]
- Lee, K.-W.; Liu, B.; Ma, L.; Li, H.; Bang, P.; Koeffler, H.P.; Cohen, P. Cellular Internalization of Insulin-like Growth Factor Binding Protein-3 Distinct Endocytic Pathways Facilitate Re-Uptake and Nuclear Localization. J. Biol. Chem. 2004, 279, 469–476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Micutkova, L.; Hermann, M.; Offterdinger, M.; Hess, M.W.; Matscheski, A.; Pircher, H.; Muck, C.; Ebner, H.L.; Laich, A.; Ferrando-May, E.; et al. Analysis of the Cellular Uptake and Nuclear Delivery of Insulin-like Growth Factor Binding Protein-3 in Human Osteosarcoma Cells. Int. J. Cancer 2012, 130, 1544–1557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Silva, H.C.; Lin, M.Z.; Phillips, L.; Martin, J.L.; Baxter, R.C. IGFBP-3 Interacts with NONO and SFPQ in PARP-Dependent DNA Damage Repair in Triple-Negative Breast Cancer. Cell. Mol. Life Sci. 2019, 76, 2015–2030. [Google Scholar] [CrossRef]
- Baxter, R.C. Nuclear Actions of Insulin-like Growth Factor Binding Protein-3. Gene 2015, 569, 7–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhardwaj, A.; Pathak, K.A.; Shrivastav, A.; Shrivastav, S.V. Insulin-Like Growth Factor Binding Protein-3 Binds to Histone 3. Int. J. Mol. Sci. 2021, 22, 407. [Google Scholar] [CrossRef]
- Kuemmerle, J.F.; Murthy, K.S.; Bowers, J.G. IGFBP-3 Activates TGF-β Receptors and Directly Inhibits Growth in Human Intestinal Smooth Muscle Cells. Am. J. Physiol.-Gastrointest. Liver Physiol. 2004, 287, G795–G802. [Google Scholar] [CrossRef]
- Ingermann, A.R.; Yang, Y.F.; Han, J.; Mikami, A.; Garza, A.E.; Mohanraj, L.; Fan, L.; Idowu, M.; Ware, J.L.; Kim, H.S.; et al. Identification of a Novel Cell Death Receptor Mediating IGFBP-3-Induced Anti-Tumor Effects in Breast and Prostate Cancer. J. Biol. Chem. 2010, 285, 30233–30246. [Google Scholar] [CrossRef] [Green Version]
- Jorissen, R.N.; Walker, F.; Pouliot, N.; Garrett, T.P.; Ward, C.W.; Burgess, A.W.W. Epidermal Growth Factor Receptor: Mechanisms of Activation and Signalling. Exp. Cell Res. 2003, 284, 31–53. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.S.L.; Cui, W. Proliferation, Survival and Metabolism: The Role of PI3K/AKT/MTOR Signalling in Pluripotency and Cell Fate Determination. Development 2016, 143, 3050–3060. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yano, T.; Ferlito, M.; Aponte, A.; Kuno, A.; Miura, T.; Murphy, E.; Steenbergen, C. Pivotal Role of MTORC2 and Involvement of Ribosomal Protein S6 in Cardioprotective Signaling. Circ. Res. 2014, 114, 1268–1280. [Google Scholar] [CrossRef]
- Sheikh, M.E.; Mesalam, A.; Mesalam, A.A.; Idrees, M.; Lee, K.-L.; Kong, I.-K. Melatonin Abrogates the Anti-Developmental Effect of the AKT Inhibitor SH6 in Bovine Oocytes and Embryos. Int. J. Mol. Sci. 2019, 20, 2956. [Google Scholar] [CrossRef] [Green Version]
- Panić, L.; Tamarut, S.; Sticker-Jantscheff, M.; Barkić, M.; Solter, D.; Uzelac, M.; Grabušić, K.; Volarević, S. Ribosomal Protein S6 Gene Haploinsufficiency Is Associated with Activation of a P53-Dependent Checkpoint during Gastrulation. Mol. Cell. Biol. 2006, 26, 8880–8891. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Granata, R.; Trovato, L.; Garbarino, G.; Taliano, M.; Ponti, R.; Sala, G.; Ghidoni, R.; Ghigo, E. Dual Effects of IGFBP-3 on Endothelial Cell Apoptosis and Survival: Involvement of the Sphingolipid Signaling Pathways. FASEB J. 2004, 18, 1456–1458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yaqoob, U.; Luo, F.; Greuter, T.; Sakrikar, N.J.; Sehrawat, T.S.; Lu, J.; Hu, X.; Gao, J.; Kostallari, E.; Chen, J.; et al. GIPC-Regulated IGFBP-3 Promotes HSC Migration In Vitro and Portal Hypertension In Vivo through a Β1-Integrin Pathway. Cell. Mol. Gastroenterol. Hepatol. 2020, 10, 545–559. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, S.E.; Milstien, S.; Spiegel, S. Autocrine and Paracrine Roles of Sphingosine-1-Phosphate. Trends Endocrinol. Metab. 2007, 18, 300–307. [Google Scholar] [CrossRef] [PubMed]
- Herr, D.R.; Lee, C.W.; Wang, W.; Ware, A.; Rivera, R.; Chun, J. Sphingosine 1-Phosphate Receptors Are Essential Mediators of Eyelid Closure during Embryonic Development. J. Biol. Chem. 2013, 288, 29882–29889. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daub, H.; Weiss, F.U.; Wallasch, C.; Ullrich, A. Role of Transactivation of the EGF Receptor in Signalling by G-Protein-Coupled Receptors. Nature 1996, 379, 557–560. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Green, C.J.; Span, M.; Rayhanna, M.H.; Perera, M.; Day, M.L. Insulin-like Growth Factor Binding Protein 3 Increases Mouse Preimplantation Embryo Cleavage Rate by Activation of IGF1R and EGFR Independent of IGF1 Signalling. Cells 2022, 11, 3762. https://doi.org/10.3390/cells11233762
Green CJ, Span M, Rayhanna MH, Perera M, Day ML. Insulin-like Growth Factor Binding Protein 3 Increases Mouse Preimplantation Embryo Cleavage Rate by Activation of IGF1R and EGFR Independent of IGF1 Signalling. Cells. 2022; 11(23):3762. https://doi.org/10.3390/cells11233762
Chicago/Turabian StyleGreen, Charmaine J., Miriam Span, Monique H. Rayhanna, Marisa Perera, and Margot L. Day. 2022. "Insulin-like Growth Factor Binding Protein 3 Increases Mouse Preimplantation Embryo Cleavage Rate by Activation of IGF1R and EGFR Independent of IGF1 Signalling" Cells 11, no. 23: 3762. https://doi.org/10.3390/cells11233762