
Binding and Detoxification of Insecticides by Potentially Probiotic Lactic Acid Bacteria Isolated from Honeybee (Apis mellifera L.) Environment—An In Vitro Study


Abstract
:
1. Introduction
2. Materials and Methods
2.1. Culture Vessels, Chemicals, and Other Materials
2.2. Lactic Acid Bacteria Strains and Growth Conditions
2.3. Pesticide Stocks Preparation and Storage
2.4. Effect of Pesticides on the Growth of LAB
2.5. Pesticide Binding Assay
2.5.1. Whole Live LAB Cells
2.5.2. Thermally Inactivated LAB Cells
2.5.3. Intracellular Extracts (ICEs) and Membrane Extracts (MEs)
2.6. HPLC Analysis
2.7. Cell Line Cultures (Sf-9, Caco-2, and IEC-6)
2.8. MTT Assay
2.9. Single-Cell Gel Electrophoresis Assay (Comet Assay)
2.10. Statistical Analysis
3. Results and Discussion
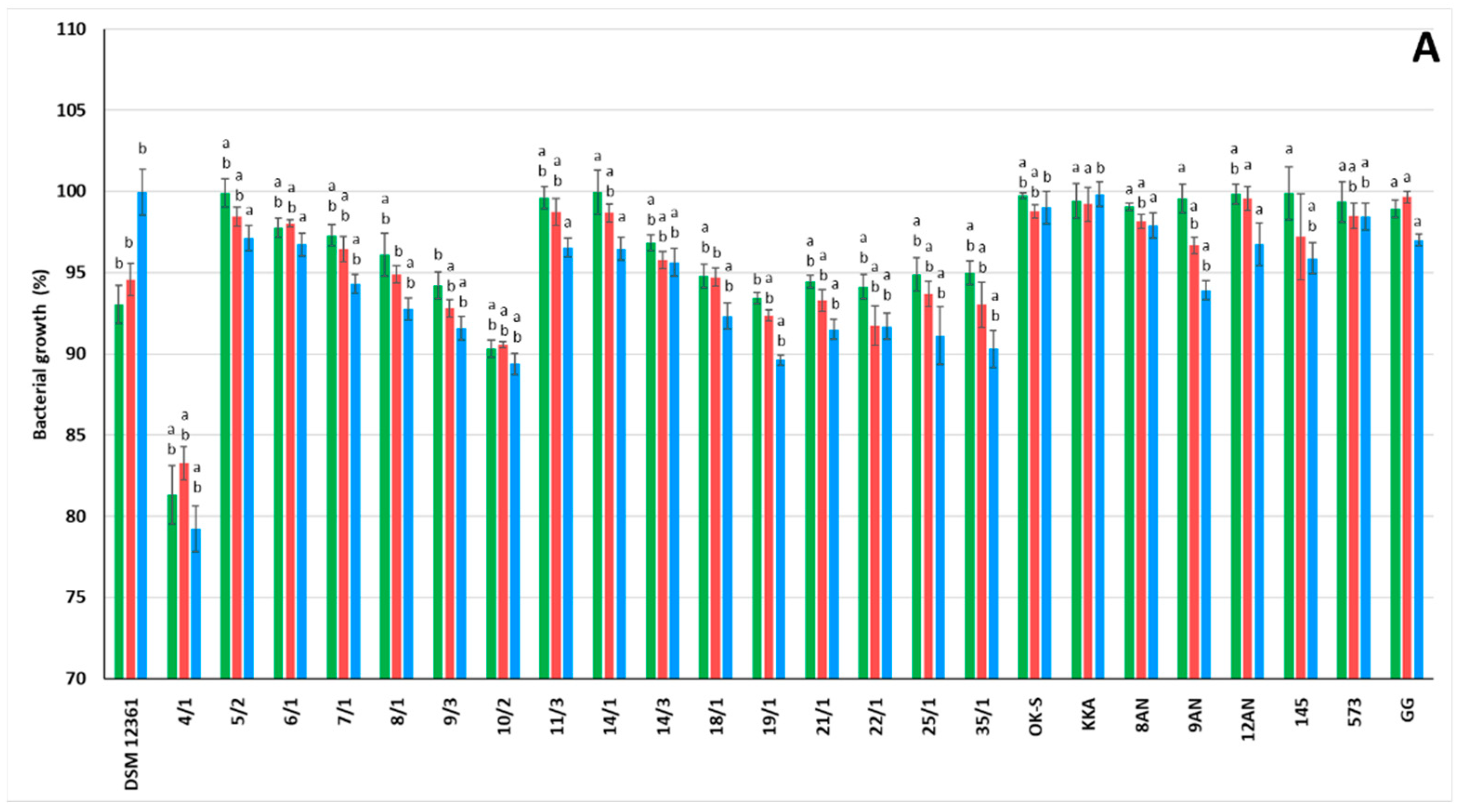
3.1. LAB Growth in the Presence of Pesticides
3.2. Binding of Pesticides to the Cell Wall of LAB
3.3. Detoxification of Pesticides by LAB
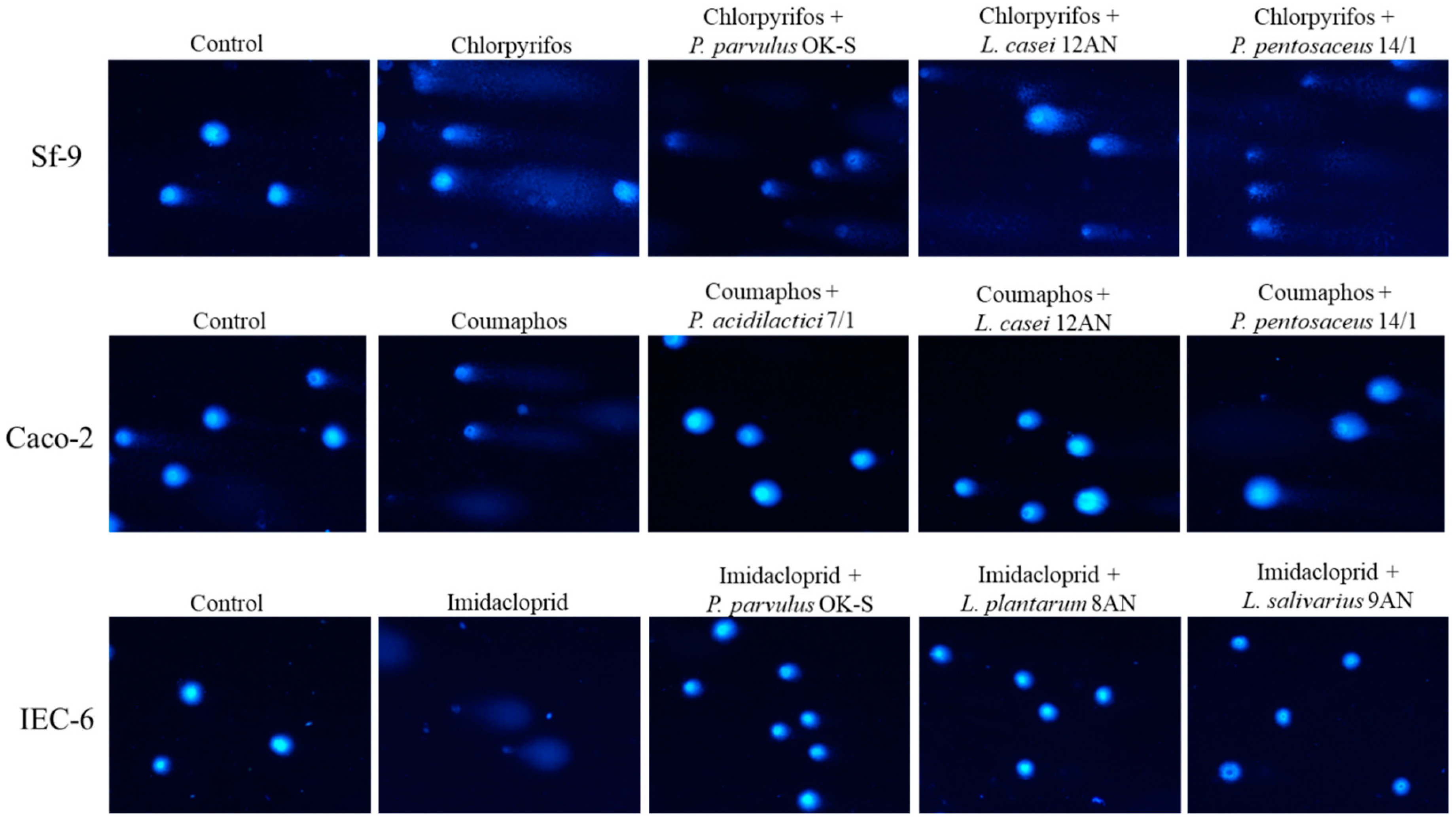
3.3.1. Decrease in cytotoxicity
3.3.2. Decrease in Genotoxicity
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Abhilash, P.C.; Singh, N. Pesticide use and application: An Indian scenario. J. Hazard. Mater. 2009, 165, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Fries, G.F.; Marrow, G.S.; Gordon, C.H. Comparative excretion and retention of DDT analogs by Dairy Cows. J. Dairy Sci. 1969, 52, 1800–1805. [Google Scholar] [CrossRef] [PubMed]
- Osman, K.A. Pesticides and human health. Pesticides in the Modern World—Effects of Pesticides Exposure; IntechOpen: London, UK, 2011. [Google Scholar] [CrossRef] [Green Version]
- Aktar, W.; Sengupta, D.; Chowdhury, A. Impact of pesticides use in agriculture: Their benefits and Hazards. Interdiscip. Toxicol. 2009, 2, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fenner-Crisp, P.A. Risk assessment and risk management. In Hayes’ Handbook of Pesticide Toxicology; Elsevier: Amsterdam, The Netherlands, 2010; pp. 1371–1380. [Google Scholar] [CrossRef]
- Feazel-Orr, H.K.; Catalfamo, K.M.; Brewster, C.C.; Fell, R.D.; Anderson, T.D.; Traver, B.E. Effects of Pesticide Treatments on Nutrient Levels in Worker Honey Bees (Apis mellifera). Insects 2016, 7, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, J.Y.; Anelli, C.M.; Sheppard, W.S. Sub-lethal effects of pesticide residues in brood comb on worker honey bee (Apis mellifera) development and longevity. PLoS ONE 2011, 6, e14720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wahl, O.; Ulm, K. Influence of pollen feeding and physiological condition on pesticide sensitivity of the honey bee Apis mellifera carnica. Oecologia 1983, 59, 106–128. [Google Scholar] [CrossRef] [PubMed]
- Claudianos, C.; Ranson, H.; Johnson, R.M.; Biswas, S.; Schuler, M.A.; Berenbaum, M.R.; Feyereisen, R.; Oakeshott, J.G. A deficit of detoxification enzymes: Pesticide sensitivity and environmental response in the honeybee. Insect Mol. Biol. 2006, 15, 615–636. [Google Scholar] [CrossRef] [Green Version]
- Kumar, G.; Singh, S.; Pramod Kodigenahalli Nagarajaiah, R. Detailed review on pesticidal toxicity to honey bees and its management. In Modern Beekeeping—Bases for Sustainable Production; IntechOpen: London, UK, 2020. [Google Scholar] [CrossRef]
- Martelli, F.; Zhongyuan, Z.; Wang, J.; Wong, C.-O.; Karagas, N.E.; Roessner, U.; Rupasinghe, T.; Venkatachalam, K.; Perry, T.; Bellen, H.J.; et al. Low doses of the neonicotinoid insecticide imidacloprid induce ROS triggering neurological and metabolic impairments in Drosophila. Proc. Natl. Acad. Sci. USA 2020, 117, 25840–25850. [Google Scholar] [CrossRef]
- Tan, J.; Galligan, J.J.; Hollingworth, R.M. Agonist actions of neonicotinoids on nicotinic acetylcholine receptors expressed by cockroach neurons. NeuroToxicology 2007, 28, 829–842. [Google Scholar] [CrossRef]
- Tosi, S.; Nieh, J.C.; Sgolastra, F.; Cabbri, R.; Medrzycki, P. Neonicotinoid pesticides and nutritional stress synergistically reduce survival in honey bees. Proc. R. Soc. B. 2017, 284, 20171711. [Google Scholar] [CrossRef] [Green Version]
- Laycock, I.; Lenthall, K.M.; Barratt, A.T.; Cresswell, J.E. Effects of imidacloprid, a neonicotinoid pesticide, on reproduction in worker bumble bees (Bombus terrestris). Ecotoxicology 2012, 21, 1937–1945. [Google Scholar] [CrossRef] [PubMed]
- Urlacher, E.; Monchanin, C.; Rivière, C.; Richard, F.-J.; Lombardi, C.; Michelsen-Heath, S.; Hageman, K.; Mercer, A. Measurements of chlorpyrifos levels in forager bees and comparison with levels that disrupt honey bee odor-mediated learning under laboratory conditions. J. Chem. Ecol. 2016, 42, 127–138. [Google Scholar] [CrossRef] [PubMed]
- Cutler, G.C.; Purdy, J.; Giesy, J.P.; Solomon, K.R. Risk to pollinators from the use of chlorpyrifos in the United States. In Ecological Risk Assessment for Chlorpyrifos in Terrestrial and Aquatic Systems in the United States; Springer: Berlin/Heidelberg, Germany, 2014; pp. 219–265. [Google Scholar] [CrossRef] [Green Version]
- Darko, G.; Addai Tabi, J.; Adjaloo, M.K.; Borquaye, L.S. Pesticide residues in honey from the major honey producing forest belts in Ghana. J. Environ. Public Health 2017, 2017, 7957431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hites, R.A. The rise and fall of chlorpyrifos in the United States. Environ. Sci. Technol. 2021, 55, 1354–1358. [Google Scholar] [CrossRef]
- Ola-Davies, O.E.; Azeez, O.I.; Oyagbemi, A.A.; Abatan, M.O. Acute coumaphos organophosphate exposure in the domestic dogs: Its implication on haematology and liver functions. Int. J. Vet. Sci. Med. 2018, 6, 103–112. [Google Scholar] [CrossRef]
- Berry, J.A.; Hood, W.M.; Pietravalle, S.; Delaplane, K.S. Field-level sublethal effects of approved bee hive chemicals on honey bees (Apis mellifera L). PLoS ONE 2013, 8, e76536. [Google Scholar] [CrossRef]
- Rouzé, R.; Moné, A.; Delbac, F.; Belzunces, L.; Blot, N. The honeybee gut microbiota is altered after chronic exposure to different families of insecticides and infection by Nosema ceranae. Microbes Environ. 2019, 34, 226–233. [Google Scholar] [CrossRef] [Green Version]
- Wang, K.; Chen, H.; Fan, R.-L.; Lin, Z.-G.; Niu, Q.-S.; Wang, Z.; Ji, T. Effect of Carbendazim on Honey Bee Health: Assessment of survival, pollen consumption, and gut microbiome composition. Ecotoxicol. Environ. Saf. 2022, 239, 113648. [Google Scholar] [CrossRef]
- Balbuena, S.; Castelli, L.; Zunino, P.; Antúnez, K. Effect of chronic exposure to sublethal doses of imidacloprid and Nosema ceranae on immunity, gut microbiota, and survival of Africanized honey bees. Microb. Ecol. 2022, 1–13. [Google Scholar] [CrossRef]
- Kakumanu, M.L.; Reeves, A.M.; Anderson, T.D.; Rodrigues, R.R.; Williams, M.A. Honey bee gut microbiome is altered by in-hive pesticide exposures. Front. Microbiol. 2016, 7, 1255. [Google Scholar] [CrossRef] [Green Version]
- Motta, E.V.; Raymann, K.; Moran, N.A. Glyphosate perturbs the gut microbiota of Honey Bees. Proc. Natl. Acad. Sci. USA 2018, 115, 10305–10310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cuesta-Maté, A.; Renelies-Hamilton, J.; Kryger, P.; Jensen, A.B.; Sinotte, V.M.; Poulsen, M. Resistance and vulnerability of honeybee (Apis mellifera) gut bacteria to commonly used pesticides. Front. Microbiol. 2021, 12, 2428. [Google Scholar] [CrossRef] [PubMed]
- Mathialagan, M.; Thangaraj Edward, Y.S.J.; David, P.M.M.; Senthilkumar, M.; Srinivasan, M.R.; Mohankumar, S. Isolation, characterization and identification of probiotic lactic acid bacteria (LAB) from honey bees. Int. J. Curr. Microbiol. Appl. Sci. 2018, 7, 894–906. [Google Scholar] [CrossRef]
- Nowak, A.; Szczuka, D.; Górczyńska, A.; Motyl, I.; Kręgiel, D. Characterization of Apis mellifera Gastrointestinal Microbiota and Lactic Acid Bacteria for Honeybee Protection—A Review. Cells 2021, 10, 701. [Google Scholar] [CrossRef] [PubMed]
- Pop, O.L.; Suharoschi, R.; Gabbianelli, R. Biodetoxification and protective properties of probiotics. Microorganisms 2022, 10, 1278. [Google Scholar] [CrossRef]
- Petrova, P.; Arsov, A.; Tsvetanova, F.; Parvanova-Mancheva, T.; Vasileva, E.; Tsigoriyna, L.; Petrov, K. The complex role of lactic acid bacteria in food detoxification. Nutrients 2022, 14, 2038. [Google Scholar] [CrossRef]
- Alcántara, C.; Jadán-Piedra, C.; Vélez, D.; Devesa, V.; Zúñiga, M.; Monedero, V. Characterization of the binding capacity of mercurial species in Lactobacillus strains. J. Sci. Food Agric. 2017, 97, 5107–5113. [Google Scholar] [CrossRef] [Green Version]
- Sadiq, F.A.; Yan, B.; Tian, F.; Zhao, J.; Zhang, H.; Chen, W. Lactic acid bacteria as antifungal and anti-mycotoxigenic agents: A comprehensive review. Compr. Rev. Food. Sci. Food Saf. 2019, 18, 1403–1436. [Google Scholar] [CrossRef] [Green Version]
- Nowak, A.; Libudzisz, Z. Ability of probiotic Lactobacillus casei DN 114001 to bind or/and metabolise heterocyclic aromatic amines in vitro. Eur. J. Nutr. 2009, 48, 419–427. [Google Scholar] [CrossRef]
- Nowak, A.; Czyżowska, A.; Stańczyk, M. Protective activity of probiotic bacteria against2-amino-3-methyl-3H-imidazo[4,5-f]quinoline (IQ) and 2-amino-1-methyl-6-phenyl-1H-imidazo[4,5-b]pyridine (PhIP)—An in vitro study. Food Addit. Contam. A 2015, 32, 1927–1938. [Google Scholar] [CrossRef]
- Yousefi, M.; Khorshidian, N.; Hosseini, H. The ability of probiotic Lactobacillus strains in removal of benzo[a]pyrene: A response surface methodology study. Probiotics Antimicrob. 2021, 14, 464–475. [Google Scholar] [CrossRef] [PubMed]
- Shoukat, S. Potential anti-carcinogenic effect of probiotic and lactic acid bacteria in detoxification of benzo[a]pyrene: A Review. Trends Food Sci. Technol. 2020, 99, 450–459. [Google Scholar] [CrossRef]
- Yousefi, M.; Shariatifar, N.; Tajabadi Ebrahimi, M.; Mortazavian, A.M.; Mohammadi, A.; Khorshidian, N.; Arab, M.; Hosseini, H. In vitro removal of polycyclic aromatic hydrocarbons by lactic acid bacteria. J. Appl. Microbiol. 2019, 126, 954–964. [Google Scholar] [CrossRef] [PubMed]
- Lili, Z.; Junyan, W.; Hongfei, Z.; Baoqing, Z.; Bolin, Z. Detoxification of cancerogenic compounds by lactic acid bacteria strains. Crit. Rev. Food. Sci. Nutr. 2017, 58, 2727–2742. [Google Scholar] [CrossRef] [PubMed]
- Barbasz, A.; Kreczmer, B.; Skórka, M.; Czyżowska, A. Toxicity of pesticides toward human immune cells U-937 and HL-60. J. Environ. Sci. Health B 2020, 55, 719–725. [Google Scholar] [CrossRef]
- Cestonaro, L.V.; Macedo, S.M.; Piton, Y.V.; Garcia, S.C.; Arbo, M.D. Toxic effects of pesticides on cellular and humoral immunity: An overview. Immunopharmacol. Immunotoxicol. 2022, 1–16. [Google Scholar] [CrossRef]
- Kumar, N.; Pathera, A.; Saini, P.; Kumar, M. Harmful effects of pesticides on human health. Ann. Agric. Bio. Res. 2012, 17, 165–168. [Google Scholar]
- Issaragrisil, S.; Chansung, K.; Kaufman, D.W.; Sirijirachai, J.; Thamprasit, T.; Young, N.S. Aplastic anemia in rural Thailand: Its association with grain farming and agricultural pesticide exposure. aplastic anemia study group. Am. J. Public Health 1997, 87, 1551–1554. [Google Scholar] [CrossRef] [Green Version]
- Naughton, S.X.; Terry, A.V. Neurotoxicity in acute and repeated organophosphate exposure. Toxicology 2018, 408, 101–112. [Google Scholar] [CrossRef]
- Nasrollahzadeh, A.; Mokhtari, S.; Khomeiri, M.; Saris, P. Mycotoxin detoxification of food by lactic acid bacteria. Int. J. Food Contam. 2022, 9, 1. [Google Scholar] [CrossRef]
- Chmiel, J.A.; Daisley, B.A.; Pitek, A.P.; Thompson, G.J.; Reid, G. Understanding the effects of sublethal pesticide exposure on honey bees: A role for probiotics as mediators of environmental stress. Front. Ecol. Evol. 2020, 8, 22. [Google Scholar] [CrossRef] [Green Version]
- Leska, A.; Nowak, A.; Motyl, I. Isolation and Some Basic Characteristics of Lactic Acid Bacteria from Honeybee (Apis mellifera L.) Environment—A Preliminary Study. Agriculture 2022, 12, 1562. [Google Scholar] [CrossRef]
- Leska, A.; Nowak, A.; Szulc, J.; Motyl, I.; Czarnecka-Chrebelska, K.H. Antagonistic Activity of Potentially Probiotic Lactic Acid Bacteria against Honeybee (Apis mellifera L.) Pathogens. Pathogens 2022, 11, 1367. [Google Scholar] [CrossRef]
- Lye, H.S.; Balakrishnan, K.; Thiagarajah, K.; Mohd Ismail, N.I.; Ooi, S.Y. Beneficial properties of probiotics. Trop. Life Sci. Res. 2016, 27, 73–90. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.-Y.; Tsen, H.-Y.; Lin, C.-L.; Lin, C.-K.; Chuang, L.-T.; Chen, C.-S.; Chiang, Y.-C. Enhancement of the immune response against Salmonella infection of mice by heat-killed multispecies combinations of lactic acid bacteria. J. Med. Microbiol. 2013, 62, 1657–1664. [Google Scholar] [CrossRef]
- Hafeez, A.; Iqbal, S.; Tawab, I.A.; Bhutto, A.; Uddin, R.; Anwar, F. Liquid chromatographic separation and quantification of imidacloprid in different modes of formulations. Am. Eurasian J. Agric. Env. Sci. 2015, 15, 2381–2386. [Google Scholar] [CrossRef]
- Harishankar, M.K.; Sasikala, C.; Ramya, M. Efficiency of the intestinal bacteria in the degradation of the toxic pesticide, chlorpyrifos. 3 Biotech 2012, 3, 137–142. [Google Scholar] [CrossRef]
- Trinder, M.; McDowell, T.W.; Daisley, B.A.; Ali, S.N.; Leong, H.S.; Sumarah, M.W.; Reid, G. Probiotic Lactobacillus rhamnosus reduces organophosphate pesticide absorption and toxicity to Drosophila melanogaster. Appl. Environ. Microbiol. 2016, 82, 6204–6213. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.-S.; Wu, T.-H.; Yang, Y.; Zhu, C.-L.; Ding, C.-L.; Dai, C.-C. Binding and detoxification of chlorpyrifos by lactic acid bacteria on rice straw silage fermentation. J. Environ. Sci. Health B 2016, 51, 316–325. [Google Scholar] [CrossRef]
- Lee, J.H.; Lee, H.Y.; Cho, D.Y.; Kim, M.J.; Jung, J.G.; Jeong, E.H.; Haque, M.A.; Cho, K.M. Biodegradable properties of organophosphorus insecticides by the potential probiotic Lactobacillus plantarum WCP931 with a degrading gene (opdc). Appl. Biol. Chem. 2021, 64, 62. [Google Scholar] [CrossRef]
- Daisley, B.A.; Trinder, M.; McDowell, T.W.; Welle, H.; Dube, J.S.; Ali, S.N.; Leong, H.S.; Sumarah, M.W.; Reid, G. Neonicotinoid-induced pathogen susceptibility is mitigated by Lactobacillus plantarum immune stimulation in a Drosophila melanogaster model. Sci. Rep. 2017, 7, 2703. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.; Sun, W.; An, J.; Xu, H.; Liu, Y.; Yan, C. Copper and chlorpyrifos stress affect the gut microbiota of chironomid larvae (Propsilocerus akamusi). Ecotoxicol. Environ. Saf. 2022, 244, 114027. [Google Scholar] [CrossRef] [PubMed]
- Kumral, A.; Kumral, N.A.; Gurbuz, O. Chlorpyrifos and Deltamethrin degradation potentials of two Lactobacillus plantarum (Orla-Jensen, 1919) (Lactobacillales: Lactobacillaceae) strains. Turk. Entomol. Derg. 2020, 44, 165–176. [Google Scholar] [CrossRef] [Green Version]
- Pinto, G.D.A.; Castro, I.M.; Miguel, M.A.L.; Koblitz, M.G.B. Lactic acid bacteria–Promising technology for organophosphate degradation in food: A pilot study. LWT 2019, 110, 353–359. [Google Scholar] [CrossRef]
- Cho, K.M.; Math, R.K.; Islam, S.M.; Lim, W.J.; Hong, S.Y.; Kim, J.M.; Yun, M.G.; Cho, J.J.; Yun, H.D. Biodegradation of chlorpyrifos by lactic acid bacteria during kimchi fermentation. J. Agric. Food Chem. 2009, 57, 1882–1889. [Google Scholar] [CrossRef] [PubMed]
- Cycoń, M.; Piotrowska-Seget, Z. Biochemical and microbial soil functioning after application of the insecticide imidacloprid. J. Environ. Sci. 2015, 27, 147–158. [Google Scholar] [CrossRef]
- Cycoń, M.; Markowicz, A.; Borymski, S.; Wójcik, M.; Piotrowska-Seget, Z. Imidacloprid induces changes in the structure, genetic diversity and catabolic activity of soil microbial communities. J. Environ. Manag. 2013, 131, 55–65. [Google Scholar] [CrossRef]
- Wang, M.-C.; Liu, Y.-H.; Wang, Q.; Gong, M.; Hua, X.-M.; Pang, Y.-J.; Hu, S.; Yang, Y.-H. Impacts of methamidophos on the biochemical, catabolic, and genetic characteristics of soil microbial communities. Soil Biol. Biochem. 2008, 40, 778–788. [Google Scholar] [CrossRef]
- Đorđević, T.M.; Šiler-Marinković, S.S.; Đurović-Pejčev, R.D.; Dimitrijević-Branković, S.I.; Gajić Umiljendić, J.S. Dissipation of pirimiphos-methyl during wheat fermentation by Lactobacillus plantarum. Lett. Appl. Microbiol. 2013, 57, 412–419. [Google Scholar] [CrossRef]
- Jia, H.-R.; Wu, Y.-Y.; Dai, P.-L.; Wang, Q.; Zhou, T. Effects of the sublethal doses of imidacloprid on the bacterial diversity in the midgut of Apis mellifera ligustica (Hymenoptera: Apidae)(In English). Acta Entomol. Sin. 2015, 58, 139–146. [Google Scholar] [CrossRef]
- Raymann, K.; Motta, E.V.; Girard, C.; Riddington, I.M.; Dinser, J.A.; Moran, N.A. Imidacloprid decreases honey bee survival rates but does not affect the gut microbiome. Appl. Environ. Microbiol. 2018, 84, e00545-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Almasri, H.; Liberti, J.; Brunet, J.-L.; Engel, P.; Belzunces, L.P. Mild chronic exposure to pesticides alters physiological markers of honey bee health without perturbing the core gut microbiota. Sci. Rep. 2022, 12, 4281. [Google Scholar] [CrossRef] [PubMed]
- Abdou, S.M.; Abdel-Gawaad, A.A.; Abo-El-Amaiem, E.; El-Alfy, A.A. Effect of some organochlorine insecticides on some species of bacteria used in the dairy industry. Egypt J. Dairy Sci. 1983, 11, 205–213. [Google Scholar]
- Alberoni, D.; Favaro, R.; Baffoni, L.; Angeli, S.; Di Gioia, D. Neonicotinoids in the agroecosystem: In-field long-term assessment on Honeybee colony strength and microbiome. Sci. Total Environ. 2021, 762, 144116. [Google Scholar] [CrossRef] [PubMed]
- Pachla, A.; Ptaszyńska, A.A.; Wicha, M.; Kunat, M.; Wydrych, J.; Oleńska, E.; Małek, W. Insight into probiotic properties of lactic acid bacterial endosymbionts of Apis mellifera L. derived from the Polish apiary. Saudi J. Biol. Sci. 2021, 28, 1890–1899. [Google Scholar] [CrossRef] [PubMed]
- Malfatti, A.; Mallmann, G.C.; Oliveira Filho, L.C.; Carniel, L.S.; Cruz, S.P.; Klauberg-Filho, O. Ecotoxicological test to assess effects of herbicides on spore germination of Rhizophagus clarus and Gigaspora albida. Ecotoxicol. Environ. Saf. 2021, 207, 111599. [Google Scholar] [CrossRef] [PubMed]
- Berber, İ.; Çökmüş, C.; Atalan, E. Effects of Some Pesticides on Spore Germination and Larvicidal Activity of Bacillus thuringiensis var. israelensis and Bacillus sphaericus 2362 Strain. Turk. J. Biol. 2004, 28, 15–21. [Google Scholar]
- Abou Ayana, I.A.A.; El Deen, A.A.; El-Metwall, M.A. Behavior of certain lactic acid bacteria in the presence of pesticides residues. Int. J. Dairy Sci. 2010, 6, 44–57. [Google Scholar] [CrossRef] [Green Version]
- Pettis, J.S.; Collins, A.M.; Wilbanks, R.; Feldlaufer, M.F. Effects of coumaphos on queen rearing in the honey bee, Apis mellifera. Apidologie 2004, 35, 605–610. [Google Scholar] [CrossRef]
- Haarmann, T.; Spivak, M.; Weaver, D.; Weaver, B.; Glenn, T. Effects of fluvalinate and coumaphos on queen honey bees (hymenoptera: Apidae) in two commercial queen rearing operations. J. Econ. Entomol. 2002, 95, 28–35. [Google Scholar] [CrossRef] [PubMed]
- Yoon, K.Y.; Woodams, E.E.; Hang, Y.D. Probiotication of tomato juice by lactic acid bacteria. Korean J. Microbiol. 2004, 42, 315–318. [Google Scholar]
- Zhang, M.; Ming, Y.; Guo, H.; Zhu, Y.; Yang, Y.; Chen, S.; He, L.; Ao, X.; Liu, A.; Zhou, K.; et al. Screening of lactic acid bacteria for their capacity to bind cypermethrin in vitro and the binding characteristics and its application. Food Chem. 2021, 347, 129000. [Google Scholar] [CrossRef] [PubMed]
- Rezaei, F.; Nejati, R.; Sayadi, M.; Nematillahi, A. Diazinon reduction in apple juice using probiotic bacteria during fermentation and refrigerated storage. Environ. Sci. Pollut. Res. 2021, 28, 61213–61224. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Ma, Y.; Mi, Z.; Huo, R.; Zhou, T.; Hai, H.; Kwok, L.; Sun, Z.; Chen, Y.; Zhang, H. Screening for lactobacillus plantarum strains that possess organophosphorus pesticide-degrading activity and metabolomic analysis of phorate degradation. Front. Microbiol. 2018, 9, 2048. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, S.; Li, C.; Yu, H.; Xie, Y.; Guo, Y.; Yao, W. Selective uptake determines the variation in degradation of organophosphorus pesticides by Lactobacillus plantarum. Food Chem. 2021, 360, 130106. [Google Scholar] [CrossRef] [PubMed]
- Sreekumar, O.; Hosono, A. Antimutagenicity and the influence of physical factors in binding Lactobacillus gasseri and Bifidobacterium longum cells to amino acid pyrolysates. J. Dairy Sci. 1998, 81, 1508–1516. [Google Scholar] [CrossRef] [PubMed]
- Islam, S.M.; Math, R.K.; Cho, K.M.; Lim, W.J.; Hong, S.Y.; Kim, J.M.; Yun, M.G.; Cho, J.J.; Yun, H.D. Organophosphorus hydrolase (OpdB) of Lactobacillus brevis WCP902 from kimchi is able to degrade organophosphorus pesticides. J. Agric. Food Chem. 2010, 58, 5380–5386. [Google Scholar] [CrossRef]
- Zhao, X.-H.; Wang, J. A brief study on the degradation kinetics of seven organophosphorus pesticides in skimmed milk cultured with Lactobacillus spp. at 42 °C. Food Chem. 2012, 131, 300–304. [Google Scholar] [CrossRef]
- Kostić, A.; Milinčić, D.; Petrović, T.; Krnjaja, V.; Stanojević, S.; Barać, M.; Tešić, Ž.; Pešić, M. Mycotoxins and mycotoxin producing fungi in pollen: Review. Toxins 2019, 11, 64. [Google Scholar] [CrossRef] [Green Version]
- Asurmendi, P.; Gerbaldo, G.; Pascual, L.; Barberis, L. Lactic acid bacteria with promising AFB1 binding properties as an alternative strategy to mitigate contamination on brewers’ grains. J. Environ. Health B 2020, 55, 1002–1008. [Google Scholar] [CrossRef]
- Genersch, E.; Gisder, S.; Hedtke, K.; Hunter, W.B.; Möckel, N.; Müller, U. Standard methods for cell cultures in Apis mellifera research. J. Apic. Res. 2013, 52, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Panyo, J.; Matsunami, K.; Panichayupakaranant, P. Bioassay-guided isolation and evaluation of antimicrobial compounds from Ixora megalophylla against some oral pathogens. Pharm. Biol. 2016, 54, 1522–1527. [Google Scholar] [CrossRef] [PubMed]
- Ilboudo, S.; Fouche, E.; Rizzati, V.; Toé, A.M.; Gamet-Payrastre, L.; Guissou, P.I. In vitro impact of five pesticides alone or in combination on human intestinal cell line Caco-2. Toxicol. Rep. 2014, 1, 474–489. [Google Scholar] [CrossRef] [PubMed]
- Abhishek, A.; Ansari, N.G.; Shankhwar, S.N.; Jain, A.; Singh, V. In vitro toxicity evaluation of low doses of pesticides in individual and mixed condition on human keratinocyte cell line. Bioinformation 2014, 10, 716–720. [Google Scholar] [CrossRef] [Green Version]
- Abdel-Halim, K.Y.; Osman, S.R. Cytotoxicity and oxidative stress responses of imidacloprid and glyphosate in human prostate epithelial wpm-Y.1 cell line. J. Toxicol. 2020, 2020, 4364650. [Google Scholar] [CrossRef]
- Yun, X.; Huang, Q.; Rao, W.; Xiao, C.; Zhang, T.; Mao, Z.; Wan, Z. A comparative assessment of cytotoxicity of commonly used agricultural insecticides to human and insect cells. Ecotoxicol. Environ. Saf. 2017, 137, 179–185. [Google Scholar] [CrossRef]
- Pandya, N.; Thakkar, B.; Pandya, P.; Parikh, P. Evaluation of insecticidal potential of organochemicals on SF9 cell line. J. Basic Appl. Zool. 2021, 82, 60. [Google Scholar] [CrossRef]
- Barabadi, H.; Najafi, M.; Samadian, H.; Azarnezhad, A.; Vahidi, H.; Mahjoub, M.; Koohiyan, M.; Ahmadi, A. A systematic review of the genotoxicity and Antigenotoxicity of biologically synthesized metallic nanomaterials: Are green nanoparticles safe enough for clinical marketing? Medicina 2019, 55, 439. [Google Scholar] [CrossRef] [Green Version]
- Ren, N.; Atyah, M.; Chen, W.-Y.; Zhou, C.-H. The various aspects of genetic and epigenetic toxicology: Testing methods and clinical applications. J. Transl. Med. 2017, 15, 110. [Google Scholar] [CrossRef] [Green Version]
- Kızılet, H.; Yilmaz, B.; Uysal, H. Herbal Medicine against genotoxicity of dimethoate, an insecticide, in mammalian somatic cells. Heliyon 2019, 5, e01337. [Google Scholar] [CrossRef]
- Kara, M.; ÖztaŞ, E.; Özhan, G. Acetamiprid-induced Cyto- and Genotoxicity in the AR42J Pancreatic Cell Line. Turk. J. Pharm. Sci. 2020, 17, 474–479. [Google Scholar] [CrossRef] [PubMed]
- Khodabandeh, Z.; Etebari, M.; Aliomrani, M. Study of the probable genotoxic effects of zolone (phosalone) exposure in mice bone marrow derived cells. Genes Environ. 2021, 43, 18. [Google Scholar] [CrossRef] [PubMed]
- Chandrakar, T.R.; Singh, A.P.; Chandra Sarkhel, B.; Nath Bagchi, S. In vitro cytotoxicity and genotoxicity assessments of carbofuran and malathion pesticides on cat (Felis catus) fibroblast cells. Biomed. Pharmacol. J. 2020, 13, 1157–1168. [Google Scholar] [CrossRef]
- Anguiano-Vega, G.A.; Cazares-Ramirez, L.H.; Rendon-Von Osten, J.; Santillan-Sidon, A.P.; Vazquez-Boucard, C.G. Risk of genotoxic damage in schoolchildren exposed to organochloride pesticides. Sci. Rep. 2020, 10, 11644. [Google Scholar] [CrossRef] [PubMed]
- Saleh, M.; Din, D.; Al-Masri, A. In vitro genotoxicity study of the lambda-cyhalothrin insecticide on Sf9 insect cells line using Comet assay. Jordan J. Bio. Sci. 2021, 14, 213–217. [Google Scholar] [CrossRef]
- El-Nahhal, Y. Pesticide residues in honey and their potential reproductive toxicity. Sci. Total Environ. 2020, 741, 139953. [Google Scholar] [CrossRef]
- González-Martín, M.I.; Revilla, I.; Vivar-Quintana, A.M.; Betances Salcedo, E.V. Pesticide residues in propolis from Spain and Chile. An approach using near infrared spectroscopy. Talanta 2017, 165, 533–539. [Google Scholar] [CrossRef]
- Friedle, C.; Wallner, K.; Rosenkranz, P.; Martens, D.; Vetter, W. Pesticide residues in daily bee pollen samples (April-July) from an intensive agricultural region in Southern Germany. Environ. Sci. Pollut. Res. Int. 2021, 28, 22789–22803. [Google Scholar] [CrossRef]
- Villalba, A.; Maggi, M.; Ondarza, P.M.; Szawarski, N.; Miglioranza, K.S.B. Influence of land use on chlorpyrifos and persistent organic pollutant levels in honey bees, bee bread and honey: Beehive exposure assessment. Sci. Total Environ. 2020, 713, 136554. [Google Scholar] [CrossRef]
- Pareja, L.; Colazzo, M.; Pérez-Parada, A.; Niell, S.; Carrasco-Letelier, L.; Besil, N.; Cesio, M.V.; Heinzen, H. Detection of Pesticides in Active and Depopulated Beehives in Uruguay. Int. J. Environ. Res. Public Health 2011, 8, 3844–3858. [Google Scholar] [CrossRef]
- Balayiannis, G.; Balayiannis, P. Bee Honey as an Environmental Bioindicator of Pesticides’ Occurrence in Six Agricultural Areas of Greece. Arch. Environ. Contam. Toxicol. 2008, 55, 462–470. [Google Scholar] [CrossRef] [PubMed]
- Woodcock, B.A.; Ridding, L.; Freeman, S.N.; Pereira, M.G.; Sleep, D.; Redhead, J.; Aston, D.; Carreck, N.L.; Shore, R.F.; Bullock, J.M.; et al. Neonicotinoid residues in UK Honey despite European Union moratorium. PLoS ONE 2018, 13, e0189681. [Google Scholar] [CrossRef] [PubMed]
- Valdovinos-Flores, C.; Gaspar-Ramírez, O.; Heras-Ramírez, M.E.; Lara-Álvarez, C.; Dorantes-Ugalde, J.A.; Saldaña-Loza, L.M. Boron and Coumaphos residues in hive materials following treatments for the Control of Aethina tumida Murray. PLoS ONE 2016, 11, e0153551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tao, Y.; Dong, F.; Xu, J.; Phung, D.; Liu, Q.; Li, R.; Liu, X.; Wu, X.; He, M.; Zheng, Y. Characteristics of neonicotinoid imidacloprid in urine following exposure of humans to orchards in China. Environ. Int. 2019, 132, 105079. [Google Scholar] [CrossRef]
- Mohamed, F.; Gawarammana, I.; Robertson, T.A.; Roberts, M.S.; Palangasinghe, C.; Zawahir, S.; Jayamanne, S.; Kandasamy, J.; Eddleston, M.; Buckley, N.A.; et al. Acute human self-poisoning with imidacloprid compound: A neonicotinoid insecticide. PLoS ONE 2009, 4, e5127. [Google Scholar] [CrossRef]
- Taheri, E.; Amin, M.M.; Daniali, S.S.; Abdollahpour, I.; Fatehizadeh, A.; Kelishadi, R. Health risk assessment of exposure to chlorpyrifos in pregnant women using deterministic and probabilistic approaches. PLoS ONE 2022, 17, e0262127. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Source | Remaining Pesticide Concentration (µg/mL) ± SD | ||
|---|---|---|---|---|
| Chlorpyrifos | Coumaphos | Imidacloprid | ||
| A. kunkeei DSM 12361 | Honeybee gut | 94.44 ± 0.34 | 70.29 ± 0.30 * | 97.70 ± 0.25 |
| L. rhamnosus GG | Human feces | 72.26 ± 0.45 * | 87.28 ± 0.41 * | 94.83 ± 0.31 |
| L. plantarum 8AN | Chicken feces | 85.68 ± 0.31 * | 83.60 ± 0.38 * | 81.88 ± 0.55 * |
| L. salivarius 9AN | Chicken feces | 94.21 ± 0.45 | 67.37 ± 0.31 * | 63.78 ± 0.45 * |
| L. casei 12AN | Human feces | 47.98 ± 0.23 * | 58.95 ± 0.20 * | 90.74 ± 0.25 |
| P. parvulus OK-S | Fermented cucumbers | 36.11 ± 0.25 * | 93.30 ± 0.29 | 73.12 ± 0.21 * |
| L. brevis KKA | Fermented cabbage | 66.98 ± 0.44 * | 77.85 ± 0.38 * | 99.10 ± 0.42 |
| L. plantarum 145 | Vegetable silage | 65.46 ± 0.38 * | 75.05 ± 0.20 * | 95.91 ± 0.45 |
| L. acidophilus 573 | Nd a | 100.31 ± 0.48 | 77.97 ± 0.51 * | 87.30 ± 0.61 * |
| P. acidilactici 4/1 | Robinia pseudoaccacia L. b | 74.75 ± 0.40 * | 92.15 ± 0.35 | 90.96 ± 0.31 |
| P. acidilactici 5/2 | Weigela florida DC. b | 38.26 ± 0.20 * | 60.28 ± 0.25 * | 97.54 ± 0.21 |
| P. acidilactici 6/1 | Centaurea jacea L. b | 84.88 ± 0.45 * | 60.70 ± 0.50 * | 84.22 ± 0.48 * |
| P. acidilactici 7/1 | Papaver rhoeas L. b | 62.04 ± 0.30 * | 49.38 ± 0.30 * | 95.54 ± 0.33 |
| P. acidilactici 8/1 | Sinapis arvensis L. b | 82.36 ± 0.41 * | 61.37 ± 0.35 * | 94.19 ± 0.27 |
| P. pentosaceus 9/3 | Trifolium pratense L. b | 99.08 ± 0.50 | 68.23 ± 0.40 * | 98.47 ± 0.42 |
| P. pentosaceus 10/2 | Sambucus nigra L. b | 69.18 ± 0.33 * | 97.84 ± 0.50 | 92.24 ± 0.40 |
| P. pentosaceus 11/3 | Philadelphus coronaries L. b | 64.91 ± 0.30 * | 100.97 ± 0.89 | 97.23 ± 0.29 |
| P. pentosaceus 14/1 | Lavandula augustifolia L. b | 36.07 ± 0.24 * | 42.96 ± 0.31 * | 98.32 ± 0.29 |
| L. plantarum 14/3 | Lavandula augustifolia L. b | 61.84 ± 0.30 * | 46.28 ± 0.22 * | 97.91 ± 0.51 |
| L. plantarum 18/1 | Buddleja davidii L. b | 58.65 ± 0.28 * | 82.96 ± 0.44 * | 99.08 ± 0.18 |
| P. pentosaceus 19/1 | Calluna vulgaris L. b | 98.99 ± 0.47 | 69.20 ± 0.30 * | 93.40 ± 0.39 |
| L. plantarum 21/1 | Fermented (spoiled) honey | 61.56 ± 0.29 * | 94.08 ± 0.29 | 97.45 ± 0.38 |
| P. acidilactici 22/1 | Royal jelly | 92.94 ± 0.46 | 91.83 ± 0.67 | 91.13 ± 0.68 |
| P. acidilactici 25/1 | Heather nectar honey | 74.18 ± 0.31 * | 89.23 ± 0.81 * | 98.42 ± 0.40 |
| P. acidilactici 35/1 | Goldenrod honey | 96.18 ± 0.48 | 50.81 ± 0.32 * | 84.22 ± 0.36 * |
| Analytical standard | - | 100.00 ± 0.86 | 100.00 ± 0.99 | 100.00 ± 0.29 |
| Strain | Remaining Chlorpyrifos Concentration (µg/mL) ± SD | |
|---|---|---|
| Intracellular Extracts | Membrane Extracts | |
| L. casei 12AN | 0.31 ± 0.07 | 0.03 ± 0.01 |
| P. parvulus OK-S | 21.27 ± 0.39 | 2.87 ± 0.18 |
| L. brevis KKA | 2.49 ± 0.13 | 0.28 ± 0.02 |
| L. plantarum 145 | 1.42 ± 0.10 | 0.19 ± 0.05 |
| P. acidilactici 5/2 | 6.86 ± 0.15 | 0.72 ± 0.07 |
| P. acidilactici 7/1 | 9.50 ± 0.27 | 2.09 ± 0.10 |
| P. pentosaceus 10/2 | 0.27 ± 0.03 | 0.11 ± 0.03 |
| P. pentosaceus 11/3 | 0.74 ± 0.09 | 0.11 ± 0.04 |
| P. pentosaceus 14/1 | 2.70 ± 0.17 | 0.19 ± 0.01 |
| L. plantarum 14/3 | 2.02 ± 0.15 | 0.41 ± 0.04 |
| L. plantarum 18/1 | 7.37 ± 0.25 | 0.62 ± 0.10 |
| L. plantarum 21/1 | 1.10 ± 0.12 | 0.10 ± 0.03 |
| Analytical standard | 100.00 ± 0.45 | |
| Strain | Remaining Coumaphos Concentration (µg/mL) ± SD | |
|---|---|---|
| Intracellular Extracts | Membrane Extracts | |
| L. salivarius 9AN | 1.45 ± 0.07 | 0.11 ± 0.05 |
| L. casei 12AN | 1.81 ± 0.10 | 0.13 ± 0.05 |
| P. acidilactici 5/2 | 2.23 ± 0.13 | 0.25 ± 0.07 |
| P. acidilactici 6/1 | 1.53 ± 0.12 | 0.13 ± 0.03 |
| P. acidilactici 7/1 | 1.71 ± 0.10 | 0.09 ± 0.02 |
| P. acidilactici 8/1 | 1.13 ± 0.08 | 0.17 ± 0.05 |
| P. pentosaceus 9/3 | 1.40 ± 0.06 | 0.13 ± 0.03 |
| P. pentosaceus 14/1 | 0.61 ± 0.04 | 0.12 ± 0.04 |
| L. plantarum 14/3 | 0.79 ± 0.06 | 0.02 ± 0.01 |
| P. pentosaceus 19/1 | 0.89 ± 0.05 | 0.04 ± 0.01 |
| P. acidilactici 35/1 | 1.89 ± 0.10 | 0.19 ± 0.03 |
| Analytical standard | 100.00 ± 0.66 | |
| Strain | Remaining Imidacloprid Concentration (µg/mL) ± SD | |
|---|---|---|
| Intracellular Extracts | Membrane Extracts | |
| L. plantarum 8AN | 2.44 ± 0.13 | 0.60 ± 0.08 |
| L. salivarius 9AN | 7.02 ± 0.11 | 1.28 ± 0.10 |
| P. parvulus OK-S | 5.99 ± 0.19 | 1.34 ± 0.11 |
| Analytical standard | 100.00 ± 0.72 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Leska, A.; Nowak, A.; Miśkiewicz, K.; Rosicka-Kaczmarek, J. Binding and Detoxification of Insecticides by Potentially Probiotic Lactic Acid Bacteria Isolated from Honeybee (Apis mellifera L.) Environment—An In Vitro Study. Cells 2022, 11, 3743. https://doi.org/10.3390/cells11233743
Leska A, Nowak A, Miśkiewicz K, Rosicka-Kaczmarek J. Binding and Detoxification of Insecticides by Potentially Probiotic Lactic Acid Bacteria Isolated from Honeybee (Apis mellifera L.) Environment—An In Vitro Study. Cells. 2022; 11(23):3743. https://doi.org/10.3390/cells11233743
Chicago/Turabian StyleLeska, Aleksandra, Adriana Nowak, Karolina Miśkiewicz, and Justyna Rosicka-Kaczmarek. 2022. "Binding and Detoxification of Insecticides by Potentially Probiotic Lactic Acid Bacteria Isolated from Honeybee (Apis mellifera L.) Environment—An In Vitro Study" Cells 11, no. 23: 3743. https://doi.org/10.3390/cells11233743