

Probiotics Mechanism of Action on Immune Cells and Beneficial Effects on Human Health
 ,
, 
Abstract
:1. Introduction
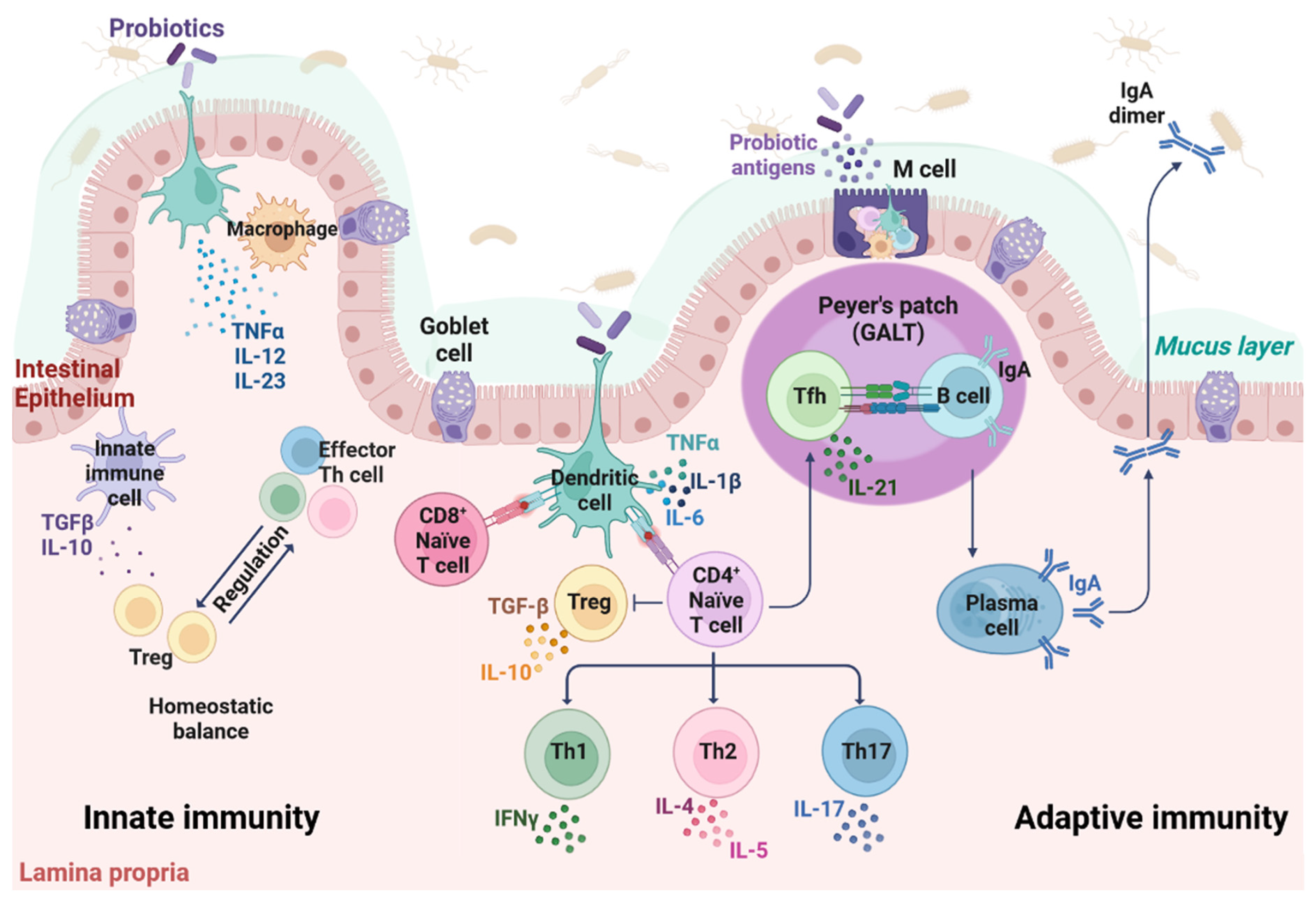
2. The Immune System of the Gut
3. Probiotic Bacteria
The Gut-Central Nervous System and Gut-Respiratory System Axes
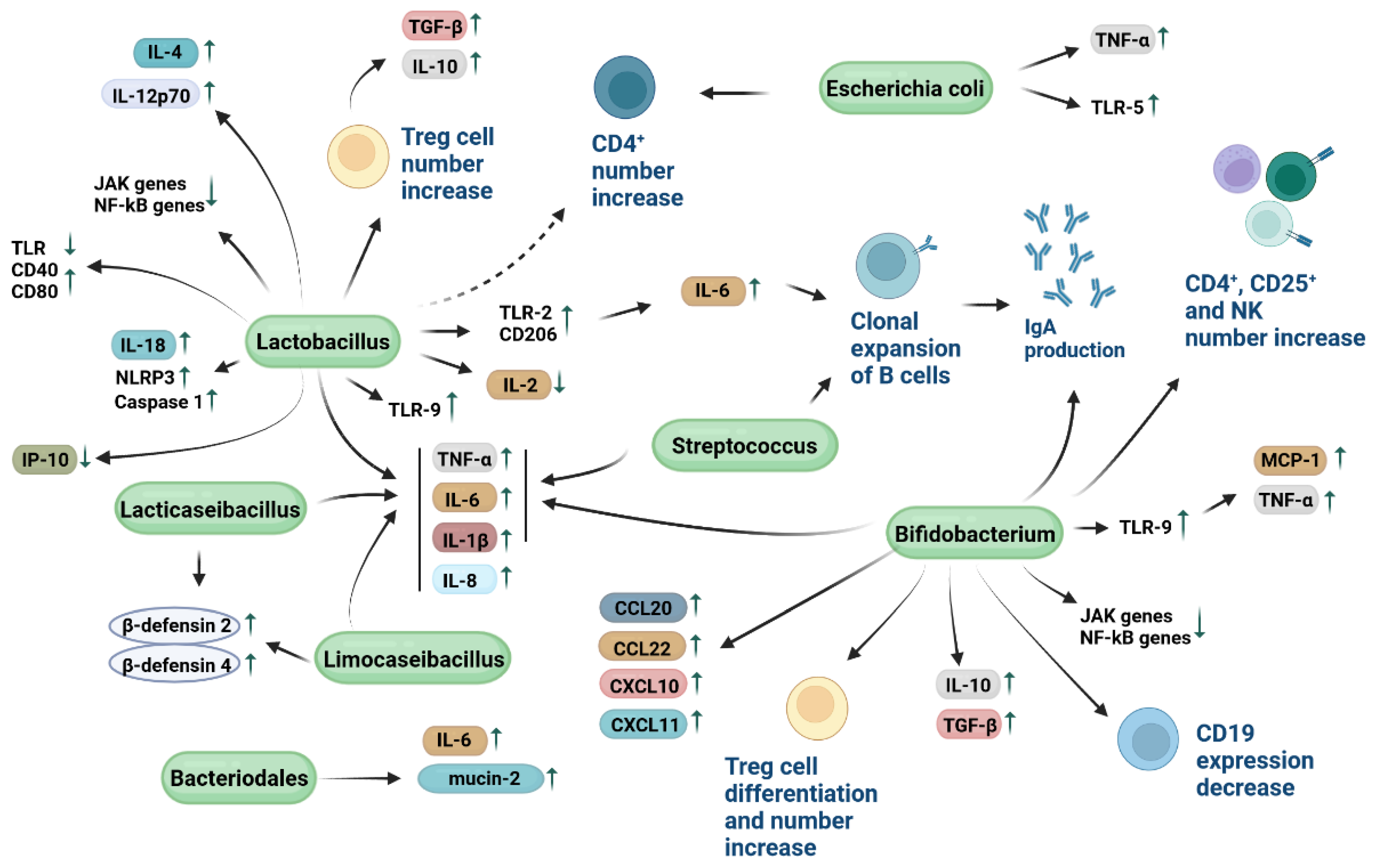
4. Immune Modulatory Mechanisms of Probiotic Bacteria
5. Host Cytokine Release and Probiotics
6. Animal Model-Based Studies with Probiotics
7. Guidelines on the Use of Probiotics in Clinical Practice and Probiotic-Based Clinical Trials
8. Probiotic Industrial Production Challenges
9. Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhou, B.; Yuan, Y.; Zhang, S.; Guo, C.; Li, X.; Li, G.; Xiong, W.; Zeng, Z. Intestinal Flora and Disease Mutually Shape the Regional Immune System in the Intestinal Tract. Front. Immunol. 2020, 11, 575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharifi-Rad, J.; Rodrigues, C.F.; Stojanović-Radić, Z.; Dimitrijević, M.; Aleksić, A.; Neffe-Skocińska, K.; Zielińska, D.; Kołożyn-Krajewska, D.; Salehi, B.; Prabu, S.M.; et al. Probiotics: Versatile Bioactive Components in Promoting Human Health. Medicina 2020, 56, 433. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.-Y.; Tsolis, R.M.; Bäumler, A.J. The microbiome and gut homeostasis. Science 2022, 37, eabp9960. [Google Scholar] [CrossRef] [PubMed]
- Chinda, D.; Takada, T.; Mikami, T.; Shimizu, K.; Oana, K.; Arai, T.; Akitaya, K.; Sakuraba, H.; Katto, M.; Nagara, Y.; et al. Spatial distribution of live gut microbiota and bile acid metabolism in various parts of human large intestine. Sci. Rep. 2022, 12, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Yan, F.; Polk, D.B. Probiotics and immune health. Curr. Opin. Gastroenterol. 2011, 27, 496–501. [Google Scholar] [CrossRef] [Green Version]
- Serek, P.; Oleksy-Wawrzyniak, M. The Effect of Bacterial Infections, Probiotics and Zonulin on Intestinal Barrier Integrity. Int. J. Mol. Sci. 2021, 22, 11359. [Google Scholar] [CrossRef] [PubMed]
- Morelli, L.; Capurso, L. FAO/WHO guidelines on probiotics: 10 years later. J. Clin. Gastroenterol. 2012, 46, S1–S2. [Google Scholar] [CrossRef]
- Ashraf, R.; Shah, N.P. Immune System Stimulation by Probiotic Microorganisms. Crit. Rev. Food Sci. Nutr. 2014, 54, 938–956. [Google Scholar] [CrossRef]
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef] [Green Version]
- Adel, M.; El-Sayed, A.F.M.; Yeganeh, S.; Dadar, M.; Giri, S.S. Effect of Potential Probiotic Lactococcus lactis Subsp. lactis on Growth Performance, Intestinal Microbiota, Digestive Enzyme Activities, and Disease Resistance of Litopenaeus vannamei. Probiotics Antimicrob. Proteins 2017, 9, 150–156. [Google Scholar] [CrossRef] [PubMed]
- Azcárate-Peril, M.A.; Sikes, M.; Bruno-Bárcena, J.M. The intestinal microbiota, gastrointestinal environment and colorectal cancer: A putative role for probiotics in prevention of colorectal cancer? Am. J. Physiol. Gastrointest. Liver Physiol. 2011, 301, G401–G424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maldonado Galdeano, C.; Cazorla, S.I.; Lemme Dumit, J.M.; Vélez, E.; Perdigón, G. Beneficial Effects of Probiotic Consumption on the Immune System. Ann. Nutr. Metab. 2019, 74, 115–124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kałużna-Czaplińska, J.; Gątarek, P.; Chartrand, M.S.; Dadar, M.; Bjørklund, G. Is there a relationship between intestinal microbiota, dietary compounds, and obesity? Trends Food Sci. Technol. 2017, 70, 105–113. [Google Scholar] [CrossRef]
- Umair, M.; Jabbar, S.; Zhaoxin, L.; Jianhao, Z.; Abid, M.; Khan, K.-U.R.; Korma, S.A.; Alghamdi, M.A.; El-Saadony, M.T.; Abd El-Hack, M.E.; et al. Probiotic-Based Bacteriocin: Immunity Supplementation Against Viruses. An Updated Review. Front. Microbiol. 2022, 13, 1633. [Google Scholar] [CrossRef]
- Peng, X.; Ed-Dra, A.; Song, Y.; Elbediwi, M.; Nambiar, R.B.; Zhou, X.; Yue, M. Lacticaseibacillus rhamnosus alleviates intestinal inflammation and promotes microbiota-mediated protection against Salmonella fatal infections. Front. Immunol. 2022, 13, 973224. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A. Importance of Probiotics in Cancer Prevention and Treatment. Recent Dev. Appl. Microbiol. Biochem. 2019, 33–45. [Google Scholar]
- Smith, D.; Jheeta, S.; Fuentes, H.V.; Palacios-Pérez, M. Feeding Our Microbiota: Stimulation of the Immune/Semiochemical System and the Potential Amelioration of Non-Communicable Diseases. Life 2022, 12, 1197. [Google Scholar] [CrossRef] [PubMed]
- Anand, A.; Sato, M.; Aoyagi, H. Screening of Phosphate-accumulating Probiotics for Potential Use in Chronic Kidney Disorder. Food Sci. Technol. Res. 2019, 25, 89–96. [Google Scholar] [CrossRef]
- Cervin, A.U. The potential for topical probiotic treatment of chronic rhinosinusitis, a personal perspective. Front. Cell. Infect. Microbiol. 2018, 7, 530. [Google Scholar] [CrossRef]
- Kim, Y.-K.; Shin, C. The Microbiota-Gut-Brain Axis in Neuropsychiatric Disorders: Pathophysiological Mechanisms and Novel Treatments. Curr. Neuropharmacol. 2018, 16, 559–573. [Google Scholar] [CrossRef]
- Schemczssen-Graeff, Z.; Pileggi, M. Probiotics and live biotherapeutic products aiming at cancer mitigation and patient recover. Front. Genet. 2022, 13, 921972. [Google Scholar] [CrossRef] [PubMed]
- Beterams, A.; De Paepe, K.; Maes, L.; Wise, I.J.; De Keersmaecker, H.; Rajkovic, A.; Laukens, D.; Van de Wiele, T.; Calatayud Arroyo, M. Versatile human in vitro triple coculture model coincubated with adhered gut microbes reproducibly mimics pro-inflammatory host-microbe interactions in the colon. FASEB J. 2021, 35, e21992. [Google Scholar] [CrossRef]
- Kumar, H.; Schütz, F.; Bhardwaj, K.; Sharma, R.; Nepovimova, E.; Dhanjal, D.S.; Verma, R.; Kumar, D.; Kuča, K.; Cruz-Martins, N. Recent advances in the concept of paraprobiotics: Nutraceutical/functional properties for promoting children health. Crit Rev Food Sci Nutr. 2021, 8, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Ke, A.; Parreira, V.R.; Goodridge, L.; Farber, J.M. Current and Future Perspectives on the Role of Probiotics, Prebiotics, and Synbiotics in Controlling Pathogenic Cronobacter Spp. in Infants. Front. Microbiol. 2021, 12, 3158. [Google Scholar] [CrossRef] [PubMed]
- Fidanza, M.; Panigrahi, P.; Kollmann, T.R. Lactiplantibacillus plantarum–Nomad and Ideal Probiotic. Front. Microbiol. 2021, 12, 2911. [Google Scholar] [CrossRef]
- Hunyady, B.; Mezey, E.; Palkovits, M. Gastrointestinal immunology: Cell types in the lamina propria—A morphological review. Acta Physiol Hung 2000, 87, 305–328. [Google Scholar]
- Donaldson, D.S.; Else, K.J.; Mabbott, N.A. The Gut-Associated Lymphoid Tissues in the Small Intestine, Not the Large Intestine, Play a Major Role in Oral Prion Disease Pathogenesis. J. Virol. 2015, 89, 9532–9547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mörbe, U.M.; Jørgensen, P.B.; Fenton, T.M.; von Burg, N.; Riis, L.B.; Spencer, J.; Agace, W.W. Human gut-associated lymphoid tissues (GALT); diversity, structure, and function. Mucosal Immunol. 2021, 14, 793–802. [Google Scholar] [CrossRef]
- Liu, K. Dendritic Cells. Encycl. Cell Biol. 2016, 3, 741–749. [Google Scholar]
- Reinholdt, J.; Husby, S. IgA and Mucosal Homeostasis; Landes Bioscience: Austin, TX, USA, 2013. [Google Scholar]
- Rotondo, J.C.; Martini, F.; Maritati, M.; Caselli, E.; Gallenga, C.E.; Guarino, M.; De Giorgio, R.; Mazziotta, C.; Tramarin, M.L.; Badiale, G.; et al. Advanced Molecular and Immunological Diagnostic Methods to Detect SARS-CoV-2 Infection. Microorganisms 2022, 10, 1193. [Google Scholar] [CrossRef] [PubMed]
- Nogueira, D.S.; De Oliveira, L.M.; Amorim, C.C.O.; Gazzinelli-Guimaraes, A.C.; Barbosa, F.S.; Oliveira, F.M.S.; Kraemer, L.; Mattos, M.; Cardoso, M.S.; Resende, N.M.; et al. Eosinophils mediate SIgA production triggered by TLR2 and TLR4 to control Ascaris suum infection in mice. PLoS Pathog. 2021, 17, e1010067. [Google Scholar] [CrossRef] [PubMed]
- Hansen, I.S.; Baeten, D.L.P.; den Dunnen, J. The inflammatory function of human IgA. Cell. Mol. Life Sci. 2018, 76, 1041–1055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hollenberg, M.D.; Epstein, M. The innate immune response, microenvironment proteinases, and the COVID-19 pandemic: Pathophysiologic mechanisms and emerging therapeutic targets. Kidney Int. Suppl. 2022, 12, 48–62. [Google Scholar] [CrossRef] [PubMed]
- Primorac, D.; Vrdoljak, K.; Brlek, P.; Pavelić, E.; Molnar, V.; Matišić, V.; Erceg Ivkošić, I.; Parčina, M. Adaptive Immune Responses and Immunity to SARS-CoV-2. Front. Immunol. 2022, 13, 848582. [Google Scholar] [CrossRef]
- Malagutti, N.; Rotondo, J.C.; Cerritelli, L.; Melchiorri, C.; De Mattei, M.; Selvatici, R.; Oton-Gonzalez, L.; Stomeo, F.; Mazzoli, M.; Borin, M.; et al. High human papillomavirus DNA loads in inflammatory middle ear diseases. Pathogens 2020, 9, 224. [Google Scholar] [CrossRef] [Green Version]
- Rigo, A.; Ferrarini, I.; Lorenzetto, E.; Darra, E.; Liparulo, I.; Bergamini, C.; Sissa, C.; Cavalieri, E.; Vinante, F. BID and the α-bisabolol-triggered cell death program: Converging on mitochondria and lysosomes. Cell Death Dis. 2019, 10, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Mazziotta, C.; Pellielo, G.; Tognon, M.; Martini, F.; Rotondo, J.C. Significantly low levels of IgG antibodies against oncogenic Merkel cell polyomavirus in sera from females affected by spontaneous abortion. Front. Microbiol. 2021, 12, 789991. [Google Scholar] [CrossRef]
- Guihot, A.; Litvinova, E.; Autran, B.; Debré, P.; Vieillard, V. Cell-Mediated Immune Responses to COVID-19 Infection. Front. Immunol. 2020, 11, 1662. [Google Scholar] [CrossRef]
- Rotondo, J.C.; Mazziotta, C.; Lanzillotti, C.; Tognon, M.; Martini, F. Epigenetic Dysregulations in Merkel Cell Polyomavirus-Driven Merkel Cell Carcinoma. Int. J. Mol. Sci. 2021, 22, 11464. [Google Scholar] [CrossRef]
- Mazziotta, C.; Lanzillotti, C.; Gafà, R.; Touzé, A.; Durand, M.-A.; Martini, F.; Rotondo, J.C. The role of histone post-translational modifications in Merkel cell carcinoma. Front. Oncol. 2022, 12, 832047. [Google Scholar] [CrossRef]
- Golubovskaya, V.; Wu, L. Different Subsets of T Cells, Memory, Effector Functions, and CAR-T Immunotherapy. Cancers 2016, 8, 36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luckheeram, R.V.; Zhou, R.; Verma, A.D.; Xia, B. CD4 +T cells: Differentiation and functions. Clin. Dev. Immunol. 2012, 2012, 925135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kany, S.; Vollrath, J.T.; Relja, B. Cytokines in Inflammatory Disease. Int. J. Mol. Sci. 2019, 20, 6008. [Google Scholar] [CrossRef]
- Corazza, M.; Oton-Gonzalez, L.; Scuderi, V.; Rotondo, J.C.; Lanzillotti, C.; Di Mauro, G.; Tognon, M.; Martini, F.; Borghi, A. Tissue cytokine/chemokine profile in vulvar lichen sclerosus: An observational study on keratinocyte and fibroblast cultures. J. Dermatol. Sci. 2020, 100, 223–226. [Google Scholar] [CrossRef] [PubMed]
- Ferrarini, I.; Rigo, A.; Zamò, A.; Vinante, F. Classical Hodgkin lymphoma cells may promote an IL-17-enriched microenvironment. Leuk. Lymphoma 2019, 60, 3395–3405. [Google Scholar] [CrossRef] [PubMed]
- Krawiec, P.; Pawłowska-Kamieniak, A.; Pac-Kożuchowska, E. Interleukin 10 and interleukin 10 receptor in paediatric inflammatory bowel disease: From bench to bedside lesson. J. Inflamm. 2021, 18, 13. [Google Scholar] [CrossRef]
- Mazziotta, C.; Rotondo, J.C.; Lanzillotti, C.; Campione, G.; Martini, F.; Tognon, M. Cancer biology and molecular genetics of A3 adenosine receptor. Oncogene 2021, 41, 301–308. [Google Scholar] [CrossRef]
- Rotondo, J.C.; Mazziotta, C.; Lanzillotti, C.; Stefani, C.; Badiale, G.; Campione, G.; Martini, F.; Tognon, M. The Role of Purinergic P2X7 Receptor in Inflammation and Cancer: Novel Molecular Insights and Clinical Applications. Cancers 2022, 14, 1116. [Google Scholar] [CrossRef]
- La Fata, G.; Weber, P.; Mohajeri, M.H. Probiotics and the Gut Immune System: Indirect Regulation. Probiotics Antimicrob. Proteins 2018, 10, 11–21. [Google Scholar] [CrossRef]
- Zeinali, T.; Faraji, N.; Joukar, F.; Khan Mirzaei, M.; Kafshdar Jalali, H.; Shenagari, M.; Mansour-Ghanaei, F. Gut bacteria, bacteriophages, and probiotics: Tripartite mutualism to quench the SARS-CoV2 storm. Microb. Pathog. 2022, 170, 105704. [Google Scholar] [CrossRef]
- Jung, J.H.; Kim, S.-E.; Suk, K.T.; Kim, D.J. Gut microbiota-modulating agents in alcoholic liver disease: Links between host metabolism and gut microbiota. Front. Med. 2022, 9, 2171. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; An, Y.; Qin, X.; Wu, X.; Wang, X.; Hou, H.; Song, X.; Liu, T.; Wang, B.; Huang, X.; et al. Gut Microbiota-Derived Metabolites in Colorectal Cancer: The Bad and the Challenges. Front. Oncol. 2021, 11, 4287. [Google Scholar] [CrossRef] [PubMed]
- de Vos, W.M.; Tilg, H.; Van Hul, M.; Cani, P.D. Gut microbiome and health: Mechanistic insights. Gut 2022, 71, 1020–1032. [Google Scholar] [CrossRef] [PubMed]
- Merchak, A.; Gaultier, A. Microbial metabolites and immune regulation: New targets for major depressive disorder. Brain Behav. Immun.-Health 2020, 9, 100169. [Google Scholar] [CrossRef] [PubMed]
- Nakov, R.; Velikova, T. Chemical Metabolism of Xenobiotics by Gut Microbiota. Curr. Drug Metab. 2020, 21, 260–269. [Google Scholar] [CrossRef] [PubMed]
- Fakharian, F.; Asgari, B.; Nabavi-Rad, A.; Sadeghi, A.; Soleimani, N.; Yadegar, A.; Zali, M.R. The interplay between Helicobacter pylori and the gut microbiota: An emerging driver influencing the immune system homeostasis and gastric carcinogenesis. Front. Cell. Infect. Microbiol. 2022, 12, 953718. [Google Scholar] [CrossRef]
- Thursby, E.; Juge, N. Introduction to the human gut microbiota. Biochem. J. 2017, 474, 1823–1836. [Google Scholar] [CrossRef]
- Yang, F.; Chen, H.; Gao, Y.; An, N.; Li, X.; Pan, X.; Yang, X.; Tian, L.; Sun, J.; Xiong, X.; et al. Gut microbiota-derived short-chain fatty acids and hypertension: Mechanism and treatment. Biomed. Pharmacother. 2020, 130, 110503. [Google Scholar] [CrossRef]
- Markowiak-Kopeć, P.; Śliżewska, K. The Effect of Probiotics on the Production of Short-Chain Fatty Acids by Human Intestinal Microbiome. Nutrients 2020, 12, 1107. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Liu, F.; Lu, J.; Shi, J.; Guan, J.; Yan, F.; Li, B.; Huo, G. Probiotic Mixture of Lactobacillus plantarum Strains Improves Lipid Metabolism and Gut Microbiota Structure in High Fat Diet-Fed Mice. Front. Microbiol. 2020, 11, 512. [Google Scholar] [CrossRef] [Green Version]
- Eslami, M.; Bahar, A.; Keikha, M.; Karbalaei, M.; Kobyliak, N.M.; Yousefi, B. Probiotics function and modulation of the immune system in allergic diseases. Allergol. Immunopathol. 2020, 48, 771–788. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.; Zhang, J. Role of intestinal microbiota and metabolites on gut homeostasis and human diseases. BMC Immunol. 2017, 18, 1–25. [Google Scholar] [CrossRef] [Green Version]
- Tan, B.; Liu, Y.; Tang, H.; Chen, D.; Xu, Y.; Chen, M.; Li, Y.; Wang, M.; Qian, J. Gut microbiota shed new light on the management of immune-related adverse events. Thorac. Cancer 2022, 13, 2681–2691. [Google Scholar] [CrossRef]
- Chen, W.; Wang, J.; Du, L.; Chen, J.; Zheng, Q.; Li, P.; Du, B.; Fang, X.; Liao, Z. Kefir microbiota and metabolites stimulate intestinal mucosal immunity and its early development. Crit. Rev. Food Sci. Nutr. 2022, 30, 1–14. [Google Scholar] [CrossRef]
- Li, Q.; Li, N.; Cai, W.; Xiao, M.; Liu, B.; Zeng, F. Fermented natural product targeting gut microbiota regulate immunity and anti-inflammatory activity: A possible way to prevent COVID-19 in daily diet. J. Funct. Foods 2022, 97, 105229. [Google Scholar] [CrossRef]
- Vijay, K. Toll-like receptors in immunity and inflammatory diseases: Past, present, and future. Int. Immunopharmacol. 2018, 59, 391–412. [Google Scholar] [CrossRef]
- Claes, A.K.; Zhou, J.Y.; Philpott, D.J. NOD-like receptors: Guardians of intestinal mucosal barriers. Physiology 2015, 30, 241–250. [Google Scholar] [CrossRef] [Green Version]
- Belkaid, Y.; Hand, T.W. Role of the Microbiota in Immunity and Inflammation. Cell 2014, 157, 121–141. [Google Scholar] [CrossRef] [Green Version]
- Zheng, D.; Liwinski, T.; Elinav, E. Interaction between microbiota and immunity in health and disease. Cell Res. 2020, 30, 492–506. [Google Scholar] [CrossRef]
- Kogut, M.H.; Lee, A.; Santin, E. Microbiome and pathogen interaction with the immune system. Poult. Sci. 2020, 99, 1906–1913. [Google Scholar] [CrossRef]
- Postler, T.S.; Ghosh, S. Cell Metabolism Review Understanding the Holobiont: How Microbial Metabolites Affect Human Health and Shape the Immune System. Cell Metab. 2017, 26, 110–130. [Google Scholar] [CrossRef] [Green Version]
- Azad, M.A.K.; Sarker, M.; Wan, D. Immunomodulatory Effects of Probiotics on Cytokine Profiles. Biomed Res. Int. 2018, 2018, 8063647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cristofori, F.; Dargenio, V.N.; Dargenio, C.; Miniello, V.L.; Barone, M.; Francavilla, R. Anti-Inflammatory and Immunomodulatory Effects of Probiotics in Gut Inflammation: A Door to the Body. Front. Immunol. 2021, 12, 178. [Google Scholar] [CrossRef] [PubMed]
- Damián, M.R.; Cortes-Perez, N.G.; Quintana, E.T.; Ortiz-Moreno, A.; Noguez, C.G.; Cruceño-Casarrubias, C.E.; Pardo, M.E.S.; Bermúdez-Humarán, L.G. Functional Foods, Nutraceuticals and Probiotics: A Focus on Human Health. Microorganisms 2022, 10, 1065. [Google Scholar] [CrossRef]
- Yadav, M.K.; Kumari, I.; Singh, B.; Sharma, K.K.; Tiwari, S.K. Probiotics, prebiotics and synbiotics: Safe options for next-generation therapeutics. Appl. Microbiol. Biotechnol. 2022, 106, 505–521. [Google Scholar] [CrossRef]
- Miri, S.T.; Sotoodehnejadnematalahi, F.; Amiri, M.M.; Pourshafie, M.R.; Rohani, M. The impact of Lactobacillus and Bifidobacterium probiotic cocktail on modulation of gene expression of gap junctions dysregulated by intestinal pathogens. Arch. Microbiol. 2022, 204, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Araujo, L.D.C.; Furlaneto, F.A.C.; da Silva, L.A.B.; Kapila, Y.L. Use of the Probiotic Bifidobacterium animalis subsp. lactis HN019 in Oral Diseases. Int. J. Mol. Sci. 2022, 23, 9334. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Liang, X.; Wu, Y.; Wang, R.; Liu, T.; Yi, H.; Yu, Z.; Zhang, Z.; Gong, P.; Zhang, L. Lanwei Bifidobacterium animalis sup F1-7 Acts as an Effective Activator to Regulate Immune Response Via Casepase-3 and Bak of FAS/CD95 Pathway. Probiotics Antimicrob. Proteins 2022, 1, 1–16. [Google Scholar]
- Ding, M.; Zheng, Y.; Liu, F.; Tian, F.; Ross, R.P.; Stanton, C.; Yu, R.; Zhao, J.; Zhang, H.; Yang, B.; et al. Lactation time influences the composition of Bifidobacterium and Lactobacillus at species level in human breast milk. Benef. Microbes 2022, 13, 319–330. [Google Scholar] [CrossRef]
- Mourand, G.; Paboeuf, F.; Grippon, P.; Lucas, P.; Bougeard, S.; Denamur, E.; Kempf, I. Impact of Escherichia coli probiotic strains ED1a and Nissle 1917 on the excretion and gut carriage of extended-spectrum beta-lactamase-producing E. coli in pigs. Vet. Anim. Sci. 2021, 14, 100217. [Google Scholar] [CrossRef]
- Nami, Y.; Bakhshayesh, R.V.; Jalaly, H.M.; Lotfi, H.; Eslami, S.; Hejazi, M.A. Probiotic properties of enterococcus isolated from artisanal dairy products. Front. Microbiol. 2019, 10, 300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baccouri, O.; Boukerb, A.M.; Farhat, L.B.; Zébré, A.; Zimmermann, K.; Domann, E.; Cambronel, M.; Barreau, M.; Maillot, O.; Rincé, I.; et al. Probiotic Potential and Safety Evaluation of Enterococcus faecalis OB14 and OB15, Isolated from Traditional Tunisian Testouri Cheese and Rigouta, Using Physiological and Genomic Analysis. Front. Microbiol. 2019, 10, 881. [Google Scholar] [CrossRef]
- Jiang, S.; Cai, L.; Lv, L.; Li, L. Pediococcus pentosaceus, a future additive or probiotic candidate. Microb. Cell Factories 2021, 20, 45. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.-R.; Lee, C.-M.; Lee, S.-H.; Baik, S.-H.; Fusco, V. Evaluation of Probiotic Properties of Pediococcus acidilactici M76 Producing Functional Exopolysaccharides and Its Lactic Acid Fermentation of Black Raspberry Extract Probiotic Properties of Pediococcus acidilactici M76 Producing Functional Exopolysaccharides and Its Lactic. Microorganisms 2021, 9, 1364. [Google Scholar] [PubMed]
- Wassenaar, T.M. Insights from 100 years of research with probiotic E. coli. Eur. J. Microbiol. Immunol. 2016, 6, 147–161. [Google Scholar] [CrossRef] [Green Version]
- Abid, R.; Waseem, H.; Ali, J.; Ghazanfar, S.; Ali, G.M.; Elasbali, A.M.; Alharethi, S.H. Probiotic Yeast Saccharomyces: Back to Nature to Improve Human Health. J. Fungi 2022, 8, 444. [Google Scholar] [CrossRef]
- Hanchi, H.; Mottawea, W.; Sebei, K.; Hammami, R. The genus Enterococcus: Between probiotic potential and safety concerns-an update. Front. Microbiol. 2018, 9, 1791. [Google Scholar] [CrossRef]
- Qi, Y.; Huang, L.; Zeng, Y.; Li, W.; Zhou, D.; Xie, J.; Xie, J.; Tu, Q.; Deng, D.; Yin, J. Pediococcus pentosaceus: Screening and Application as Probiotics in Food Processing. Front. Microbiol. 2021, 12, 3827. [Google Scholar] [CrossRef]
- Khan, R.; Petersen, F.C.; Shekhar, S. Commensal bacteria: An emerging player in defense against respiratory pathogens. Front. Immunol. 2019, 10, 1203. [Google Scholar] [CrossRef] [Green Version]
- Bidossi, A.; De Grandi, R.; Toscano, M.; Bottagisio, M.; De Vecchi, E.; Gelardi, M.; Drago, L. Probiotics Streptococcus salivarius 24SMB and Streptococcus oralis 89a interfere with biofilm formation of pathogens of the upper respiratory tract. BMC Infect. Dis. 2018, 18, 653. [Google Scholar] [CrossRef]
- Passali, D.; Passali, G.C.; Vesperini, E.; Cocca, S.; Visconti, I.C.; Ralli, M.; Bellussi, L.M. The efficacy and tolerability of Streptococcus salivarius 24SMB and Streptococcus oralis 89a administered as nasal spray in the treatment of recurrent upper respiratory tract infections in children. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 67–72. [Google Scholar] [PubMed]
- George Kerry, R.; Patra, J.K.; Gouda, S.; Park, Y.; Shin, H.S.; Das, G. Benefaction of probiotics for human health: A review. J. Food Drug Anal. 2018, 26, 927–939. [Google Scholar] [CrossRef] [Green Version]
- Pandey, K.R.; Naik, S.R.; Vakil, B.V. Probiotics, prebiotics and synbiotics- a review. J. Food Sci. Technol. 2015, 52, 7577–7587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vamanu, E.; Gatea, F. Correlations between Microbiota Bioactivity and Bioavailability of Functional Compounds: A Mini-Review. Biomedicines 2020, 8, 39. [Google Scholar] [CrossRef]
- Oak, S.J.; Jha, R. The effects of probiotics in lactose intolerance: A systematic review. Crit. Rev. Food Sci. Nutr. 2018, 59, 1675–1683. [Google Scholar] [CrossRef] [PubMed]
- Ding, S.; Yan, W.; Ma, Y.; Fang, J. The impact of probiotics on gut health via alternation of immune status of monogastric animals. Anim. Nutr. 2021, 7, 24–30. [Google Scholar] [CrossRef]
- Hevia, A.; Delgado, S.; Sánchez, B.; Margolles, A. Molecular players involved in the interaction between beneficial bacteria and the immune system. Front. Microbiol. 2015, 80, 1363–1376. [Google Scholar] [CrossRef] [Green Version]
- Boger, M.C.L.; van Bueren, A.L.; Dijkhuizen, L. Cross-Feeding among Probiotic Bacterial Strains on Prebiotic Inulin Involves the Extracellular exo-Inulinase of Lactobacillus paracasei Strain W20. Appl. Environ. Microbiol. 2018, 84, e01539-18. [Google Scholar] [CrossRef] [Green Version]
- Turroni, F.; Serafini, F.; Foroni, E.; Duranti, S.; Motherway, M.O.C.; Taverniti, V.; Mangifesta, M.; Milani, C.; Viappiani, A.; Roversi, T.; et al. Role of sortase-dependent pili of Bifidobacterium bifidum PRL2010 in modulating bacterium-host interactions. Proc. Natl. Acad. Sci. USA 2013, 110, 11151–11156. [Google Scholar] [CrossRef] [Green Version]
- Macpherson, A.J.; Harris, N.L. Interactions between commensal intestinal bacteria and the immune system. Nat. Rev. Immunol. 2004, 4, 478–485. [Google Scholar] [CrossRef]
- Chénard, T.; Prévost, K.; Dubé, J.; Massé, E. Immune System Modulations by Products of the Gut Microbiota. Vaccines 2020, 8, 461. [Google Scholar] [CrossRef]
- Cremon, C.; Barbaro, M.R.; Ventura, M.; Barbara, G. Pre- and probiotic overview. Curr. Opin. Pharmacol. 2018, 43, 87–92. [Google Scholar] [CrossRef]
- Du, T.; Lei, A.; Zhang, N.; Zhu, C. The Beneficial Role of Probiotic Lactobacillus in Respiratory Diseases. Front. Immunol. 2022, 13, 2582. [Google Scholar] [CrossRef]
- Kopacz, K.; Phadtare, S. Probiotics for the Prevention of Antibiotic-Associated Diarrhea. Healthcare 2022, 10, 1450. [Google Scholar] [CrossRef] [PubMed]
- Farah, N.; Chin, V.K.; Chong, P.P.; Lim, W.F.; Lim, C.W.; Basir, R.; Chang, S.K.; Lee, T.Y. Riboflavin as a promising antimicrobial agent? A multi-perspective review. Curr. Res. Microb. Sci. 2022, 3, 100111. [Google Scholar] [CrossRef]
- Sánchez, B.; Delgado, S.; Blanco-Míguez, A.; Lourenço, A.; Gueimonde, M.; Margolles, A. Probiotics, gut microbiota, and their influence on host health and disease. Mol. Nutr. Food Res. 2017, 61, 1600240. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Lin, J.; Xiao, L.; Zhang, X.; Zhao, L.; Wang, M.; Li, L. Gut microbiota in systemic lupus erythematosus: A fuse and a solution. J. Autoimmun. 2022, 132, 102867. [Google Scholar] [CrossRef]
- Ranjha, M.M.A.N.; Shafique, B.; Batool, M.; Kowalczewski, P.Ł.; Shehzad, Q.; Usman, M.; Manzoor, M.F.; Zahra, S.M.; Yaqub, S.; Aadil, R.M. Nutritional and Health Potential of Probiotics: A Review. Appl. Sci. 2021, 11, 11204. [Google Scholar] [CrossRef]
- Snigdha, S.; Ha, K.; Tsai, P.; Dinan, T.G.; Bartos, J.D.; Shahid, M. Probiotics: Potential novel therapeutics for microbiota-gut-brain axis dysfunction across gender and lifespan. Pharmacol. Ther. 2022, 231, 107978. [Google Scholar] [CrossRef] [PubMed]
- Balta, I.; Butucel, E.; Mohylyuk, V.; Criste, A.; Dezmirean, D.S.; Stef, L.; Pet, I.; Corcionivoschi, N. Novel Insights into the Role of Probiotics in Respiratory Infections, Allergies, Cancer, and Neurological Abnormalities. Diseases 2021, 9, 60. [Google Scholar] [CrossRef]
- Dang, A.T.; Marsland, B.J. Microbes, metabolites, and the gut–lung axis. Mucosal Immunol. 2019, 12, 843–850. [Google Scholar] [CrossRef] [Green Version]
- Banfi, D.; Moro, E.; Bosi, A.; Bistoletti, M.; Cerantola, S.; Crema, F.; Maggi, F.; Giron, M.C.; Giaroni, C.; Baj, A. Impact of Microbial Metabolites on Microbiota–Gut–Brain Axis in Inflammatory Bowel Disease. Int. J. Mol. Sci. 2021, 22, 1623. [Google Scholar] [CrossRef]
- Bistoletti, M.; Bosi, A.; Banfi, D.; Giaroni, C.; Baj, A. The microbiota-gut-brain axis: Focus on the fundamental communication pathways. Prog. Mol. Biol. Transl. Sci. 2020, 176, 43–110. [Google Scholar]
- Carabotti, M.; Scirocco, A.; Antonietta Maselli, M.; Severi, C. The gut-brain axis: Interactions between enteric microbiota, central and enteric nervous systems. Ann. Gastroenterol. 2015, 28, 203–209. [Google Scholar]
- Huffnagle, G.B.; Dickson, R.P.; Lukacs, N.W. The respiratory tract microbiome and lung inflammation: A two-way street. Mucosal Immunol. 2017, 10, 299. [Google Scholar] [CrossRef] [Green Version]
- Bulanda, E.; Wypych, T.P. Bypassing the Gut–Lung Axis via Microbial Metabolites: Implications for Chronic Respiratory Diseases. Front. Microbiol. 2022, 13, 1522. [Google Scholar] [CrossRef]
- Humayun Kober, A.K.M.; Rajoka, M.S.R.; Mehwish, H.M.; Villena, J.; Kitazawa, H. Immunomodulation Potential of Probiotics: A Novel Strategy for Improving Livestock Health, Immunity, and Productivity. Microorganisms 2022, 10, 388. [Google Scholar] [CrossRef]
- Vitini, E.; Alvares, S.; Medina, M.; Perdigon, G. Gut mucosal immunostimulation by lactic acid bacteria. Biocell 2000, 24, 223–232. [Google Scholar]
- Vinderola, G.; Matar, C.; Perdigon, G. Role of intestinal epithelial cells in immune effects mediated by gram-positive probiotic bacteria: Involvement of toll-like receptors. Clin. Diagn. Lab. Immunol. 2005, 12, 1075–1084. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, S.; Massami, S.M.; Martins, K.; Gierus, M.; Farid, A.; Azizi, N.; Uemura, R.; Omori, M.; Sueyoshi, M.; Yasuda, M. Effects of Probiotics on Growth and Immunity of Piglets. Animals 2022, 12, 1786. [Google Scholar]
- Li, A.; Wang, Y.; Li, Z.; Qamar, H.; Mehmood, K.; Zhang, L.; Liu, J.; Zhang, H.; Li, J. Probiotics isolated from yaks improves the growth performance, antioxidant activity, and cytokines related to immunity and inflammation in mice. Microb. Cell Fact. 2019, 18, 112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ménard, O.; Gafa, V.; Kapel, N.; Rodriguez, B.; Butel, M.J.; Waligora-Dupriet, A.J. Characterization of immunostimulatory CpG-rich sequences from different Bifidobacterium species. Appl. Environ. Microbiol. 2010, 76, 2846–2855. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bouvier, M.; Meance, S.; Bouley, C.; Berta, J.-L.; Grimaud, J.-C. Effects of Consumptionof a Milk Fermented by the Probiotic Strain Bifidobacterium animalis DN-173 010 on Colonic Transit Times in Healthy Humans. Biosci. Microflora 2001, 20, 43–48. [Google Scholar] [CrossRef] [Green Version]
- Ebrahimpour-Koujan, S.; Milajerdi, A.; Larijani, B.; Esmaillzadeh, A. Effects of probiotics on salivary cytokines and immunoglobulines: A systematic review and meta-analysis on clinical trials. Sci. Rep. 2020, 10, 11800. [Google Scholar] [CrossRef] [PubMed]
- Blais, L.L.; Montgomery, T.L.; Amiel, E.; Deming, P.B.; Krementsov, D.N. Probiotic and commensal gut microbial therapies in multiple sclerosis and its animal models: A comprehensive review. Gut Microbes 2021, 13, 1943289. [Google Scholar] [CrossRef]
- Hougee, S.; Vriesema, A.J.M.; Wijering, S.C.; Knippels, L.M.J.; Folkerts, G.; Nijkamp, F.P.; Knol, J.; Garssen, J. Oral Treatment with Probiotics Reduces Allergic Symptoms in Ovalbumin-Sensitized Mice: A Bacterial Strain Comparative Study. Int. Arch. Allergy Immunol. 2010, 151, 107–117. [Google Scholar] [CrossRef]
- Fanning, S.; Hall, L.J.; Cronin, M.; Zomer, A.; MacSharry, J.; Goulding, D.; Motherway, M.O.C.; Shanahan, F.; Nally, K.; Dougan, G.; et al. Bifidobacterial surface-exopolysaccharide facilitates commensal-host interaction through immune modulation and pathogen protection. Proc. Natl. Acad. Sci. USA 2012, 109, 2108–2113. [Google Scholar] [CrossRef] [Green Version]
- Hidalgo-Cantabrana, C.; Sánchez, B.; Milani, C.; Ventura, M.; Margolles, A.; Ruas-Madiedo, P. Genomic overview and biological functions of exopolysaccharide biosynthesis in Bifidobacterium spp. Appl. Environ. Microbiol. 2014, 80, 9–18. [Google Scholar] [CrossRef]
- Djaldetti, M.; Bessler, H. Probiotic strains modulate cytokine production and the immune interplay between human peripheral blood mononucear cells and colon cancer cells. FEMS Microbiol. Lett. 2017, 364, 14. [Google Scholar] [CrossRef] [Green Version]
- Seifati, S.M.; Zaker, E.; Fesahat, F.; Zare, F.; Hekmatimoghaddam, S. Modulatory Effect of Probiotics on Proinflammatory Cytokine Levels in Acrylamide-Treated Rats. Biochem. Res. Int. 2021, 2021, 2268770. [Google Scholar] [CrossRef]
- López, P.; González-Rodríguez, I.; Sánchez, B.; Ruas-Madiedo, P.; Suárez, A.; Margolles, A.; Gueimonde, M. Interaction of Bifidobacterium bifidum LMG13195 with HT29 Cells Influences regulatory-T-cell-associated chemokine receptor expression. Appl. Environ. Microbiol. 2012, 78, 2850–2857. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lyons, A.; O’Mahony, D.; O’Brien, F.; MacSharry, J.; Sheil, B.; Ceddia, M.; Russell, W.M.; Forsythe, P.; Bienenstock, J.; Kiely, B.; et al. Bacterial strain-specific induction of Foxp3+ T regulatory cells is protective in murine allergy models. Clin. Exp. Allergy 2010, 40, 811–819. [Google Scholar] [CrossRef] [PubMed]
- Taverniti, V.; Cesari, V.; Gargari, G.; Rossi, U.; Biddau, C.; Lecchi, C.; Fiore, W.; Arioli, S.; Toschi, I.; Guglielmetti, S. Probiotics Modulate Mouse Gut Microbiota and Influence Intestinal Immune and Serotonergic Gene Expression in a Site-Specific Fashion. Front. Microbiol. 2021, 12, 2336. [Google Scholar] [CrossRef] [PubMed]
- Arunachalam, K.; Gill, H.S.; Chandra, R.K. Enhancement of natural immune function by dietary consumption of Bifidobacterium lactis (HN019). Eur. J. Clin. Nutr. 2000, 54, 263–267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ivanov, D.; Emonet, C.; Foata, F.; Affolter, M.; Delley, M.; Fisseha, M.; Blum-Sperisen, S.; Kochhar, S.; Arigoni, F. A serpin from the gut bacterium Bifidobacterium longum inhibits eukaryotic elastase-like serine proteases. J. Biol. Chem. 2006, 281, 17246–17252. [Google Scholar] [CrossRef] [Green Version]
- McCarthy, J.; O’Mahony, L.; O’Callaghan, L.; Sheil, B.; Vaughan, E.E.; Fitzsimons, N.; Fitzgibbon, J.; O’Sullivan, G.C.; Kiely, B.; Collins, J.K.; et al. Double blind, placebo controlled trial of two probiotic strains in interleukin 10 knockout mice and mechanistic link with cytokine balance. Gut 2003, 52, 975–980. [Google Scholar] [CrossRef]
- Oh, J.Z.; Ravindran, R.; Chassaing, B.; Carvalho, F.A.; Maddur, M.S.; Bower, M.; Hakimpour, P.; Gill, K.P.; Nakaya, H.I.; Yarovinsky, F.; et al. TLR5-Mediated Sensing of Gut Microbiota Is Necessary for Antibody Responses to Seasonal Influenza Vaccination. Immunity 2014, 41, 478–492. [Google Scholar] [CrossRef] [Green Version]
- Abdulamir, A.S.; Zukhrufuz, M.; Hafidh, R.R.; Abu, F. The Role of Diet, Prebiotic and Probiotic in the Development and Management of Inflammatory Bowel Diseases (IBD). In Inflammatory Bowel Disease-Advances in Pathogenesis and Management; IntechOpen: Rijeka, Croatia, 2012. [Google Scholar]
- Dogi, C.A.; Weill, F.; Perdigón, G. Immune response of non-pathogenic Gram(+) and Gram(−) bacteria in inductive sites of the intestinal mucosa: Study of the pathway of signaling involved. Immunobiology 2010, 215, 60–69. [Google Scholar] [CrossRef]
- Kang, S.-J.; Yang, J.; Lee, N.-Y.; Lee, C.-H.; Park, I.-B.; Park, S.-W.; Lee, H.J.; Park, H.-W.; Yun, H.S.; Chun, T. Monitoring Cellular Immune Responses after Consumption of Selected Probiotics in Immunocompromised Mice. Food Sci. Anim. Resour. 2022, 42, 903–914. [Google Scholar] [CrossRef]
- Konstantinov, S.R.; Smidt, H.; De Vos, W.M.; Bruijns, S.C.M.; Singh, S.K.; Valence, F.; Molle, D.; Lortal, S.; Altermann, E.; Klaenhammer, T.R.; et al. S layer protein A of Lactobacillus acidophilus NCFM regulates immature dendritic cell and T cell functions. Proc. Natl. Acad. Sci. USA 2008, 105, 19474–19479. [Google Scholar] [CrossRef] [Green Version]
- Kourelis, A.; Zinonos, I.; Kakagianni, M.; Christidou, A.; Christoglou, N.; Yiannaki, E.; Testa, T.; Kotzamanidis, C.; Litopoulou-Tzanetaki, E.; Tzanetakis, N.; et al. Validation of the dorsal air pouch model to predict and examine immunostimulatory responses in the gut. J. Appl. Microbiol. 2010, 108, 274–284. [Google Scholar] [CrossRef] [PubMed]
- Jacouton, E.; Michel, M.L.; Torres-Maravilla, E.; Chain, F.; Langella, P.; Bermúdez-Humarán, L.G. Elucidating the immune-related mechanisms by which probiotic strain lactobacillus casei BL23 displays anti-tumoral properties. Front. Microbiol. 2019, 9, 3281. [Google Scholar] [CrossRef] [PubMed]
- Galdeano, C.M.; Perdigón, G. Role of viability of probiotic strains in their persistence in the gut and in mucosal immune stimulation. J. Appl. Microbiol. 2004, 97, 673–681. [Google Scholar] [CrossRef] [PubMed]
- Sichetti, M.; De Marco, S.; Pagiotti, R.; Traina, G.; Pietrella, D. Anti-inflammatory effect of multistrain probiotic formulation (L. rhamnosus, B. lactis, and B. longum). Nutrition 2018, 53, 95–102. [Google Scholar] [CrossRef] [PubMed]
- Maldonado Galdeano, C.; Perdigón, G. The probiotic bacterium Lactobacillus casei induces activation of the gut mucosal immune system through innate immunity. Clin. Vaccine Immunol. 2006, 13, 219–226. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.T.; Xu, H.; Ye, J.Z.; Wu, W.R.; Shi, D.; Fang, D.Q.; Liu, Y.; Li, L.J. Efficacy of Lactobacillus rhamnosus GG in treatment of acute pediatric diarrhea: A systematic review with meta-analysis. World J. Gastroenterol. 2019, 25, 4999–5016. [Google Scholar] [CrossRef]
- Dou, X.; Qiao, L.; Chang, J.; Yan, S.; Song, X.; Chen, Y.; Xu, Q.; Xu, C. Lactobacillus casei ATCC 393 and it’s metabolites alleviate dextran sulphate sodium-induced ulcerative colitis in mice through the NLRP3-(Caspase-1)/IL-1β pathway. Food Funct. 2021, 12, 12022–12035. [Google Scholar] [CrossRef]
- Rocha-Ramírez, L.M.; Pérez-Solano, R.A.; Castañón-Alonso, S.L.; Moreno Guerrero, S.S.; Ramírez Pacheco, A.; García Garibay, M.; Eslava, C. Probiotic Lactobacillus Strains Stimulate the Inflammatory Response and Activate Human Macrophages. J. Immunol. Res. 2017, 2017, 4607491. [Google Scholar] [CrossRef]
- Haller, D.; Bode, C.; Hammes, W.P.; Pfeifer, A.M.A.; Schiffrin, E.J.; Blum, S. Non-pathogenic bacteria elicit a differential cytokine response by intestinal epithelial cell/leucocyte co-cultures. Gut 2000, 47, 79–87. [Google Scholar] [CrossRef] [Green Version]
- Jaffar, N.; Okinaga, T.; Nishihara, T.; Maeda, T. Enhanced phagocytosis of Aggregatibacter actinomycetemcomitans cells by macrophages activated by a probiotic Lactobacillus strain. J. Dairy Sci. 2018, 101, 5789–5798. [Google Scholar] [CrossRef] [Green Version]
- Park, M.R.; Shin, M.; Mun, D.; Jeong, S.Y.; Jeong, D.Y.; Song, M.; Ko, G.; Unno, T.; Kim, Y.; Oh, S. Probiotic Lactobacillus fermentum strain JDFM216 improves cognitive behavior and modulates immune response with gut microbiota. Sci. Rep. 2020, 10, 21701. [Google Scholar] [CrossRef] [PubMed]
- Sakai, F.; Hosoya, T.; Ono-Ohmachi, A.; Ukibe, K.; Ogawa, A.; Moriya, T.; Kadooka, Y.; Shiozaki, T.; Nakagawa, H.; Nakayama, Y.; et al. Lactobacillus gasseri SBT2055 Induces TGF-β Expression in Dendritic Cells and Activates TLR2 Signal to Produce IgA in the Small Intestine. PLoS ONE 2014, 9, e105370. [Google Scholar] [CrossRef] [PubMed]
- Von Schillde, M.A.; Hörmannsperger, G.; Weiher, M.; Alpert, C.A.; Hahne, H.; Bäuerl, C.; Van Huynegem, K.; Steidler, L.; Hrncir, T.; Pérez-Martínez, G.; et al. Lactocepin secreted by Lactobacillus exerts anti-inflammatory effects by selectively degrading proinflammatory chemokines. Cell Host Microbe 2012, 11, 387–396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peña, J.A.; Rogers, A.B.; Ge, Z.; Ng, V.; Li, S.Y.; Fox, J.G.; Versalovic, J. Probiotic Lactobacillus spp. diminish Helicobacter hepaticus-induced inflammatory bowel disease in interleukin-10-deficient mice. Infect. Immun. 2005, 73, 912–920. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Hassi, H.O.; Mann, E.R.; Sanchez, B.; English, N.R.; Peake, S.T.C.; Landy, J.; Man, R.; Urdaci, M.; Hart, A.L.; Fernandez-Salazar, L.; et al. Altered human gut dendritic cell properties in ulcerative colitis are reversed by Lactobacillus plantarum extracellular encrypted peptide STp. Mol. Nutr. Food Res. 2014, 58, 1132–1143. [Google Scholar] [CrossRef] [Green Version]
- Grangette, C.; Nutten, S.; Palumbo, E.; Morath, S.; Hermann, C.; Dewulf, J.; Pot, B.; Hartung, T.; Hols, P.; Mercenier, A. Enhanced antiinflammatory capacity of a Lactobacillus plantarum mutant synthesizing modified teichoic acids. Proc. Natl. Acad. Sci. USA 2005, 102, 10321–10326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murofushi, Y.; Villena, J.; Morie, K.; Kanmani, P.; Tohno, M.; Shimazu, T.; Aso, H.; Suda, Y.; Hashiguchi, K.; Saito, T.; et al. The toll-like receptor family protein RP105/MD1 complex is involved in the immunoregulatory effect of exopolysaccharides from Lactobacillus plantarum N14. Mol. Immunol. 2015, 64, 63–75. [Google Scholar] [CrossRef] [PubMed]
- Anukam, K.C.; Osazuwa, E.O.; Osadolor, H.B.; Bruce, A.W.; Reid, G. Yogurt containing probiotic Lactobacillus rhamnosus GR-1 and L. reuteri RC-14 helps resolve moderate diarrhea and increases CD4 count in HIV/AIDS patients. J. Clin. Gastroenterol. 2008, 42, 239–243. [Google Scholar] [CrossRef]
- Feleszko, W.; Jaworska, J.; Rha, R.D.; Steinhausen, S.; Avagyan, A.; Jaudszus, A.; Ahrens, B.; Groneberg, D.A.; Wahn, U.; Hamelmann, E. Probiotic-induced suppression of allergic sensitization and airway inflammation is associated with an increase of T regulatory-dependent mechanisms in a murine model of asthma. Clin. Exp. Allergy 2007, 37, 498–505. [Google Scholar] [CrossRef]
- Azizah, N.; Qonitun, U.; Yudani, T.; Raras, M.; Prawiro, S.R. Probiotics Lactobacillus reuteri increase levels of β-Defensin1, sIgA and decrease number of Staphylococcus aureus bacteria colonies in vaginal mucosa on puerperal mice model infected with Staphylococcus aureus. J. Kebidanan 2020, 9, 71–80. [Google Scholar] [CrossRef]
- Shornikova, A.V.; Casas, I.A.; Mykkänen, H.; Salo, E.; Vesikari, T. Bacteriotherapy with Lactobacillus reuteri in rotavirus gastroenteritis. Pediatr. Infect. Dis. J. 1997, 16, 1103–1107. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Tian, X.; He, B.; Hoang, T.K.; Taylor, C.M.; Blanchard, E.; Freeborn, J.; Park, S.; Luo, M.; Couturier, J.; et al. Lactobacillus reuteri DSM 17938 feeding of healthy newborn mice regulates immune responses while modulating gut microbiota and boosting beneficial metabolites. Am. J. Physiol. Gastrointest. Liver Physiol. 2019, 317, G824–G838. [Google Scholar] [CrossRef] [PubMed]
- Livingston, M.; Loach, D.; Wilson, M.; Tannock, G.W.; Baird, M. Gut commensal Lactobacillus reuteri 100-23 stimulates an immunoregulatory response. Immunol. Cell Biol. 2010, 88, 99–102. [Google Scholar] [CrossRef] [PubMed]
- Pahumunto, N.; Duangnumsawang, Y.; Teanpaisan, R. Effects of potential probiotics on the expression of cytokines and human β-defensins in human gingival epithelial cells and in vivo efficacy in a dog model. Arch. Oral Biol. 2022, 142, 105513. [Google Scholar] [CrossRef] [PubMed]
- Velez, E.M.M.; Maldonado Galdeano, C.; Carmuega, E.; Weill, R.; Bibas Bonet, M.E.; Perdigón, G. Probiotic fermented milk consumption modulates the allergic process induced by ovoalbumin in mice. Br. J. Nutr. 2015, 114, 566–576. [Google Scholar] [CrossRef] [Green Version]
- Kwon, H.K.; Lee, C.G.; So, J.S.; Chae, C.S.; Hwang, J.S.; Sahoo, A.; Nam, J.H.; Rhee, J.H.; Hwang, K.C.; Im, S.H. Generation of regulatory dendritic cells and CD4+Foxp3 + T cells by probiotics administration suppresses immune disorders. Proc. Natl. Acad. Sci. USA 2010, 107, 2159–2164. [Google Scholar] [CrossRef] [Green Version]
- Ohshima-Terada, Y.; Higuchi, Y.; Kumagai, T.; Hagihara, A.; Nagata, M. Complementary effect of oral administration of Lactobacillus paracasei K71 on canine atopic dermatitis. Vet. Dermatol. 2015, 26, 350-e75. [Google Scholar] [CrossRef]
- Choi, S.H.; Oh, J.W.; Ryu, J.S.; Kim, H.M.; Im, S.H.; Kim, K.P.; Kim, M.K. IRT5 Probiotics Changes Immune Modulatory Protein Expression in the Extraorbital Lacrimal Glands of an Autoimmune Dry Eye Mouse Model. Investig. Ophthalmol. Vis. Sci. 2020, 61, 42. [Google Scholar] [CrossRef]
- D’Mello, C.; Ronaghan, N.; Zaheer, R.; Dicay, M.; Le, T.; MacNaughton, W.K.; Surrette, M.G.; Swain, M.G. Probiotics Improve Inflammation-Associated Sickness Behavior by Altering Communication between the Peripheral Immune System and the Brain. J. Neurosci. 2015, 35, 10821–10830. [Google Scholar] [CrossRef] [Green Version]
- Yousefi, B.; Eslami, M.; Ghasemian, A.; Kokhaei, P.; Salek Farrokhi, A.; Darabi, N. Probiotics importance and their immunomodulatory properties. J. Cell. Physiol. 2019, 234, 8008–8018. [Google Scholar] [CrossRef]
- Gutzeit, C.; Magri, G.; Cerutti, A. Intestinal IgA production and its role in host-microbe interaction. Immunol. Rev. 2014, 260, 76–85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caballero-Franco, C.; Keller, K.; De Simone, C.; Chadee, K. The VSL#3 probiotic formula induces mucin gene expression and secretion in colonic epithelial cells. Am. J. Physiol. Gastrointest. Liver Physiol. 2007, 292, 315–322. [Google Scholar]
- Monteagudo-Mera, A.; Rastall, R.A.; Gibson, G.R.; Charalampopoulos, D.; Chatzifragkou, A. Adhesion mechanisms mediated by probiotics and prebiotics and their potential impact on human health. Appl. Microbiol. Biotechnol. 2019, 103, 6463–6472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salminen, S.; Isolauri, E. Intestinal colonization, microbiota, and probiotics. J. Pediatr. 2006, 149, S115–S120. [Google Scholar] [CrossRef]
- Reid, G.; Younes, J.A.; Van Der Mei, H.C.; Gloor, G.B.; Knight, R.; Busscher, H.J. Microbiota restoration: Natural and supplemented recovery of human microbial communities. Nat. Rev. Microbiol. 2010, 9, 27–38. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, J.; Wu, C. Modulation of Gut Microbiota and Immune System by Probiotics, Pre-biotics, and Post-biotics. Front. Nutr. 2022, 8, 1155. [Google Scholar] [CrossRef]
- D’Angelo, C.; Reale, M.; Costantini, E. Microbiota and Probiotics in Health and HIV Infection. Nutrients 2017, 9, 615. [Google Scholar] [CrossRef] [Green Version]
- Reikvam, D.H.; Meyer-Myklestad, M.H.; Trøseid, M.; Stiksrud, B. Probiotics to manage inflammation in HIV infection. Curr. Opin. Infect. Dis. 2020, 33, 34–43. [Google Scholar] [CrossRef]
- LeClair, C.E.; McConnell, K.A. Rotavirus; StatPearls: Treasure Island, FL, USA, 2022. [Google Scholar]
- Guandalini, S. Probiotics for Children With Diarrhea. J. Clin. Gastroenterol. 2008, 42, S53–S57. [Google Scholar] [CrossRef]
- Kim, A.H.J.; Hogarty, M.P.; Harris, V.C.; Baldridge, M.T. The Complex Interactions Between Rotavirus and the Gut Microbiota. Front. Cell. Infect. Microbiol. 2021, 10, 820. [Google Scholar] [CrossRef]
- Cruchet, S.; Furnes, R.; Maruy, A.; Hebel, E.; Palacios, J.; Medina, F.; Ramirez, N.; Orsi, M.; Rondon, L.; Sdepanian, V.; et al. The Use of Probiotics in Pediatric Gastroenterology: A Review of the Literature and Recommendations by Latin-American Experts. Pediatr. Drugs 2015, 17, 199–216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lo Vecchio, A.; Nunziata, F.; Bruzzese, D.; Conelli, M.L.; Guarino, A. Rotavirus immunisation status affects the efficacy of Lacticaseibacillus rhamnosus GG for the treatment of children with acute diarrhoea: A meta-analysis. Benef. Microbes 2022, 13, 283–294. [Google Scholar] [CrossRef] [PubMed]
- Peroni, D.G.; Morelli, L. Probiotics as Adjuvants in Vaccine Strategy: Is There More Room for Improvement? Vaccines 2021, 9, 811. [Google Scholar] [CrossRef] [PubMed]
- Ciabattini, A.; Olivieri, R.; Lazzeri, E.; Medaglini, D. Role of the microbiota in the modulation of vaccine immune responses. Front. Microbiol. 2019, 10, 1305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vlasova, A.N.; Takanashi, S.; Miyazaki, A.; Rajashekara, G.; Saif, L.J. How the gut microbiome regulates host immune responses to viral vaccines. Curr. Opin. Virol. 2019, 37, 16–25. [Google Scholar] [CrossRef] [PubMed]
- Popov, J.; Caputi, V.; Nandeesha, N.; Rodriguez, D.A.; Pai, N. Microbiota-Immune Interactions in Ulcerative Colitis and Colitis Associated Cancer and Emerging Microbiota-Based Therapies. Int. J. Mol. Sci. 2021, 22, 11365. [Google Scholar] [CrossRef] [PubMed]
- Fava, F.; Danese, S. Intestinal microbiota in inflammatory bowel disease: Friend of foe? World J. Gastroenterol. 2011, 17, 557–566. [Google Scholar] [CrossRef]
- Kapoor, B.; Gulati, M.; Rani, P.; Gupta, R. Psoriasis: Interplay between dysbiosis and host immune system. Autoimmun. Rev. 2022, 21, 103169. [Google Scholar] [CrossRef]
- Travers, M.A.; Florent, I.; Kohl, L.; Grellier, P. Probiotics for the control of parasites: An overview. J. Parasitol. Res. 2011, 2011, 610769. [Google Scholar] [CrossRef]
- Yao, P.; Tan, F.; Gao, H.; Wang, L.; Yang, T.; Cheng, Y. Effects of probiotics on Toll-like receptor expression in ulcerative colitis rats induced by 2,4,6-trinitro-benzene sulfonic acid. Mol. Med. Rep. 2017, 15, 1973–1980. [Google Scholar] [CrossRef] [Green Version]
- Yiu, J.H.C.; Dorweiler, B.; Woo, C.W. Interaction between gut microbiota and toll-like receptor: From immunity to metabolism. J. Mol. Med. 2017, 95, 13–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reunanen, J.; von Ossowski, I.; Hendrickx, A.P.A.; Palva, A.; de Vosa, W.M. Characterization of the SpaCBA pilus fibers in the probiotic Lactobacillus rhamnosus GG. Appl. Environ. Microbiol. 2012, 78, 2337–2344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mazziotta, C.; Cervellera, C.F.; Lanzillotti, C.; Touzé, A.; Gaboriaud, P.; Tognon, M.; Martini, F. MicroRNA dysregulations in Merkel cell carcinoma: Molecular mechanisms and clinical applications. J. Med. Virol. 2022. [Google Scholar] [CrossRef] [PubMed]
- Wang, I.K.; Wu, Y.Y.; Yang, Y.F.; Ting, I.W.; Lin, C.C.; Yen, T.H.; Chen, J.H.; Wang, C.H.; Huang, C.C.; Lin, H.C. The effect of probiotics on serum levels of cytokine and endotoxin in peritoneal dialysis patients: A randomised, double-blind, placebo-controlled trial. Benef. Microbes 2015, 6, 423–430. [Google Scholar] [CrossRef] [PubMed]
- Angurana, S.K.; Bansal, A.; Singhi, S.; Aggarwal, R.; Jayashree, M.; Salaria, M.; Mangat, N.K. Evaluation of effect of probiotics on cytokine levels in critically Ill children with severe sepsis: A double-blind, placebo-controlled trial. Crit. Care Med. 2018, 46, 1656–1664. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Peng, Y.; Li, Z.; Christensen, B.; Heckmann, A.B.; Lagerqvist, C.; Stenlund, H.; Lonnerdal, B.; Hernell, O.; West, C.E. Serum cytokine patterns are modulated in infants fed formula with probiotics or milk fat globule membranes: A randomized controlled trial. PLoS ONE 2021, 16, e0251293. [Google Scholar] [CrossRef]
- Meir, D.; Chiya Moshe, L.; Hanna, B. Multi-Species Probiotic Modulates Cytokine Production and the Interplay between Immune and Colon Cancer Cells. OBM Hepatol. Gastroenterol. 2020, 4, 1–15. [Google Scholar]
- Wang, X.; Zhang, P.; Zhang, X. Probiotics Regulate Gut Microbiota: An Effective Method to Improve Immunity. Molecules 2021, 26, 6076. [Google Scholar] [CrossRef]
- Kawashima, T.; Ikari, N.; Kouchi, T.; Kowatari, Y.; Kubota, Y.; Shimojo, N.; Tsuji, N.M. The molecular mechanism for activating IgA production by Pediococcus acidilactici K15 and the clinical impact in a randomized trial. Sci. Rep. 2018, 8, 5065. [Google Scholar] [CrossRef] [Green Version]
- Bungau, S.G.; Behl, T.; Singh, A.; Sehgal, A.; Singh, S.; Chigurupati, S.; Vijayabalan, S.; Das, S.; Palanimuthu, V.R. Targeting Probiotics in Rheumatoid Arthritis. Nutrients 2021, 13, 3376. [Google Scholar] [CrossRef]
- Verrucci, M.; Iacobino, A.; Fattorini, L.; Marcoaldi, R.; Maggio, A.; Piccaro, G. Use of probiotics in medical devices applied to some common pathologies. Annali dell’Istituto Superiore di Sanità 2019, 55, 380–385. [Google Scholar]
- Borruel, N.; Carol, M.; Casellas, F.; Antolín, M.; De Lara, F.; Espín, E.; Naval, J.; Guarner, F.; Malagelada, J.R. Increased mucosal tumour necrosis factor α production in Crohn’s disease can be downregulated ex vivo by probiotic bacteria. Gut 2002, 51, 659–664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reséndiz-Albor, A.A.; Reina-Garfias, H.; Rojas-Hernández, S.; Jarillo-Luna, A.; Rivera-Aguilar, V.; Miliar-García, A.; Campos-Rodríguez, R. Regionalization of pIgR expression in the mucosa of mouse small intestine. Immunol. Lett. 2010, 128, 59–67. [Google Scholar] [CrossRef]
- Brunelli, L.; De Vitis, V.; Ferrari, R.; Minuzzo, M.; Fiore, W.; Jäger, R.; Taverniti, V.; Guglielmetti, S. In vitro assessment of the probiotic properties of an industrial preparation containing Lacticaseibacillus paracasei in the context of athlete health. Front. Pharmacol. 2022, 13, 857987. [Google Scholar] [CrossRef] [PubMed]
- Aghamohammad, S.; Sepehr, A.; Miri, S.T.; Najafi, S.; Rohani, M.; Pourshafiea, M.R. The effects of the probiotic cocktail on modulation of the NF-kB and JAK/STAT signaling pathways involved in the inflammatory response in bowel disease model. BMC Immunol. 2022, 23, 8. [Google Scholar] [CrossRef] [PubMed]
- Kaur, H.; Ali, S.A. Probiotics and gut microbiota: Mechanistic insights into gut immune homeostasis through TLR pathway regulation. Food Funct. 2022, 13, 7423–7447. [Google Scholar] [CrossRef]
- Guo, Y.; Wang, B.; Wang, T.; Gao, L.; Yang, Z.J.; Wang, F.F.; Shang, H.W.; Hua, R.; Xu, J.D. Biological characteristics of il-6 and related intestinal diseases. Int. J. Biol. Sci. 2020, 17, 204–219. [Google Scholar] [CrossRef]
- Savino, F.; Galliano, I.; Savino, A.; Daprà, V.; Montanari, P.; Calvi, C.; Bergallo, M. Lactobacillus reuteri DSM 17938 probiotics may increase CC-chemokine receptor 7 expression in infants treated with for colic. Front. Pediatr. 2019, 7, 292. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.; Mao, J.; Zhang, S. Effect of Intervention of Probiotics in Advance on Treg/Th17 in Premature Mice. Biomed Res. Int. 2022, 2022, 6131069. [Google Scholar] [CrossRef]
- Patel, S.; Shukla, R.; Goyal, A. Probiotics in valorization of innate immunity across various animal models. J. Funct. Foods 2015, 14, 549–561. [Google Scholar] [CrossRef]
- Tang, S.; Liu, J.; Xu, C.; Shang, D.; Chen, H.; Zhang, G. Effects of probiotics on the improvement and regulation of intestinal barrier dysfunction and immune imbalance in intra-abdominal infections (Review). Int. J. Funct. Nutr. 2021, 2, 12. [Google Scholar] [CrossRef]
- Mohamed, F.M.; Thabet, M.H.; Ali, M.F. The Use of Probiotics to Enhance Immunity of Broiler Chicken Against Some Intestinal Infection Pathogens. SVU-Int. J. Vet. Sci. 2019, 2, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Raabis, S.; Li, W.; Cersosimo, L. Effects and immune responses of probiotic treatment in ruminants. Vet. Immunol. Immunopathol. 2019, 208, 58–66. [Google Scholar] [CrossRef] [PubMed]
- Lei, S.; Twitchell, E.; Yuan, L. Pathogenesis, Immunity and the Role of Microbiome/Probiotics in Enteric Virus Infections in Humans and Animal Models. In Mechanisms Underlying Host-Microbiome Interactions in Pathophysiology of Human Diseases; Springer: Boston, MA, USA, 2018; pp. 55–78. [Google Scholar]
- Lunghi, B.; Morfini, M.; Martinelli, N.; Balestra, D.; Linari, S.; Frusconi, S.; Branchini, A.; Cervellera, C.F.; Marchetti, G.; Castaman, G.; et al. The Asialoglycoprotein Receptor Minor Subunit Gene Contributes to Pharmacokinetics of Factor VIII Concentrates in Hemophilia A. Thromb. Haemost. 2022, 122, 715–725. [Google Scholar] [CrossRef] [PubMed]
- Martín, R.; Chain, F.; Miquel, S.; Motta, J.-P.; Vergnolle, N.; Sokol, H.; Langella, P. Using murine colitis models to analyze probiotics-host interactions. FEMS Microbiol. Rev. 2017, 035, 49–70. [Google Scholar] [CrossRef]
- Chen, L.L.; Wang, X.H.; Cui, Y.; Lian, G.H.; Zhang, J.; Ouyang, C.H.; Lu, F.G. Therapeutic effects of four strains of probiotics on experimental colitis in mice. World J. Gastroenterol. 2009, 15, 321–327. [Google Scholar] [CrossRef]
- Xu, L.; Liu, B.; Huang, L.; Li, Z.; Cheng, Y.; Tian, Y.; Pan, G.; Li, H.; Xu, Y.; Wu, W.; et al. Probiotic Consortia and Their Metabolites Ameliorate the Symptoms of Inflammatory Bowel Diseases in a Colitis Mouse Model. Microbiol. Spectr. 2022, 10, e00657-22. [Google Scholar] [CrossRef]
- Novotny Núñez, I.; Maldonado Galdeano, C.; de LeBlanc, A.d.M.; Perdigón, G. Lactobacillus casei CRL 431 administration decreases inflammatory cytokines in a diet-induced obese mouse model. Nutrition 2015, 31, 1000–1007. [Google Scholar] [CrossRef]
- Aragón, F.; Carino, S.; Perdigón, G.; De Moreno de LeBlanc, A. The administration of milk fermented by the probiotic Lactobacillus casei CRL 431 exerts an immunomodulatory effect against a breast tumour in a mouse model. Immunobiology 2014, 219, 457–464. [Google Scholar] [CrossRef]
- Karamese, M.; Aydin, H.; Sengul, E.; Gelen, V.; Sevim, C.; Ustek, D.; Karakus, E. The Immunostimulatory Effect of Lactic Acid Bacteria in a Rat Model. Iran. J. Immunol. 2016, 13, 220–228. [Google Scholar]
- Galdeano, C.M.; de Leblanc, A.d.M.; Carmuega, E.; Weill, R.; Perdigón, G. Mechanisms involved in the immunostimulation by probiotic fermented milk. J. Dairy Res. 2009, 76, 446–454. [Google Scholar] [CrossRef]
- Valizadeh, S.; Majdi Seghinsara, A.; Maleki Chollou, K.; Bahadori, A.; Abbaszadeh, S.; Taghdir, M.; Behniafar, H.; Riahi, S.M. The efficacy of probiotics in experimental autoimmune encephalomyelitis (an animal model for MS): A systematic review and meta-analysis. Lett. Appl. Microbiol. 2021, 73, 408–417. [Google Scholar] [CrossRef] [PubMed]
- Frank, M.G.; Fonken, L.K.; Watkins, L.R.; Maier, S.F.; Lowry, C.A. Could Probiotics Be Used to Mitigate Neuroinflammation? ACS Chem. Neurosci. 2019, 10, 13–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Argenio, V.; Sarnataro, D. Probiotics, prebiotics and their role in Alzheimer’s disease. Neural Regen. Res. 2021, 16, 1768–1769. [Google Scholar] [CrossRef]
- Mora, D.; Filardi, R.; Arioli, S.; Boeren, S.; Aalvink, S.; de Vos, W.M. Development of omics-based protocols for the microbiological characterization of multi-strain formulations marketed as probiotics: The case of VSL#3. Microb. Biotechnol. 2019, 12, 1371–1386. [Google Scholar] [PubMed] [Green Version]
- Lalonde, R.; Strazielle, C. Probiotic effects on anxiety-like behavior in animal models. Rev. Neurosci. 2022, 33, 691–701. [Google Scholar] [CrossRef]
- Ting, N.L.N.; Lau, H.C.H.; Yu, J. Cancer pharmacomicrobiomics: Targeting microbiota to optimise cancer therapy outcomes. Gut 2022, 71, 1412–1425. [Google Scholar] [CrossRef]
- Prakash, S.; Nicoletti, C.; McAleer, J.P.; Su, H.; Huang, J.; Zhang, J.; Wang, X.; Jin, Z.; Zhang, P.; Sun, X. Effect of Probiotics on Respiratory Tract Allergic Disease and Gut Microbiota. Front. Nutr. 2022, 9, 821900. [Google Scholar]
- Casaro, M.B.; Thomas, A.M.; Mendes, E.; Fukumori, C.; Ribeiro, W.R.; Oliveira, F.A.; Crisma, A.R.; Murata, G.M.; Bizzarro, B.; Sá-Nunes, A.; et al. A probiotic has differential effects on allergic airway inflammation in A/J and C57BL/6 mice and is correlated with the gut microbiome. Microbiome 2021, 9, 134. [Google Scholar] [CrossRef]
- Dudek-Wicher, R.; Junka, A.; Paleczny, J.; Bartoszewicz, M. Clinical Trials of Probiotic Strains in Selected Disease Entities. Int. J. Microbiol. 2020, 2020, 8854119. [Google Scholar] [CrossRef]
- Generally Recognized as Safe (GRAS)|FDA. Available online: https://www.fda.gov/food/food-ingredients-packaging/generally-recognized-safe-gras (accessed on 3 October 2022).
- Szajewska, H.; Guarino, A.; Hojsak, I.; Indrio, F.; Kolacek, S.; Shamir, R.; Vandenplas, Y.; Weizman, Z. Use of probiotics for management of acute gastroenteritis: A position paper by the ESPGHAN working group for probiotics and prebiotics. J. Pediatr. Gastroenterol. Nutr. 2014, 58, 531–539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shane, A.L.; Mody, R.K.; Crump, J.A.; Tarr, P.I.; Steiner, T.S.; Kotloff, K.; Langley, J.M.; Wanke, C.; Warren, C.A.; Cheng, A.C.; et al. 2017 Infectious Diseases Society of America Clinical Practice Guidelines for the Diagnosis and Management of Infectious Diarrhea. Clin. Infect. Dis. 2017, 65, e45–e80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guarino, A.; Ashkenazi, S.; Gendrel, D.; Lo Vecchio, A.; Shamir, R.; Szajewska, H. European society for pediatric gastroenterology, hepatology, and nutrition/european society for pediatric infectious diseases evidence-based guidelines for the management of acute gastroenteritis in children in Europe: Update 2014. J. Pediatr. Gastroenterol. Nutr. 2014, 59, 132–152. [Google Scholar] [CrossRef] [PubMed]
- Su, G.L.; Ko, C.W.; Bercik, P.; Falck-Ytter, Y.; Sultan, S.; Weizman, A.V.; Morgan, R.L. AGA Clinical Practice Guidelines on the Role of Probiotics in the Management of Gastrointestinal Disorders. Gastroenterology 2020, 159, 697–705. [Google Scholar] [CrossRef]
- Preidis, G.A.; Weizman, A.V.; Kashyap, P.C.; Morgan, R.L. AGA Technical Review on the Role of Probiotics in the Management of Gastrointestinal Disorders. Gastroenterology 2020, 159, 708–738.e4. [Google Scholar] [CrossRef]
- Dronkers, T.M.G.; Ouwehand, A.C.; Rijkers, G.T. Global analysis of clinical trials with probiotics. Heliyon 2017, 6, e04467. [Google Scholar] [CrossRef]
- Brüssow, H.; Ontario, G.R.; London, C.; Merenstein, D. Open Peer Review Probiotics and prebiotics in clinical tests: An update [version 1; peer review: 2 approved]. F1000Research 2019, 8. [Google Scholar] [CrossRef] [Green Version]
- Rijkers, G.T.; Andriessen, S.Q.; van Overveld, F.J. Death and the Miser: Microbiota regulate the outcome of checkpoint inhibition immunotherapy. Expert Rev. Anticancer Ther. 2019, 19, 831–834. [Google Scholar] [CrossRef] [Green Version]
- Hasley, T.; Ologun, G.; Wargo, J.; Jenq, R.R. Uncovering the role of the gut microbiota in immune checkpoint blockade therapy: A mini-review. Semin Hematol. 2020, 57, 13–18. [Google Scholar]
- Wang, T.; Zheng, N.; Luo, Q.; Jiang, L.; He, B.; Yuan, X.; Shen, L. Probiotics lactobacillus reuteriabrogates immune checkpoint blockade-associated colitis by inhibiting group 3 innate lymphoid cells. Front. Immunol. 2019, 10, 1235. [Google Scholar] [CrossRef]
- Chiang, B.L.; Sheih, Y.H.; Wang, L.H.; Liao, C.K.; Gill, H.S. Enhancing immunity by dietary consumption of a probiotic lactic acid bacterium (Bifidobacterium lactis HN019): Optimization and definition of cellular immune responses. Eur. J. Clin. Nutr. 2000, 54, 849–855. [Google Scholar] [CrossRef] [PubMed]
- Childs, C.E.; Röytiö, H.; Alhoniemi, E.; Fekete, A.A.; Forssten, S.D.; Hudjec, N.; Lim, Y.N.; Steger, C.J.; Yaqoob, P.; Tuohy, K.M.; et al. Xylo-oligosaccharides alone or in synbiotic combination with Bifidobacterium animalis subsp. lactis induce bifidogenesis and modulate markers of immune function in healthy adults: A double-blind, placebo-controlled, randomised, factorial cross-over study. Br. J. Nutr. 2014, 111, 1945–1956. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prescott, S.L.; Wickens, K.; Westcott, L.; Jung, W.; Currie, H.; Black, P.N.; Stanley, T.; Mitchell, E.A.; Fitzharris, P.; Siebers, R.; et al. Supplementation with Lactobacillus rhamnosus or Bifidobacterium lactis probiotics in pregnancy increases cord blood interferon-γ and breast milk transforming growth factor-β and immunoglobin A detection. Clin. Exp. Allergy 2008, 38, 1606–1614. [Google Scholar] [CrossRef] [PubMed]
- Konieczna, P.; Groeger, D.; Ziegler, M.; Frei, R.; Ferstl, R.; Shanahan, F.; Quigley, E.M.M.; Kiely, B.; Akdis, C.A.; O’Mahony, L. Bifidobacterium infantis 35624 administration induces Foxp3 T regulatory cells in human peripheral blood: Potential role for myeloid and plasmacytoid dendritic cells. Gut 2012, 61, 354–366. [Google Scholar] [CrossRef]
- Groeger, D.; O’Mahony, L.; Murphy, E.F.; Bourke, J.F.; Dinan, T.G.; Kiely, B.; Shanahan, F.; Quigley, E.M.M. Bifidobacterium infantis 35624 modulates host inflammatory processes beyond the gut. Gut Microbes 2013, 4, 325–339. [Google Scholar] [CrossRef] [Green Version]
- Schiffrin, E.J.; Rochat, F.; Link-Amster, H.; Aeschlimann, J.M.; Donnet-Hughes, A. Immunomodulation of human blood cells following the ingestion of lactic acid bacteria. J. Dairy Sci. 1995, 78, 491–497. [Google Scholar] [CrossRef]
- Klein, A.; Friedrich, U.; Vogelsang, H.; Jahreis, G. Lactobacillus acidophilus 74-2 and Bifidobacterium animalis subsp lactis DGCC 420 modulate unspecific cellular immune response in healthy adults. Eur. J. Clin. Nutr. 2007, 62, 584–593. [Google Scholar] [CrossRef] [Green Version]
- Castex, M.; Durand, H.; Okeke, B. Issues with Industrial Probiotic Scale-up. Aquac. Nutr. Gut Health Probiotics Prebiotics 2014, 13, 347–359. [Google Scholar]
- Fenster, K.; Freeburg, B.; Hollard, C.; Wong, C.; Laursen, R.R.; Ouwehand, A.C. The Production and Delivery of Probiotics: A Review of a Practical Approach. Microorganisms 2019, 7, 83. [Google Scholar] [CrossRef] [Green Version]
- Chang, C.P.; Liew, S.L. Growth Medium Optimization for Biomass Production of a Probiotic Bacterium, Lactobacillus rhamnosus ATCC 7469. J. Food Biochem. 2013, 37, 536–543. [Google Scholar] [CrossRef]
- Dang, T.D.; Yong, C.C.; Rheem, S.; Oh, S. Optimizing the composition of the medium for the viable cells of Bifidobacterium animalis subsp. lactis JNU306 using response surface methodology. J. Anim. Sci. Technol. 2021, 63, 603–613. [Google Scholar] [CrossRef] [PubMed]
- Martín, M.J.; Lara-Villoslada, F.; Ruiz, M.A.; Morales, M.E. Microencapsulation of bacteria: A review of different technologies and their impact on the probiotic effects. Innov. Food Sci. Emerg. Technol. 2015, 27, 15–25. [Google Scholar] [CrossRef]
- Jankovic, I.; Sybesma, W.; Phothirath, P.; Ananta, E.; Mercenier, A. Application of probiotics in food products—Challenges and new approaches. Curr. Opin. Biotechnol. 2010, 21, 175–181. [Google Scholar] [CrossRef]
- Shah, N.P.; Ding, W.K.; Fallourd, M.J.; Leyer, G. Improving the Stability of Probiotic Bacteria in Model Fruit Juices Using Vitamins and Antioxidants. J. Food Sci. 2010, 75, M278–M282. [Google Scholar] [CrossRef]
- Heidebach, T.; Först, P.; Kulozik, U. Microencapsulation of Probiotic Cells for Food Applications. Crit. Rev. Food Sci. Nutr. 2012, 52, 291–311. [Google Scholar] [CrossRef] [PubMed]
- Damin, M.R.; Minowa, E.; AlcÂntara, M.R.; Oliveira, M.N. Effect of cold storage on culture viability and some rheological properties of fermented milk prepared with yogurt and probiotic bacteria. J. Text. Stud. 2008, 39, 40–55. [Google Scholar] [CrossRef]
- Gueimonde, M.; Sánchez, B. Enhancing probiotic stability in industrial processes. Microb. Ecol. Health Dis. 2012, 23, 18562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fayol-Messaoudi, D.; Berger, C.N.; Coconnier-Polter, M.H.; Liévin-Le Moal, V.; Servin, A.L. pH-, lactic acid-, and non-lactic acid-dependent activities of probiotic lactobacilli against Salmonella enterica serovar typhimurium. Appl. Environ. Microbiol. 2005, 71, 6008–6013. [Google Scholar] [CrossRef] [Green Version]
- Kolaček, S.; Hojsak, I.; Berni Canani, R.; Guarino, A.; Indrio, F.; Orel, R.; Pot, B.; Shamir, R.; Szajewska, H.; Vandenplas, Y.; et al. Commercial Probiotic Products: A Call for Improved Quality Control. A Position Paper by the ESPGHAN Working Group for Probiotics and Prebiotics. J. Pediatr. Gastroenterol. Nutr. 2017, 65, 117–124. [Google Scholar] [CrossRef]
- Andreoletti, O.; Lau Baggesen, D.; Bolton, D.; Butaye, P.; Cook, P.; Griffin, J.; Davies, R.; Fernández Escámez, P.S.; Griffin, J.; Hald, T.; et al. Scientific Opinion on the maintenance of the list of QPS biological agents intentionally added to food and feed (2013 update). EFSA J. 2013, 11, 3449. [Google Scholar]
- Koutsoumanis, K.; Allende, A.; Alvarez-Ordóñez, A.; Bolton, D.; Bover-Cid, S.; Chemaly, M.; Davies, R.; De Cesare, A.; Hilbert, F.; Lindqvist, R.; et al. Update of the list of QPS-recommended biological agents intentionally added to food or feed as notified to EFSA 15: Suitability of taxonomic units notified to EFSA until September 2021. EFSA J. 2022, 20, 7045. [Google Scholar]
- Arora, M.; Baldi, A. Regulatory categories of probiotics across the globe: A review representing existing and recommended categorization. Indian J. Med. Microbiol. 2015, 33, S2–S10. [Google Scholar] [CrossRef] [PubMed]
- Markowiak, P.; Ślizewska, K. Effects of Probiotics, Prebiotics, and Synbiotics on Human Health. Nutrients 2017, 9, 1021. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.-T.; Hu, Y.-F.; Lee, B.-H.; Huang, C.-Y.; Lin, Y.-R.; Huang, S.-N.; Chen, Y.-Y.; Chang, J.-J.; Nan, F.-H. Dietary of Lactobacillus paracasei and Bifidobacterium longum improve nonspecific immune responses, growth performance, and resistance against Vibrio parahaemolyticus in Penaeus vannamei. Fish Shellfish. Immunol. 2022, 128, 307–315. [Google Scholar] [CrossRef]
- Bilal, M.; Ashraf, S.; Zhao, X. Dietary Component-Induced Inflammation and Its Amelioration by Prebiotics, Probiotics, and Synbiotics. Front. Nutr. 2022, 9, 931458. [Google Scholar] [CrossRef]
- Siddique, F.; Akram, K.; Alghamdi, E.S.; Arshad, Q.; Siddique, A. The Immunomodulatory Role of Probiotics. In Prebiotics and Probiotics-From Food to Health; IntechOpen: Rijeka, Croatia, 2022. [Google Scholar]
- Amdekar, S.; Dwivedi, D.; Roy, P.; Kushwah, S.; Singh, V. Probiotics: Multifarious oral vaccine against infectious traumas. FEMS Immunol. Med. Microbiol. 2010, 58, 299–306. [Google Scholar] [CrossRef]
- Reid, G.; Bruce, A.W.; Fraser, N.; Heinemann, C.; Owen, J.; Henning, B. Oral probiotics can resolve urogenital infections. FEMS Immunol. Med. Microbiol. 2001, 30, 49–52. [Google Scholar] [CrossRef]
- Van Hoang, V.; Ochi, T.; Kurata, K.; Arita, Y.; Ogasahara, Y.; Enomoto, K. Nisin-induced expression of recombinant T cell epitopes of major Japanese cedar pollen allergens in Lactococcus lactis. Appl. Microbiol. Biotechnol. 2018, 102, 261–268. [Google Scholar] [CrossRef]
- Ye¸silyurt, N.Y.; Yılmaz, B.; Gagündüz, D.A.; Capasso, R.; Apostolopoulos, V. Involvement of Probiotics and Postbiotics in the Immune System Modulation. Biologics 2021, 1, 89–110. [Google Scholar] [CrossRef]
- Correale, J.; Hohlfeld, R.; Baranzini, S.E. The role of the gut microbiota in multiple sclerosis. Nat. Rev. Neurol. 2022, 18, 544–558. [Google Scholar] [CrossRef]
- e Silva, N.O.; de Brito, B.B.; da Silva, F.A.F.; Santos, M.L.C.; de Melo, F.F. Probiotics in inflammatory bowel disease: Does it work? World J. Meta-Anal. 2020, 8, 54–66. [Google Scholar] [CrossRef]
- Phan, J.; Nair, D.; Jain, S.; Montagne, T.; Flores, D.V.; Nguyen, A.; Dietsche, S.; Gombar, S.; Cotter, P. Alterations in Gut Microbiome Composition and Function in Irritable Bowel Syndrome and Increased Probiotic Abundance with Daily Supplementation. mSystems 2021, 6, e01215-21. [Google Scholar] [CrossRef] [PubMed]
- Didari, T.; Mozaffari, S.; Nikfar, S.; Abdollahi, M. Effectiveness of probiotics in irritable bowel syndrome: Updated systematic review with meta-analysis. World J. Gastroenterol. 2015, 21, 3072–3084. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Li, M.; Chen, Q.; Li, X.; Chen, L.; Dong, Z.; Zhu, W.; Yang, Y.; Liu, Z.; Chen, Q. Programmable probiotics modulate inflammation and gut microbiota for inflammatory bowel disease treatment after effective oral delivery. Nat. Commun. 2022, 13, 3432. [Google Scholar] [CrossRef]
- Mandal, A.; Sahi, P.K. Probiotics for Diarrhea in Children. J. Med. Res. Innov. 2017, 1, AV5–AV12. [Google Scholar] [CrossRef] [Green Version]
- Raheem, A.; Liang, L.; Zhang, G.; Cui, S. Modulatory Effects of Probiotics During Pathogenic Infections With Emphasis on Immune Regulation. Front. Immunol. 2021, 12, 616713. [Google Scholar] [CrossRef]
- Li, Y.; Ye, Z.; Zhu, J.; Fang, S.; Meng, L.; Zhou, C. Effects of Gut Microbiota on Host Adaptive Immunity Under Immune Homeostasis and Tumor Pathology State. Front. Immunol. 2022, 13, 807. [Google Scholar] [CrossRef]
- Huang, J.; Liu, W.; Kang, W.; He, Y.; Yang, R.; Mou, X.; Zhao, W. Effects of microbiota on anticancer drugs: Current knowledge and potential applications. eBioMedicine 2022, 83, 104197. [Google Scholar] [CrossRef]
- Górska, A.; Przystupski, D.; Niemczura, M.J.; Kulbacka, J. Probiotic Bacteria: A Promising Tool in Cancer Prevention and Therapy. Curr. Microbiol. 2019, 76, 939. [Google Scholar] [CrossRef] [Green Version]
- Lu, K.; Dong, S.; Wu, X.; Jin, R.; Chen, H. Probiotics in Cancer. Front. Oncol. 2021, 11, 408. [Google Scholar] [CrossRef]
- Plaza-Diaz, J.; Ruiz-Ojeda, F.J.; Gil-Campos, M.; Gil, A. Mechanisms of Action of Probiotics. Adv. Nutr. 2019, 10, S49–S66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Javanshir, N.; Hosseini, G.N.G.; Sadeghi, M.; Esmaeili, R.; Satarikia, F.; Ahmadian, G.; Allahyari, N. Evaluation of the Function of Probiotics, Emphasizing the Role of their Binding to the Intestinal Epithelium in the Stability and their Effects on the Immune System. Biol. Proced. Online 2021, 23, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.; Walker, A.R.; Chakraborty, B.; Kaspar, J.R.; Nascimento, M.M.; Burne, R.A. Novel probiotic mechanisms of the oral bacterium Streptococcus sp. A12 as explored with functional genomics. Appl. Environ. Microbiol. 2019, 85, e01335-19. [Google Scholar] [CrossRef] [PubMed]
- Mahesh, R.; Ilangovan, P.; Nongbri, D.; Suchiang, K. Probiotics Interactions and the Modulation of Major Signalling Pathways in Host Model Organism Caenorhabditis elegans. Indian J. Microbiol. 2021, 61, 404–416. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Probiotic Strains | Associated Health Benefits | Experimental Model | References |
|---|---|---|---|
| Bacillus | |||
| Bacillus mesentericus | Immunestimulation | in vivo | [121] |
| Bacillus subtilis | Attenuates inflammation, Immunestimulation | in vivo | [122] |
| Bacillus velezensis | Attenuates inflammation, Immunestimulation | in vivo | [122] |
| Bifidobacterium | |||
| Bifidobacterium | Immunestimulation | in vitro | [123] |
| Bifidobacterium animalis DN-173 010 | Immunestimulation | Elderly subjects | [124] |
| Bifidobacterium animalis | Immunestimulation (salivary cytokine release) | Healthy adults | [125] |
| Bifidobacterium animalis | Multiple sclerosis therapy | in vivo | [126] |
| Bifidobacterium animalis NumRes252/-253 | Improve lung function, Immunestimulation | in vivo | [127] |
| Bifidobacterium animalis subsp. Lactis | Imunnemodulation | in vitro | [98,128,129] |
| Bifidobacterium bifidum | Immunestimulation (cytokine release) | in vitro | [130] |
| Bifidobacterium breve | Immunestimulation (cytokine release) | in vivo | [131] |
| Bifidobacterium breve IPLA 20004 | Improves intestinal barrier function | in vitro | [132] |
| Bifidobacterium breve M-16V | Improves lung function, Immunestimulation | in vivo | [127] |
| Bifidobacterium breve AH1205 | Immunestimulation | in vivo | [133] |
| Bifidobacterium breve UCC2003 | Imunnemodulation | in vitro, in vivo | [98,128,129] |
| Bifidobacterium bifidum MIMBb23sg | Attenuates inflammation | in vivo | [134] |
| Bifidobacterium bifidum LMG13195 | Improves intestinal barrier function | in vitro | [132] |
| Bifidobacterium lactis HN019 | Immunestimulation | Elderly subjects | [135] |
| Bifidobacterium longum | Immunestimulation (cytokine release) | in vitro | [130] |
| Bifidobacterium longum | Imunnemodulation | in vitro | [136] |
| Bifidobacterium longum AH1206 | Immunestimulation | in vivo | [133] |
| Bifidobacterium infantis NumRes251 | Improves lung function, Immunestimulation | in vivo | [127] |
| Bifdobacterium infantis | Attenuates colitis, Immunestimulation (cytokine release) | in vivo, in vivo | [131,137] |
| Clostridium butyricum | Immunestimulation | in vivo | [121] |
| Escherichia coli | Immunestimulation to inactivated influenza vaccine | in vivo | [138] |
| Escherichia coli | Immunestimulation (cytokine release) | in vitro | [139] |
| Escherichia coli 129 | Imunnemodulation | in vivo | [140] |
| Escherichia coli 13-7 | Imunnemodulation | in vivo | [140] |
| Escherichia coli Nissle 1917 | Multiple sclerosis therapy | in vivo | [126] |
| Lactiplantibacillus plantarum CJLP243/-45/-W55-10 | Immunestimulation (cytokine release) | in vivo | [141] |
| Lactobacillus | |||
| Lactobacillus acidophilus | Immunestimulation (cytokine release) | in vitro, in vivo | [130,131] |
| Lactobacillus acidophilus | Immunestimulation (IgA-producing cells increase) | Healthy adults | [119] |
| Lactobacillus acidophilus CRL 1462 | Imunnemodulation | in vivo | [140] |
| Lactobacillus acidophilus A9 | Imunnemodulation | in vivo | [140] |
| Lactobacillus acidophilus NCFM | Immunestimulation (cytokine release) | in vitro | [142] |
| Lactobacillus acidophilus NCFB 1748 | Increased chemotaxis of polymorphonuclear cells | in vivo | [143] |
| Lactobacillus bulgaricus | Immunestimulation (cytokine release) | in vitro, in vivo | [131,139] |
| Lactobacillus casei BL23 | Antitumor proprieties | in vivo | [144] |
| Lactobacillus casei CRL 431 | Imunnemodulation | in vivo | [140] |
| Lactobacillus casei CRL 431 | Immunestimulation | in vitro | [120,145,146] |
| Lactobacillus casei | Immunestimulation (cytokine release) | in vitro, in vivo | [130,131,139,147] |
| Lactobacillus casei | Reduces symptoms of rotavirus diarrhea | Children with rotavirus diarrhea | [148] |
| Lactobacillus casei | Immunestimulation (IgA-producing cells increase) | Healthy adults | [119] |
| Lactobacillus casei ATCC 393 | Attenuates colitis, Immunestimulation | in vivo | [149] |
| Lactobacillus casei Shirota | Immunestimulation (salivary cytokine release) | Healthy adults | [125] |
| Lactobacillus casei IMAU60214 | Immunestimulation (cytokine release) | in vitro | [150] |
| Lactobacillus crispatus | Immunestimulation (cytokine release) | in vitro | [139] |
| Lactobacillus delbrueckii subsp. bulgaricus | Immunestimulation (IgA-producing cells increase) | Healthy adults | [119] |
| Lactobacillus johnsonii | Immunestimulation (cytokine release) | in vitro | [151] |
| Lactobacillus johnsonii NBRC 13952 | Immunestimulation (cytokine release) | in vitro | [152] |
| Lactobacillus fermentum | Immunestimulation (salivary cytokine release) | Healthy adults | [125] |
| Lactobacillus fermentum JDFM216 | Increases mouse behavior, Immunestimulation | in vivo | [153] |
| Lactobacillus gasseri SBT2055 | Immunestimulation (IgA-producing cells increase) | in vivo | [154] |
| Lactobacillus helveticus R389 | Immunestimulation | in vitro | [120] |
| Lactobacillus helveticus IMAU70129 | Immunestimulation (cytokine release) | in vitro | [150] |
| Lactobacillus lactis | Immunestimulation (IgA-producing cells increase) | Healthy adults | [119] |
| Lactobacillus salivarius | Attenuates colitis | in vivo | [137] |
| Lactobacillus paracasei | Multiple sclerosis therapy | in vivo | [126] |
| Lactobacillus paracasei | Immunestimulation | in vitro, in vivo | [155] |
| Lactobacillus paracasei CNCM I-1518 | Immunestimulation | in vitro, in vivo | [145] |
| Lactobacillus paracasei + reuteri | Attenuates inflammation and colitis | in vivo | [156] |
| Lactobacillus paracasei subsp. Paracasei | Increases chemotaxis of polymorphonuclear cells | in vivo | [143] |
| Lactobacillus plantarum | Immunestimulation (IgA-producing cells increase) | Healthy adults | [119] |
| Lactobacillus plantarum | Immunestimulation (CD40 and CD80 expression increase)in vitro | [157] | |
| Lactobacillus plantarum | Immunestimulation | in vitro, in vivo | [98,158] |
| Lactobacillus plantarum N14 | Immunestimulation | in vitro | [159] |
| Lactobacillus plantarum NumRes8 | Improve lung function, Immunestimulation | in vivo | [127] |
| Lactobacillus rhamnosus | Immunestimulation (cytokine release) | in vitro, in vivo | [130,131] |
| Lactobacillus rhamnosus | Immunestimulation (IgA-producing cells increase) | Healthy adults | [119] |
| Lactobacillus rhamnosus GR-1 | Immunestimulation | HIV/AIDS-affected patient | [160] |
| Lactobacillus rhamnosus GG | Reduces symptoms of rotavirus diarrhea | Children with rotavirus diarrhea | [148] |
| Lactobacillus rhamnosus GG | Immunestimulation (cytokine release) | in vitro | [150] |
| Lactobacillus rhamnosus KLSD | Immunestimulation (cytokine release) | in vitro | [150] |
| Lactobacillus rhamnosus NumRes6 | Improves lung function, Immunestimulation | in vivo | [127] |
| Lactobacillus rhamnosus | Immunestimulation (salivary cytokine release) | Healthy adults | [125] |
| Lactobacillus rhamnosus + lactis | Attenuates allergic disease | in vivo | [161] |
| Lactobacillus reuteri | Immunestimulation | in vitro | [162] |
| Lactobacillus reuteri RC-14 | Immunestimulation | HIV/AIDS-affected patient | [160] |
| Lactobacillus reuteri ATCC 55730 | Reduces symptoms of rotavirus diarrhea | Children with rotavirus diarrhea | [163] |
| Lactobacillus reuteri DSM 17938 | Gut microbiota diversity increase, Immunestimulation | in vivo | [164] |
| Lactobacillus reuteri 100-23 | Immunestimulation (cytokine release) | in vitro | [165] |
| Lactobacillus sakei | Immunestimulation (cytokine release) | in vitro | [151] |
| Lactocaseibacillus | |||
| Lacticaseibacillus paracasei SD1 | Immunestimulation (cytokine release) | in vitro | [166] |
| Lacticaseibacillus rhamnosus SD4 | Immunestimulation (cytokine release) | in vitro | [166] |
| Lacticaseibacillus rhamnosus SD11 | Immunestimulation (cytokine release) | in vitro | [166] |
| Limosilactobacillus fermentum SD7 | Immunestimulation (cytokine release) | in vitro | [166] |
| Prevotella histicola | Multiple sclerosis therapy | in vivo | [126] |
| Streptococcus | |||
| Streptococcus thermophilus | Immunestimulation (cytokine release) | in vitro, in vivo | [130,131] |
| Streptococcus thermophilus | Immunestimulation (IgA-producing cells increase) | Healthy adults | [119] |
| Streptococcus faecalis | Immunestimulation | in vivo | [121] |
| Probiotic Mixtures | |||
| Lactobacilli+ Streptococchi | Immunestimulation | in vivo | [167] |
| Lactobacilli+ Streptococchi + | |||
| Bifidobacteria | Imunnemodulation | in vitro | [168] |
| Bifidobacteria + Lactobacilli | Attenuates allergic disease | in vivo | [169] |
| IRT5 | Immunestimulation | in vivo | [170] |
| VSL#3 | Attenuates sickness behavior development | in vivo | [171] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mazziotta, C.; Tognon, M.; Martini, F.; Torreggiani, E.; Rotondo, J.C. Probiotics Mechanism of Action on Immune Cells and Beneficial Effects on Human Health. Cells 2023, 12, 184. https://doi.org/10.3390/cells12010184
Mazziotta C, Tognon M, Martini F, Torreggiani E, Rotondo JC. Probiotics Mechanism of Action on Immune Cells and Beneficial Effects on Human Health. Cells. 2023; 12(1):184. https://doi.org/10.3390/cells12010184
Chicago/Turabian StyleMazziotta, Chiara, Mauro Tognon, Fernanda Martini, Elena Torreggiani, and John Charles Rotondo. 2023. "Probiotics Mechanism of Action on Immune Cells and Beneficial Effects on Human Health" Cells 12, no. 1: 184. https://doi.org/10.3390/cells12010184