
Premature Macrophage Activation by Stored Red Blood Cell Transfusion Halts Liver Regeneration Post-Partial Hepatectomy in Rats

Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Experiments
2.1.1. RBC Collection and Storage
2.1.2. PHx, Controlled Bleeding and Resuscitation Procedures
2.1.3. Experimental Groups
2.1.4. KCs Depletion and HO-1 Inhibitor Administration
2.1.5. Isolation of Hepatic Monocytes by FACS Sorting
2.2. Histological Evaluation
2.2.1. Histology and Immunohistochemistry
2.2.2. Measurement of Cell Size
2.3. Bioinformatics Analysis
2.3.1. Gene Expression Profiling
2.3.2. qRT-PCR
2.3.3. Protein Extraction and Western Blotting
2.4. In Vitro Analysis
Co-Cultures of Human RBCs and Macrophages
2.5. Statistical Analysis
3. Results
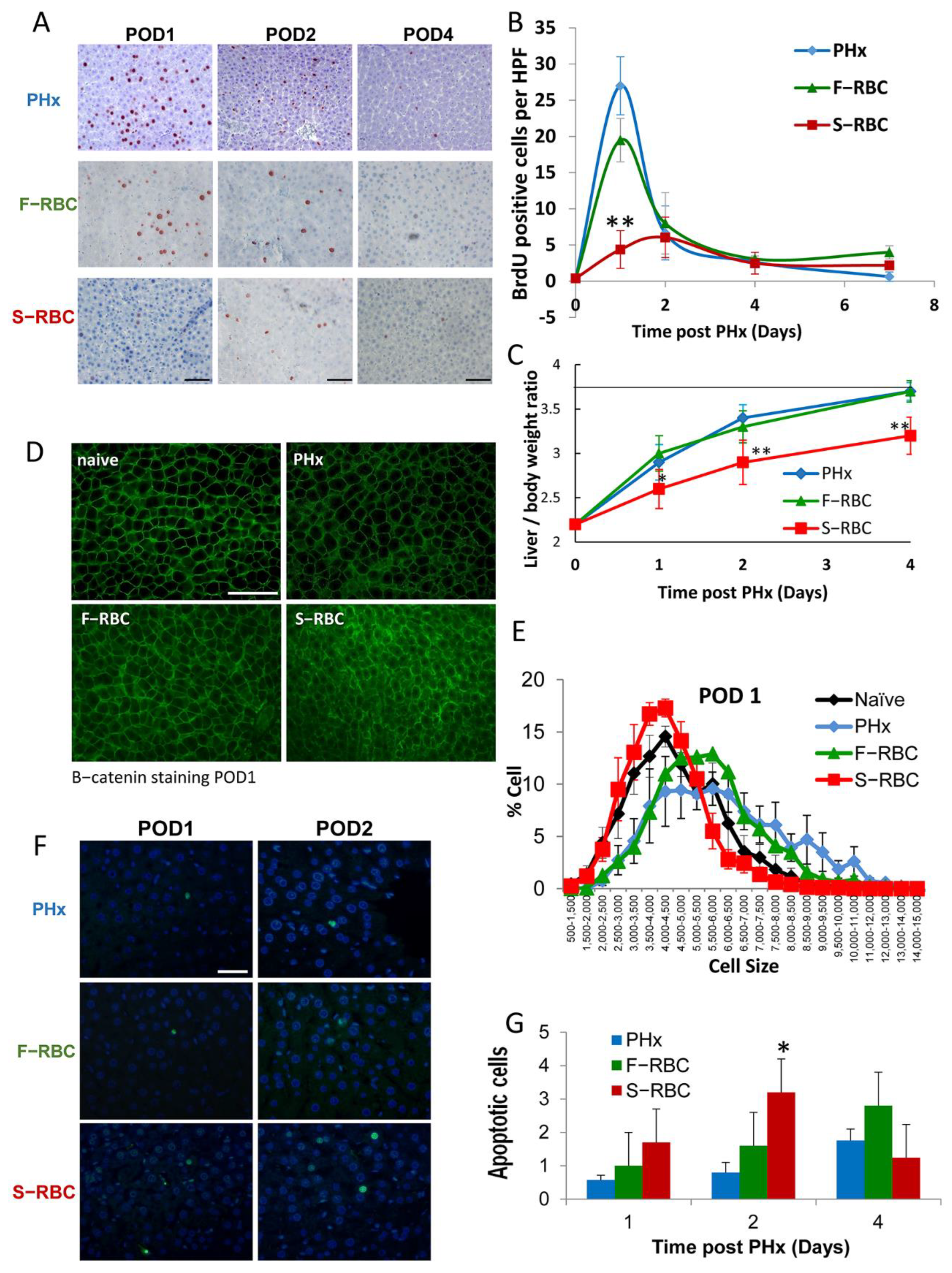
3.1. The Effect of RBC Storage Time on Hepatocyte Proliferation and Hypertrophy
3.2. Blood Count and Liver Enzymes following PHx and Blood Transfusion
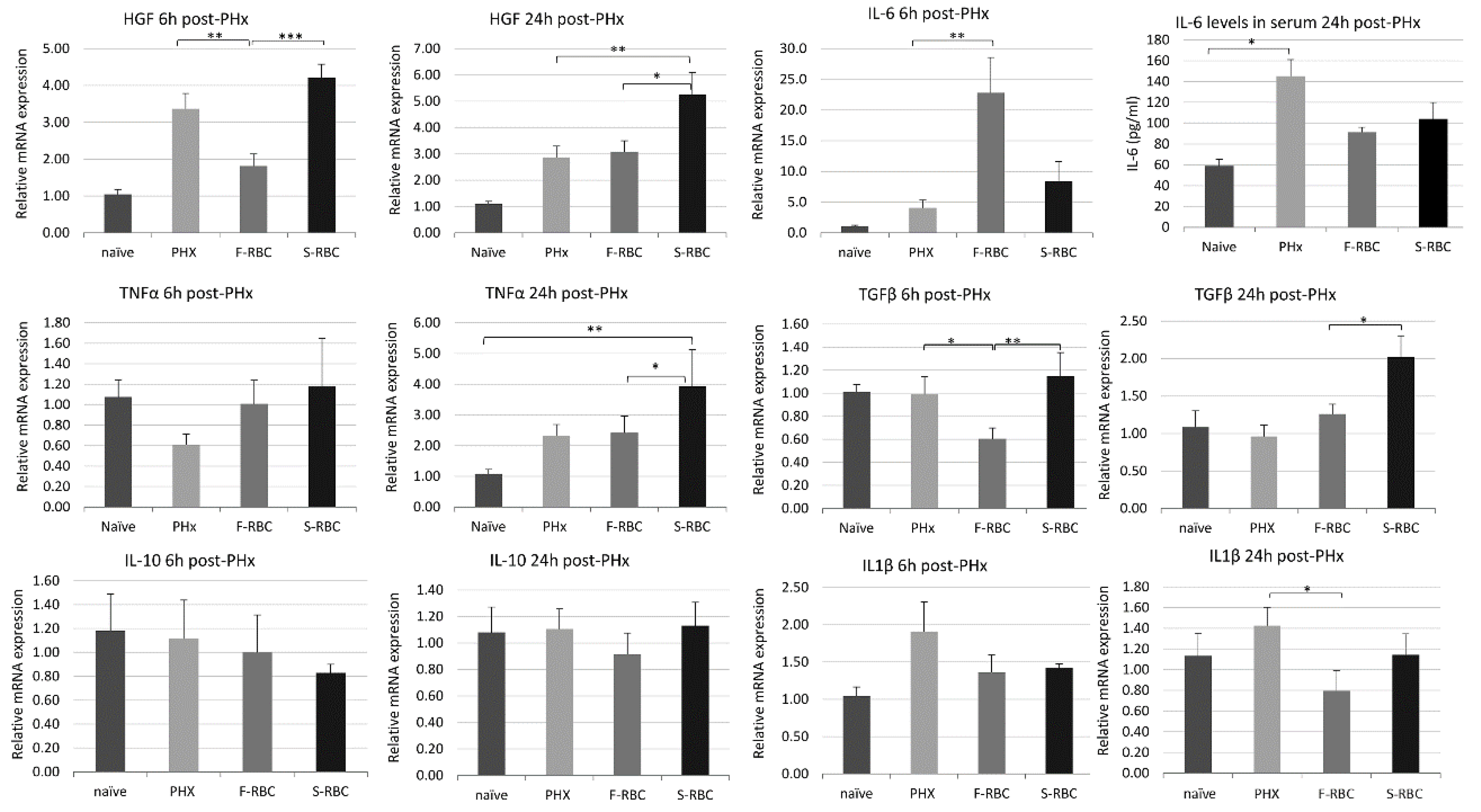
3.3. Initiation and Termination of Cytokine Levels in Livers following PHx and RBC Transfusion
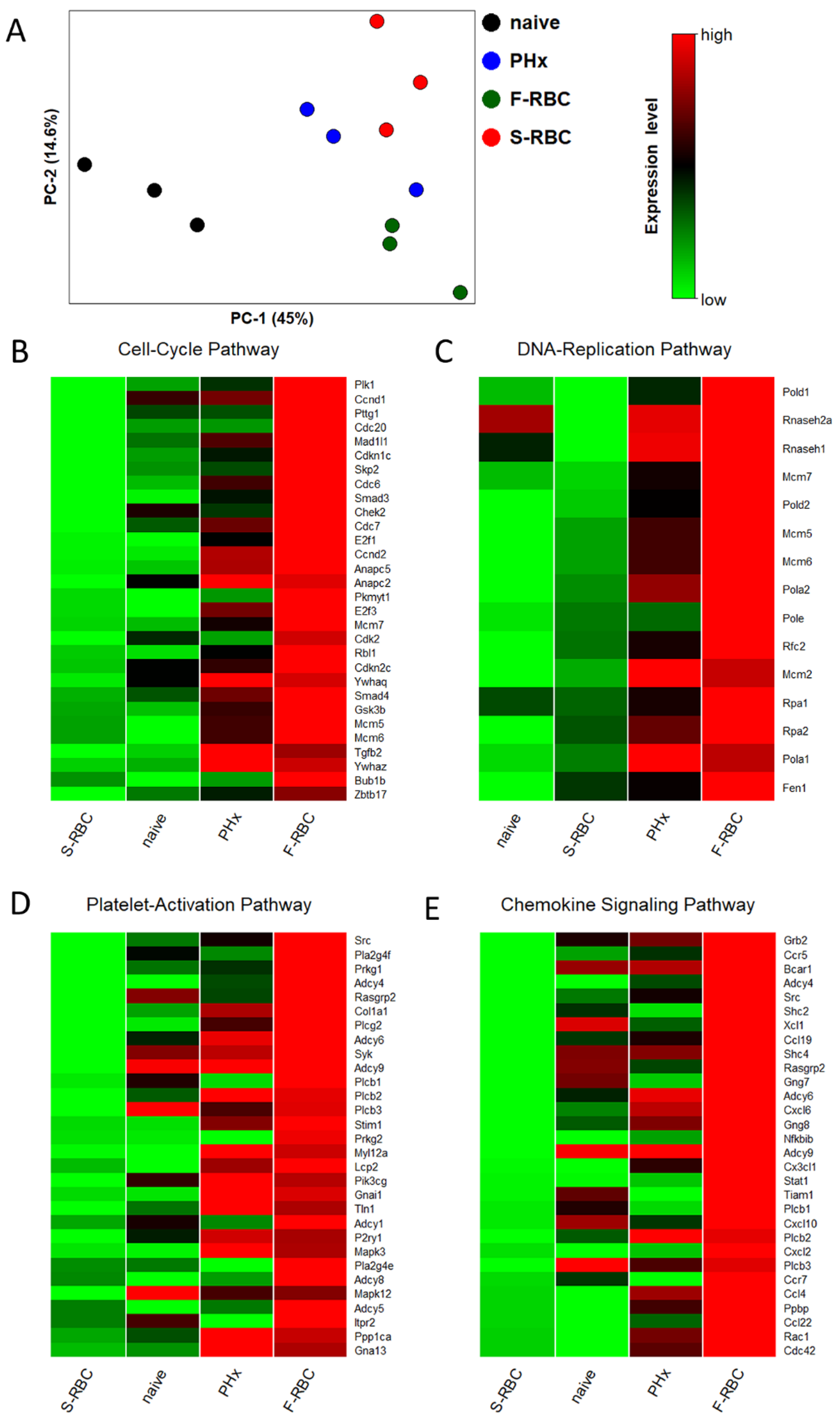
3.4. Storage Time of Transfused RBCs Affects Expression of Genes Associated with Tissue Regeneration
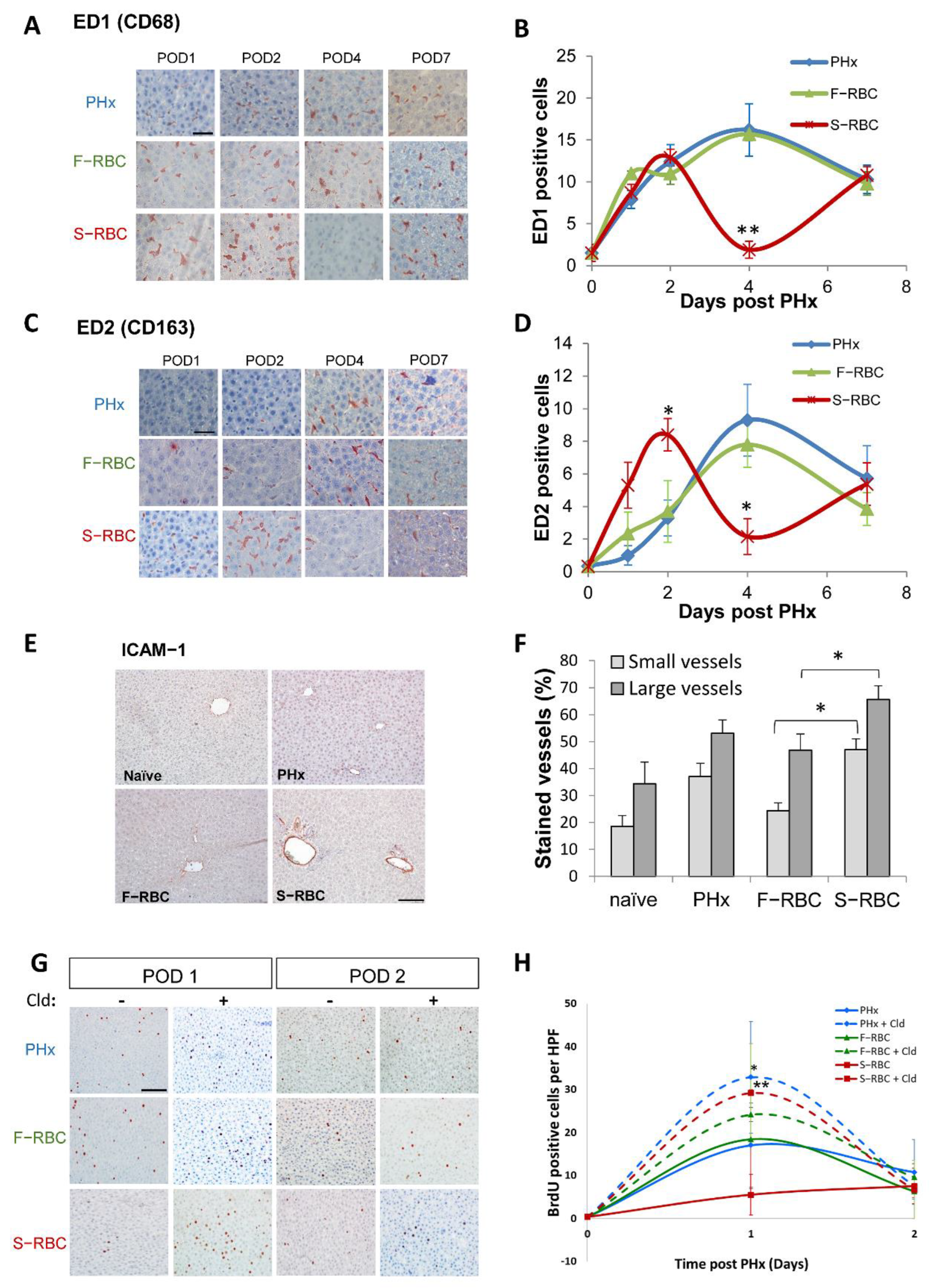
3.5. Expedited Macrophage Activation following Transfusion with S-RBCs
3.6. The Role of Macrophages in Liver Regeneration following PHx
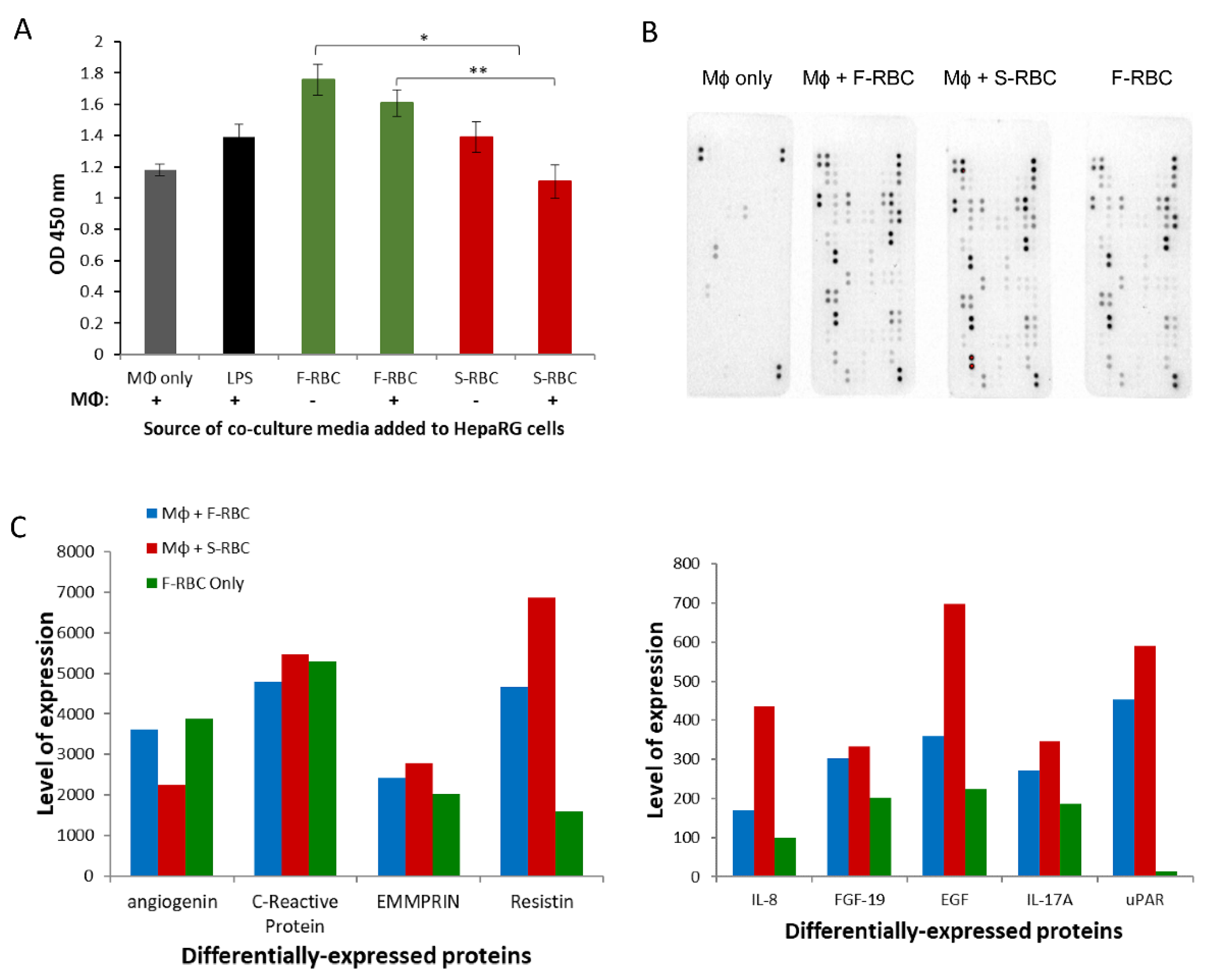
3.7. The Effect of Conditioned Media (CM) from Macrophage-RBC Co-Cultures on Hepatocyte Proliferation
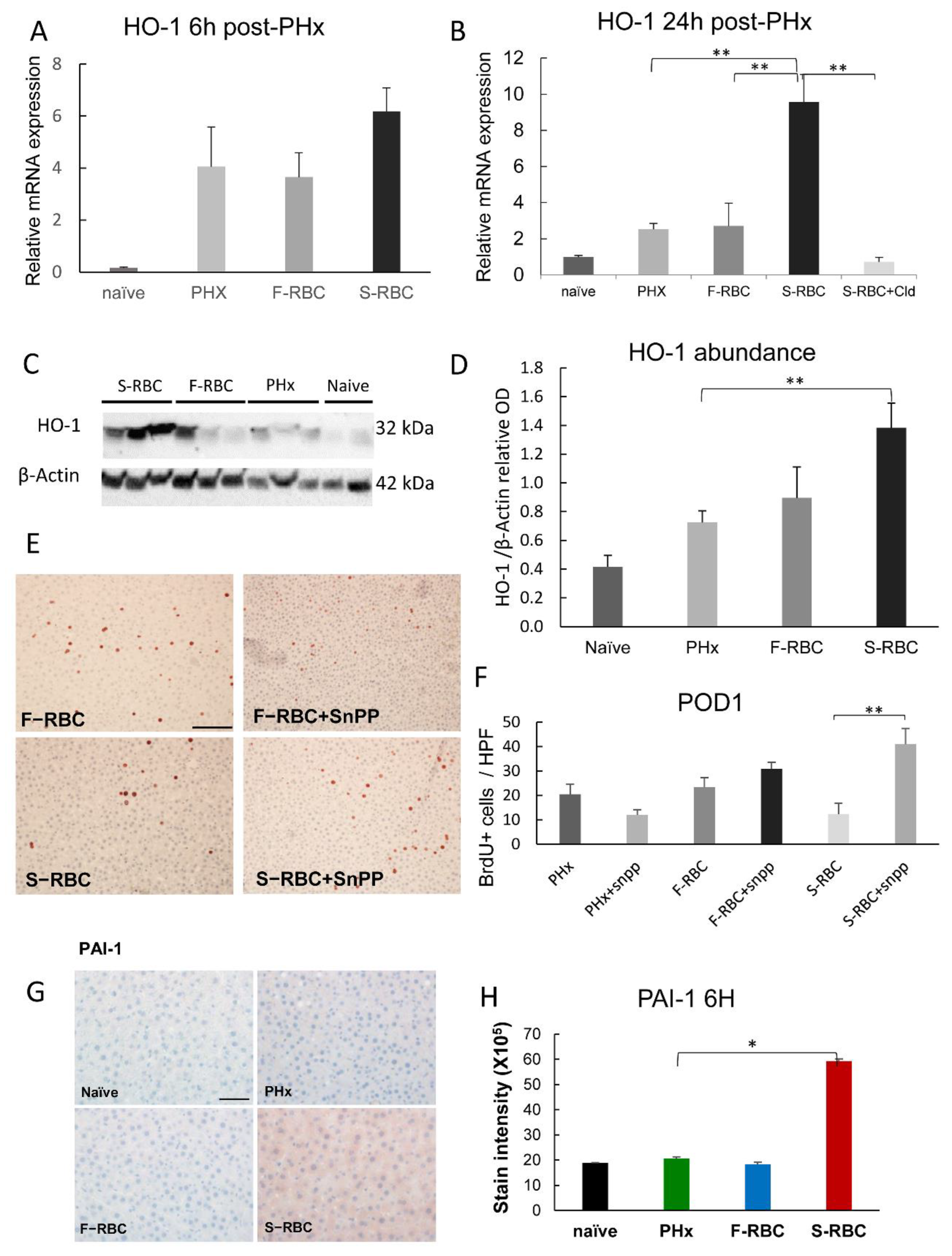
3.8. The Function of Heme Oxygenase-1 (HO-1) in Liver Regeneration
3.9. Elevated PAI-1 Expression following Transfusion with S-RBCs
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Forbes, S.J.; Newsome, P.N. Liver regeneration—mechanisms and models to clinical application. Nat. Rev. Gastroenterol. Hepatol. 2016, 13, 473–485. [Google Scholar] [CrossRef]
- Yang, T.; Zhang, J.; Lu, J.H.; Yang, G.S.; Wu, M.C.; Yu, W.F. Risk factors influencing postoperative outcomes of major hepatic resection of hepatocellular carcinoma for patients with underlying liver diseases. World J. Surg. 2011, 35, 2073–2082. [Google Scholar] [CrossRef]
- Michalopoulos, G.K.; Bhushan, B. Liver regeneration: Biological and pathological mechanisms and implications. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 40–55. [Google Scholar] [CrossRef]
- Clavien, P.A.; Petrowsky, H.; DeOliveira, M.L.; Graf, R. Strategies for safer liver surgery and partial liver transplantation. N. Engl. J. Med. 2007, 356, 1545–1559. [Google Scholar] [CrossRef]
- Feng, K.; Yan, J.; Li, X.; Xia, F.; Ma, K.; Wang, S.; Bie, P.; Dong, J. A randomized controlled trial of radiofrequency ablation and surgical resection in the treatment of small hepatocellular carcinoma. J. Hepatol. 2012, 57, 794–802. [Google Scholar] [CrossRef]
- Huang, J.; Yan, L.; Cheng, Z.; Wu, H.; Du, L.; Wang, J.; Xu, Y.; Zeng, Y. A randomized trial comparing radiofrequency ablation and surgical resection for HCC conforming to the Milan criteria. Ann. Surg. 2010, 252, 903–912. [Google Scholar] [CrossRef]
- Kopetz, S.; Chang, G.J.; Overman, M.J.; Eng, C.; Sargent, D.; Larson, D.W.; Grothey, A.; Vauthey, J.N.; Nagorney, D.M. Improved survival in metastatic colorectal cancer is associated with adoption of hepatic resection and improved chemotherapy. J. Clin. Oncol. 2009, 27, 3677–3683. [Google Scholar] [CrossRef]
- Petrowsky, H.; Fritsch, R.; Guckenberger, M.; De Oliveira, M.L.; Dutkowski, P.; Clavien, P.A. Modern therapeutic approaches for the treatment of malignant liver tumours. Nat. Rev. Gastroenterol. Hepatol. 2020, 17, 755–772. [Google Scholar] [CrossRef]
- Shirabe, K.; Kajiyama, K.; Harimoto, N.; Tsujita, E.; Wakiyama, S.; Maehara, Y. Risk factors for massive bleeding during major hepatectomy. World J. Surg. 2010, 34, 1555–1562. [Google Scholar] [CrossRef]
- Huntington, J.T.; Royall, N.A.; Schmidt, C.R. Minimizing blood loss during hepatectomy: A literature review. J. Surg. Oncol. 2014, 109, 81–88. [Google Scholar]
- Simon, E.R. Adenine in blood banking. Transfusion 1977, 17, 317–325. [Google Scholar] [CrossRef]
- Bennett-Guerrero, E.; Veldman, T.H.; Doctor, A.; Telen, M.J.; Ortel, T.L.; Reid, T.S.; Mulherin, M.A.; Zhu, H.; Buck, R.D.; Califf, R.M.; et al. Evolution of adverse changes in stored RBCs. Proc. Natl. Acad. Sci. USA 2007, 104, 17063–17068. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, T.; Prudent, M.; D’Alessandro, A. Red blood cell storage lesion: Causes and potential clinical consequences. Blood Transfus. 2019, 17, 27–52. [Google Scholar]
- Stack, G.; Baril, L.; Napychank, P.; Snyder, E.L. Cytokine generation in stored, white cell-reduced, and bacterially contaminated units of red cells. Transfusion 1995, 35, 199–203. [Google Scholar] [CrossRef]
- Koch, C.G.; Li, L.; Sessler, D.I.; Figueroa, P.; Hoeltge, G.A.; Mihaljevic, T.; Blackstone, E.H. Duration of red-cell storage and complications after cardiac surgery. N. Engl. J. Med. 2008, 358, 1229–1239. [Google Scholar] [CrossRef] [Green Version]
- Lelubre, C.; Piagnerelli, M.; Vincent, J.L. Association between duration of storage of transfused red blood cells and morbidity and mortality in adult patients: Myth or reality? Transfusion 2009, 49, 1384–1394. [Google Scholar] [CrossRef]
- Vamvakas, E.C. Meta-analysis of clinical studies of the purported deleterious effects of “old” (versus “fresh”) red blood cells: Are we at equipoise? Transfusion 2010, 50, 600–610. [Google Scholar] [CrossRef]
- Matot, I.; Cohen, K.; Pappo, O.; Barash, H.; Abramovitch, R. Liver response to hemorrhagic shock and subsequent resuscitation: MRI analysis. Shock 2008, 29, 16–24. [Google Scholar] [CrossRef]
- Matot, I.; Katz, M.; Pappo, O.; Zelig, O.; Corchia, N.; Yedgar, S.; Barshtein, G.; Guerrero, E.B.; Abramovitch, R. Resuscitation with aged blood exacerbates liver injury in a hemorrhagic rat model. Crit. Care Med. 2012, 41, 842–849. [Google Scholar] [CrossRef]
- Inoue, Y.; Ishii, M.; Fujii, K.; Kitada, K.; Kuramoto, T.; Takano, Y.; Matsuo, K.; Tomioka, A.; Yamaguchi, T.; Yokohama, K.; et al. The effects of allogeneic blood transfusion in hepatic resection. Am. Surg. 2021, 87, 228–234. [Google Scholar] [CrossRef]
- Lu, Q.; Zhang, J.; Gao, W.M.; Lv, Y.; Zhang, X.F.; Liu, X.M. Intraoperative Blood Transfusion and Postoperative Morbidity Following Liver Resection. Med. Sci. Monit. 2018, 24, 8469–8480. [Google Scholar] [CrossRef]
- Barash, H.; Gross, E.; Edrei, Y.; Pappo, O.; Spira, G.; Vlodavsky, I.; Galun, E.; Matot, I.; Abramovitch, R. Functional magnetic resonance imaging monitoring of pathological changes in rodent livers during hyperoxia and hypercapnia. Hepatology 2008, 48, 1232–1241. [Google Scholar] [CrossRef]
- Barash, H.; Gross, E.; Matot, I.; Edrei, Y.; Tsarfaty, G.; Spira, G.; Vlodavsky, I.; Galun, E.; Abramovitch, R. Functional MR imaging during hypercapnia and hyperoxia: Noninvasive tool for monitoring changes in liver perfusion and hemodynamics in a rat model. Radiology 2007, 243, 727–735. [Google Scholar] [CrossRef]
- Matot, I.; Nachmansson, N.; Duev, O.; Schulz, S.; Schroeder-Stein, K.; Frede, S.; Abramovitch, R. Impaired liver regeneration after hepatectomy and bleeding is associated with a shift from hepatocyte proliferation to hypertrophy. FASEB J. 2017, 31, 5283–5295. [Google Scholar] [CrossRef] [Green Version]
- Taub, R. Liver regeneration: From myth to mechanism. Nat. Rev. Mol. Cell Biol. 2004, 5, 836–847. [Google Scholar] [CrossRef]
- Meijer, C.; Wiezer, M.J.; Diehl, A.M.; Yang, S.-Q.; Schouten, H.J.; Meijer, S.; Van Rooijen, N.; Van Lambalgen, A.A.; Dijkstra, C.D.; Van Leeuwen, P.A.M. Kupffer cell depletion by CI2MDP-liposomes alters hepatic cytokine expression and delays liver regeneration after partial hepatectomy. Liver 2000, 20, 66–77. [Google Scholar] [CrossRef]
- Rai, R.M.; Yang, S.Q.; McClain, C.; Karp, C.L.; Klein, A.S.; Diehl, A.M. Kupffer cell depletion by gadolinium chloride enhances liver regeneration after partial hepatectomy in rats. Am. J. Physiol. 1996, 270, G909–G918. [Google Scholar] [CrossRef]
- Xu, C.S.; Jiang, Y.; Zhang, L.X.; Chang, C.F.; Wang, G.P.; Shi, R.J.; Yang, Y.J. The role of Kupffer cells in rat liver regeneration revealed by cell-specific microarray analysis. J. Cell. Biochem. 2012, 113, 229–237. [Google Scholar] [CrossRef]
- Selzner, N.; Selzner, M.; Odermatt, B.; Tian, Y.; Van Rooijen, N.; Clavien, P.A. ICAM-1 triggers liver regeneration through leukocyte recruitment and Kupffer cell-dependent release of TNF-alpha/IL-6 in mice. Gastroenterology 2003, 124, 692–700. [Google Scholar] [CrossRef]
- Lee, S.H.; Kim, K.W.; Min, K.M.; Kim, K.W.; Chang, S.I.; Kim, J.C. Angiogenin reduces immune inflammation via inhibition of TANK-binding kinase 1 expression in human corneal fibroblast cells. Mediat. Inflamm. 2014, 2014, 861435. [Google Scholar] [CrossRef] [Green Version]
- Blandino-Rosano, M.; Perez-Arana, G.; Mellado-Gil, J.M.; Segundo, C.; Aguilar-Diosdado, M. Anti-proliferative effect of pro-inflammatory cytokines in cultured beta cells is associated with extracellular signal-regulated kinase 1/2 pathway inhibition: Protective role of glucagon-like peptide-1. J. Mol. Endocrinol. 2008, 41, 35–44. [Google Scholar] [CrossRef] [Green Version]
- Martinez, F.O.; Gordon, S. The M1 and M2 paradigm of macrophage activation: Time for reassessment. F1000Prime Rep. 2014, 6, 13. [Google Scholar] [CrossRef]
- Hammond, J.S.; Guha, I.N.; Beckingham, I.J.; Lobo, D.N. Prediction, prevention and management of postresection liver failure. Br. J. Surg. 2011, 98, 1188–1200. [Google Scholar] [CrossRef]
- Jarnagin, W.R.; Gonen, M.; Fong, Y.; DeMatteo, R.P.; Ben-Porat, L.; Little, S.; Corvera, C.; Weber, S.; Blumgart, L.H. Improvement in perioperative outcome after hepatic resection: Analysis of 1803 consecutive cases over the past decade. Ann. Surg. 2002, 236, 397–407. [Google Scholar] [CrossRef]
- García-Roa, M.; Vicente-Ayuso, M.D.C.; Bobes, A.M.; Pedraza, A.C.; González-Fernández, A.; Martín, M.P.; Sáez, I.; Seghatchian, J.; Gutiérrez, L. Red blood cell storage time and transfusion: Current practice, concerns and future perspectives. Blood Transfus 2017, 15, 222–231. [Google Scholar]
- Elchaninov, A.V.; Fatkhudinov, T.K.; Vishnyakova, P.A.; Lokhonina, A.V.; Sukhikh, G.T. Phenotypical and functional polymorphism of liver resident macrophages. Cells 2019, 8, 1032. [Google Scholar] [CrossRef] [Green Version]
- Abshagen, K.; Eipel, C.; Kalff, J.C.; Menger, M.D.; Vollmar, B. Loss of NF-kappaB activation in Kupffer cell-depleted mice impairs liver regeneration after partial hepatectomy. Am. J. Physiol. Gastrointest. Liver Physiol. 2007, 292, G1570–G1577. [Google Scholar] [CrossRef] [Green Version]
- Araujo, J.A.; Zhang, M.; Yin, F. Heme oxygenase-1, oxidation, inflammation, and atherosclerosis. Front. Pharmacol 2012, 3, 119. [Google Scholar] [CrossRef] [Green Version]
- Naito, Y.; Takagi, T.; Higashimura, Y. Heme oxygenase-1 and anti-inflammatory M2 macrophages. Arch. Biochem. Biophys. 2014, 564, 83–88. [Google Scholar] [CrossRef] [Green Version]
- Pibiri, M.; Leoni, V.P.; Atzori, L. Heme oxygenase-1 inhibitor tin-protoporphyrin improves liver regeneration after partial hepatectomy. Life Sci. 2018, 204, 9–14. [Google Scholar] [CrossRef]
- Roselli, H.T.; Su, M.; Washington, K.; Kerins, D.M.; Vaughan, D.E.; Russell, W.E. Liver regeneration is transiently impaired in urokinase-deficient mice. Am. J. Physiol. 1998, 275, G1472–G1479. [Google Scholar] [CrossRef]
- Yamashita, M.; Darlington, D.N.; Weeks, E.J.; Jones, R.O.; Gann, D.S. Plasminogen activator inhibitor-1 rises after hemorrhage in rats. Am. J. Physiol. 1995, 268, E1065–E1069. [Google Scholar] [CrossRef]
- Long, X.; Schafer, A.I. Inhibition of plasminogen activator inhibitor-1 expression in vascular smooth muscle cells by protoporphyrins through a heme oxygenase-independent mechanism. Mol. Cell. Biochem. 2008, 312, 93–101. [Google Scholar] [CrossRef]
- Boyle, J.J.; Harrington, H.A.; Piper, E.; Elderfield, K.; Stark, J.; Landis, R.C.; Haskard, D.O. Coronary intraplaque hemorrhage evokes a novel atheroprotective macrophage phenotype. Am. J. Pathol. 2009, 174, 1097–1108. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target | Forward | Reverse |
|---|---|---|
| HGF | GATCAGGACCTTGTGAGGGA | ATGGCACATCCACGACCA |
| IL-6 | CTTCCAAACTGGATATAACCAGGAA | CTTCACAAACTCCAGGTAGAAACG |
| TNFα | CCCAGACCCTCACACTCAGATC | CTCCGCTTGGTGGTTTGCTA |
| TGFβ | GATACGCCTGAGTGGCTGTCT | CGAAAGCCCTGTATTCCGTCT |
| IL1β | GCTGTGGCAGCTACCTATGTCTT | GTCACAGAGGACGGGCTCTTC |
| IL10 | GATACAGCTGCGACGCTGTCA | CCTTGTAGACACCTTTGTCTTGGA |
| HO-1 | GTCCAGGGAAGGCTTTAAGCT | GGCATAGACTGGGTTCTGCTT |
| HPRT | GAGCACTTCAGGGATTTGAATCAT | GTAGATTCAACTTGCCGCTGTCT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abudi, N.; Duev, O.; Asraf, T.; Blank, S.; Matot, I.; Abramovitch, R. Premature Macrophage Activation by Stored Red Blood Cell Transfusion Halts Liver Regeneration Post-Partial Hepatectomy in Rats. Cells 2022, 11, 3522. https://doi.org/10.3390/cells11213522
Abudi N, Duev O, Asraf T, Blank S, Matot I, Abramovitch R. Premature Macrophage Activation by Stored Red Blood Cell Transfusion Halts Liver Regeneration Post-Partial Hepatectomy in Rats. Cells. 2022; 11(21):3522. https://doi.org/10.3390/cells11213522
Chicago/Turabian StyleAbudi, Nathalie, Omri Duev, Tal Asraf, Simcha Blank, Idit Matot, and Rinat Abramovitch. 2022. "Premature Macrophage Activation by Stored Red Blood Cell Transfusion Halts Liver Regeneration Post-Partial Hepatectomy in Rats" Cells 11, no. 21: 3522. https://doi.org/10.3390/cells11213522