
Multi-Channel Metabolomics Analysis Identifies Novel Metabolite Biomarkers for the Early Detection of Fatty Liver Disease in Dairy Cows

Abstract
:1. Introduction
2. Materials and Methods
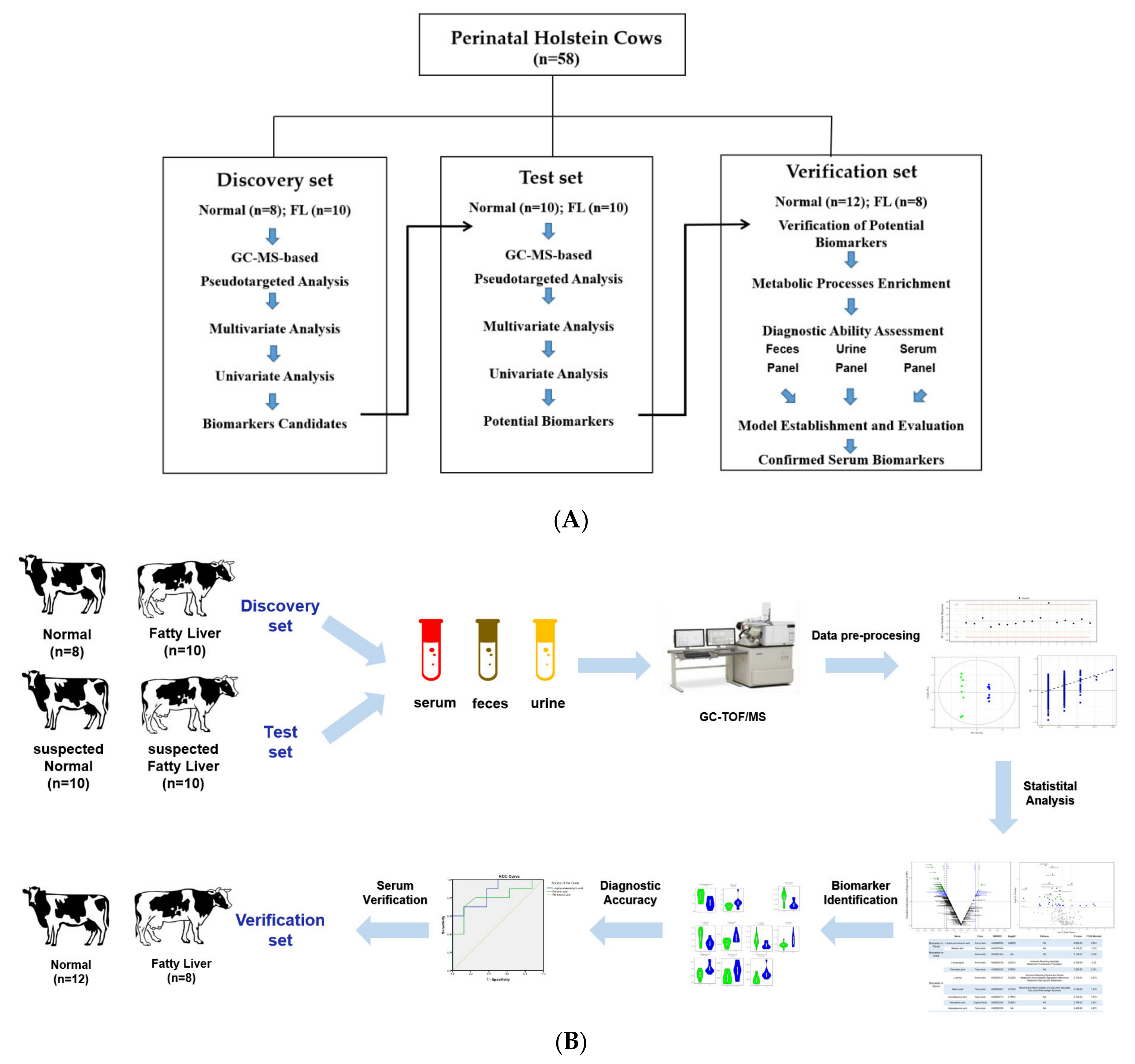
2.1. Study Design and Animal Participants
2.2. Serum, Urine, and Feces Sample Collection and Serum Biochemical Indicator Measurement
2.3. Sample Pre-Treatment and Non-Target GC–MS Analysis
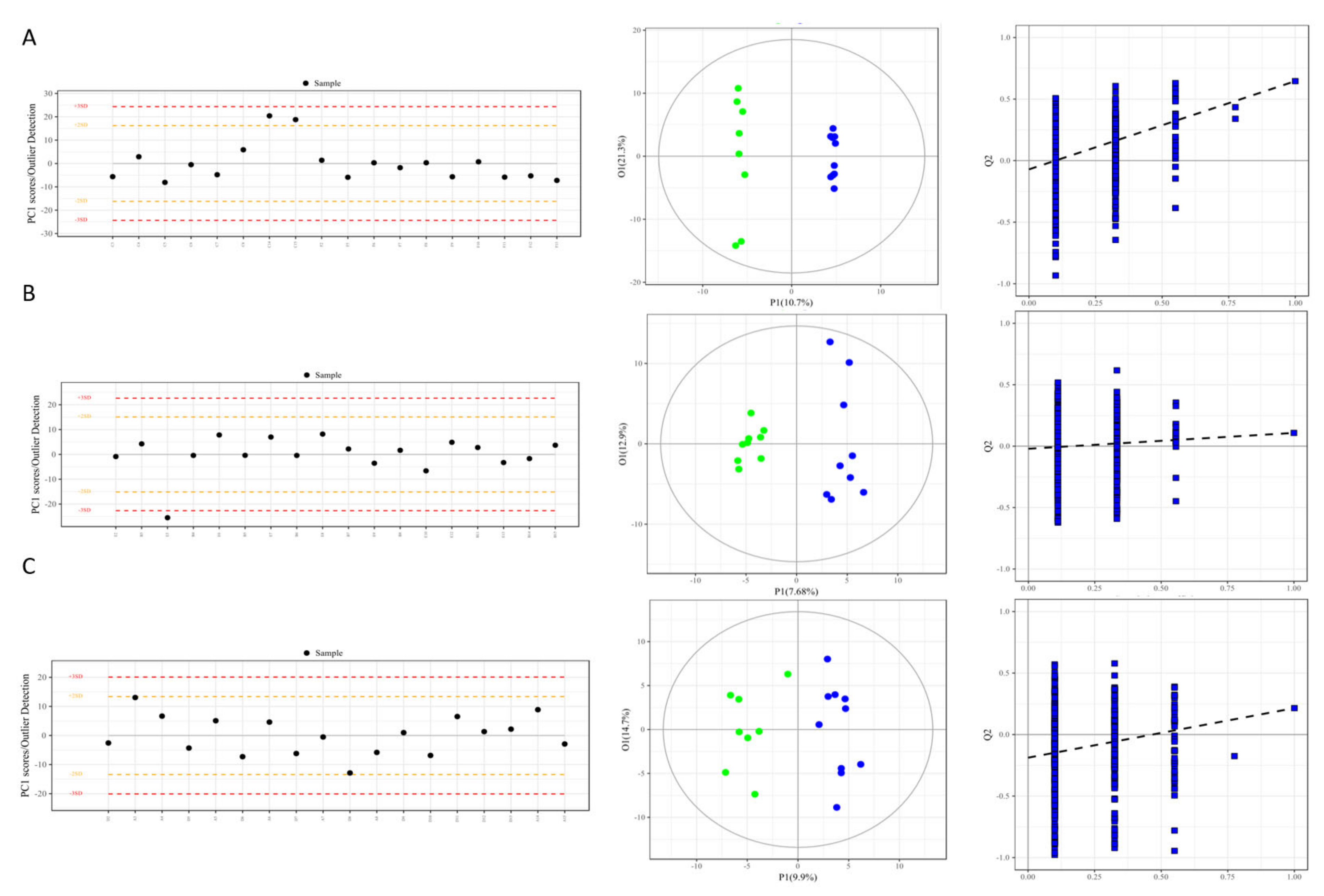
2.4. Quality Control Analysis and Model Reliability Testing
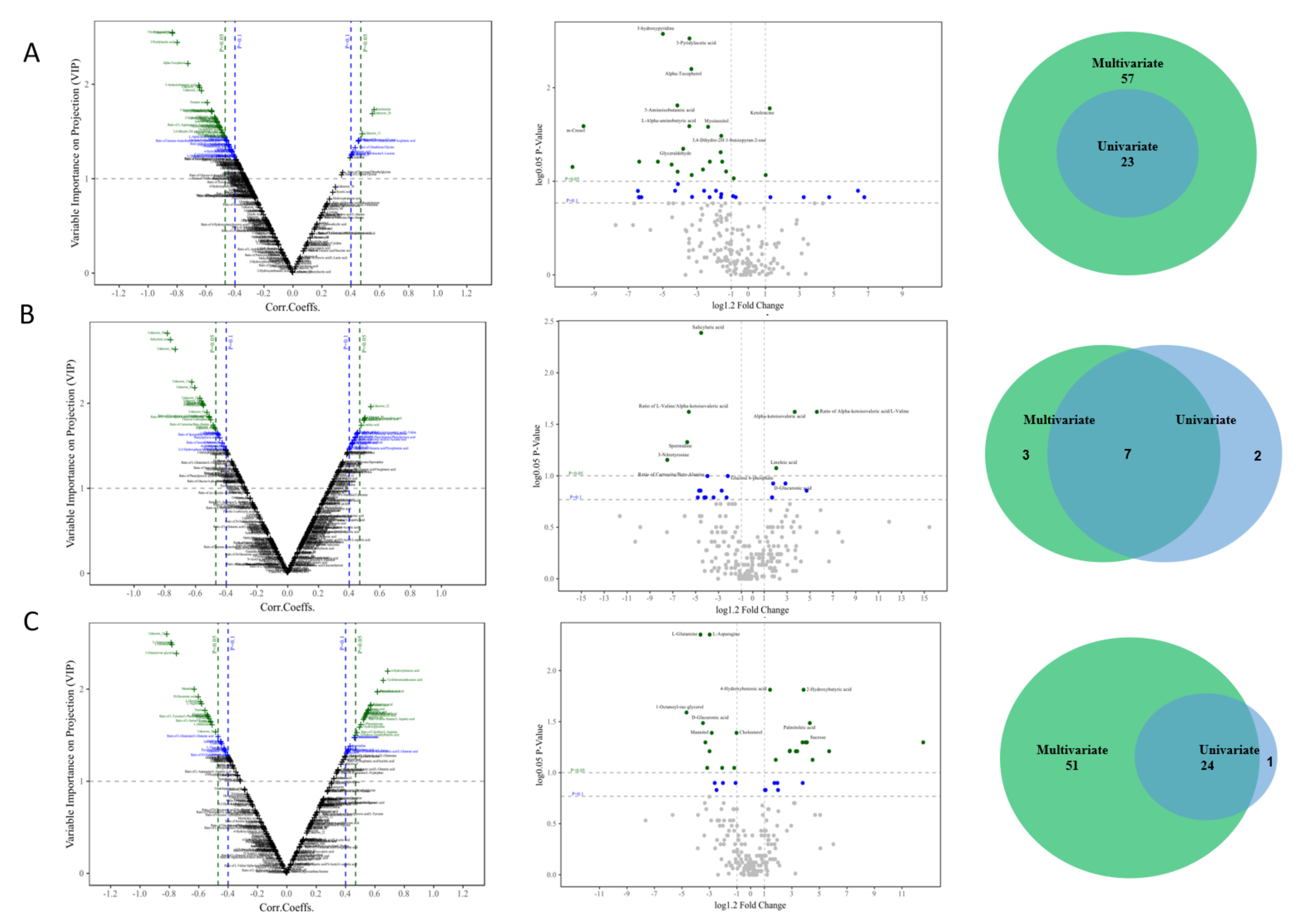
2.5. Data Analysis and Identification of Differentially Expressed Metabolites
3. Results
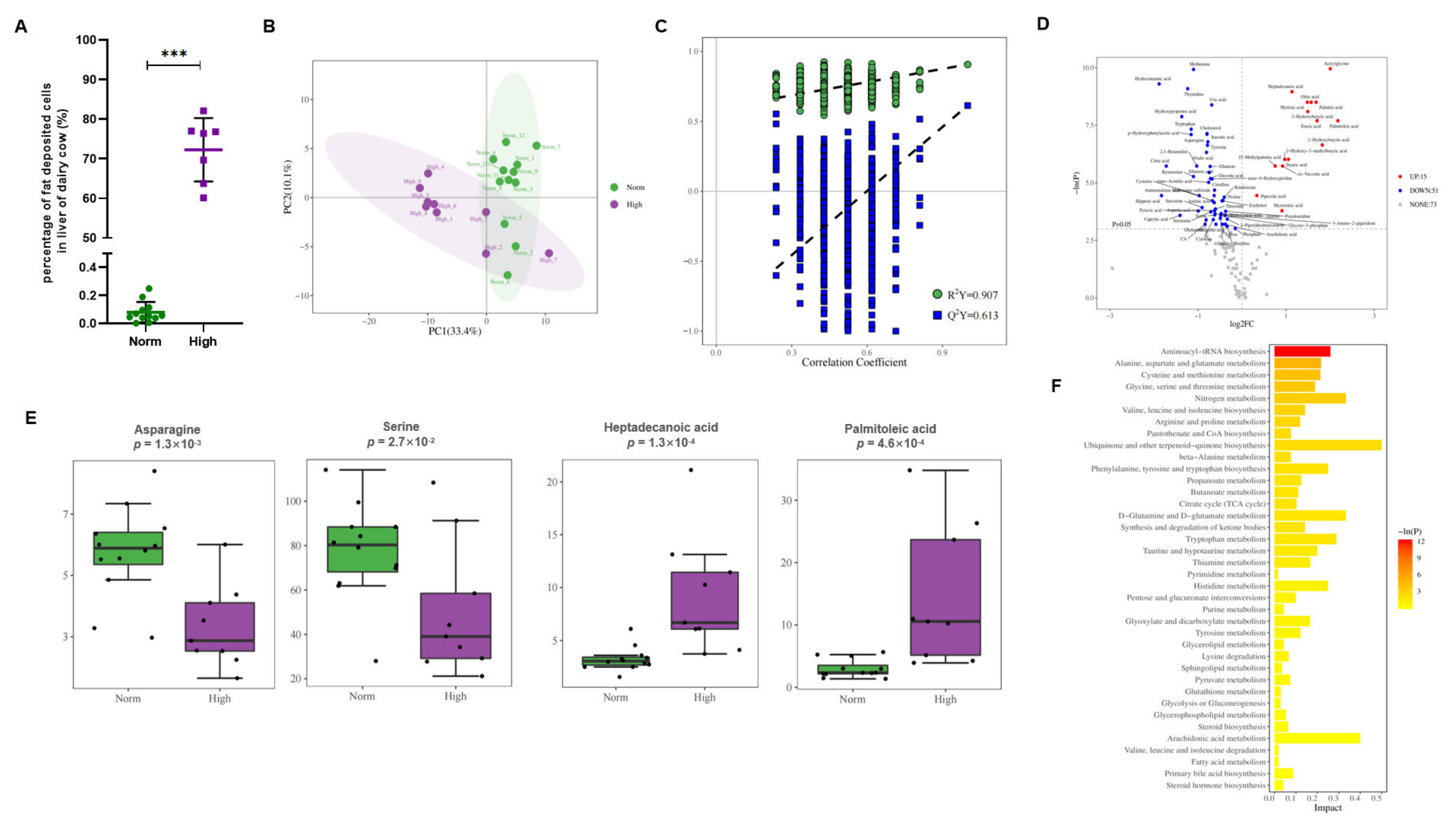
3.1. Workflow of the Study and the Participants Cohort
3.2. Metabolic Profiling, Model Establishment and Evaluation
3.3. Identification of Candidate Metabolic Biomarkers in Feces, Urine, and Serum in the Discovery Set
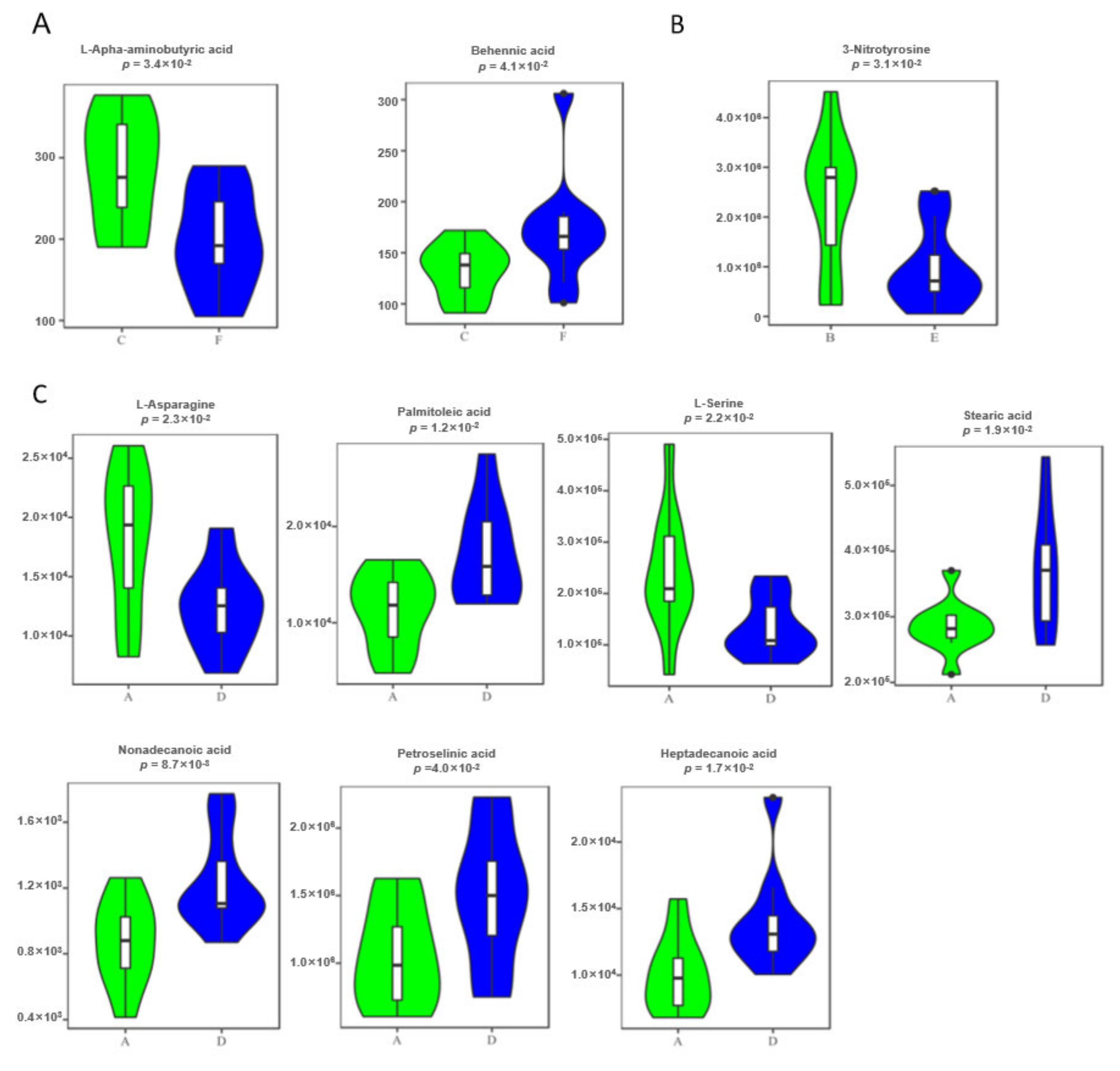
3.4. Defining Potential Metabolic Biomarkers for Fatty Liver Disease
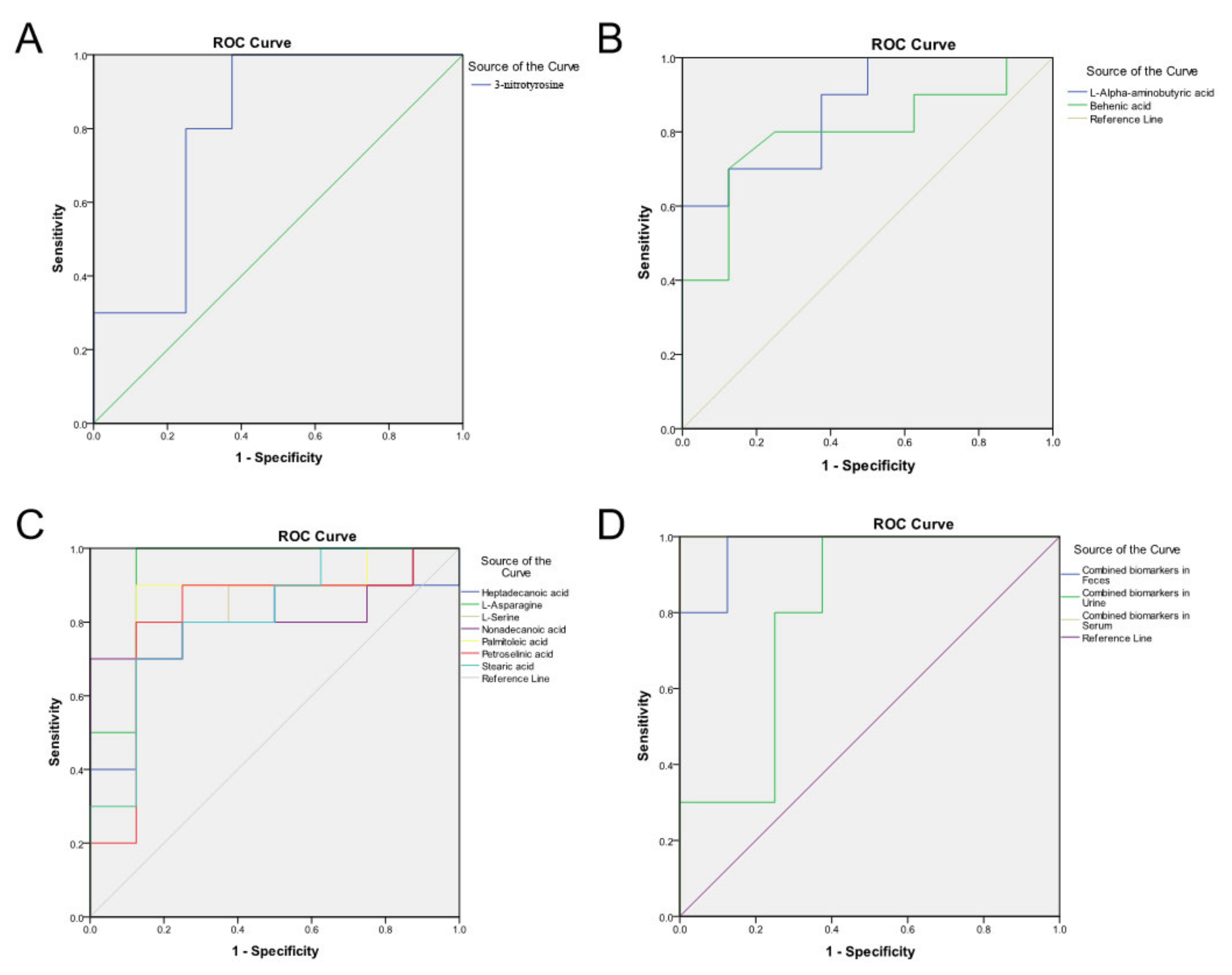
3.5. Validation of the Metabolic Marker Panel in the Test Set
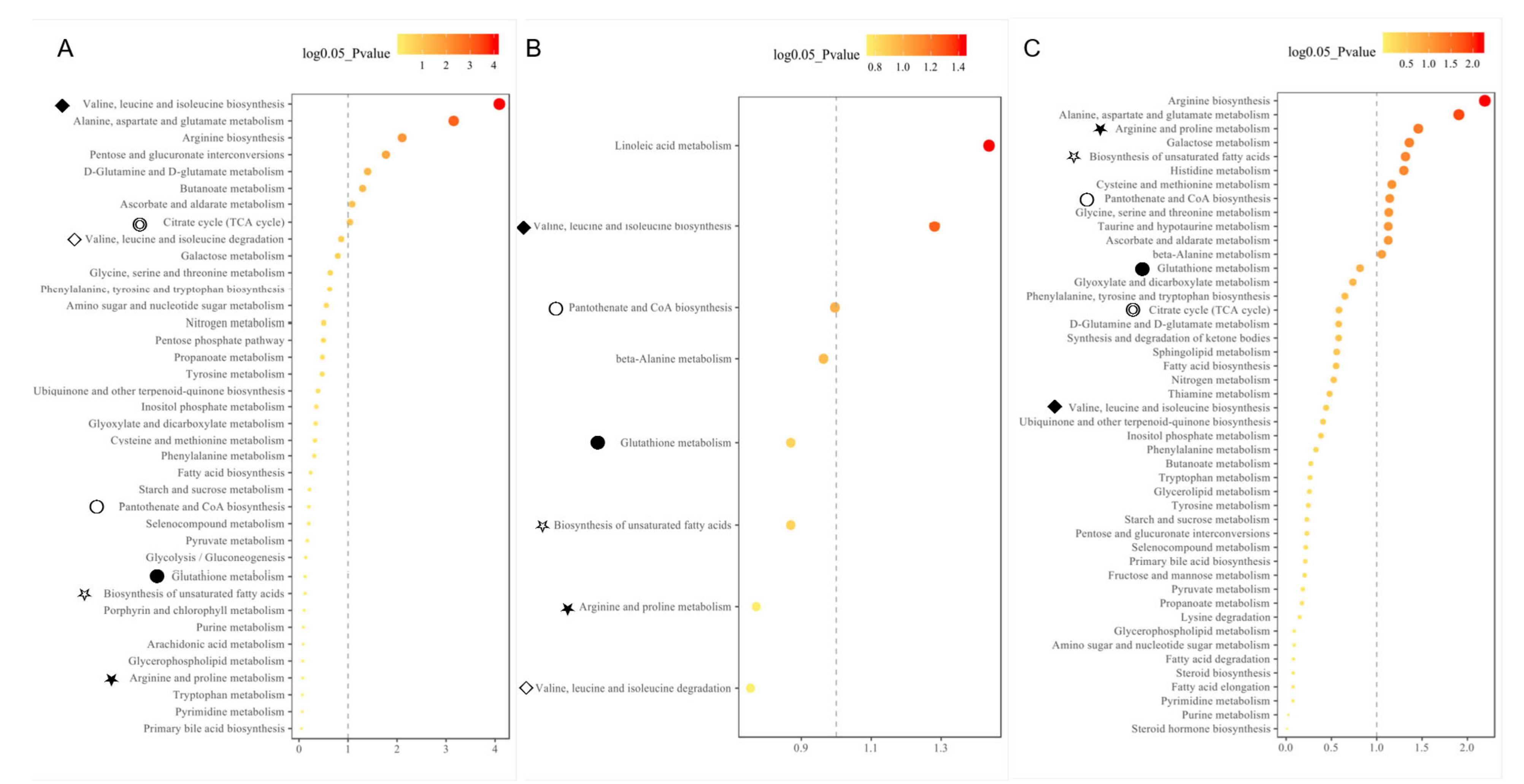
3.6. Associated Biological Pathways of Metabolite Biomarkers with Fatty Liver Syndrome in Cattle
3.7. Verification of Serum Biomarkers in a Third Liver Biopsy-Diagnosed Holstein Population
4. Discussion
4.1. Desirable and Novel Metabolite Biomarkers (Panels) to Early Diagnose Fatty Liver Cattle Were Strictly Identified in the Study
4.2. Dysregulated Fatty Acid Metabolism and Impaired Metabolism Capacity Were Accompanied with Fatty Liver Cattle
4.3. Common Biological Pathways Were Underlying the Pathogenesis of Fatty Liver Syndrome in Cattle
4.4. The Identified Serum Biomarkers Were Confirmed by a Third Biopsied Population
4.5. Potentiality of the Study
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Bobe, G.; Young, J.W.; Beitz, D.C. Invited review: Pathology, etiology, prevention, and treatment of fatty liver in dairy cows. J. Dairy Sci. 2004, 87, 3105–3124. [Google Scholar] [CrossRef]
- Shi, K.R.; Li, R.R.; Xu, Z.J.; Zhang, Q. Identification of crucial genetic factors, such as PPARγ, that regulate the pathogenesis of fatty liver disease in dairy cows is imperative for the sustainable development of dairy industry. Animals 2020, 10, 639. [Google Scholar] [CrossRef] [PubMed]
- Shi, K.R.; Niu, F.; Zhang, Q.; Ning, C.; Yue, S.J.; Hu, C.Z.; Xu, Z.J.; Wang, S.X.; Li, R.R.; Wang, Z.H. Identification of whole-genome significant single nucleotide polymorphisms in candidate genes associated with serum biochemical traits in chinese holstein cattle. Front. Genet. 2020, 11, 163. [Google Scholar] [PubMed]
- Katoh, N. Relevance of apolipoproteins in the development of fatty liver and fatty liver-related peripartum diseases in dairy cows. J. Vet. Med. Sci. 2002, 64, 293–307. [Google Scholar] [CrossRef]
- Bradford, B.J.; Yuan, K.; Farney, J.K.; Mamedova, L.K.; Carpenter, A.J. Invited review: Inflammation during the transition to lactation: New adventures with an old flame. J. Dairy Sci. 2015, 98, 6631–6650. [Google Scholar] [CrossRef]
- Carpenter, A.J.; Lioja, C.M.Y.; Vargas, C.F.; Mamedova, L.K.; Mendonça, L.G.; Coetzee, J.F.; Hollis, L.; Gehring, R.; Bradford, B. Hot topic: Early postpartum treatment of commercial dairy cows with nonsteroidal antiinflammatory drugs increases whole-lactation milk yield. J. Dairy Sci. 2016, 99, 672–679. [Google Scholar] [CrossRef]
- Zhang, L.T.; Liu, T.J.; Hu, C.Z.; Zhang, X.; Zhang, Q.; Shi, K.R. Proteome analysis identified proteins associated with mitochondrial function and inflammation activation crucially regulating the pathogenesis of fatty liver disease. BMC Genom. 2021, 22, 640. [Google Scholar]
- Adewuyi, A.A.; Gruys, E.; van Eerdenburg, F.J. Non esterified fatty acids (NEFA) in dairy cattle. A review. Vet. Q. 2005, 27, 117–126. [Google Scholar]
- Shen, Y.; Chen, L.; Yang, W.; Wang, X. Exploration of serum sensitive biomarkers of fatty liver in dairy cows. Sci. Rep. 2018, 8, 13574. [Google Scholar] [CrossRef]
- Gaal, T.; Reid, I.M.; Collins, R.A.; Roberts, C.J.; Pike, B.V. Comparison of biochemical and histological methods of estimating fat content of liver of dairy cows. Res. Vet. Sci. 1983, 34, 245–248. [Google Scholar]
- Reid, I.M.; Rowlands, G.J.; Dew, A.M.; Collins, R.A.; Roberts, C.J.; Manston, R. The relationship between post-parturient fatty liver and blood composition in dairy cows. J. Agric. Sci. 1983, 101, 473–480. [Google Scholar]
- Starke, A.; Haudum, A.; Weijers, G.; Herzog, K.; Wohlsein, P.; Beyerbach, M.; Thijssen, J.; Rehage, J.; De Korte, C. Noninvasive detection of hepatic lipidosis in dairy cows with calibrated ultrasonographic image analysis. J. Dairy Sci. 2010, 93, 2952–2965. [Google Scholar]
- Starke, A.; Schmidt, S.; Haudum, A.; Scholbach, T.; Wohlsein, P.; Beyerbach, M.; Rehage, J. Evaluation of portal blood flow using transcutaneous and intraoperative Doppler ultrasonography in dairy cows with fatty liver. J. Dairy Sci. 2011, 94, 2964–2971. [Google Scholar]
- Zhang, G.; Deng, Q.; Mandal, R.; Wishart, D.S.; Ametaj, B.N. DI/LC-MS/MS-Based Metabolic Profiling for Identification of Early Predictive Serum Biomarkers of Metritis in Transition Dairy Cows. J. Agric. Food Chem. 2017, 65, 8510–8521. [Google Scholar]
- Dervishi, E.; Zhang, G.; Zwierzchowski, G.; Mandal, R.; Wishart, D.S.; Ametaj, B.N. Serum metabolic fingerprinting of pre-lameness dairy cows by GC-MS reveals typical profiles that can identify susceptible cows. J. Proteomics. 2020, 213, 103620. [Google Scholar] [CrossRef]
- Eckel, E.F.; Zhang, G.; Dervishi, E.; Zwierzchowski, G.; Mandal, R.; Wishart, D.S.; Ametaj, B.N. Urinary metabolomics fingerprinting around parturition identifies metabolites that differentiate lame dairy cows from healthy ones. Animal 2020, 14, 2138–2149. [Google Scholar]
- Zhang, G.; Ametaj, B.N. Ketosis an old story under a new approach. Dairy 2020, 1, 42–60. [Google Scholar]
- Zhang, G.; Zwierzchowski, G.; Mandal, R.; Wishart, D.S.; Ametaj, B.N. Serum metabolomics identifies metabolite panels that differentiate lame dairy cows from healthy ones. Metabolomics 2020, 16, 73. [Google Scholar]
- Zhang, L.T.; Hu, C.Z.; Zhang, X.; Zhang, Q.; Yan, Z.G.; Wei, Q.; Wang, S.-X.; Xu, Z.-J.; Li, R.-R.; Liu, T.-J.; et al. Protein acetylation in mitochondria plays critical functions in the pathogenesis of fatty liver disease. BMC Genom. 2020, 21, 435. [Google Scholar]
- Zwierzchowski, G.; Zhang, G.; Mandal, R.; Wishart, D.S.; Ametaj, B.N. Milk metabotyping identifies metabolite alterations in the whole raw milk of dairy cows with lameness. J. Agric. Food Chem. 2020, 68, 4507–4514. [Google Scholar]
- Pan, L.; Qiu, Y.; Chen, T.; Lin, J.; Chi, Y.; Su, M.; Zhao, A.; Jia, W. An optimized procedure for metabonomic analysis of rat liver tissue using gas chromatography/time-of-flight mass spectrometry. J. Pharm. Biomed. Anal. 2010, 52, 589–596. [Google Scholar] [CrossRef]
- Wang, J.H.; Chen, W.L.; Li, J.M.; Wu, S.F.; Chen, T.L.; Zhu, Y.M.; Zhang, W.-N.; Li, Y.; Qiu, Y.-P.; Zhao, A.-H.; et al. Prognostic significance of 2-hydroxyglutarate levels in acute myeloid leukemia in China. Proc. Natl. Acad. Sci. USA 2013, 110, 17017–17022. [Google Scholar] [CrossRef] [Green Version]
- Luo, P.; Yin, P.Y.; Hua, R.; Tan, Y.X.; Li, Z.F.; Qiu, G.K.; Yin, Z.; Xie, X.; Wang, X.; Chen, W.; et al. A large-scale, multi-center serum metabolite biomarker identification study for the early detection of hepatocellular carcinoma. Hepatology 2018, 67, 662–674. [Google Scholar] [CrossRef]
- Sun, H.Z.; Wang, B.; Wang, J.K.; Liu, H.Y.; Liu, J.X. Biomarker and pathway analyses of urine metabolomics in dairy cows when corn stover replaces alfalfa hay. J. Anim. Sci. Biotechnol. 2016, 7, 49. [Google Scholar] [CrossRef]
- Tian, H.; Zheng, N.; Wang, W.; Cheng, J.; Li, S.; Zhang, Y.; Wang, J. Integrated metabolomics study of the milk of heat-stressed lactating dairy cows. Sci. Rep. 2016, 6, 24208. [Google Scholar] [CrossRef]
- Hanley, J.A.; McNeil, B.J. The meaning and use of the area under a receiver operating characteristic (ROC) curve. Radiology 1982, 143, 29–36. [Google Scholar] [CrossRef]
- Pencina, M.J.; D’Agostino, R.B., Sr.; D’Agostino, R.B., Jr.; Vasan, R.S. Evaluating the added predictive ability of a new marker: From area under the ROC curve to reclassification and beyond. Stat. Med. 2008, 27, 157–172. [Google Scholar] [CrossRef] [PubMed]
- Xia, J.; Wishart, D.S. MetPA: A web-based metabolomics tool for pathway analysis and visualization. Bioinformatics 2010, 26, 2342–2344. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Tobolski, D.; Zwierzchowski, G.; Mandal, R.; Wishart, D.S.; Ametaj, B.N. A targeted serum metabolomics GC-MS approach identifies predictive blood biomarkers for retained placenta in Holstein dairy cows. Metabolites 2021, 11, 633. [Google Scholar] [CrossRef] [PubMed]
- Yuan, M.; Breitkopf, S.B.; Yang, X.M.; Asara, J.M. A positive/negative ion-switching, targeted mass spectrometry based metabolomics platform for bodily fluids, cells, and fresh and fixed tissue. Nat. Protoc. 2012, 7, 872–881. [Google Scholar] [CrossRef] [PubMed]
- Goff, J.P.; Horst, R.L. Physiological changes at parturition and their relationship to metabolic disorders. J. Dairy Sci. 1997, 80, 1260–1268. [Google Scholar] [CrossRef]
- Herr, M.; Bostedt, H.; Failing, K. IgG and IgM levels in dairy cows during the periparturient period. Theriogenology 2011, 75, 377–385. [Google Scholar] [CrossRef]
- Herdt, T.H. Fatty liver in dairy cows. Vet. Clin. North Am. Food Anim. Pract. 1988, 4, 269–287. [Google Scholar] [CrossRef]
- Farid, A.S.; Honkawa, F.; Fath, E.M. Serum paraoxonase-1 as biomarker for improved diagnosis of fatty liver in dairy cows. BMC Vet. Res. 2013, 9, 73. [Google Scholar] [CrossRef]
- Imhasly, S.; Naegeli, H.; Baumann, S.; Bergen, M.V.; Luch, A.; Jungnickel, H.; Potratz, S.; Gerspach, C. Metabolomic biomarkers correlating with hepatic lipidosis in dairy cows. BMC Vet. Res. 2014, 10, 122. [Google Scholar] [CrossRef]
- Pullen, D.L.; Liesman, J.S.; Emery, R.S. A species comparison of liver slice synthesis and secretion of triacylglycerol from nonesterified fatty acids in media. J. Anim. Sci. 1990, 68, 1395–1399. [Google Scholar] [CrossRef]
- Williamson, J.R.; Browning, E.T.; Scholz, R.; Kreisberg, R.A.; Fritz, I.B. Inhibition of fatty acid stimulation of gluconeogenesis by (+)-decanoylcarnitine in perfused rat liver. Diabetes 1968, 17, 194–208. [Google Scholar] [CrossRef]
- Avila, M.A.; Berasain, C.; Torres, L.; Martín-Duce, A.; Corrales, F.J.; Yang, H.; Prieto, J.; Lu, S.C.; Caballería, J.; Rodés, J.; et al. Reduced mRNA abundance of the main enzymes involved in methionine metabolism in human liver cirrhosis and hepatocellular carcinoma. J. Hepatol. 2000, 33, 907–914. [Google Scholar]
- Mato, J.M.; Martinez-Chantar, M.L.; Lu, S.C. Methionine metabolism and liver disease. Annu. Rev. Nutr. 2008, 28, 273–293. [Google Scholar]
- Mantena, S.K.; Vaughn, D.P.; Andringa, K.K.; Eccleston, H.B.; King, A.L.; Abrams, G.A.; Doeller, J.E.; Kraus, D.W.; Darley-Usmar, V.M.; Bailey, S.M. High fat diet induces dysregulation of hepatic oxygen gradients and mitochondrial function in vivo. Biochem. J. 2009, 417, 183–193. [Google Scholar]
- Zhang, G.; Hailemariam, D.; Dervishi, E.; Goldansaz, S.A.; Deng, Q.; Dunn, S.M.; Ametaj, B.N. Dairy cows affected by ketosis show alterations in innate immunity and lipid and carbohydrate metabolism during the dry off period and postpartum. Res. Vet. Sci. 2016, 107, 246–256. [Google Scholar] [CrossRef]
- Myher, J.J.; Marai, L.; Kuksis, A.; Yousef, I.M.; Fisher, M.M. Identification of ornithine and arginine conjugates of cholic acid by mass spectrometry. Can. J. Biochem. 1975, 53, 583–590. [Google Scholar] [CrossRef]
- Voloshin, I.; Hahn-Obercyger, M.; Anavi, S.; Tirosh, Q. L-arginine conjugates of bile acids-a possible treatment for non-alcoholic fatty liver disease. Lipids Health Dis. 2014, 13, 69. [Google Scholar] [CrossRef]
- Clifford, B.L.; Sedgeman, L.R.; Williams, K.J.; Morand, P.; Cheng, A.; Jarrett, K.E.; Chan, A.P.; Brearley-Sholto, M.C.; Wahlström, A.; Ashby, J.W.; et al. FXR activation protects against NAFLD via bile-acid-dependent reductions in lipid absorption. Cell Metab. 2021, 33, 1671–1684.e4. [Google Scholar] [CrossRef]
- Adinolfi, L.E.; Ingrosso, D.; Cesaro, G.; Cimmino, A.; D’Antò, M.; Capasso, R.; Zappia, V.; Ruggiero, G. Hyperhomocysteinemia and the MTHFR C677T polymorphism promote steatosis and fibrosis in chronic hepatitis C patients. Hepatology 2005, 41, 995–1003. [Google Scholar] [CrossRef]
- Varela-Rey, M.; Martínez-López, N.; Fernández-Ramos, D.; Embade, N.; Calvisi, D.F.; Woodhoo, A.; Rodríguez, J.; Fraga, M.F.; Julve, J.; Rodríguez-Millán, E.; et al. Fatty liver and fibrosis in glycine N-methyltransferase knockout mice is prevented by nicotinamide. Hepatology 2010, 52, 105–114. [Google Scholar] [CrossRef]
- Martinez-Chantar, M.L.; Vázquez-Chantada, M.; Ariz, U.; Martínez, N.; Varela, M.; Luka, Z.; Capdevila, A.; Rodríguez, J.; Aransay, A.M.; Matthiesen, R.; et al. Loss of the glycine N-methyltransferase gene leads to steatosis and hepatocellular carcinoma in mice. Hepatology 2008, 47, 1191–1199. [Google Scholar] [CrossRef]
- Chen, Y.M.; Shiu, J.Y.; Tzeng, S.J.; Shih, L.S.; Chen, Y.J.; Lui, W.Y.; Chen, P.-H. Characterization of glycine-N-methyltransferase-gene expression in human hepatocellular carcinoma. Int. J. Cancer 1998, 75, 787–793. [Google Scholar] [CrossRef]
- Mudd, S.H.; Cerone, R.; Schiaffino, M.C.; Fantasia, A.R.; Minniti, G.; Caruso, U.; Lorini, R.; Watkins, D.; Matiaszuk, N.; Rosenblatt, D.S.; et al. Glycine N-methyltransferase deficiency: A novel inborn error causing persistent isolated hypermethioninaemia. J. Inherit. Metab. Dis. 2001, 24, 448–464. [Google Scholar] [CrossRef]
- Augoustides-Savvopoulou, P.; Luka, Z.; Karyda, S.; Stabler, S.P.; Allen, R.H.; Patsiaoura, K.; Wagner, C.; Mudd, S.H. Glycine N-methyltransferase deficiency: A new patient with a novel mutation. J. Inherit. Metab. Dis. 2003, 26, 745–759. [Google Scholar] [CrossRef]
- Nasr, P.; Ignatova, S.; Kechagias, S.; Ekstedt, M. Natural history of nonalcoholic fatty liver disease: A prospective follow-up study with serial biopsies. Hepatol. Commun. 2018, 2, 199–210. [Google Scholar] [CrossRef] [PubMed]
- Demir, M.; Lang, S.; Steffen, H.M. Nonalcoholic fatty liver disease—Current status and future directions. J. Dig. Dis. 2015, 16, 541–557. [Google Scholar] [CrossRef] [PubMed]
- Younossi, Z.M.; Loomba, R.; Rinella, M.E.; Bugianesi, E.; Marchesini, G.; Neuschwander-Tetri, B.A.; Serfaty, L.; Negro, F.; Caldwell, S.H.; Ratziu, V.; et al. Current and future therapeutic regimens for nonalcoholic fatty liver disease and nonalcoholic steatohepatitis. Hepatology 2018, 68, 361–371. [Google Scholar] [CrossRef] [PubMed]
- Zhou, F.; Zhou, J.H.; Wang, W.X.; Zhang, X.J.; Ji, Y.X.; Zhang, P.; She, Z.G.; Zhu, L.; Cai, J.; Li, H. Unexpected rapid increase in the burden of NAFLD in China from 2008 to 2018: A systematic review and meta-analysis. Hepatology 2019, 70, 1119–1133. [Google Scholar]
- Caussy, C.; Hsu, C.; Lo, M.T.; Liu, A.; Bettencourt, R.; Ajmera, V.H.; Bassirian, S.; Hooker, J.; Sy, E.; Richards, L.; et al. Link between gut-microbiome derived metabolite and shared gene-effects with hepatic steatosis and fibrosis in NAFLD. Hepatology 2018, 68, 918–932. [Google Scholar] [CrossRef]
- Zhang, A.; Sun, H.; Han, Y.; Yuan, Y.; Wang, P.; Song, G.; Yuan, X.; Zhang, M.; Xie, N.; Wang, X. Exploratory urinary metabolic biomarkers and pathways using UPLC-Q-TOF-HDMS coupled with pattern recognition approach. Analyst 2012, 137, 4200–4208. [Google Scholar] [CrossRef]
- Ajmera, V.; Perito, E.R.; Bass, N.M.; Terrault, N.A.; Yates, K.P.; Gill, R.; Loomba, R.; Diehl, A.M.; Aouizerat, B.E. Novel plasma biomarkers associated with liver disease severity in adults with nonalcoholic fatty liver disease. Hepatology 2017, 65, 65–77. [Google Scholar] [CrossRef] [Green Version]
- Huang, Q.; Tan, Y.; Yin, P.; Ye, G.; Gao, P.; Lu, X.; Wang, H.; Xu, G. Metabolic characterization of hepatocellular carcinoma using nontargeted tissue metabolomics. Cancer Res. 2013, 73, 4992–5002. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Biomarker Source | Biomarker Name | Class | FC (FL/Norm) 3 | p-Value | HMDB ID 4 | KEGG ID 5 | Associated Pathways |
|---|---|---|---|---|---|---|---|
| Feces | L-Alpha-aminobutyric acid | Amino Acid | 0.534 | 8.50 × 10−3 | HMDB00452 | C02356 | / |
| Behenic acid | Fatty Acids | 1.203 | 4.10 × 10−2 | HMDB00944 | / | ||
| Urine | 3-Nitrotyrosine | Amino Acid | 0.256 | 3.10 × 10−2 | HMDB01904 | / | / |
| Serum | L-Asparagine | Amino Acid | 0.58 | 8.70 × 10−4 | HMDB00168 | C00152 | Ammonia Recycling; Aspartate Metabolism; Transcription/Translation |
| Palmitoleic acid | Fatty Acids | 2.191 | 1.20 × 10−2 | HMDB03229 | C08362 | / | |
| L-Serine | Amino Acid | 0.579 | 2.70 × 10−2 | HMDB00187 | C00065 | Ammonia Recycling; Glycine and Serine Metabolism; Homocysteine Degradation; Methionine Metabolism; Sphingolipid Metabolism | |
| Stearic acid | Fatty Acids | 1.819 | 2.70 × 10−2 | HMDB00827 | C01530 | Mitochondrial Beta-Oxidation of Long Chain Saturated Fatty Acids; Plasmalogen Synthesis | |
| Nonadecanoic acid | Fatty Acids | 1.678 | 2.70 × 10−2 | HMDB00772 | C16535 | / | |
| Petroselinic acid | Organic Acids | 2.831 | 2.70 × 10−2 | HMDB02080 | C08363 | / | |
| Heptadecanoic acid | Fatty Acids | 2.272 | 3.40 × 10−2 | HMDB02259 | / | / |
| Biomarker Source | Biomarker Name | AUC in Discovery Set (95% CI) 2 | AUC in Test Set (95% CI) 3 | p-Value in Violin Chart 4 |
|---|---|---|---|---|
| Traditional Biochemical Indicator in Serum 1 | AST | 0.756 | / | / |
| UREA | 0.5 | / | / | |
| ALB | 0.469 | / | / | |
| INS | 0.363 | / | / | |
| UA | 0.338 | / | / | |
| TP | 0.313 | / | / | |
| TG | 0.294 | / | / | |
| TCHO | 0.25 | / | / | |
| GLU | 0.112 | / | / | |
| Biomarker in Feces | L-Alpha-aminobutyric acid | 0.863 | 0.825 | 3.40 × 10−2 |
| Behenic acid | 0.794 | 0.929 | 4.10 × 10−2 | |
| Combined biomarkers in Feces | / | 0.975 | 1 | / |
| Biomarker in Urine | 3-Nitrotyrosine | 0.802 | 0.841 | 3.10 × 10−2 |
| Combined biomarkers in Feces and Urine | / | 0.988 | 1 | / |
| Biomarker in Serum | L-Asparagine | 0.938 | 0.76 | 2.30 × 10−2 |
| Palmitoleic acid | 0.85 | 0.81 | 1.20 × 10−2 | |
| L-Serine | 0.812 | 0.79 | 2.20 × 10−2 | |
| Stearic acid | 0.813 | 0.79 | 1.90 × 10−2 | |
| Nonadecanoic acid | 0.813 | 0.84 | 8.70 × 10−3 | |
| Petroselinic acid | 0.813 | 0.76 | 4.00 × 10−2 | |
| Heptadecanoic acid | 0.8 | 0.84 | 1.70 × 10−2 | |
| Combined Biomarkers in Serum | / | 1 | 1 | / |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, X.; Liu, T.; Hou, X.; Hu, C.; Zhang, L.; Wang, S.; Zhang, Q.; Shi, K. Multi-Channel Metabolomics Analysis Identifies Novel Metabolite Biomarkers for the Early Detection of Fatty Liver Disease in Dairy Cows. Cells 2022, 11, 2883. https://doi.org/10.3390/cells11182883
Zhang X, Liu T, Hou X, Hu C, Zhang L, Wang S, Zhang Q, Shi K. Multi-Channel Metabolomics Analysis Identifies Novel Metabolite Biomarkers for the Early Detection of Fatty Liver Disease in Dairy Cows. Cells. 2022; 11(18):2883. https://doi.org/10.3390/cells11182883
Chicago/Turabian StyleZhang, Xuan, Tingjun Liu, Xianpeng Hou, Chengzhang Hu, Letian Zhang, Shengxuan Wang, Qin Zhang, and Kerong Shi. 2022. "Multi-Channel Metabolomics Analysis Identifies Novel Metabolite Biomarkers for the Early Detection of Fatty Liver Disease in Dairy Cows" Cells 11, no. 18: 2883. https://doi.org/10.3390/cells11182883