
The Role of Crosstalk between Adipose Cells and Myocytes in the Pathogenesis of Sarcopenic Obesity in the Elderly


Abstract
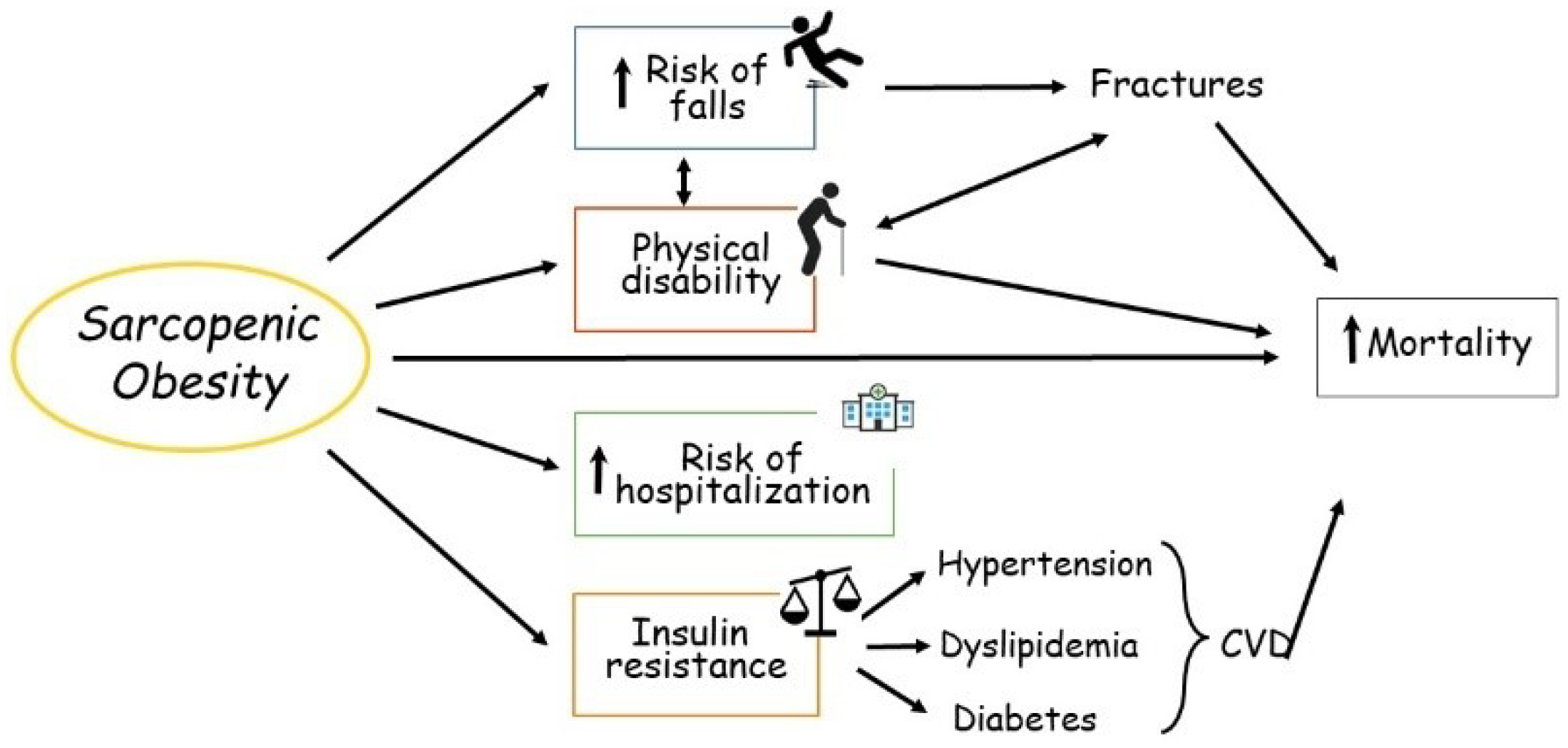
:1. Introduction
2. Adipose Tissue as Endocrine Organ
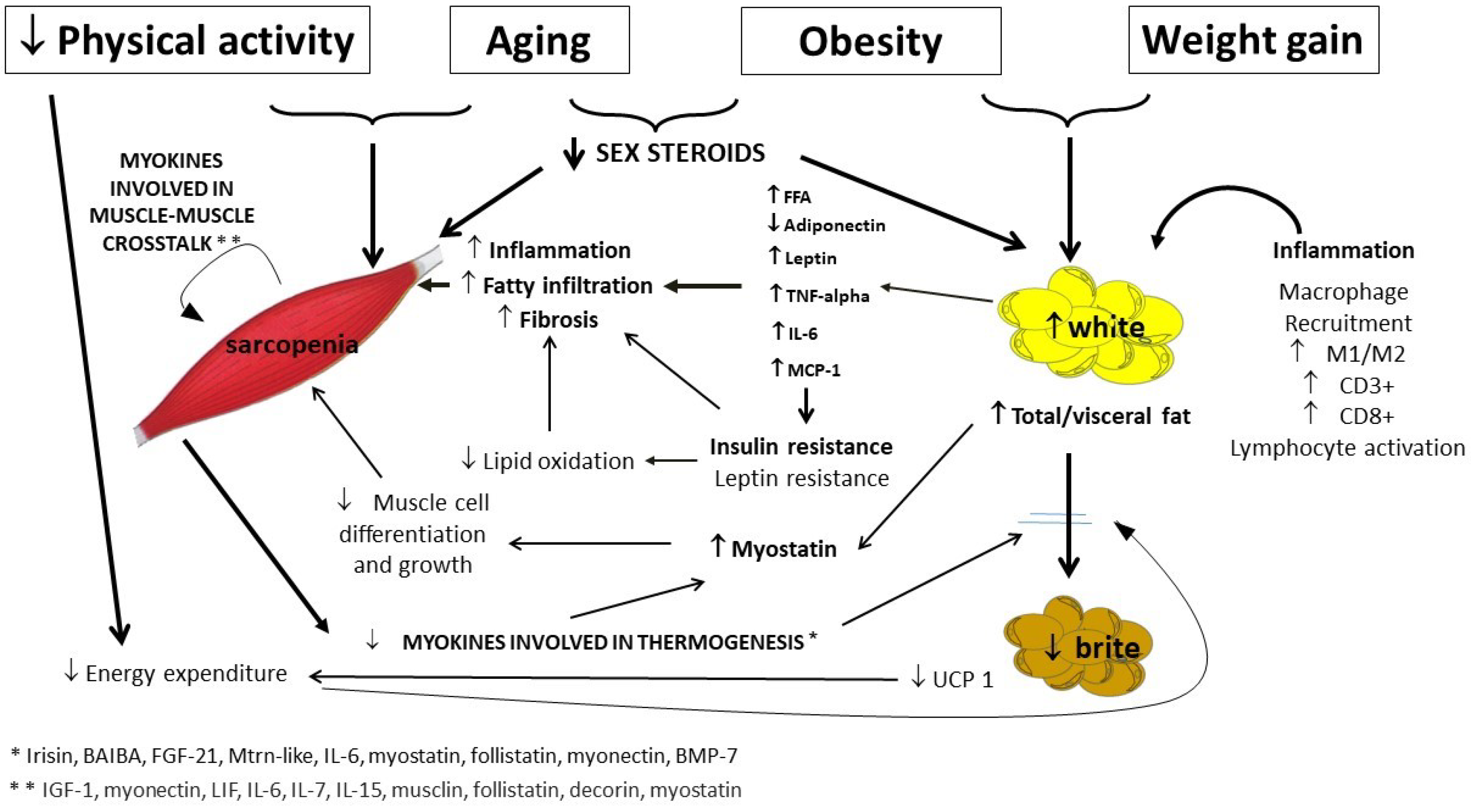
3. Adipose Tissue Changes across Aging
4. Muscle as Endocrine Organ

{kind=link}
{kind=link}
| Function | Myokine | Aging | References |
|---|---|---|---|
| Myogenesis and muscle hypertrophy | myostatin LIF IL-6 IL-7 IL-15 musclin follistatin decorin myonectin IGF-1 musclin | ⇑ ? ⇑ ⇓ ⇓ ? ⇓ ⇓ ⇓ ⇓ ? | [61,74] [67] [66,75] [58] [76] [77] [64,78] [79,80] [81] [56] [82] |
| Muscle-cell FFA oxidation | IL-6 BDNF irisin myonectin | ⇑ ⇓ ⇓ ⇓ | [69,83] [68] [84] [81] |
| Insulin sensitivity | IL-6 IL-15 SPARC LIF BMP-7 mitsugumin 53 | ⇑ ⇓ ⇓ ? ⇓ ? | [7,69] [85,86] [87] [88] [89] [90] |
| Osteogenesis | IGF-1 decorin IL-6 | ⇓ ⇓ ⇑ | [91] [92] [93] |
| Browning of WAT | Irisin IL-6 meteorin-like FGF-21 BAIBA follistatin myonectin BMP-7 | ⇓ ⇑ ? ⇓ ⇓ ⇓ ? ⇓ | [84] [94] [95] [96] [97,98] [99] [100] [101] |
| Lipolysis | IL-6 FGF-21 ANGPTL-4 | ⇑ ⇓ ? | [7] [102] [103] |
| Muscle innervation | BDNF FGFBP-1 | ⇓ ? | [104] [105] |
| Muscle angiogenesis | IL-8 VEGF-A | ⇓ ? | [76] [106,107] |
5. Muscle Mass Changes across Aging
6. Muscle–Adipose Tissue Crosstalk: Role of Myokines
7. Adipose Tissue Muscle Crosstalk: Role of Adipokines
8. Adipomyokines and MicroRNA in the Crosstalk between AT and Muscle
| Adipomyokine | Effects–Skeletal Muscle | Effects—Adipose Tissue | Aging | References |
|---|---|---|---|---|
| IL-6 | +muscle hypertrophy +glucose uptake +glycogenolysis, lipolysis | +lipolysis +free fatty acid (FFA) oxidation browning of WAT | ⇑ | [7,69] |
| Irisin | +glucose uptake +muscle trophism | +lipolysis browning of WAT | ⇓ | [176,177] |
| IL-15 | +glucose uptake +mitochondrial activity | −lipid accumulation +adiponectin secretion | ⇓ | [169,171,178] |
| BAIBA | +mitochondrial FFA oxidation +insulin sensitivity | +mitochondrial FFA oxidation | ⇓ | [97,139] |
| Meteorin-like | +energy expenditure +glucose tolerance | browning of WAT | ? | [95] |
| LIF | +muscle hypertrophy +satellite cell proliferation regeneration after muscle damage | +adipocyte differentiation | ⇓ | [67] |
| Myostatin | - muscle hypertrophy | +adipogenesis | ⇑ | [60,161,179,180] |
| Apelin | improves muscle metabolism | glucose uptake −lipid storage | ⇓ | [129,130,131,132] |
| ANGPTL4 | +FFA oxidation | +lipolysis | ? | [103,181] |
| FGF-21 | +thermogenesis | +glucose uptake | ⇓ | [182,183] |
| Follistatin-like 1 | +endothelial cells function and survival | ? | [184,185] | |
| IL-8 | +insulin resistance | +insulin resistance | ⇓ | [186,187] |
| MCP-1 | −glucose uptake | ? | [187] | |
| PEDF | +insulin resistance +ectopic lipid deposition | +insulin resistance +pro-inflammatory pathway | ? | [188,189] |
9. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Donini, L.M.; Busetto, L.; Bischoff, S.C.; Cederholm, T.; Ballesteros-Pomar, M.D.; Batsis, J.A.; Bauer, J.M.; Boirie, Y.; Cruz-Jentoft, A.J.; Dicker, D.; et al. Definition and Diagnostic Criteria for Sarcopenic Obesity: ESPEN and EASO Consensus Statement. Obes. Facts 2022, 15, 321–335. [Google Scholar] [CrossRef] [PubMed]
- Zamboni, M.; Mazzali, G.; Fantin, F.; Rossi, A.; di Francesco, V. Sarcopenic Obesity: A New Category of Obesity in the Elderly. Nutr. Metab. Cardiovasc. Dis. 2008, 18, 388–395. [Google Scholar] [CrossRef] [PubMed]
- Batsis, J.A.; Villareal, D.T. Sarcopenic Obesity in Older Adults: Aetiology, Epidemiology and Treatment Strategies. Nat. Rev. Endocrinol. 2018, 14, 513–537. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.J.; Tamadon, A.; Park, H.T.; Kim, H.; Ku, S.-Y. The Role of Sex Steroid Hormones in the Pathophysiology and Treatment of Sarcopenia. Osteoporos. Sarcopenia 2016, 2, 140–155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wawrzkiewicz-Jałowiecka, A.; Lalik, A.; Soveral, G. Recent Update on the Molecular Mechanisms of Gonadal Steroids Action in Adipose Tissue. Int. J. Mol. Sci. 2021, 22, 5226. [Google Scholar] [CrossRef]
- Funcke, J.-B.; Scherer, P.E. Beyond Adiponectin and Leptin: Adipose Tissue-Derived Mediators of Inter-Organ Communication. J. Lipid. Res. 2019, 60, 1648–1684. [Google Scholar] [CrossRef]
- Pedersen, B.K.; Febbraio, M.A. Muscle as an Endocrine Organ: Focus on Muscle-Derived Interleukin-6. Physiol. Rev. 2008, 88, 1379–1406. [Google Scholar] [CrossRef] [Green Version]
- Kershaw, E.E.; Flier, J.S. Adipose Tissue as an Endocrine Organ. J. Clin. Endocrinol. Metab. 2004, 89, 2548–2556. [Google Scholar] [CrossRef]
- Bosello, O.; Zamboni, M. Visceral Obesity and Metabolic Syndrome. Obes. Rev. 2000, 1, 47–56. [Google Scholar] [CrossRef]
- Kwon, H.; Kim, D.; Kim, J.S. Body Fat Distribution and the Risk of Incident Metabolic Syndrome: A Longitudinal Cohort Study. Sci. Rep. 2017, 7, 10955. [Google Scholar] [CrossRef]
- Hube, F.; Lietz, U.; Igel, M.; Jensen, P.; Tornqvist, H.; Joost, H.-G.; Hauner, H. Difference in Leptin MRNA Levels Between Omental and Subcutaneous Abdominal Adipose Tissue From Obese Humans. Horm. Metab. Res. 1996, 28, 690–693. [Google Scholar] [CrossRef]
- Lefebvre, A.-M.; Laville, M.; Vega, N.; Riou, J.P.; van Gaal, L.; Auwerx, J.; Vidal, H. Depot-Specific Differences in Adipose Tissue Gene Expression in Lean and Obese Subjects. Diabetes 1998, 47, 98–103. [Google Scholar] [CrossRef] [PubMed]
- Trayhurn, P.; Bing, C. Appetite and Energy Balance Signals from Adipocytes. Philos. Trans. R. Soc. B Biol. Sci. 2006, 361, 1237–1249. [Google Scholar] [CrossRef]
- Zhao, S.; Kusminski, C.M.; Scherer, P.E. Adiponectin, Leptin and Cardiovascular Disorders. Circ. Res. 2021, 128, 136–149. [Google Scholar] [CrossRef] [PubMed]
- Goldstein, B.J.; Scalia, R. Adiponectin: A Novel Adipokine Linking Adipocytes and Vascular Function. J. Clin. Endocrinol. Metab. 2004, 89, 2563–2568. [Google Scholar] [CrossRef] [Green Version]
- Guenther, M.; James, R.; Marks, J.; Zhao, S.; Szabo, A.; Kidambi, S. Adiposity Distribution Influences Circulating Adiponectin Levels. Transl. Res. 2014, 164, 270–277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryo, M.; Nakamura, T.; Kihara, S.; Kumada, M.; Shibazaki, S.; Takahashi, M.; Nagai, M.; Matsuzawa, Y.; Funahashi, T. Adiponectin as a Biomarker of the Metabolic Syndrome. Circ. J. 2004, 68, 975–981. [Google Scholar] [CrossRef] [Green Version]
- Weisberg, S.P.; McCann, D.; Desai, M.; Rosenbaum, M.; Leibel, R.L.; Ferrante, A.W. Obesity Is Associated with Macrophage Accumulation in Adipose Tissue. J. Clin. Investig. 2003, 112, 1796–1808. [Google Scholar] [CrossRef]
- Colleluori, G.; Villareal, D.T. Aging, Obesity, Sarcopenia and the Effect of Diet and Exercise Intervention. Exp. Gerontol. 2021, 155, 111561. [Google Scholar] [CrossRef]
- Hotamisligil, G.S. Inflammation, Metaflammation and Immunometabolic Disorders. Nature 2017, 542, 177–185. [Google Scholar] [CrossRef]
- Cinti, S.; Mitchell, G.; Barbatelli, G.; Murano, I.; Ceresi, E.; Faloia, E.; Wang, S.; Fortier, M.; Greenberg, A.S.; Obin, M.S. Adipocyte Death Defines Macrophage Localization and Function in Adipose Tissue of Obese Mice and Humans. J. Lipid. Res. 2005, 46, 2347–2355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zamboni, M.; Mazzali, G.; Zoico, E.; Harris, T.B.; Meigs, J.B.; di Francesco, V.; Fantin, F.; Bissoli, L.; Bosello, O. Health Consequences of Obesity in the Elderly: A Review of Four Unresolved Questions. Int. J. Obes. 2005, 29, 1011–1029. [Google Scholar] [CrossRef] [Green Version]
- Cruz-Jentoft, A.J.; Bahat, G.; Bauer, J.; Boirie, Y.; Bruyère, O.; Cederholm, T.; Cooper, C.; Landi, F.; Rolland, Y.; Sayer, A.A.; et al. Sarcopenia: Revised European Consensus on Definition and Diagnosis. Age Ageing 2019, 48, 16–31. [Google Scholar] [CrossRef] [Green Version]
- Zamboni, M.; Rossi, A.P.; Fantin, F.; Budui, S.L.; Zoico, E.; Zamboni, G.A.; Mazzali, G. Predictors of Ectopic Fat in Humans. Curr. Obes. Rep. 2014, 3, 404–413. [Google Scholar] [CrossRef] [PubMed]
- Zamboni, M.; Nori, N.; Brunelli, A.; Zoico, E. How Does Adipose Tissue Contribute to Inflammageing? Exp. Gerontol. 2021, 143, 111162. [Google Scholar] [CrossRef]
- Santoro, A.; Bientinesi, E.; Monti, D. Immunosenescence and Inflammaging in the Aging Process: Age-Related Diseases or Longevity? Ageing Res. Rev. 2021, 71, 101422. [Google Scholar] [CrossRef]
- Johnson, K.O.; Shannon, O.M.; Matu, J.; Holliday, A.; Ispoglou, T.; Deighton, K. Differences in Circulating Appetite-Related Hormone Concentrations between Younger and Older Adults: A Systematic Review and Meta-Analysis. Aging Clin. Exp. Res. 2020, 32, 1233–1244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Picard, F.; Carter, S.; Caron, A.; Richard, D. Role of Leptin Resistance in the Development of Obesity in Older Patients. Clin. Interv. Aging 2013, 8, 829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kizer, J.R.; Arnold, A.M.; Jenny, N.S.; Cushman, M.; Strotmeyer, E.S.; Ives, D.G.; Ding, J.; Kritchevsky, S.B.; Chaves, P.H.M.; Hirsch, C.H.; et al. Longitudinal Changes in Adiponectin and Inflammatory Markers and Relation to Survival in the Oldest Old: The Cardiovascular Health Study All Stars Study. J. Gerontol. A Biol. Sci. Med. Sci. 2011, 66, 1100–1107. [Google Scholar] [CrossRef]
- Arai, Y.; Kamide, K.; Hirose, N. Adipokines and Aging: Findings From Centenarians and the Very Old. Front. Endocrinol. 2019, 10, 142. [Google Scholar] [CrossRef]
- Mancuso, P.; Bouchard, B. The Impact of Aging on Adipose Function and Adipokine Synthesis. Front. Endocrinol. 2019, 10, 137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baker, J.F.; Newman, A.B.; Kanaya, A.; Leonard, M.B.; Zemel, B.; Miljkovic, I.; Long, J.; Weber, D.; Harris, T.B. The Adiponectin Paradox in the Elderly: Associations With Body Composition, Physical Functioning, and Mortality. J. Gerontol. Ser. A 2019, 74, 247–253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Y.-H.; Zhu, H. Chronological Changes in Metabolism and Functions of Cultured Adipocytes: A Hypothesis for Cell Aging in Mature Adipocytes. Am. J. Physiol. Endocrinol. Metab. 2004, 286, E402–E410. [Google Scholar] [CrossRef]
- Zoico, E.; Rizzatti, V.; Policastro, G.; Tebon, M.; Darra, E.; Rossi, A.P.; Mazzali, G.; Fantin, F.; Zamboni, M. In Vitro Model of Chronological Aging of Adipocytes: Interrelationships with Hypoxia and Oxidation. Exp. Gerontol. 2019, 121, 81–90. [Google Scholar] [CrossRef]
- Zoico, E.; di Francesco, V.; Olioso, D.; Fratta Pasini, A.M.; Sepe, A.; Bosello, O.; Cinti, S.; Cominacini, L.; Zamboni, M. In Vitro Aging of 3T3-L1 Mouse Adipocytes Leads to Altered Metabolism and Response to Inflammation. Biogerontology 2010, 11, 111–122. [Google Scholar] [CrossRef]
- Palmer, A.K.; Kirkland, J.L. Aging and Adipose Tissue: Potential Interventions for Diabetes and Regenerative Medicine. Exp. Gerontol. 2016, 86, 97–105. [Google Scholar] [CrossRef] [Green Version]
- Lumeng, C.N.; Saltiel, A.R. Inflammatory Links between Obesity and Metabolic Disease. J. Clin. Investig. 2011, 121, 2111–2117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garg, S.K.; Delaney, C.; Shi, H.; Yung, R. Changes in Adipose Tissue Macrophages and T Cells during Aging. Crit. Rev. Immunol. 2014, 34, 1–14. [Google Scholar] [CrossRef]
- Nakagami, H. Cellular Senescence and Senescence-Associated T Cells as a Potential Therapeutic Target. Geriatr. Gerontol. Int. 2020, 20, 97–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalathookunnel Antony, A.; Lian, Z.; Wu, H. T Cells in Adipose Tissue in Aging. Front. Immunol. 2018, 9, 2945. [Google Scholar] [CrossRef]
- Kirkland, J.L.; Tchkonia, T.; Pirtskhalava, T.; Han, J.; Karagiannides, I. Adipogenesis and Aging: Does Aging Make Fat Go MAD? Exp. Gerontol. 2002, 37, 757–767. [Google Scholar] [CrossRef]
- Carobbio, S.; Pellegrinelli, V.; Vidal-Puig, A. Adipose Tissue Function and Expandability as Determinants of Lipotoxicity and the Metabolic Syndrome. Adv. Exp. Med. Biol. 2017, 960, 161–196. [Google Scholar] [CrossRef]
- Sepe, A.; Tchkonia, T.; Thomou, T.; Zamboni, M.; Kirkland, J.L. Aging and Regional Differences in Fat Cell Progenitors—A Mini-Review. Gerontology 2011, 57, 66–75. [Google Scholar] [CrossRef] [Green Version]
- Donato, A.J.; Henson, G.D.; Hart, C.R.; Layec, G.; Trinity, J.D.; Bramwell, R.C.; Enz, R.A.; Morgan, R.G.; Reihl, K.D.; Hazra, S.; et al. The Impact of Ageing on Adipose Structure, Function and Vasculature in the B6D2F1 Mouse: Evidence of Significant Multisystem Dysfunction. J. Physiol. 2014, 592, 4083–4096. [Google Scholar] [CrossRef]
- Tardif, N.; Salles, J.; Guillet, C.; Tordjman, J.; Reggio, S.; Landrier, J.-F.; Giraudet, C.; Patrac, V.; Bertrand-Michel, J.; Migne, C.; et al. Muscle Ectopic Fat Deposition Contributes to Anabolic Resistance in Obese Sarcopenic Old Rats through EIF2α Activation. Aging Cell 2014, 13, 1001–1011. [Google Scholar] [CrossRef]
- Khan, T.; Muise, E.S.; Iyengar, P.; Wang, Z.V.; Chandalia, M.; Abate, N.; Zhang, B.B.; Bonaldo, P.; Chua, S.; Scherer, P.E. Metabolic Dysregulation and Adipose Tissue Fibrosis: Role of Collagen VI. Mol. Cell. Biol. 2009, 29, 1575–1591. [Google Scholar] [CrossRef] [Green Version]
- Tchkonia, T.; Lenburg, M.; Thomou, T.; Giorgadze, N.; Frampton, G.; Pirtskhalava, T.; Cartwright, A.; Cartwright, M.; Flanagan, J.; Karagiannides, I.; et al. Identification of Depot-Specific Human Fat Cell Progenitors through Distinct Expression Profiles and Developmental Gene Patterns. Am. J. Physiol. Endocrinol. Metab. 2007, 292, E298–E307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tchkonia, T.; Kirkland, J.L. Aging, Cell Senescence, and Chronic Disease: Emerging Therapeutic Strategies. JAMA 2018, 320, 1319–1320. [Google Scholar] [CrossRef]
- Severinsen, M.C.K.; Pedersen, B.K. Muscle-Organ Crosstalk: The Emerging Roles of Myokines. Endocr. Rev. 2020, 41, 594–609. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, B.K.; Febbraio, M.A. Muscles, Exercise and Obesity: Skeletal Muscle as a Secretory Organ. Nat. Rev. Endocrinol. 2012, 8, 457–465. [Google Scholar] [CrossRef]
- Henningsen, J.; Rigbolt, K.T.G.; Blagoev, B.; Pedersen, B.K.; Kratchmarova, I. Dynamics of the Skeletal Muscle Secretome during Myoblast Differentiation. Mol. Cell. Proteom. 2010, 9, 2482–2496. [Google Scholar] [CrossRef] [Green Version]
- Henningsen, J.; Pedersen, B.K.; Kratchmarova, I. Quantitative Analysis of the Secretion of the MCP Family of Chemokines by Muscle Cells. Mol. Biosyst. 2011, 7, 311–321. [Google Scholar] [CrossRef] [Green Version]
- Waldemer-Streyer, R.J.; Kim, D.; Chen, J. Muscle Cell-Derived Cytokines in Skeletal Muscle Regeneration. FEBS J. 2022, 24, 16372. [Google Scholar] [CrossRef] [PubMed]
- Florini, J.R.; Ewton, D.Z.; Coolican, S.A. Growth Hormone and the Insulin-Like Growth Factor System in Myogenesis*. Endocr. Rev. 1996, 17, 481–517. [Google Scholar] [CrossRef] [Green Version]
- Florini, J.R.; Magri, K.A.; Ewton, D.Z.; James, P.L.; Grindstaff, K.; Rotwein, P.S. “Spontaneous” Differentiation of Skeletal Myoblasts Is Dependent upon Autocrine Secretion of Insulin-like Growth Factor-II. J. Biol. Chem. 1991, 266, 15917–15923. [Google Scholar] [CrossRef]
- Yoshida, T.; Delafontaine, P. Mechanisms of IGF-1-Mediated Regulation of Skeletal Muscle Hypertrophy and Atrophy. Cells 2020, 9, 1970. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, S.S.; Ahmad, K.; Lee, E.J.; Lee, Y.-H.; Choi, I. Implications of Insulin-Like Growth Factor-1 in Skeletal Muscle and Various Diseases. Cells 2020, 9, 1773. [Google Scholar] [CrossRef] [PubMed]
- Haugen, F.; Norheim, F.; Lian, H.; Wensaas, A.J.; Dueland, S.; Berg, O.; Funderud, A.; Skålhegg, B.S.; Raastad, T.; Drevon, C.A. IL-7 Is Expressed and Secreted by Human Skeletal Muscle Cells. Am. J. Physiol. Cell Physiol. 2010, 298, C807–C816. [Google Scholar] [CrossRef] [Green Version]
- Olson, E.N.; Sternberg, E.; Hu, J.S.; Spizz, G.; Wilcox, C. Regulation of Myogenic Differentiation by Type Beta Transforming Growth Factor. J. Cell. Biol. 1986, 103, 1799–1805. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joulia, D.; Bernardi, H.; Garandel, V.; Rabenoelina, F.; Vernus, B.; Cabello, G. Mechanisms Involved in the Inhibition of Myoblast Proliferation and Differentiation by Myostatin. Exp. Cell. Res. 2003, 286, 263–275. [Google Scholar] [CrossRef]
- McPherron, A.C.; Lawler, A.M.; Lee, S.J. Regulation of Skeletal Muscle Mass in Mice by a New TGF-Beta Superfamily Member. Nature 1997, 387, 83–90. [Google Scholar] [CrossRef]
- Grobet, L.; Martin, L.J.; Poncelet, D.; Pirottin, D.; Brouwers, B.; Riquet, J.; Schoeberlein, A.; Dunner, S.; Ménissier, F.; Massabanda, J.; et al. A Deletion in the Bovine Myostatin Gene Causes the Double-Muscled Phenotype in Cattle. Nat. Genet. 1997, 17, 71–74. [Google Scholar] [CrossRef] [PubMed]
- Mosher, D.S.; Quignon, P.; Bustamante, C.D.; Sutter, N.B.; Mellersh, C.S.; Parker, H.G.; Ostrander, E.A. A Mutation in the Myostatin Gene Increases Muscle Mass and Enhances Racing Performance in Heterozygote Dogs. PLoS Genet. 2007, 3, e79. [Google Scholar] [CrossRef] [PubMed]
- Gilson, H.; Schakman, O.; Kalista, S.; Lause, P.; Tsuchida, K.; Thissen, J.-P. Follistatin Induces Muscle Hypertrophy through Satellite Cell Proliferation and Inhibition of Both Myostatin and Activin. Am. J. Physiol. Endocrinol. Metab. 2009, 297, E157–E164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- el Shafey, N.; Guesnon, M.; Simon, F.; Deprez, E.; Cosette, J.; Stockholm, D.; Scherman, D.; Bigey, P.; Kichler, A. Inhibition of the Myostatin/Smad Signaling Pathway by Short Decorin-Derived Peptides. Exp. Cell. Res. 2016, 341, 187–195. [Google Scholar] [CrossRef] [PubMed]
- Serrano, A.L.; Baeza-Raja, B.; Perdiguero, E.; Jardí, M.; Muñoz-Cánoves, P. Interleukin-6 Is an Essential Regulator of Satellite Cell-Mediated Skeletal Muscle Hypertrophy. Cell Metab. 2008, 7, 33–44. [Google Scholar] [CrossRef] [Green Version]
- Broholm, C.; Pedersen, B.K. Leukaemia Inhibitory Factor--an Exercise-Induced Myokine. Exerc. Immunol. Rev. 2010, 16, 77–85. [Google Scholar]
- Matthews, V.B.; Aström, M.-B.; Chan, M.H.S.; Bruce, C.R.; Krabbe, K.S.; Prelovsek, O.; Akerström, T.; Yfanti, C.; Broholm, C.; Mortensen, O.H.; et al. Brain-Derived Neurotrophic Factor Is Produced by Skeletal Muscle Cells in Response to Contraction and Enhances Fat Oxidation via Activation of AMP-Activated Protein Kinase. Diabetologia 2009, 52, 1409–1418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carey, A.L.; Steinberg, G.R.; Macaulay, S.L.; Thomas, W.G.; Holmes, A.G.; Ramm, G.; Prelovsek, O.; Hohnen-Behrens, C.; Watt, M.J.; James, D.E.; et al. Interleukin-6 Increases Insulin-Stimulated Glucose Disposal in Humans and Glucose Uptake and Fatty Acid Oxidation In Vitro via AMP-Activated Protein Kinase. Diabetes 2006, 55, 2688–2697. [Google Scholar] [CrossRef] [Green Version]
- Petersen, E.W.; Carey, A.L.; Sacchetti, M.; Steinberg, G.R.; Macaulay, S.L.; Febbraio, M.A.; Pedersen, B.K. Acute IL-6 Treatment Increases Fatty Acid Turnover in Elderly Humans in Vivo and in Tissue Culture in Vitro. Am. J. Physiol. Endocrinol. Metab. 2005, 288, E155–E162. [Google Scholar] [CrossRef]
- Colombo, E.; Bedogni, F.; Lorenzetti, I.; Landsberger, N.; Previtali, S.C.; Farina, C. Autocrine and Immune Cell-Derived BDNF in Human Skeletal Muscle: Implications for Myogenesis and Tissue Regeneration. J. Pathol. 2013, 231, 190–198. [Google Scholar] [CrossRef] [PubMed]
- Guo, A.; Li, K.; Xiao, Q. Sarcopenic Obesity: Myokines as Potential Diagnostic Biomarkers and Therapeutic Targets? Exp. Gerontol. 2020, 139, 111022. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Lang, N.; Cheng, Z.-F. Serum Levels of Brain-Derived Neurotrophic Factor Are Associated with Diabetes Risk, Complications, and Obesity: A Cohort Study from Chinese Patients with Type 2 Diabetes. Mol. Neurobiol. 2016, 53, 5492–5499. [Google Scholar] [CrossRef]
- Allen, D.L.; Hittel, D.S.; McPherron, A.C. Expression and Function of Myostatin in Obesity, Diabetes, and Exercise Adaptation. Med. Sci. Sports Exerc. 2011, 43, 1828–1835. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, S.; Durstine, J.L.; Koh, H.-J.; Carver, W.E.; Frizzell, N.; Carson, J.A. Acute Myotube Protein Synthesis Regulation by IL-6-Related Cytokines. Am. J. Physiol. Cell Physiol. 2017, 313, C487–C500. [Google Scholar] [CrossRef] [Green Version]
- Nielsen, A.R.; Pedersen, B.K. The Biological Roles of Exercise-Induced Cytokines: IL-6, IL-8, and IL-15. Appl. Physiol. Nutr. Metab. 2007, 32, 833–839. [Google Scholar] [CrossRef]
- Re Cecconi, A.D.; Forti, M.; Chiappa, M.; Zhu, Z.; Zingman, L.V.; Cervo, L.; Beltrame, L.; Marchini, S.; Piccirillo, R. Musclin, A Myokine Induced by Aerobic Exercise, Retards Muscle Atrophy During Cancer Cachexia in Mice. Cancers 2019, 11, 1541. [Google Scholar] [CrossRef] [Green Version]
- Kota, J.; Handy, C.R.; Haidet, A.M.; Montgomery, C.L.; Eagle, A.; Rodino-Klapac, L.R.; Tucker, D.; Shilling, C.J.; Therlfall, W.R.; Walker, C.M.; et al. Follistatin Gene Delivery Enhances Muscle Growth and Strength in Nonhuman Primates. Sci. Transl. Med. 2009, 1, 6ra15. [Google Scholar] [CrossRef] [Green Version]
- Miura, T.; Kishioka, Y.; Wakamatsu, J.; Hattori, A.; Hennebry, A.; Berry, C.J.; Sharma, M.; Kambadur, R.; Nishimura, T. Decorin Binds Myostatin and Modulates Its Activity to Muscle Cells. Biochem. Biophys. Res. Commun. 2006, 340, 675–680. [Google Scholar] [CrossRef]
- Kanzleiter, T.; Rath, M.; Görgens, S.W.; Jensen, J.; Tangen, D.S.; Kolnes, A.J.; Kolnes, K.J.; Lee, S.; Eckel, J.; Schürmann, A.; et al. The Myokine Decorin Is Regulated by Contraction and Involved in Muscle Hypertrophy. Biochem. Biophys. Res. Commun. 2014, 450, 1089–1094. [Google Scholar] [CrossRef]
- Seldin, M.M.; Peterson, J.M.; Byerly, M.S.; Wei, Z.; Wong, G.W. Myonectin (CTRP15), a Novel Myokine That Links Skeletal Muscle to Systemic Lipid Homeostasis. J. Biol. Chem. 2012, 287, 11968–11980. [Google Scholar] [CrossRef] [PubMed]
- Subbotina, E.; Sierra, A.; Zhu, Z.; Gao, Z.; Koganti, S.R.K.; Reyes, S.; Stepniak, E.; Walsh, S.A.; Acevedo, M.R.; Perez-Terzic, C.M.; et al. Musclin Is an Activity-Stimulated Myokine That Enhances Physical Endurance. Proc. Natl. Acad. Sci. USA 2015, 112, 16042–16047. [Google Scholar] [CrossRef] [Green Version]
- Bruce, C.R.; Dyck, D.J. Cytokine Regulation of Skeletal Muscle Fatty Acid Metabolism: Effect of Interleukin-6 and Tumor Necrosis Factor-α. Am. J. Physiol. Endocrinol. Metab. 2004, 287, E616–E621. [Google Scholar] [CrossRef] [PubMed]
- Boström, P.; Wu, J.; Jedrychowski, M.P.; Korde, A.; Ye, L.; Lo, J.C.; Rasbach, K.A.; Boström, E.A.; Choi, J.H.; Long, J.Z.; et al. A PGC1-α-Dependent Myokine That Drives Brown-Fat-like Development of White Fat and Thermogenesis. Nature 2012, 481, 463–468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quinn, L.; Straitbodey, L.; Anderson, B.; Argiles, J.; Havel, P. Interleukin-15 Stimulates Adiponectin Secretion by 3T3-L1 Adipocytes: Evidence for a Skeletal Muscle-to-Fat Signaling Pathway. Cell. Biol. Int. 2005, 29, 449–457. [Google Scholar] [CrossRef] [Green Version]
- Krolopp, J.E.; Thornton, S.M.; Abbott, M.J. IL-15 Activates the Jak3/STAT3 Signaling Pathway to Mediate Glucose Uptake in Skeletal Muscle Cells. Front. Physiol. 2016, 7, 626. [Google Scholar] [CrossRef] [Green Version]
- Aoi, W.; Hirano, N.; Lassiter, D.G.; Björnholm, M.; Chibalin, A.V.; Sakuma, K.; Tanimura, Y.; Mizushima, K.; Takagi, T.; Naito, Y.; et al. Secreted Protein Acidic and Rich in Cysteine (SPARC) Improves Glucose Tolerance via AMP-activated Protein Kinase Activation. FASEB J. 2019, 33, 10551–10562. [Google Scholar] [CrossRef] [Green Version]
- Brandt, N.; O’Neill, H.M.; Kleinert, M.; Schjerling, P.; Vernet, E.; Steinberg, G.R.; Richter, E.A.; Jørgensen, S.B. Leukemia Inhibitory Factor Increases Glucose Uptake in Mouse Skeletal Muscle. Am. J. Physiol. -Endocrinol. Metab. 2015, 309, E142–E153. [Google Scholar] [CrossRef] [Green Version]
- Chattopadhyay, T.; Singh, R.R.; Gupta, S.; Surolia, A. Bone Morphogenetic Protein-7 (BMP-7) Augments Insulin Sensitivity in Mice with Type II Diabetes Mellitus by Potentiating PI3K/AKT Pathway. BioFactors 2017, 43, 195–209. [Google Scholar] [CrossRef]
- Wu, H.-K.; Zhang, Y.; Cao, C.-M.; Hu, X.; Fang, M.; Yao, Y.; Jin, L.; Chen, G.; Jiang, P.; Zhang, S.; et al. Glucose-Sensitive Myokine/Cardiokine MG53 Regulates Systemic Insulin Response and Metabolic Homeostasis. Circulation 2019, 139, 901–914. [Google Scholar] [CrossRef]
- Perrini, S.; Laviola, L.; Carreira, M.C.; Cignarelli, A.; Natalicchio, A.; Giorgino, F. The GH/IGF1 Axis and Signaling Pathways in the Muscle and Bone: Mechanisms Underlying Age-Related Skeletal Muscle Wasting and Osteoporosis. J. Endocrinol. 2010, 205, 201–210. [Google Scholar] [CrossRef] [PubMed]
- Kaji, H. Effects of Myokines on Bone. Bonekey Rep. 2016, 5, 826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Axmann, R.; Böhm, C.; Krönke, G.; Zwerina, J.; Smolen, J.; Schett, G. Inhibition of Interleukin-6 Receptor Directly Blocks Osteoclast Formation In Vitro and In Vivo. Arthritis Rheum. 2009, 60, 2747–2756. [Google Scholar] [CrossRef] [PubMed]
- Kharitonenkov, A.; Shiyanova, T.L.; Koester, A.; Ford, A.M.; Micanovic, R.; Galbreath, E.J.; Sandusky, G.E.; Hammond, L.J.; Moyers, J.S.; Owens, R.A.; et al. FGF-21 as a Novel Metabolic Regulator. J. Clin. Investig. 2005, 115, 1627–1635. [Google Scholar] [CrossRef] [Green Version]
- Rao, R.R.; Long, J.Z.; White, J.P.; Svensson, K.J.; Lou, J.; Lokurkar, I.; Jedrychowski, M.P.; Ruas, J.L.; Wrann, C.D.; Lo, J.C.; et al. Meteorin-like Is a Hormone That Regulates Immune-Adipose Interactions to Increase Beige Fat Thermogenesis. Cell 2014, 157, 1279–1291. [Google Scholar] [CrossRef] [Green Version]
- Véniant, M.M.; Sivits, G.; Helmering, J.; Komorowski, R.; Lee, J.; Fan, W.; Moyer, C.; Lloyd, D.J. Pharmacologic Effects of FGF21 Are Independent of the “Browning” of White Adipose Tissue. Cell Metab. 2015, 21, 731–738. [Google Scholar] [CrossRef] [Green Version]
- Roberts, L.D.; Boström, P.; O’Sullivan, J.F.; Schinzel, R.T.; Lewis, G.D.; Dejam, A.; Lee, Y.-K.; Palma, M.J.; Calhoun, S.; Georgiadi, A.; et al. β-Aminoisobutyric Acid Induces Browning of White Fat and Hepatic β-Oxidation and Is Inversely Correlated with Cardiometabolic Risk Factors. Cell Metab. 2014, 19, 96–108. [Google Scholar] [CrossRef] [Green Version]
- Tanianskii, D.A.; Jarzebska, N.; Birkenfeld, A.L.; O’Sullivan, J.F.; Rodionov, R.N. Beta-Aminoisobutyric Acid as a Novel Regulator of Carbohydrate and Lipid Metabolism. Nutrients 2019, 11, 524. [Google Scholar] [CrossRef] [Green Version]
- Singh, R.; Braga, M.; Pervin, S. Regulation of Brown Adipocyte Metabolism by Myostatin/Follistatin Signaling. Front. Cell Dev. Biol. 2014, 2, 60. [Google Scholar] [CrossRef] [Green Version]
- Peterson, J.M.; Mart, R.; Bond, C.E. Effect of Obesity and Exercise on the Expression of the Novel Myokines, Myonectin and Fibronectin Type III Domain Containing 5. PeerJ 2014, 2, e605. [Google Scholar] [CrossRef] [Green Version]
- Tseng, Y.-H.; Kokkotou, E.; Schulz, T.J.; Huang, T.L.; Winnay, J.N.; Taniguchi, C.M.; Tran, T.T.; Suzuki, R.; Espinoza, D.O.; Yamamoto, Y.; et al. New Role of Bone Morphogenetic Protein 7 in Brown Adipogenesis and Energy Expenditure. Nature 2008, 454, 1000–1004. [Google Scholar] [CrossRef] [PubMed]
- Cuevas-Ramos, D.; Aguilar-Salinas, C.A. Modulation of Energy Balance by Fibroblast Growth Factor 21. Horm. Mol. Biol. Clin. Investig 2016, 30. [Google Scholar] [CrossRef] [PubMed]
- Staiger, H.; Haas, C.; Machann, J.; Werner, R.; Weisser, M.; Schick, F.; Machicao, F.; Stefan, N.; Fritsche, A.; Häring, H.-U. Muscle-Derived Angiopoietin-Like Protein 4 Is Induced by Fatty Acids via Peroxisome Proliferator–Activated Receptor (PPAR)-δ and Is of Metabolic Relevance in Humans. Diabetes 2009, 58, 579–589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakuma, K.; Yamaguchi, A. The Recent Understanding of the Neurotrophin’s Role in Skeletal Muscle Adaptation. J. Biomed. Biotechnol. 2011, 2011, 201696. [Google Scholar] [CrossRef] [Green Version]
- Taetzsch, T.; Tenga, M.J.; Valdez, G. Muscle Fibers Secrete FGFBP1 to Slow Degeneration of Neuromuscular Synapses during Aging and Progression of ALS. J. Neurosci. 2017, 37, 70–82. [Google Scholar] [CrossRef] [Green Version]
- Hoier, B.; Hellsten, Y. Exercise-Induced Capillary Growth in Human Skeletal Muscle and the Dynamics of VEGF. Microcirculation 2014, 21, 301–314. [Google Scholar] [CrossRef]
- Jensen, L.; Schjerling, P.; Hellsten, Y. Regulation of VEGF and BFGF MRNA Expression and Other Proliferative Compounds in Skeletal Muscle Cells. Angiogenesis 2004, 7, 255–267. [Google Scholar] [CrossRef]
- Larsson, L.; Degens, H.; Li, M.; Salviati, L.; Lee, Y.I.; Thompson, W.; Kirkland, J.L.; Sandri, M. Sarcopenia: Aging-Related Loss of Muscle Mass and Function. Physiol. Rev. 2019, 99, 427–511. [Google Scholar] [CrossRef]
- Correa-De-Araujo, R.; Rossi, A.P.; Zamboni, M.; Addison, O.; Miljkovic, I.; Goodpaster, B. Editorial: Muscle Quality in Skeletal Muscle Function Deficit: Recent Advances and Potential Clinical and Therapeutic Implications. Front. Physiol. 2022, 13, 847883. [Google Scholar] [CrossRef]
- Li, J.; Wang, Z.; Li, C.; Song, Y.; Wang, Y.; Bo, H.; Zhang, Y. Impact of Exercise and Aging on Mitochondrial Homeostasis in Skeletal Muscle: Roles of ROS and Epigenetics. Cells 2022, 11, 2086. [Google Scholar] [CrossRef]
- Huh, J.Y.; Mougios, V.; Kabasakalis, A.; Fatouros, I.; Siopi, A.; Douroudos, I.I.; Filippaios, A.; Panagiotou, G.; Park, K.H.; Mantzoros, C.S. Exercise-Induced Irisin Secretion Is Independent of Age or Fitness Level and Increased Irisin May Directly Modulate Muscle Metabolism Through AMPK Activation. J. Clin. Endocrinol. Metab. 2014, 99, E2154–E2161. [Google Scholar] [CrossRef] [PubMed]
- Elia, A.; Cannavo, A.; Gambino, G.; Cimini, M.; Ferrara, N.; Kishore, R.; Paolocci, N.; Rengo, G. Aging Is Associated with Cardiac Autonomic Nerve Fiber Depletion and Reduced Cardiac and Circulating BDNF Levels. J. Geriatr. Cardiol. 2021, 18, 549–559. [Google Scholar] [CrossRef] [PubMed]
- Dray, C.; Knauf, C.; Daviaud, D.; Waget, A.; Boucher, J.; Buléon, M.; Cani, P.D.; Attané, C.; Guigné, C.; Carpéné, C.; et al. Apelin Stimulates Glucose Utilization in Normal and Obese Insulin-Resistant Mice. Cell Metab. 2008, 8, 437–445. [Google Scholar] [CrossRef] [PubMed]
- Peng, L.-N.; Lee, W.-J.; Liu, L.-K.; Lin, M.-H.; Chen, L.-K. Healthy Community-Living Older Men Differ from Women in Associations between Myostatin Levels and Skeletal Muscle Mass. J. Cachexia Sarcopenia Muscle 2018, 9, 635–642. [Google Scholar] [CrossRef]
- Yarasheski, K.E.; Bhasin, S.; Sinha-Hikim, I.; Pak-Loduca, J.; Gonzalez-Cadavid, N.F. Serum Myostatin-Immunoreactive Protein Is Increased in 60-92 Year Old Women and Men with Muscle Wasting. J. Nutr. Health Aging 2002, 6, 343–348. [Google Scholar]
- Jeanplong, F.; Osepchook, C.C.; Falconer, S.J.; Smith, H.K.; Bass, J.J.; McMahon, C.D.; Oldham, J.M. Undernutrition Regulates the Expression of a Novel Splice Variant of Myostatin and Insulin-like Growth Factor 1 in Ovine Skeletal Muscle. Domest. Anim. Endocrinol. 2015, 52, 17–24. [Google Scholar] [CrossRef]
- Wueest, S.; Item, F.; Boyle, C.N.; Jirkof, P.; Cesarovic, N.; Ellingsgaard, H.; Böni-Schnetzler, M.; Timper, K.; Arras, M.; Donath, M.Y.; et al. Interleukin-6 Contributes to Early Fasting-Induced Free Fatty Acid Mobilization in Mice. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2014, 306, R861–R867. [Google Scholar] [CrossRef]
- Wedell-Neergaard, A.-S.; Lang Lehrskov, L.; Christensen, R.H.; Legaard, G.E.; Dorph, E.; Larsen, M.K.; Launbo, N.; Fagerlind, S.R.; Seide, S.K.; Nymand, S.; et al. Exercise-Induced Changes in Visceral Adipose Tissue Mass Are Regulated by IL-6 Signaling: A Randomized Controlled Trial. Cell Metab. 2019, 29, 844–855.e3. [Google Scholar] [CrossRef]
- Graf, C.; Ferrari, N. Metabolic Health—The Role of Adipo-Myokines. Int. J. Mol. Sci. 2019, 20, 6159. [Google Scholar] [CrossRef] [Green Version]
- Colaianni, G.; Mongelli, T.; Cuscito, C.; Pignataro, P.; Lippo, L.; Spiro, G.; Notarnicola, A.; Severi, I.; Passeri, G.; Mori, G.; et al. Irisin Prevents and Restores Bone Loss and Muscle Atrophy in Hind-Limb Suspended Mice. Sci. Rep. 2017, 7, 2811. [Google Scholar] [CrossRef] [Green Version]
- Gomarasca, M.; Banfi, G.; Lombardi, G. Myokines: The Endocrine Coupling of Skeletal Muscle and Bone. Adv. Clin. Chem. 2020, 94, 155–218. [Google Scholar] [CrossRef] [PubMed]
- Seldin, M.M.; Wong, G.W. Regulation of Tissue Crosstalk by Skeletal Muscle-Derived Myonectin and Other Myokines. Adipocyte 2012, 1, 200–202. [Google Scholar] [CrossRef] [PubMed]
- Katsu-Jiménez, Y.; Giménez-Cassina, A. Fibroblast Growth Factor-21 Promotes Ketone Body Utilization in Neurons through Activation of AMP-Dependent Kinase. Mol. Cell. Neurosci. 2019, 101, 103415. [Google Scholar] [CrossRef] [PubMed]
- Lin, Z.; Tian, H.; Lam, K.S.L.; Lin, S.; Hoo, R.C.L.; Konishi, M.; Itoh, N.; Wang, Y.; Bornstein, S.R.; Xu, A.; et al. Adiponectin Mediates the Metabolic Effects of FGF21 on Glucose Homeostasis and Insulin Sensitivity in Mice. Cell Metab. 2013, 17, 779–789. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, C.-S.; Joe, Y.; Choi, H.-S.; Back, S.H.; Park, J.W.; Chung, H.T.; Roh, E.; Kim, M.-S.; Ha, T.Y.; Yu, R. Deficiency of Fibroblast Growth Factor 21 Aggravates Obesity-Induced Atrophic Responses in Skeletal Muscle. J. Inflamm. 2019, 16, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castan-Laurell, I.; Boucher, J.; Dray, C.; Daviaud, D.; Guigné, C.; Valet, P. Apelin, a Novel Adipokine over-Produced in Obesity: Friend or Foe? Mol. Cell. Endocrinol. 2005, 245, 7–9. [Google Scholar] [CrossRef]
- Boucher, J.; Masri, B.; Daviaud, D.; Gesta, S.; Guigné, C.; Mazzucotelli, A.; Castan-Laurell, I.; Tack, I.; Knibiehler, B.; Carpéné, C.; et al. Apelin, a Newly Identified Adipokine Up-Regulated by Insulin and Obesity. Endocrinology 2005, 146, 1764–1771. [Google Scholar] [CrossRef]
- Besse-Patin, A.; Montastier, E.; Vinel, C.; Castan-Laurell, I.; Louche, K.; Dray, C.; Daviaud, D.; Mir, L.; Marques, M.-A.; Thalamas, C.; et al. Effect of Endurance Training on Skeletal Muscle Myokine Expression in Obese Men: Identification of Apelin as a Novel Myokine. Int. J. Obes. 2014, 38, 707–713. [Google Scholar] [CrossRef]
- Higuchi, K.; Masaki, T.; Gotoh, K.; Chiba, S.; Katsuragi, I.; Tanaka, K.; Kakuma, T.; Yoshimatsu, H. Apelin, an APJ Receptor Ligand, Regulates Body Adiposity and Favors the Messenger Ribonucleic Acid Expression of Uncoupling Proteins in Mice. Endocrinology 2007, 148, 2690–2697. [Google Scholar] [CrossRef]
- Son, J.S.; Kim, H.J.; Son, Y.; Lee, H.; Chae, S.A.; Seong, J.K.; Song, W. Effects of exercise-induced apelin levels on skeletal muscle and their capillarization in type 2 diabetic rats. Muscle Nerve. 2017, 56, 1155–1163. [Google Scholar] [CrossRef]
- Yue, P.; Jin, H.; Aillaud, M.; Deng, A.C.; Azuma, J.; Asagami, T.; Kundu, R.K.; Reaven, G.M.; Quertermous, T.; Tsao, P.S. Apelin Is Necessary for the Maintenance of Insulin Sensitivity. Am. J. Physiol. Endocrinol. Metab. 2010, 298, E59–E67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Attané, C.; Foussal, C.; le Gonidec, S.; Benani, A.; Daviaud, D.; Wanecq, E.; Guzmán-Ruiz, R.; Dray, C.; Bezaire, V.; Rancoule, C.; et al. Apelin Treatment Increases Complete Fatty Acid Oxidation, Mitochondrial Oxidative Capacity, and Biogenesis in Muscle of Insulin-Resistant Mice. Diabetes 2012, 61, 310–320. [Google Scholar] [CrossRef] [PubMed]
- Svärd, J.; Røst, T.H.; Sommervoll, C.E.N.; Haugen, C.; Gudbrandsen, O.A.; Mellgren, A.E.; Rødahl, E.; Fernø, J.; Dankel, S.N.; Sagen, J.V.; et al. Absence of the Proteoglycan Decorin Reduces Glucose Tolerance in Overfed Male Mice. Sci. Rep. 2019, 9, 4614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodríguez, A.; Catalán, V.; Ramírez, B.; Unamuno, X.; Portincasa, P.; Gómez-Ambrosi, J.; Frühbeck, G.; Becerril, S. Impact of Adipokines and Myokines on Fat Browning. J. Physiol. Biochem. 2020, 76, 227–240. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Kim, Y.J.; Seong, J.K. AMP-Activated Protein Kinase Activation in Skeletal Muscle Modulates Exercise-Induced Uncoupled Protein 1 Expression in Brown Adipocyte in Mouse Model. J. Physiol. 2022, 600, 2359–2376. [Google Scholar] [CrossRef]
- Severinsen, M.C.K.; Schéele, C.; Pedersen, B.K. Exercise and Browning of White Adipose Tissue—A Translational Perspective. Curr. Opin. Pharmacol. 2020, 52, 18–24. [Google Scholar] [CrossRef]
- Scheel, A.K.; Espelage, L.; Chadt, A. Many Ways to Rome: Exercise, Cold Exposure and Diet-Do They All Affect BAT Activation and WAT Browning in the Same Manner? Int. J. Mol. Sci. 2022, 23, 4759. [Google Scholar] [CrossRef]
- Kamei, Y.; Hatazawa, Y.; Uchitomi, R.; Yoshimura, R.; Miura, S. Regulation of Skeletal Muscle Function by Amino Acids. Nutrients 2020, 12, 261. [Google Scholar] [CrossRef] [Green Version]
- Jung, T.W.; Park, H.S.; Choi, G.H.; Kim, D.; Lee, T. β-Aminoisobutyric Acid Attenuates LPS-Induced Inflammation and Insulin Resistance in Adipocytes through AMPK-Mediated Pathway. J. Biomed. Sci. 2018, 25, 27. [Google Scholar] [CrossRef] [Green Version]
- Ginter, E.; Simko, V. Recent Data on Obesity Research: β-Aminoisobutyric Acid. Bratisl. Med. J. 2014, 115, 492–493. [Google Scholar] [CrossRef]
- Hangelbroek, R.W.J.; Fazelzadeh, P.; Tieland, M.; Boekschoten, M.V.; Hooiveld, G.J.E.J.; van Duynhoven, J.P.M.; Timmons, J.A.; Verdijk, L.B.; de Groot, L.C.P.G.M.; van Loon, L.J.C.; et al. Expression of Protocadherin Gamma in Skeletal Muscle Tissue Is Associated with Age and Muscle Weakness. J. Cachexia Sarcopenia Muscle 2016, 7, 604–614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baht, G.S.; Bareja, A.; Lee, D.E.; Rao, R.R.; Huang, R.; Huebner, J.L.; Bartlett, D.B.; Hart, C.R.; Gibson, J.R.; Lanza, I.R.; et al. Meteorin-like Facilitates Skeletal Muscle Repair through a Stat3/IGF-1 Mechanism. Nat. Metab. 2020, 2, 278–289. [Google Scholar] [CrossRef]
- Akinci, G.; Topaloglu, H.; Demir, T.; Danyeli, A.E.; Talim, B.; Keskin, F.E.; Kadioglu, P.; Talip, E.; Altay, C.; Yaylali, G.F.; et al. Clinical Spectra of Neuromuscular Manifestations in Patients with Lipodystrophy: A Multicenter Study. Neuromuscul. Disord. 2017, 27, 923–930. [Google Scholar] [CrossRef] [PubMed]
- Unger, R.H. Longevity, Lipotoxicity and Leptin: The Adipocyte Defense against Feasting and Famine. Biochimie 2005, 87, 57–64. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Li, Y.; Duan, Y.; Hu, C.-A.A.; Tang, Y.; Yin, Y. Myokines and Adipokines: Involvement in the Crosstalk between Skeletal Muscle and Adipose Tissue. Cytokine Growth Factor Rev. 2017, 33, 73–82. [Google Scholar] [CrossRef]
- Nicholson, T.; Church, C.; Baker, D.J.; Jones, S.W. The Role of Adipokines in Skeletal Muscle Inflammation and Insulin Sensitivity. J. Inflamm. 2018, 15, 9. [Google Scholar] [CrossRef] [Green Version]
- Lynch, G.M.; Murphy, C.H.; de Castro, E.M.; Roche, H.M. Inflammation and Metabolism: The Role of Adiposity in Sarcopenic Obesity. Proc. Nutr. Soc. 2020, 79, 435–447. [Google Scholar] [CrossRef]
- Minard, A.Y.; Tan, S.-X.; Yang, P.; Fazakerley, D.J.; Domanova, W.; Parker, B.L.; Humphrey, S.J.; Jothi, R.; Stöckli, J.; James, D.E. MTORC1 Is a Major Regulatory Node in the FGF21 Signaling Network in Adipocytes. Cell Rep. 2016, 17, 29–36. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez, A.; Catalán, V.; Gómez-Ambrosi, J.; García-Navarro, S.; Rotellar, F.; Valentí, V.; Silva, C.; Gil, M.J.; Salvador, J.; Burrell, M.A.; et al. Insulin- and Leptin-Mediated Control of Aquaglyceroporins in Human Adipocytes and Hepatocytes Is Mediated via the PI3K/Akt/MTOR Signaling Cascade. J. Clin. Endocrinol. Metab. 2011, 96, E586–E597. [Google Scholar] [CrossRef] [Green Version]
- Arounleut, P.; Bowser, M.; Upadhyay, S.; Shi, X.-M.; Fulzele, S.; Johnson, M.H.; Stranahan, A.M.; Hill, W.D.; Isales, C.M.; Hamrick, M.W. Absence of Functional Leptin Receptor Isoforms in the POUND (Lepr(Db/Lb)) Mouse Is Associated with Muscle Atrophy and Altered Myoblast Proliferation and Differentiation. PLoS ONE 2013, 8, e72330. [Google Scholar] [CrossRef]
- Collins, K.H.; Gui, C.; Ely, E.V.; Lenz, K.L.; Harris, C.A.; Guilak, F.; Meyer, G.A. Leptin Mediates the Regulation of Muscle Mass and Strength by Adipose Tissue. J. Physiol. 2022, 600, 3795–3817. [Google Scholar] [CrossRef] [PubMed]
- Krause, M.P.; Liu, Y.; Vu, V.; Chan, L.; Xu, A.; Riddell, M.C.; Sweeney, G.; Hawke, T.J. Adiponectin Is Expressed by Skeletal Muscle Fibers and Influences Muscle Phenotype and Function. Am. J. Physiol. Cell Physiol. 2008, 295, C203–C212. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Li, F.; Kong, X.; Yuan, X.; Wang, W.; Huang, R.; Li, T.; Geng, M.; Wu, G.; Yin, Y. Chemerin Regulates Proliferation and Differentiation of Myoblast Cells via ERK1/2 and MTOR Signaling Pathways. Cytokine 2012, 60, 646–652. [Google Scholar] [CrossRef] [PubMed]
- Becker, M.; Rabe, K.; Lebherz, C.; Zugwurst, J.; Göke, B.; Parhofer, K.G.; Lehrke, M.; Broedl, U.C. Expression of Human Chemerin Induces Insulin Resistance in the Skeletal Muscle but Does Not Affect Weight, Lipid Levels, and Atherosclerosis in LDL Receptor Knockout Mice on High-Fat Diet. Diabetes 2010, 59, 2898–2903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- da Silva, G.N.; Amato, A.A. Thermogenic Adipose Tissue Aging: Mechanisms and Implications. Front. Cell. Dev. Biol. 2022, 10, 955612. [Google Scholar] [CrossRef]
- de Oliveira dos Santos, A.R.; de Oliveira Zanuso, B.; Miola, V.F.B.; Barbalho, S.M.; Santos Bueno, P.C.; Flato, U.A.P.; Detregiachi, C.R.P.; Buchaim, D.V.; Buchaim, R.L.; Tofano, R.J.; et al. Adipokines, Myokines, and Hepatokines: Crosstalk and Metabolic Repercussions. Int. J. Mol. Sci. 2021, 22, 2639. [Google Scholar] [CrossRef]
- Haines, M.S.; Dichtel, L.E.; Kimball, A.; Bollinger, B.; Gerweck, A.V.; Bredella, M.A.; Miller, K.K. OR26-03 Lower Serum Myostatin Levels Are Associated with Higher Insulin Sensitivity in Adults with Overweight/Obesity. J. Endocr. Soc. 2020, 4, OR26-03. [Google Scholar] [CrossRef]
- Amor, M.; Itariu, B.K.; Moreno-Viedma, V.; Keindl, M.; Jürets, A.; Prager, G.; Langer, F.; Grablowitz, V.; Zeyda, M.; Stulnig, T.M. Serum Myostatin Is Upregulated in Obesity and Correlates with Insulin Resistance in Humans. Exp. Clin. Endocrinol. Diabetes 2019, 127, 550–556. [Google Scholar] [CrossRef]
- Hittel, D.S.; Berggren, J.R.; Shearer, J.; Boyle, K.; Houmard, J.A. Increased Secretion and Expression of Myostatin in Skeletal Muscle From Extremely Obese Women. Diabetes 2009, 58, 30–38. [Google Scholar] [CrossRef] [Green Version]
- Hjorth, M.; Pourteymour, S.; Görgens, S.W.; Langleite, T.M.; Lee, S.; Holen, T.; Gulseth, H.L.; Birkeland, K.I.; Jensen, J.; Drevon, C.A.; et al. Myostatin in Relation to Physical Activity and Dysglycaemia and Its Effect on Energy Metabolism in Human Skeletal Muscle Cells. Acta Physiol. 2016, 217, 45–60. [Google Scholar] [CrossRef]
- Lin, J.; Arnold, H.B.; Della-Fera, M.A.; Azain, M.J.; Hartzell, D.L.; Baile, C.A. Myostatin Knockout in Mice Increases Myogenesis and Decreases Adipogenesis. Biochem. Biophys. Res. Commun. 2002, 291, 701–706. [Google Scholar] [CrossRef] [PubMed]
- CONSITT, L.A.; CLARK, B.C. The vicious cycle of myostatin signaling in sarcopenic obesity: Myostatin role in skeletal muscle growth, insulin signaling and implications for clinical trials. J. Frailty Aging 2017, 7, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Shan, T.; Liang, X.; Bi, P.; Kuang, S. Myostatin Knockout Drives Browning of White Adipose Tissue through Activating the AMPK-PGC1α-Fndc5 Pathway in Muscle. FASEB J. 2013, 27, 1981–1989. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nielsen, A.R.; Mounier, R.; Plomgaard, P.; Mortensen, O.H.; Penkowa, M.; Speerschneider, T.; Pilegaard, H.; Pedersen, B.K. Expression of Interleukin-15 in Human Skeletal Muscle Effect of Exercise and Muscle Fibre Type Composition. J. Physiol. 2007, 584, 305–312. [Google Scholar] [CrossRef] [PubMed]
- Quinn, L.S.; Anderson, B.G.; Conner, J.D.; Wolden-Hanson, T. IL-15 Overexpression Promotes Endurance, Oxidative Energy Metabolism, and Muscle PPARδ, SIRT1, PGC-1α, and PGC-1β Expression in Male Mice. Endocrinology 2013, 154, 232–245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Connell, G.C.; Pistilli, E.E. Interleukin-15 Directly Stimulates pro-Oxidative Gene Expression in Skeletal Muscle in-Vitro via a Mechanism That Requires Interleukin-15 Receptor Alpha. Biochem. Biophys. Res. Commun. 2015, 458, 614–619. [Google Scholar] [CrossRef]
- Sun, H.; Liu, D. Hydrodynamic Delivery of Interleukin 15 Gene Promotes Resistance to High Fat Diet-Induced Obesity, Fatty Liver and Improves Glucose Homeostasis. Gene. Ther. 2015, 22, 341–347. [Google Scholar] [CrossRef] [Green Version]
- Carbó, N.; López-Soriano, J.; Costelli, P.; Alvarez, B.; Busquets, S.; Baccino, F.M.; Quinn, L.S.; López-Soriano, F.J.; Argilés, J.M. Interleukin-15 Mediates Reciprocal Regulation of Adipose and Muscle Mass: A Potential Role in Body Weight Control. Biochim. Biophys. Acta 2001, 1526, 17–24. [Google Scholar] [CrossRef]
- Nielsen, A.R.; Hojman, P.; Erikstrup, C.; Fischer, C.P.; Plomgaard, P.; Mounier, R.; Mortensen, O.H.; Broholm, C.; Taudorf, S.; Krogh-Madsen, R.; et al. Association between Interleukin-15 and Obesity: Interleukin-15 as a Potential Regulator of Fat Mass. J. Clin. Endocrinol. Metab. 2008, 93, 4486–4493. [Google Scholar] [CrossRef] [Green Version]
- Quinn, L.S.; Anderson, B.G.; Strait-Bodey, L.; Stroud, A.M.; Argilés, J.M. Oversecretion of Interleukin-15 from Skeletal Muscle Reduces Adiposity. Am. J. Physiol. Endocrinol. Metab. 2009, 296, E191–E202. [Google Scholar] [CrossRef] [Green Version]
- Alvarez, B.; Carbó, N.; López-Soriano, J.; Drivdahl, R.H.; Busquets, S.; López-Soriano, F.J.; Argilés, J.M.; Quinn, L.S. Effects of Interleukin-15 (IL-15) on Adipose Tissue Mass in Rodent Obesity Models: Evidence for Direct IL-15 Action on Adipose Tissue. Biochim. Et Biophys. Acta (BBA) Gen. Subj. 2002, 1570, 33–37. [Google Scholar] [CrossRef]
- Iannone, F.; Montesanto, A.; Cione, E.; Crocco, P.; Caroleo, M.C.; Dato, S.; Rose, G.; Passarino, G. Expression Patterns of Muscle-Specific MiR-133b and MiR-206 Correlate with Nutritional Status and Sarcopenia. Nutrients 2020, 12, 297. [Google Scholar] [CrossRef] [PubMed]
- Nachtigall, P.G.; Dias, M.C.; Carvalho, R.F.; Martins, C.; Pinhal, D. MicroRNA-499 Expression Distinctively Correlates to Target Genes Sox6 and Rod1 Profiles to Resolve the Skeletal Muscle Phenotype in Nile Tilapia. PLoS ONE 2015, 10, e0119804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, T.; Zhang, Y.; Liu, Y.; Zhu, D.; Yu, J.; Li, G.; Sun, Z.; Wang, W.; Jiang, H.; Hong, Z. MiR-27a Promotes Insulin Resistance and Mediates Glucose Metabolism by Targeting PPAR-γ-Mediated PI3K/AKT Signaling. Aging 2019, 11, 7510–7524. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Qiu, J.; Liu, H.; Deng, Y.; Hu, S.; Hu, J.; Wang, Y.; Wang, J. MicroRNA-33a Negatively Regulates Myoblast Proliferation by Targeting IGF1, Follistatin and Cyclin D1. Biosci. Rep. 2020, 40, BSR20191327. [Google Scholar] [CrossRef] [PubMed]
- Chen, N.; Li, Q.; Liu, J.; Jia, S. Irisin, an Exercise-Induced Myokine as a Metabolic Regulator: An Updated Narrative Review. Diabetes Metab. Res. Rev. 2016, 32, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Huh, J.Y.; Dincer, F.; Mesfum, E.; Mantzoros, C.S. Irisin Stimulates Muscle Growth-Related Genes and Regulates Adipocyte Differentiation and Metabolism in Humans. Int. J. Obes. 2014, 38, 1538–1544. [Google Scholar] [CrossRef]
- Barra, N.G.; Palanivel, R.; Denou, E.; Chew, M.V.; Gillgrass, A.; Walker, T.D.; Kong, J.; Richards, C.D.; Jordana, M.; Collins, S.M.; et al. Interleukin-15 Modulates Adipose Tissue by Altering Mitochondrial Mass and Activity. PLoS ONE 2014, 9, e114799. [Google Scholar] [CrossRef]
- Wang, M.; Yu, H.; Kim, Y.S.; Bidwell, C.A.; Kuang, S. Myostatin Facilitates Slow and Inhibits Fast Myosin Heavy Chain Expression during Myogenic Differentiation. Biochem. Biophys. Res. Commun. 2012, 426, 83–88. [Google Scholar] [CrossRef] [Green Version]
- Cleasby, M.E.; Jarmin, S.; Eilers, W.; Elashry, M.; Andersen, D.K.; Dickson, G.; Foster, K. Local Overexpression of the Myostatin Propeptide Increases Glucose Transporter Expression and Enhances Skeletal Muscle Glucose Disposal. Am. J. Physiol. Endocrinol. Metab. 2014, 306, E814–E823. [Google Scholar] [CrossRef]
- Kersten, S.; Lichtenstein, L.; Steenbergen, E.; Mudde, K.; Hendriks, H.F.J.; Hesselink, M.K.; Schrauwen, P.; Müller, M. Caloric Restriction and Exercise Increase Plasma ANGPTL4 Levels in Humans via Elevated Free Fatty Acids. Arterioscler. Thromb. Vasc. Biol. 2009, 29, 969–974. [Google Scholar] [CrossRef] [PubMed]
- Hojman, P.; Pedersen, M.; Nielsen, A.R.; Krogh-Madsen, R.; Yfanti, C.; Akerstrom, T.; Nielsen, S.; Pedersen, B.K. Fibroblast Growth Factor-21 Is Induced in Human Skeletal Muscles by Hyperinsulinemia. Diabetes 2009, 58, 2797–2801. [Google Scholar] [CrossRef] [PubMed]
- Lee, P.; Linderman, J.D.; Smith, S.; Brychta, R.J.; Wang, J.; Idelson, C.; Perron, R.M.; Werner, C.D.; Phan, G.Q.; Kammula, U.S.; et al. Irisin and FGF21 Are Cold-Induced Endocrine Activators of Brown Fat Function in Humans. Cell Metab. 2014, 19, 302–309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ouchi, N.; Oshima, Y.; Ohashi, K.; Higuchi, A.; Ikegami, C.; Izumiya, Y.; Walsh, K. Follistatin-like 1, a Secreted Muscle Protein, Promotes Endothelial Cell Function and Revascularization in Ischemic Tissue through a Nitric-Oxide Synthase-Dependent Mechanism. J. Biol. Chem. 2008, 283, 32802–32811. [Google Scholar] [CrossRef] [Green Version]
- Görgens, S.W.; Raschke, S.; Holven, K.B.; Jensen, J.; Eckardt, K.; Eckel, J. Regulation of Follistatin-like Protein 1 Expression and Secretion in Primary Human Skeletal Muscle Cells. Arch. Physiol. Biochem. 2013, 119, 75–80. [Google Scholar] [CrossRef]
- Kim, C.-S.; Park, H.-S.; Kawada, T.; Kim, J.-H.; Lim, D.; Hubbard, N.E.; Kwon, B.-S.; Erickson, K.L.; Yu, R. Circulating Levels of MCP-1 and IL-8 Are Elevated in Human Obese Subjects and Associated with Obesity-Related Parameters. Int. J. Obes. 2006, 30, 1347–1355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sell, H.; Dietze-Schroeder, D.; Kaiser, U.; Eckel, J. Monocyte Chemotactic Protein-1 Is a Potential Player in the Negative Cross-Talk between Adipose Tissue and Skeletal Muscle. Endocrinology 2006, 147, 2458–2467. [Google Scholar] [CrossRef] [Green Version]
- Sabater, M.; Moreno-Navarrete, J.M.; José Ortega, F.; Pardo, G.; Salvador, J.; Ricart, W.; Frühbeck, G.; Fernández-Real, J.M. Circulating Pigment Epithelium-Derived Factor Levels Are Associated with Insulin Resistance and Decrease after Weight Loss. J. Clin. Endocrinol. Metab. 2010, 95, 4720–4728. [Google Scholar] [CrossRef]
- Famulla, S.; Lamers, D.; Hartwig, S.; Passlack, W.; Horrighs, A.; Cramer, A.; Lehr, S.; Sell, H.; Eckel, J. Pigment Epithelium-Derived Factor (PEDF) Is One of the Most Abundant Proteins Secreted by Human Adipocytes and Induces Insulin Resistance and Inflammatory Signaling in Muscle and Fat Cells. Int. J. Obes. 2011, 35, 762–772. [Google Scholar] [CrossRef] [Green Version]
- Atkins, J.L.; Wannamathee, S.G. Sarcopenic Obesity in Ageing: Cardiovascular Outcomes and Mortality. Br. J. Nutr. 2020, 124, 1102–1113. [Google Scholar] [CrossRef]
- Chow, L.S.; Gerszten, R.E.; Taylor, J.M.; Pedersen, B.K.; van Praag, H.; Trappe, S.; Febbraio, M.A.; Galis, Z.S.; Gao, Y.; Haus, J.M.; et al. Exerkines in Health, Resilience and Disease. Nat. Rev. Endocrinol. 2022, 18, 273–289. [Google Scholar] [CrossRef] [PubMed]
- Laurens, C.; Bergouignan, A.; Moro, C. Exercise-Released Myokines in the Control of Energy Metabolism. Front. Physiol. 2020, 11, 91. [Google Scholar] [CrossRef] [PubMed]
- Zunner, B.E.M.; Wachsmuth, N.B.; Eckstein, M.L.; Scherl, L.; Schierbauer, J.R.; Haupt, S.; Stumpf, C.; Reusch, L.; Moser, O. Myokines and Resistance Training: A Narrative Review. Int. J. Mol. Sci. 2022, 23, 3501. [Google Scholar] [CrossRef] [PubMed]
- Senesi, P.; Luzi, L.; Terruzzi, I. Adipokines, Myokines, and Cardiokines: The Role of Nutritional Interventions. Int. J. Mol. Sci. 2020, 21, 8372. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zamboni, M.; Mazzali, G.; Brunelli, A.; Saatchi, T.; Urbani, S.; Giani, A.; Rossi, A.P.; Zoico, E.; Fantin, F. The Role of Crosstalk between Adipose Cells and Myocytes in the Pathogenesis of Sarcopenic Obesity in the Elderly. Cells 2022, 11, 3361. https://doi.org/10.3390/cells11213361
Zamboni M, Mazzali G, Brunelli A, Saatchi T, Urbani S, Giani A, Rossi AP, Zoico E, Fantin F. The Role of Crosstalk between Adipose Cells and Myocytes in the Pathogenesis of Sarcopenic Obesity in the Elderly. Cells. 2022; 11(21):3361. https://doi.org/10.3390/cells11213361
Chicago/Turabian StyleZamboni, Mauro, Gloria Mazzali, Anna Brunelli, Tanaz Saatchi, Silvia Urbani, Anna Giani, Andrea P. Rossi, Elena Zoico, and Francesco Fantin. 2022. "The Role of Crosstalk between Adipose Cells and Myocytes in the Pathogenesis of Sarcopenic Obesity in the Elderly" Cells 11, no. 21: 3361. https://doi.org/10.3390/cells11213361