

Metabolic Advantage of 25(OH)D3 versus 1,25(OH)2D3 Supplementation in Infantile Nephropathic Cystinosis-Associated Adipose Tissue Browning and Muscle Wasting
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design
2.2. Measuremnt of Lean and Fat Mass
2.3. Resting Metabolic Rate
2.4. Mouse Muscle Function
2.5. Serum and Blood Chemistry
2.6. Protein Assay for Muscle and Adipose Tissue
2.7. Fiber Size and Fatty Infiltration of Gastrocnemius
2.8. Muscle Cystine Content Measurement
2.9. Muscle RNAseq Analysis
2.10. Quantitative Real-Time PCR
2.11. Statistics
3. Results
3.1. Supplementation of 25(OH)D3 or 1,25(OH)2D3 Replenishes Serum 25(OH)D3 or 1,25(OH)2D3 Concentration in Ctns−/− Mice
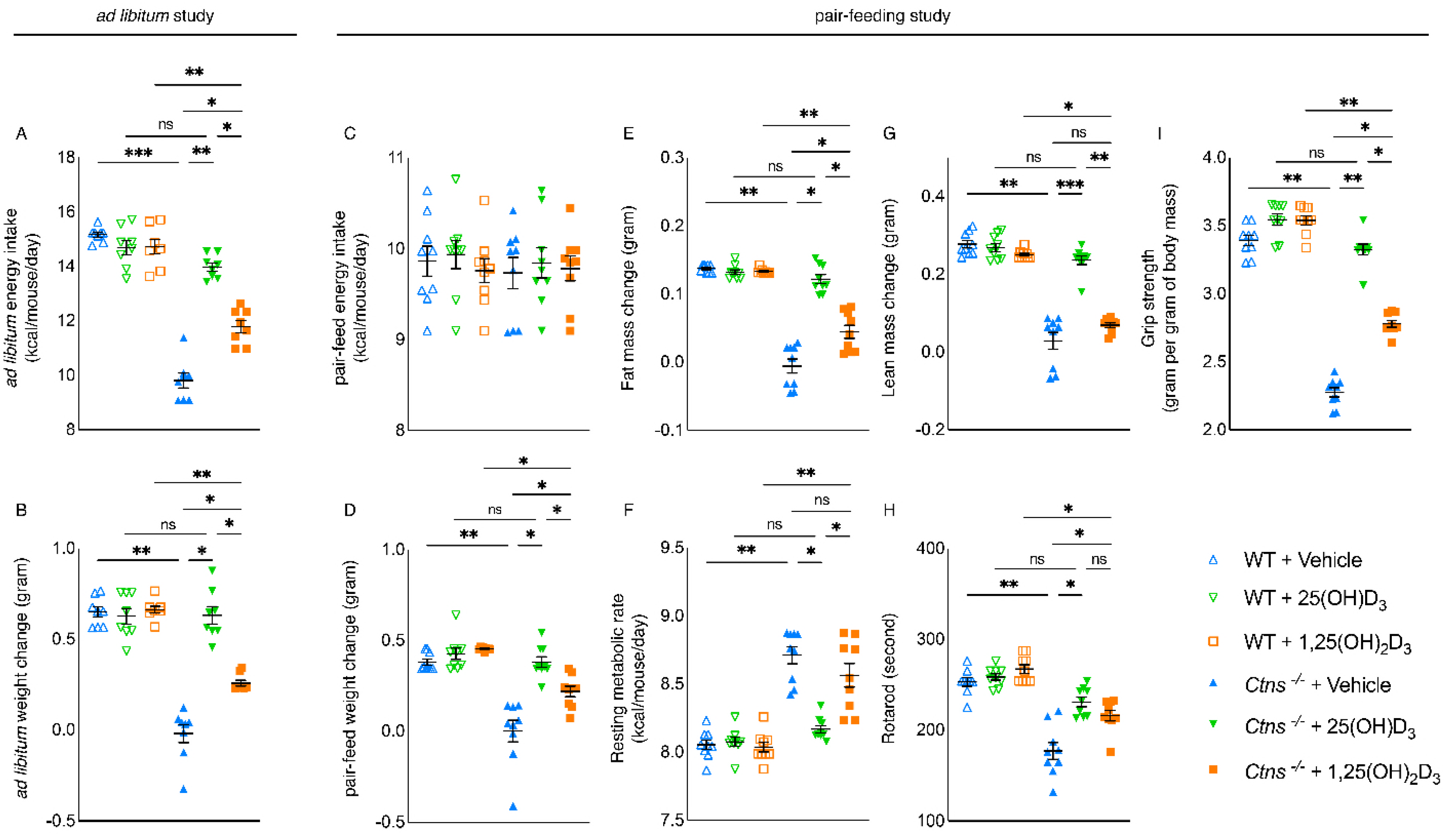
3.2. Repletion of 25-Hydroxyvitamin D3 Normalizes Caloric Intake and Weight Gain in Ctns−/− Mice
3.3. Repletion of 25-Hydroxyvitamin D3 Improves Energy Homeostasis in Ctns−/− Mice
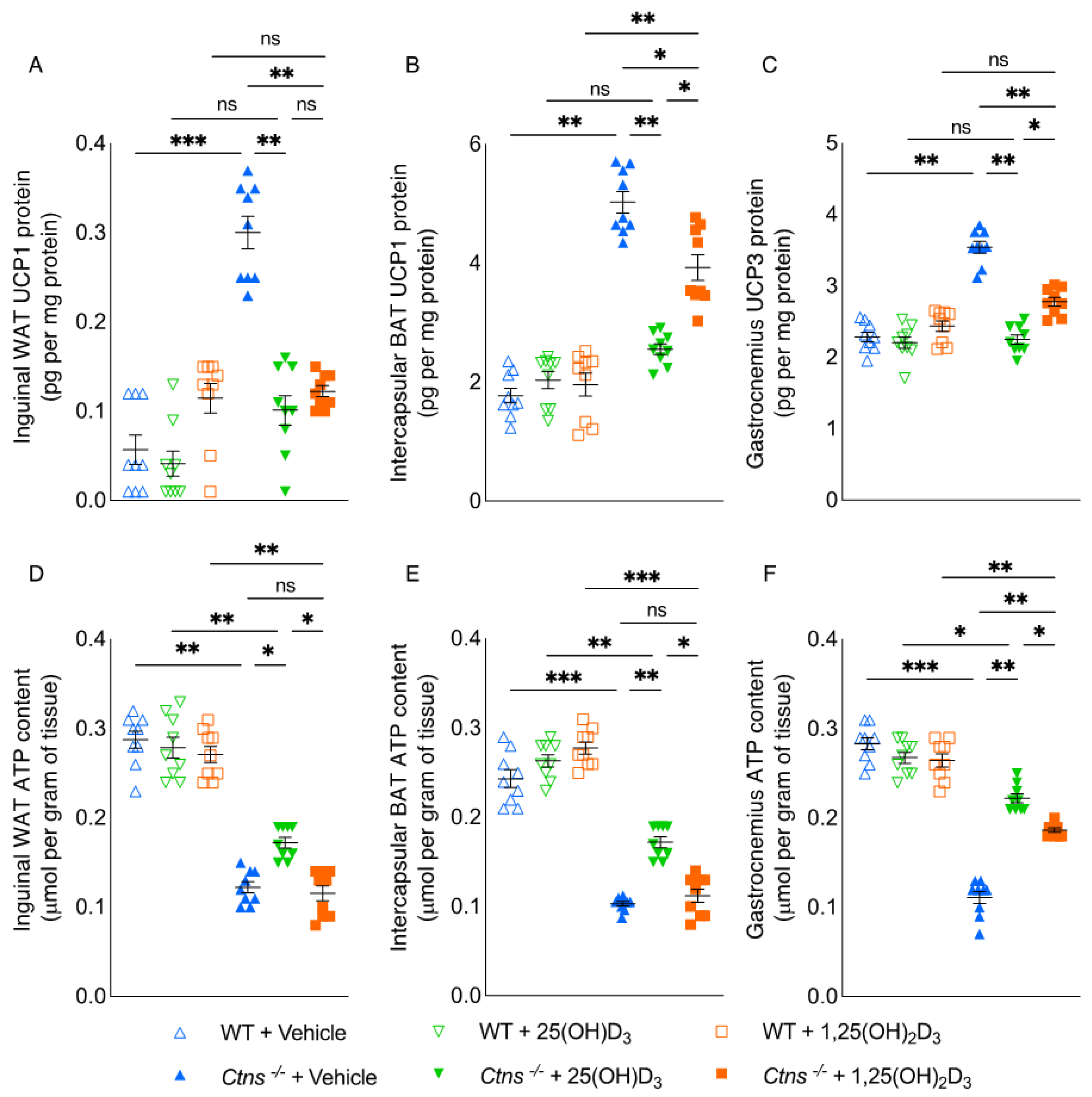
3.4. Repletion of 25-Hydroxyvitamin D3 Improves Adipose Tissue and Skeletal Muscle Energy Homeostasis in Ctns−/− Mice
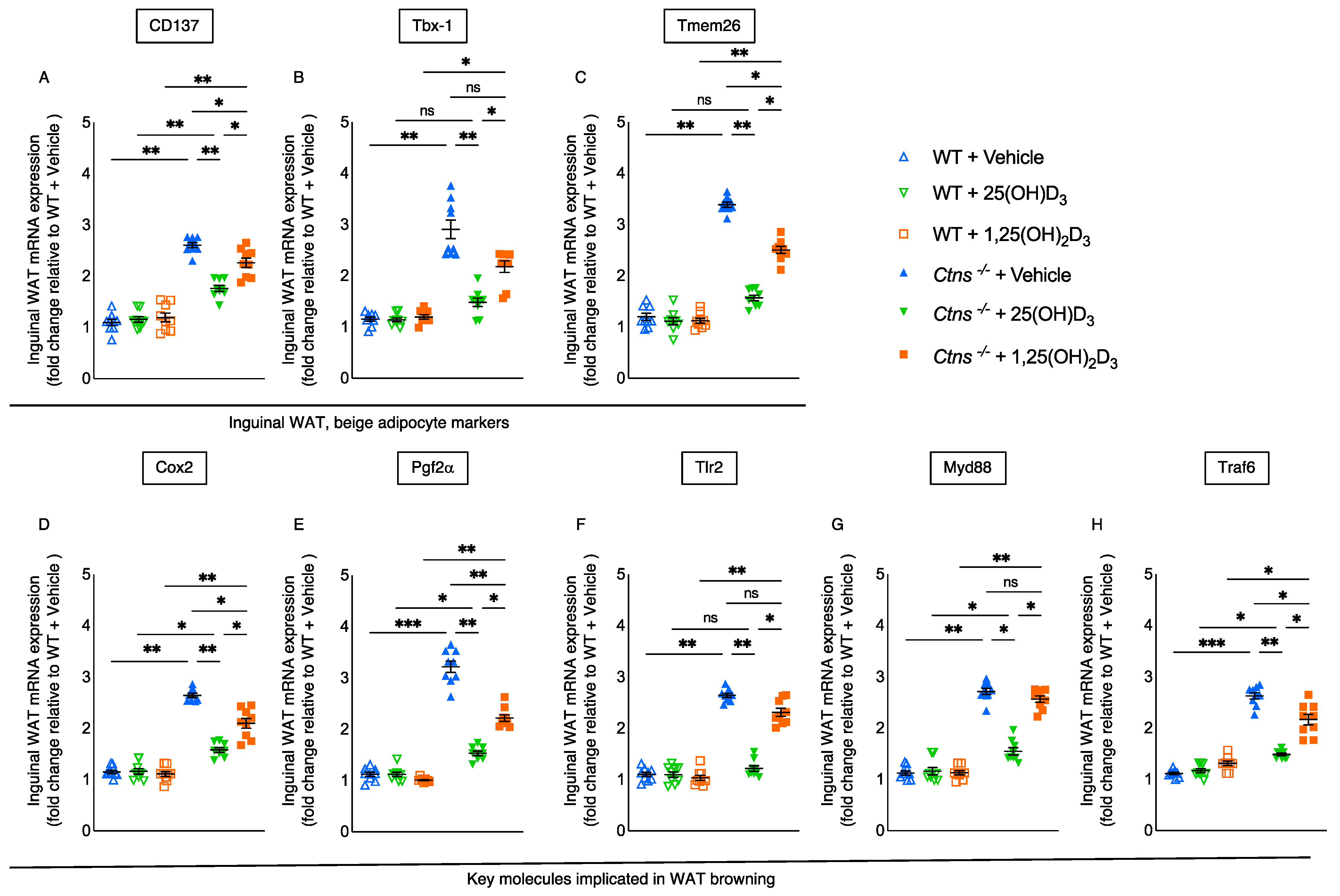
3.5. Repletion of 25-Hydroxyvitamin D3 Mitigates White Adipose Tissue Browning in Ctns−/− Mice
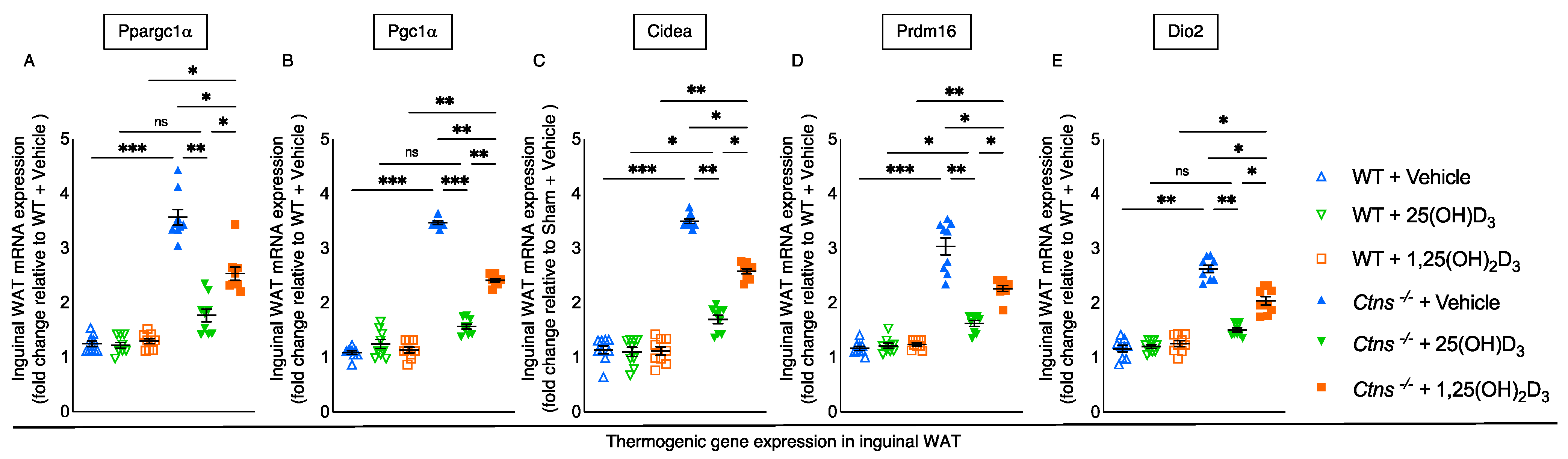
3.6. Repletion of 25-Hydroxyvitamin D3 Decreases WAT Thermogenic Gene Expression in Ctns−/− Mice
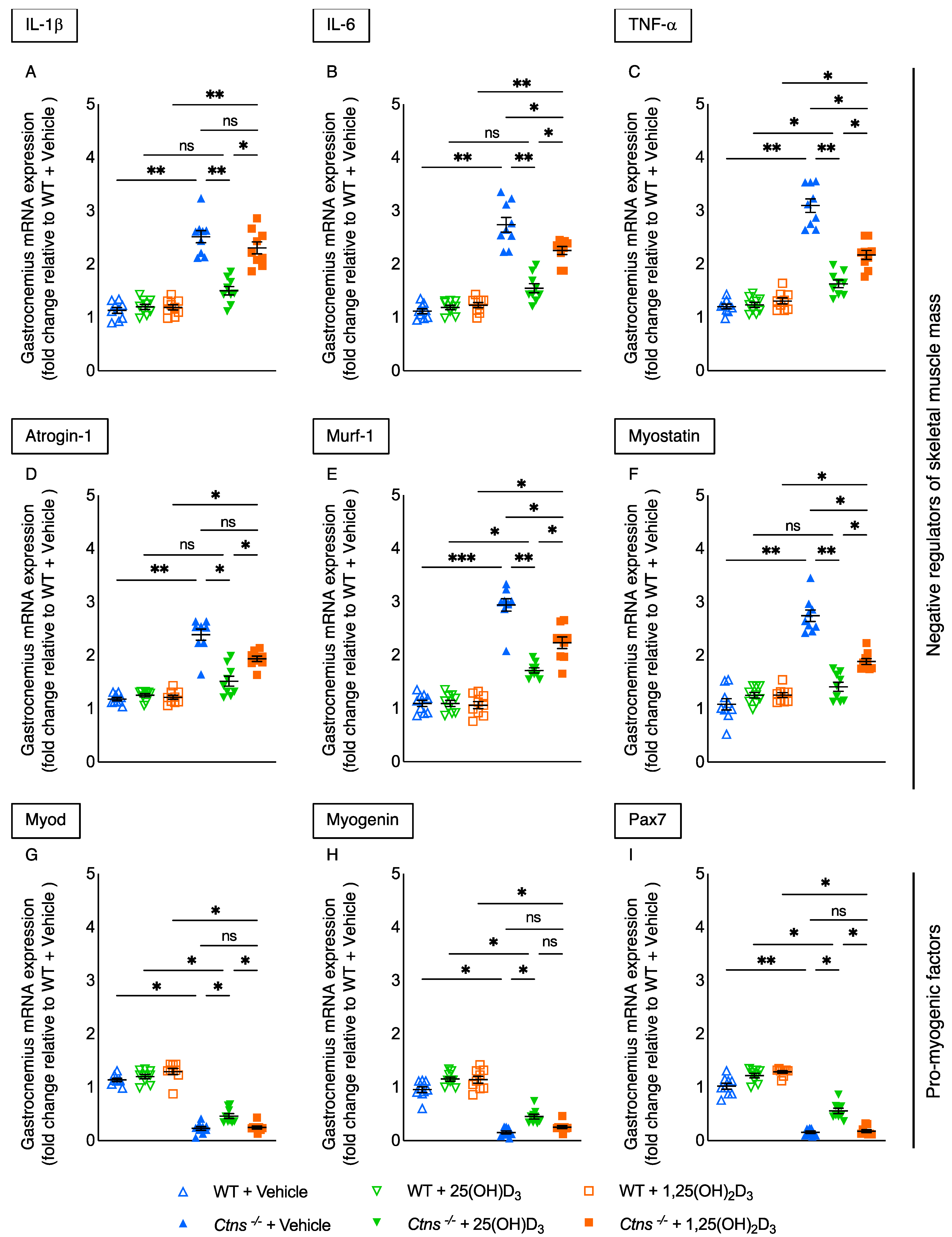
3.7. Repletion of 25-Hydroxyvitamin D3 Ameliorates Muscle Wasting Signaling Pathways in Ctns−/− Mice
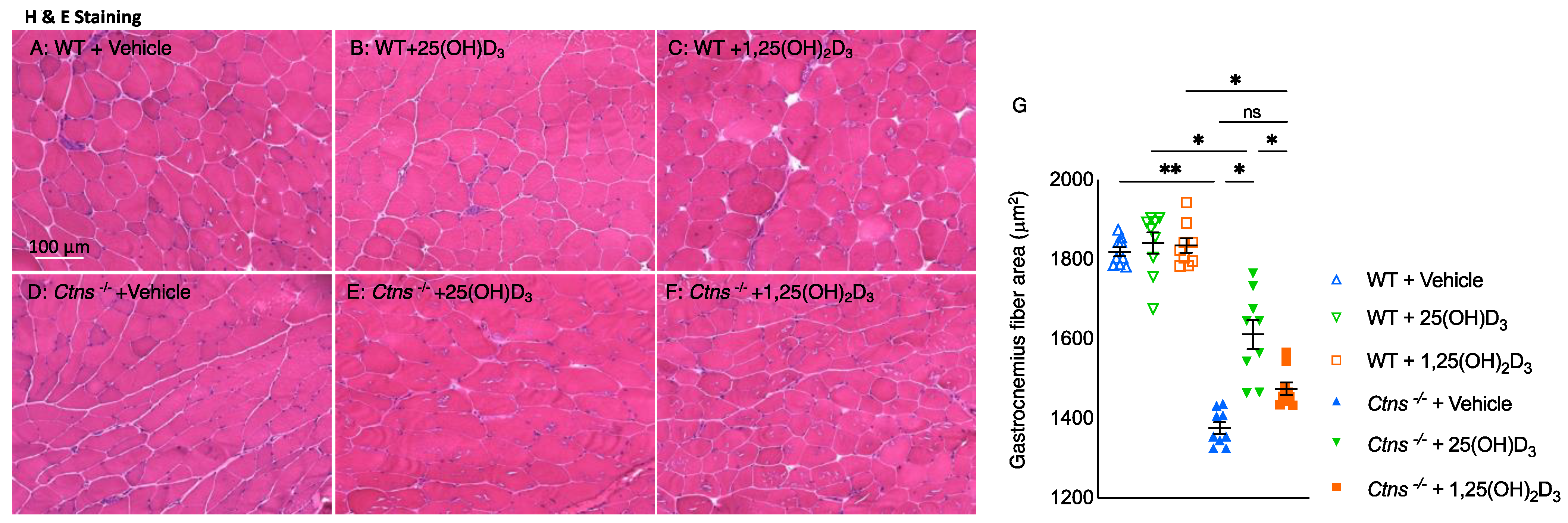
3.8. Repletion of 25-Hydroxyvitamin D3 Increases Muscle Fiber Size in Ctns−/− Mice
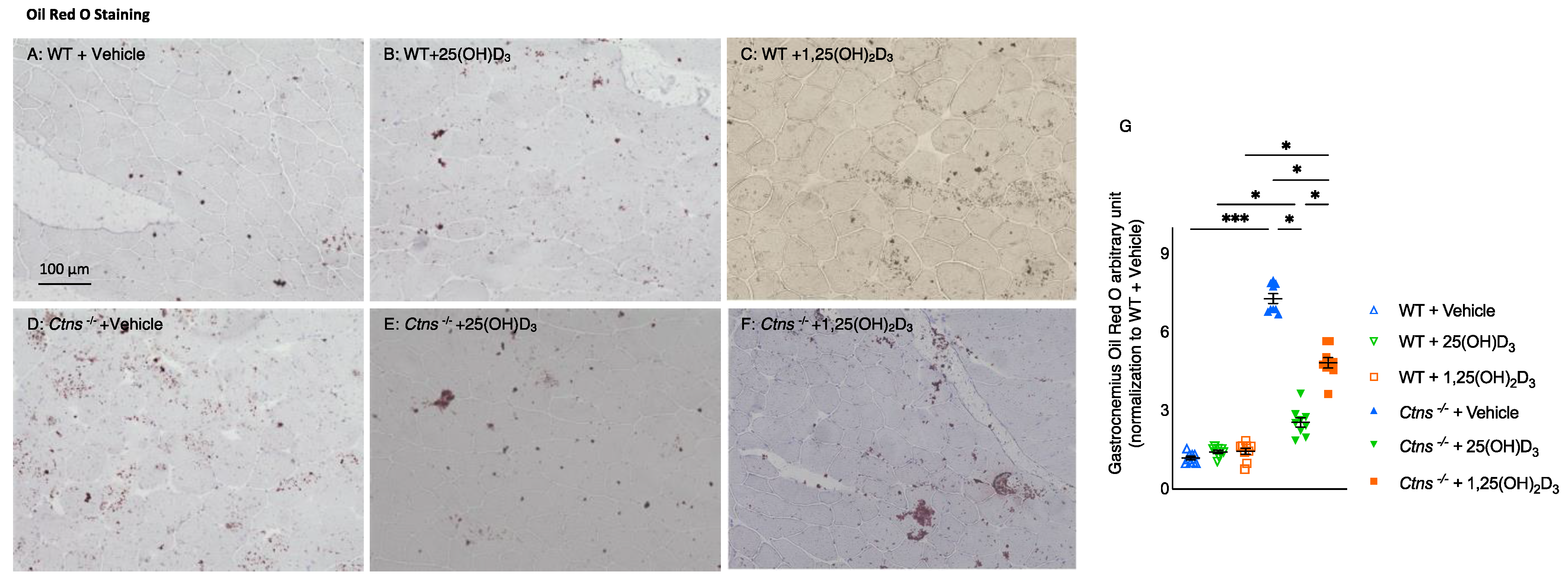
3.9. Repletion of 25-Hydroxyvitamin D3 Decreases Muscle Fat Infiltration in Ctns−/− Mice
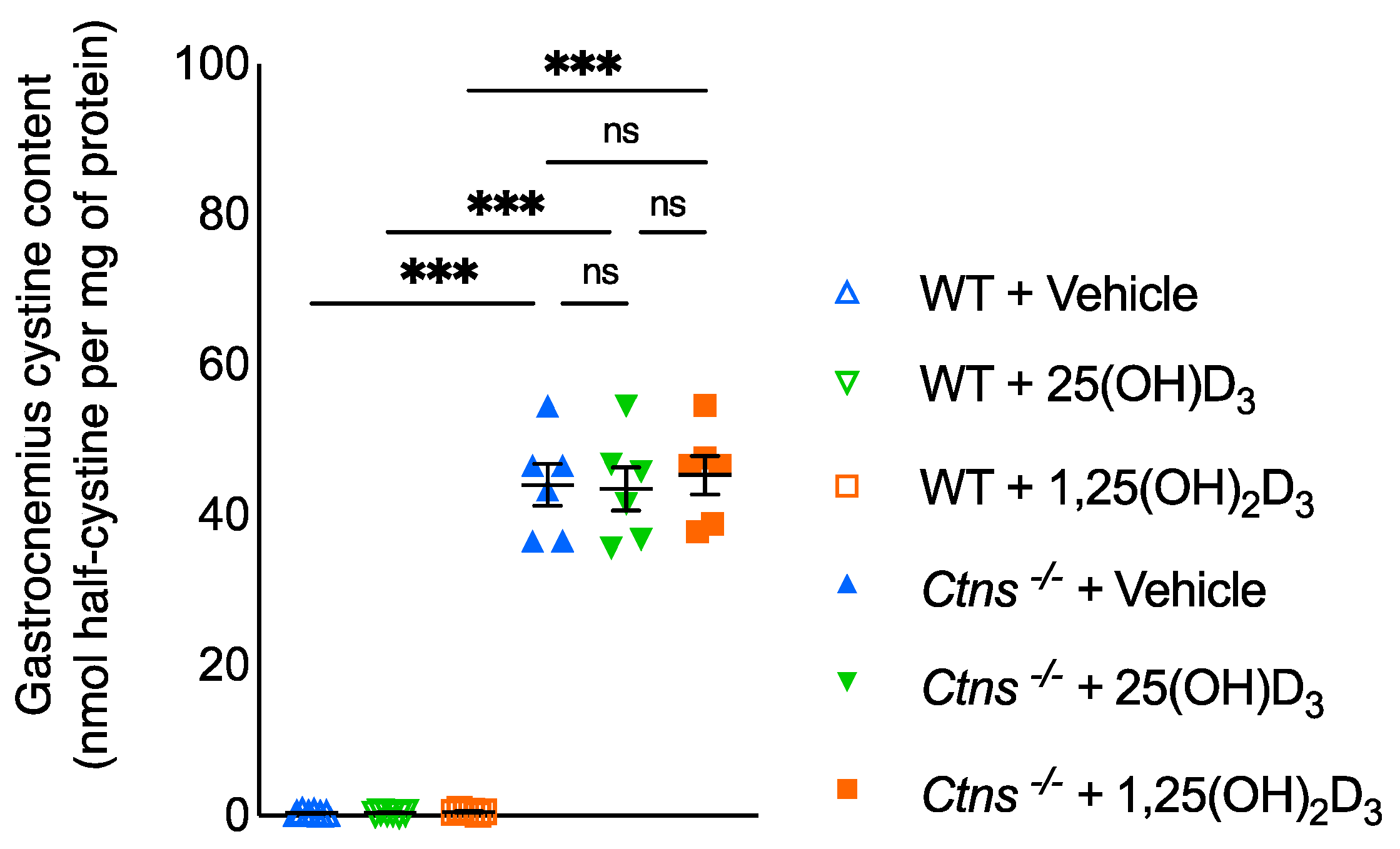
3.10. Muscle Content of Cystine in Ctns−/− Mice
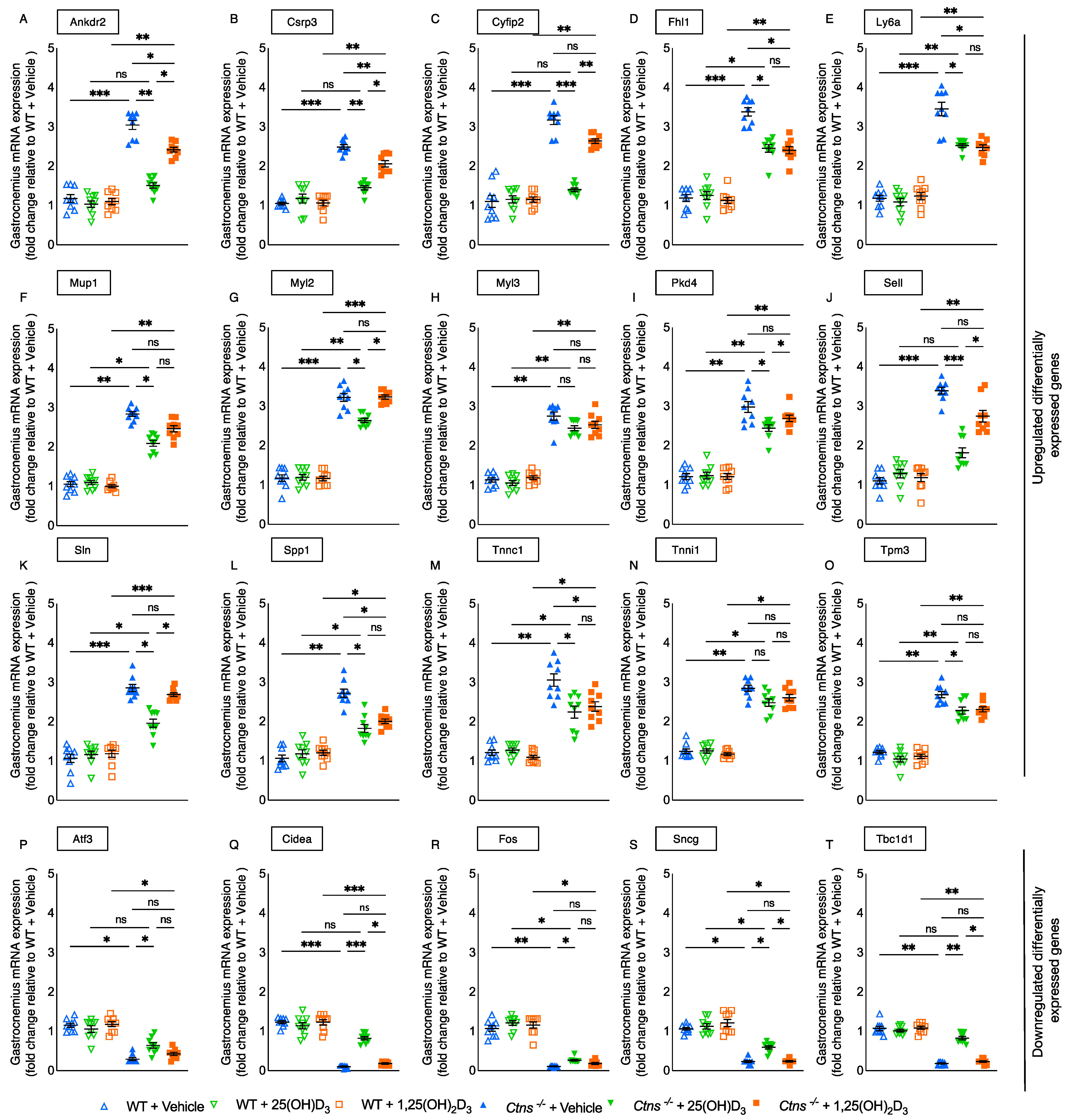
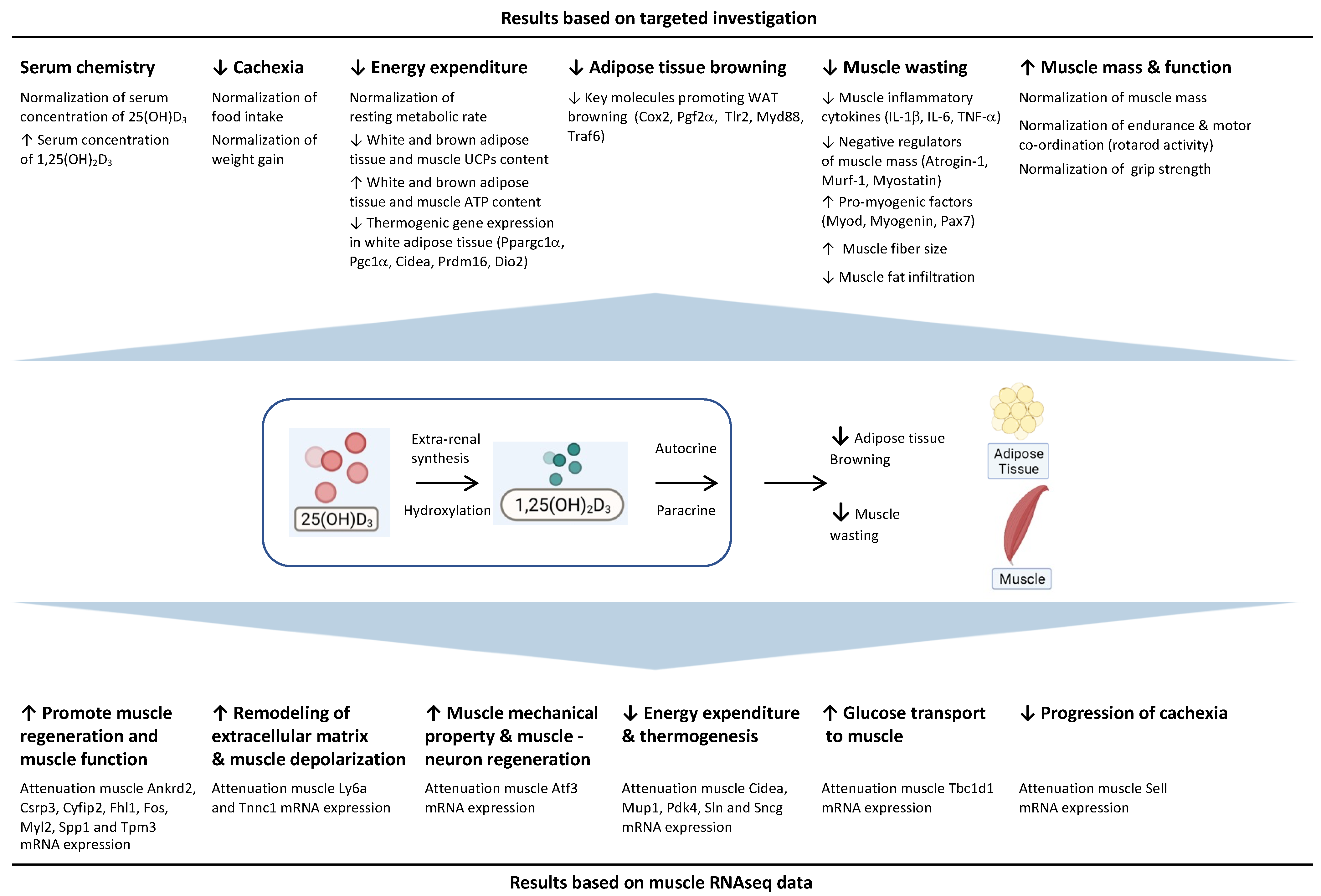
3.11. Molecular Mechanism of 25-Hydroxyvitamin D3 Repletion by RNAseq Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Town, M.M.; Jean, G.; Cherqui, S.; Attard, M.; Forestier, L.; Whitmore, S.A.; Callen, D.F.; Gribouval, O.; Broyer, M.; Bates, G.; et al. A novel gene encoding an integral membrane protein is mutated in nephropathic cystinosis. Nat. Genet. 1998, 18, 319–324. [Google Scholar] [CrossRef]
- Gahl, W.A.; Thoene, J.G.; Schneider, J.A. Cystinosis. N. Engl. J. Med. 2002, 347, 111–121. [Google Scholar] [CrossRef]
- Nesterova, G.; Gahl, W. Nephropathic cystinosis: Late complications of a multisystemic disease. Pediatr. Nephrol. 2007, 23, 863–878. [Google Scholar] [CrossRef]
- Theodoropoulos, D.S.; Krasnewich, D.; Kaiser-Kupfer, M.I.; Gahl, W.A. Classic Nephropathic Cystinosis as an Adult Disease. JAMA 1993, 270, 2200–2204. [Google Scholar] [CrossRef]
- Cheung, W.W.; Cherqui, S.; Ding, W.; Esparza, M.; Zhou, P.; Shao, J.; Lieber, R.; Mak, R.H. Muscle wasting and adipose tissue browning in infantile nephropathic cystinosis. J. Cachexia Sarcopenia Muscle 2015, 7, 152–164. [Google Scholar] [CrossRef]
- Fenzl, A.; Kiefer, F.W. Brown adipose tissue and thermogenesis. Horm. Mol. Biol. Clin. Investig. 2014, 19, 25–37. [Google Scholar] [CrossRef]
- Petruzzelli, M.; Schweiger, M.; Schreiber, R.; Campos-Olivas, R.; Tsoli, M.; Allen, J.; Swarbrick, M.; Rose-John, S.; Rincon, M.; Robertson, G.; et al. A Switch from White to Brown Fat Increases Energy Expenditure in Cancer-Associated Cachexia. Cell Metab. 2014, 20, 433–447. [Google Scholar] [CrossRef] [Green Version]
- A Vaitkus, J.; Celi, F.S. The role of adipose tissue in cancer-associated cachexia. Exp. Biol. Med. 2016, 242, 473–481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elattar, S.; Dimri, M.; Satyanarayana, A. The tumor secretory factor ZAG promotes white adipose tissue browning and energy wasting. FASEB J. 2018, 32, 4727–4743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, Y.; Liu, R.-X.; Zhu, M.-T.; Shen, W.-B.; Xie, J.; Zhang, Z.-Y.; Chen, N.; Shan, C.; Guo, X.-Z.; Lu, Y.-D.; et al. The browning of white adipose tissue and body weight loss in primary hyperparathyroidism. EBioMedicine 2019, 40, 56–66. [Google Scholar] [CrossRef] [PubMed]
- Kong, X.; Yao, T.; Zhou, P.; Kazak, L.; Tenen, D.; Lyubetskaya, A.; Dawes, B.A.; Tsai, L.; Kahn, B.B.; Spiegelman, B.M.; et al. Brown adipose tissue controls skeletal muscle function via the secretion of myostatin. Cell Metab. 2018, 28, 631–643.e3. [Google Scholar] [CrossRef] [Green Version]
- Kir, S.; White, J.P.; Kleiner, S.; Kazak, L.; Cohen, P.; Baracos, V.E.; Spiegelman, B.M. Tumour-derived PTH-related protein triggers adipose tissue browning and cancer cachexia. Nature 2014, 513, 100–104. [Google Scholar] [CrossRef] [Green Version]
- Kir, S.; Komaba, H.; Garcian, A.P.; Economopoulos, K.P.; Liu, W.; Lanske, B.; Hodin, R.A.; Spiegelman, B.M. PTH/PTHrP receptor mediates cachexia in models of kidney failure and cancer. Cell Metab. 2015, 23, 315–323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Autier, P.; Boniol, M.; Pizot, C.; Mullie, P. Vitamin D status and ill health: A systematic review. Lancet Diabetes Endocrinol. 2014, 2, 76–89. [Google Scholar] [CrossRef]
- Park, J.E.; Tirupathi Pichiah, P.B.; Cha, Y.-S. Vitamin D and Metabolic Diseases: Growing Roles of Vitamin D. J. Obes. Metab. Syndr. 2018, 27, 223–232. [Google Scholar] [CrossRef] [Green Version]
- Katzir, Z.; Shivil, Y.; Landau, H.; Kidrony, G.; Popovtzer, M.M. Nephrogenic diabetes insipidus, cystinosis, and vitamin D. Arch. Dis. Child. 1988, 63, 548–550. [Google Scholar] [CrossRef] [Green Version]
- Steinherz, R.; Chesney, R.W.; Schulman, J.D.; DeLuca, H.F.; Phelps, M. Circulating vitamin D metabolites in nephropathic cystinosis. J. Pediatr. 1983, 102, 592–594. [Google Scholar] [PubMed]
- Chesney, R.W.; Hamstra, J.; Mazess, R.B.; Rose, P.; DeLuca, H.F. Circulating vitamin D metabolites concentrations in childhood renal disease. Kidney Int. 1982, 21, 65–69. [Google Scholar] [CrossRef] [Green Version]
- Cheung, W.W.; Hao, S.; Wang, Z.; Ding, W.; Zheng, R.H.; Gonzalez, A.; Zhan, J.-Y.; Zhou, P.; Li, S.P.; Esparza, M.C.; et al. Vitamin D repletion ameliorates adipose tissue browning and muscle wasting in infantile nephropathic cystinosis-associated cachexia. J. Cachexia Sarcopenia Muscle 2020, 11, 120–134. [Google Scholar] [CrossRef] [Green Version]
- Querfeld, U.; Mak, R.H. Vitamin D deficiency and toxicity in chronic kidney disease: In search of the therapeutic window. Pediatr. Nephrol. 2010, 25, 2413–2430. [Google Scholar] [CrossRef] [PubMed]
- Dusso, A.S. Kidney disease and vitamin D levels: 25-hydroxyvitamin D, 1,25-dihydroxyvitamin D, and VDR activation. Kidney Int. Suppl. 2011, 1, 136–141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lou, Y.-R.; Laaksi, I.; Syvala, A.; Blauer, M.; Tammela, T.L.J.; YLikomi, T.; Tuohimaa, P. 25-hydroxyvitamin D3 is an active hormone in human primary prostatic stromal cells. FASEB J. 2004, 18, 332–334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, X.; Hawthorne, M.; Vaishnav, A.; St-Arnaud, R.; Mehta, R.G. 25-Hydroxyvitamin D3 is a natural chemopreventive agent against carcinogen induced precancerous lesions in mouse mammary gland organ culture. Breast Cancer Res. Treat. 2009, 113, 31–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lou, Y.-R.; Molnar, F.; Perakyla, M.; Qiao, S.; Kalueff, A.V.; St-Arnaud, R.; Carlberg, C.; Tuohimaa, P. 25-Hydroxyvitamin D(3) is an agonistic vitamin D receptor ligand. J. Steroid Biochem. Mol. Biol. 2010, 118, 162–170. [Google Scholar] [CrossRef] [PubMed]
- Ritter, C.S.; Armbrecht, H.J.; Slatopolsky, E.; Brown, A.J. 25-Hydroxyvitamin D(3) suppresses PTH synthesis and secretion by bovine parathyroid cells. Kidney Int. 2006, 70, 654–659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.L.; Ding, X.F.; Tong, J.; Li, B.Y. Partial rescue of the phenotype in 1alpha-hydroxylase gene knockout mice by vitamin D3 injection. Endocr. Res. 2011, 36, 101–108. [Google Scholar] [CrossRef] [PubMed]
- Tuohimaa, P.; Wang, J.-H.; Khan, S.; Kuuslahti, M.; Qian, K.; Manninen, T.; Auvinen, P.; Vihinen, M.; Lou, Y.-R. Gene expression profiles in human and mouse primary cells provide new insights into the differential actions of vitamin D3 metabolites. PLoS ONE 2013, 8, e75338. [Google Scholar] [CrossRef]
- Young, S.; Struys, E.; Wood, T. Quantification of creatine and guanidinoacetate using GC-MS and LC-MS/MS for the detection of cerebral creatine deficiency syndromes. Curr. Protoc. Hum. Genet. 2007, 54, 17-3. [Google Scholar] [CrossRef]
- Du bowitz, V.; Sewry, C.A.; Oldfors, A.; Lane, R.J.M. Muscle Biopsy: A Practical Approach; Elsevier Health Sciences: Amsterdam, The Netherlands, 2013; 572p. [Google Scholar]
- Mehlem, A.; Hagberg, C.; Muhl, L.; Eriksson, U.; Falkevall, A. Imaging of neutral lipids by oil red O for analyzing the metabolic status in health and disease. Nat. Protoc. 2013, 8, 1149–1154. [Google Scholar] [CrossRef] [Green Version]
- Cherqui, S.; Sevin, C.; Hamard, G.; Kaatzis, V.; Sich, M.; Pequignot, M.O.; Gogat, K.; Abitbol, M.; Broyer, M.; Gubler, M.-C.; et al. Intralysosomal cystine Accumulation in Mice Lacking Cystinosin, the Protein Defective in Cystinosis. Mol. Cell Biol. 2002, 22, 7622–7632. [Google Scholar] [CrossRef]
- Syres, K.; Harrison, F.; Tedlock, M.; Jester, J.-V.; Simpson, J.; Roy, S.; Salomon, D.R.; Cherqui, S. Successful treatment of the murine model of cystinosis using bone marrow cell transplantation. Blood 2009, 11, 2542–2552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, P. TRAF molecules in cell signaling and in human diseases. J. Mol. Signal. 2013, 8, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hassan-Smith, Z.; Jenkinson, C.; Smith, D.J.; Hernadez, I.; Morgan, S.A.; Crabtree, N.J.; Gittoes, N.J.; Keevil, B.G.; Stewart, P.M.; Hewison, M. 25-hydroxyvitamin D3 and 1,25-dihydroxyvitamin D3 exert distinct effects on human skeletal muscle function and gene expression. PLoS ONE 2017, 12, e0170665. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Atkins, G.J.; Anderson, P.H.; Findlay, D.M.; Welldon, K.J.; Vincent, C.; Zannettino, A.C.W.; O’Loughlin, P.D.; Morris, H.A. Metabolism of vitamin D3 in human osteoblasts: Evidence for autocrine and paracrine activities of 1 alpha,25-dihydroxyvitamin D3. Bone 2007, 40, 1517–1528. [Google Scholar] [CrossRef] [PubMed]
- Boullata, J.I. Vitamin D supplementation: A pharmacologic perspective. Curr. Opin. Clin. Nutr. Metab. Care 2010, 13, 677–684. [Google Scholar] [CrossRef] [PubMed]
- Askeno, A.; Saikatsu, S.; Kawane, T.; Horiuchi, N. Mouse vitamin D-24-hydroxylase: Molecular cloning, tissue distribution, and transcriptional regulation by 1alpha,25-dihydroxyvitamin D3. Endocrinology 1997, 138, 2233–2240. [Google Scholar] [CrossRef] [PubMed]
- Munetsuna, E.; Kawanami, R.; Nishikawa, M.; Ikeda, S.; Nakabayashi, S.; Yasuda, K.; Ohta, M.; Kamakura, M.; Ikushiro, S.; Sakaki, T. Anti-proliferative activity of 25-hydroxyvitamin D3 in human prostate cells. Mol. Cell. Endocrinol. 2014, 382, 960–970. [Google Scholar] [CrossRef] [PubMed]
- Horst, R.L.; Wovkulich, P.M.; Baggiolini, E.G.; Uskoković., M.R.; Engstrom, G.W.; Napoli, J.L. (23S)-1,23,25-Trihydroxyvitamin D3: Its biologic activity and role in 1 alpha,25-dihydroxyvitamin D3 26,23-lactone biosynthesis. Biochemistry 1984, 23, 3973–3979. [Google Scholar] [CrossRef]
- Ikizler, T.A.; Wingard, R.L.; Sun, M.; Harvell, J.; Parker, R.A.; Hakim, R.M. Increased energy expenditure in hemodialysis patients. J. Am. Soc. Nepohrol. 1996, 7, 2646–2653. [Google Scholar] [CrossRef]
- Wang, A.Y.-M.; Sea, M.M.-M.; Tang, N.; Sanderson, J.E.; Lui, S.-F.; Li, P.K.-T.; Woo, J. Resting energy expenditure and subsequent mortality risk in peritoneal dialysis patients. J. Am. Soc. Nephrol. 2004, 15, 3134–3143. [Google Scholar] [CrossRef]
- Rousset, S.; Alves-Guerra, M.-C.; Mozo, J.; Miroux, B.; Cassard-Doulcier, A.-M.; Bouillaud, F.; Ricquier, D. The biology of mitochondrial uncoupling proteins. Diabetes 2004, 53 (Suppl. 1), S130–S135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bing, C.; Brown, M.; King, P.; Collins, P.; Tisdsale, M.J.; Williams, G. Increased gene expression of brown fat uncoupling protein (UCP)1 and skeletal muscle UCP2 and UCP3 in MAC16-induced cancer cachexia. Cancer Res. 2000, 60, 2405–2410. [Google Scholar] [PubMed]
- Wong, K.E.; Szeto, F.L.; Zhang, W.; Ye, H.; Kong, J.; Zhang, Z.; Sun, X.J.; Li, Y.C. Involvement of the vitamin D receptor in energy metabolism: Regulation of uncoupling proteins. Am. J. Physiol. Endocrinol. Metab. 2009, 296, E820–E828. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamei, Y.; Kawada, T.; Kazuki, R.; Ono, T.; Kato, S.; Sugimoto, E. Vitamin D receptor gene expression is up-regulated by 1, 25-dihydroxyvitamin D3 in 3T3-L1 preadipocytes. Biochem. Biophys. Res. Commun. 1993, 193, 948–955. [Google Scholar] [CrossRef]
- Wang, P.-Q.; Pan, D.-X.; Hu, C.-Q.; Zhu, Y.-L.; Liu, X.-J. Vitamin D–vitamin D receptor system down-regulates expression of uncoupling proteins in brown adipocyte through interaction with hairless protein. Biosci. Rep. 2020, 40, BSR20194294. [Google Scholar] [CrossRef] [PubMed]
- Jonas, M.I.; Kurylowicz, A.; Bartoszewicz, Z.; Lisik, W.; Jonas, M.; Kozniewski, K.; Puzianowska-Kuznicka, M. Vitamin D receptor gene expression in adipose tissue of obese individuals is regulated by miRNA and correlates with the pro-inflammatory cytokine level. Int. J. Mol. Sci. 2019, 20, 5272. [Google Scholar] [CrossRef] [Green Version]
- Nimitphong, H.; Holick, M.F.; Fried, S.K.; Lee, M.-J. 25-hydroxyvitamin D3 and 1,25-dihydroxyvitamin D3 promote the differentiation of human subcutaneous preadipocytes. PLoS ONE 2012, 7, e52171. [Google Scholar] [CrossRef] [Green Version]
- Fan, Y.; Futawaka, K.; Komaya, R.; Fukuda, Y.; Hayashi, M.; Imamoto, M.; Miyawaki, T.; Kasahara, M.; Tagami, T.; Moriyama, K. Vitamin D3/VDR resists diet-induced obesity by modulating UCP3 expression in muscle. J. Biomed. Sci. 2016, 23, 56. [Google Scholar] [CrossRef] [Green Version]
- Mierzejewski, B.; Archacka, K.; Grabowska, I.; Florkowska, A.; Ciemerych, M.A.; Brzoska, E. Human and mouse skeletal muscle stem and progenitor cells in health and disease. Semin. Cell Devel. Biol. 2020, 104, 93–104. [Google Scholar] [CrossRef] [PubMed]
- Gahl, W.A.; Balog, J.Z.; Kleta, R. Nephropathic cystinosis in adults: Natural history and effects of oral cysteamine therapy. Ann. Intern. Med. 2007, 147, 242–250. [Google Scholar] [CrossRef]
- Brodin-Sartorius, A.; Tete, M.J.; Niaudet, P.; Antignac, C.; Guest, G.; Ottolenghi, C.; Charbit, M.; Moyse, D.; Legendre, C.; Lesavre, P.; et al. Cysteamine therapy delays the progression of nephropathic cystinosis in late adolescents and adults. Kidney Int. 2012, 81, 179–189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruivo, R.; Anne, C.; Sagne, C.; Gasnier, B. Molecular and cellular basis of lysosomal transmembrane protein dysfunction. Biochim. Biophys. Acta 2009, 1793, 636–649. [Google Scholar] [CrossRef] [PubMed]
- Elmonem, M.A.; Veys, K.R.P.; Prencipe, G. Nephropathic cystinosis: Pathogenic roles of inflammation and potential for new therapies. Cells 2022, 11, 190. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| WT + Vehicle (n = 8) | WT + 25(OH)D3 (n = 8) | WT + 1,25(OH)2D3 (n = 8) | Ctns−/− + Vehicle (n = 8) | Ctns−/− + 25(OH)D3 (n = 8) | Ctns−/− + 1,25(OH)2D3 (n = 8) | |
|---|---|---|---|---|---|---|
| BUN (mg/dL) | 26.3 ± 4.3 | 27.9 ± 2.6 | 23.1 ± 4.3 | 67.6 ± 12.4 a | 57.6 ± 9.8 a | 65.7 ± 7.9 a |
| Creatinine (mg/dL) | 0.09 ± 0.02 | 0.08 ± 0.03 | 0.09 ± 0.02 | 0.24 ± 0.05 a | 0.23 ± 0.04 a | 0.21 ± 0.05 a |
| Bicarbonate (mmol/L) | 27.6 ± 2.3 | 27.8 ± 2.4 | 26.7 ± 2.7 | 26.7 ± 2.3 | 27.1 ± 5.6 | 26.7 ± 2.7 |
| 25(OH)D3 (ng/mL) | 104.2 ± 13.5 | 105.3 ± 13.9 | 113.6 ± 12.8 | 43.6± 3.4 a | 109.4 ± 13.7 b | 58.9 ± 5.7 a |
| 1,25(OH)2D3 (pg/mL) | 263.6 ± 31.5 | 201.7 ± 21.5 | 243.7 ± 12.8 | 125.6 ± 17.8 a | 193.4 ± 14.3 a,b | 276.1 ± 17.8 b |
| WT + Vehicle (n = 9) | WT + 25(OH)D3 (n = 9) | WT + 1,25(OH)2D3 (n = 9) | Ctns−/− + Vehicle (n = 9) | Ctns−/− + 25(OH)D3 (n = 9) | Ctns−/− + 1,25(OH)2D3 (n = 9) | |
|---|---|---|---|---|---|---|
| BUN (mg/dL) | 27.3 ± 4.3 | 22.7 ± 6.5 | 24.5 ± 2.5 | 65.9 ± 22.1 a | 75.4 ± 11.1 a | 76.9 ± 12.7 a |
| Creatinine (mg/dL) | 0.08 ± 0.04 | 0.09 ± 0.02 | 0.08 ± 0.03 | 0.21 ± 0.06 a | 0.26 ± 0.07 a | 0.28 ± 0.04 a |
| Bicarbonate (mmol/L) | 27.5 ± 2.6 | 27.1 ± 3.3 | 27.3 ± 2.4 | 26.7 ± 2.3 | 27.5 ± 4.3 | 26.9 ± 3.3 |
| 25(OH)D3 (ng/mL) | 121.8 ± 23.5 | 124.1 ± 21.5 | 109.5 ± 17.6 | 48.2 ± 6.9 a | 125.4 ± 23.7 b | 64.5 ± 11.3 a |
| 1,25(OH)2D3 (pg/mL) | 254.3 ± 24.3 | 213.6 ± 16.5 | 235.4 ± 23.6 | 126.4 ± 24.3 a | 189.8 ± 25.4 a,b | 254.3 ± 14.3 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, P.; Cheung, W.W.; Gonzalez, A.; Vaddi, V.; Oliveira, E.A.; Mak, R.H. Metabolic Advantage of 25(OH)D3 versus 1,25(OH)2D3 Supplementation in Infantile Nephropathic Cystinosis-Associated Adipose Tissue Browning and Muscle Wasting. Cells 2022, 11, 3264. https://doi.org/10.3390/cells11203264
Zhou P, Cheung WW, Gonzalez A, Vaddi V, Oliveira EA, Mak RH. Metabolic Advantage of 25(OH)D3 versus 1,25(OH)2D3 Supplementation in Infantile Nephropathic Cystinosis-Associated Adipose Tissue Browning and Muscle Wasting. Cells. 2022; 11(20):3264. https://doi.org/10.3390/cells11203264
Chicago/Turabian StyleZhou, Ping, Wai W. Cheung, Alex Gonzalez, Venya Vaddi, Eduardo A. Oliveira, and Robert H. Mak. 2022. "Metabolic Advantage of 25(OH)D3 versus 1,25(OH)2D3 Supplementation in Infantile Nephropathic Cystinosis-Associated Adipose Tissue Browning and Muscle Wasting" Cells 11, no. 20: 3264. https://doi.org/10.3390/cells11203264