
Mechanical Stretch Induced Osteogenesis on Human Annulus Fibrosus Cells through Upregulation of BMP-2/6 Heterodimer and Activation of P38 and SMAD1/5/8 Signaling Pathways


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patients
2.2. Reagents
2.3. Culture of Cell Line of AF Cells
2.4. Cyclic Stretching
2.5. Real-Time Quantitative PCR
2.6. Western Blot Analysis
2.7. siRNA Transfection
2.8. Statistical Analysis
3. Results
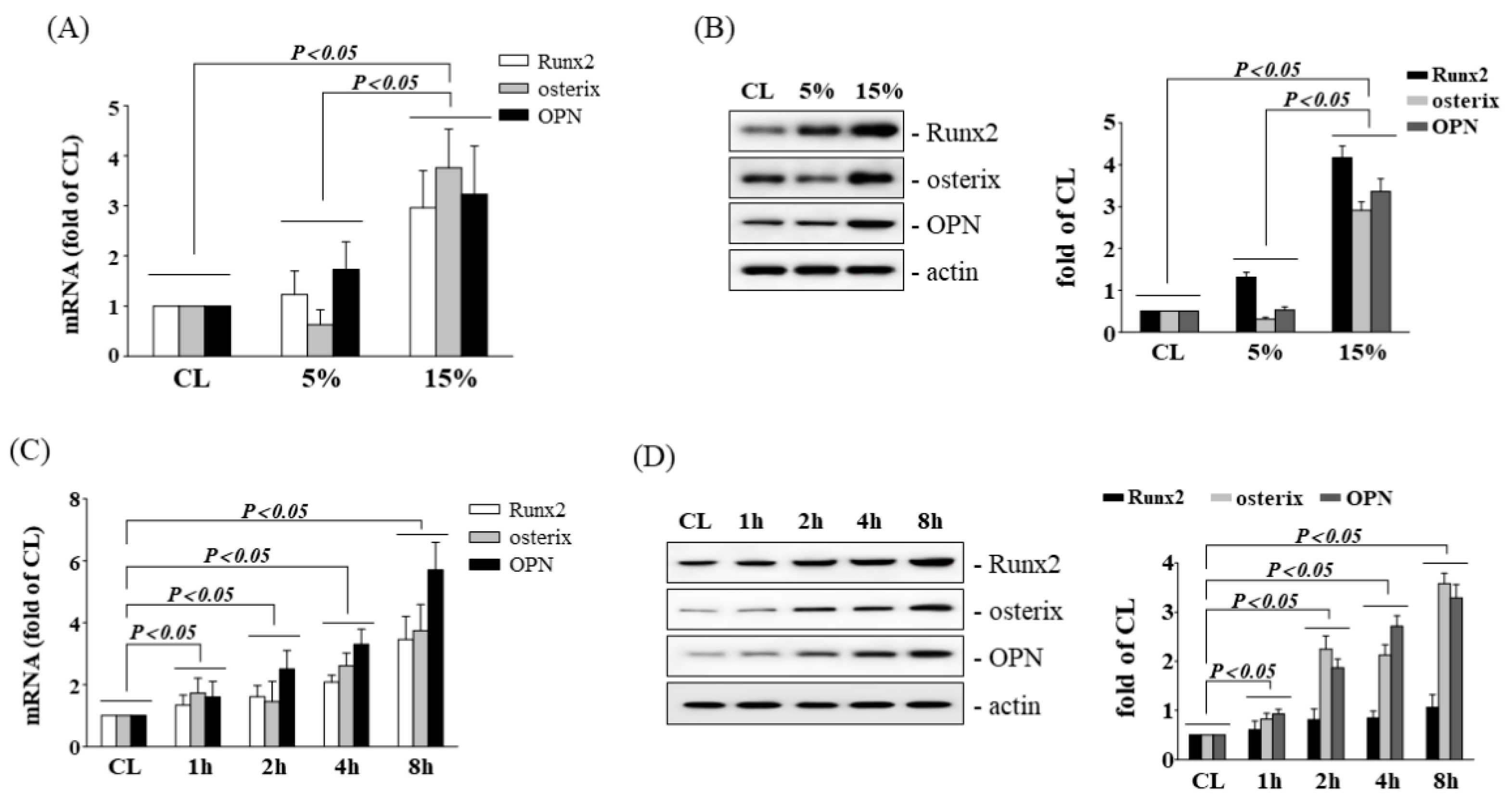
3.1. HCS Upregulates Osteogenic Gene Expression in Human AF Cells
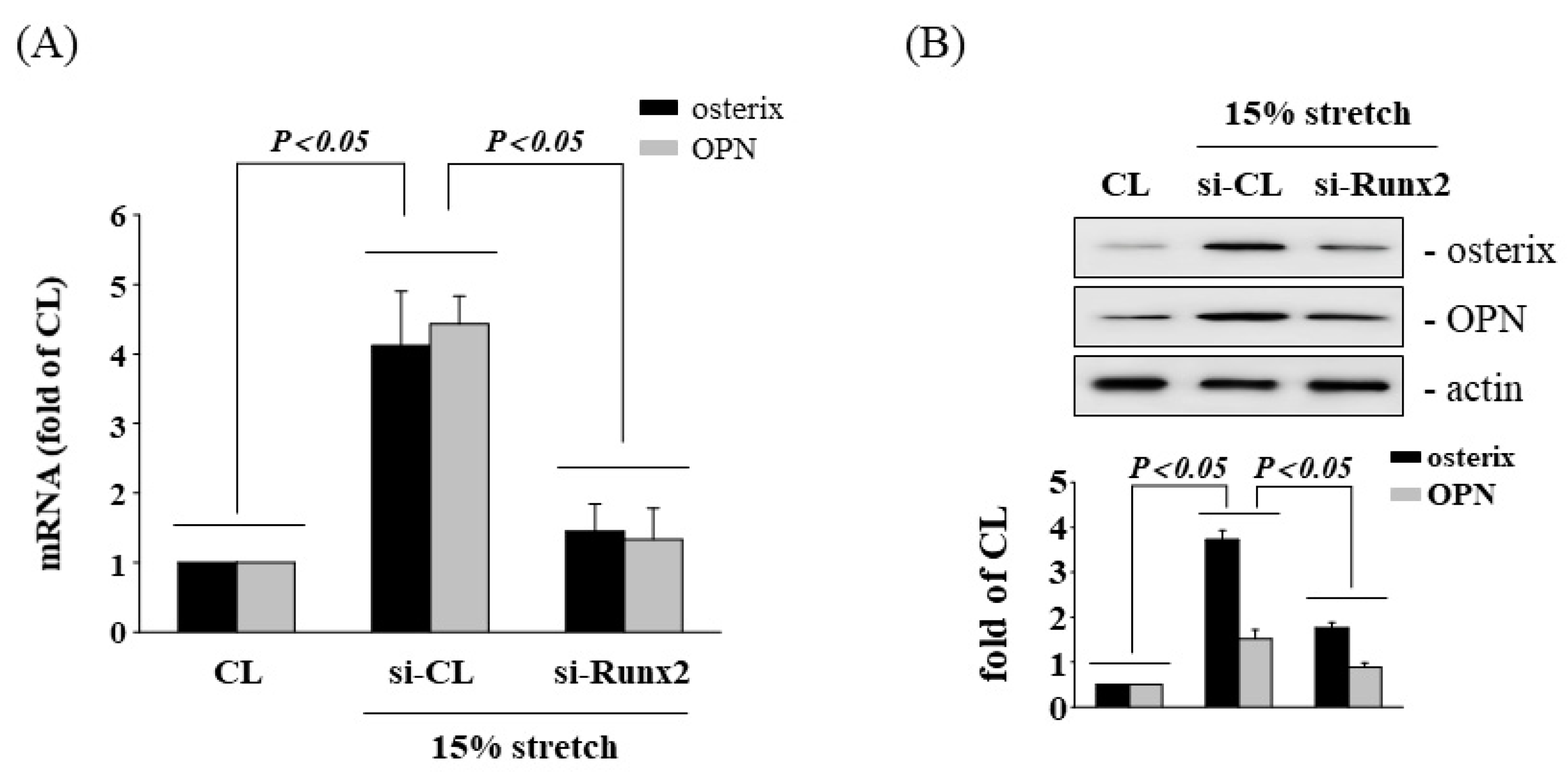
3.2. Involvement of Runx2 in HCS-Induced Osteogenesis
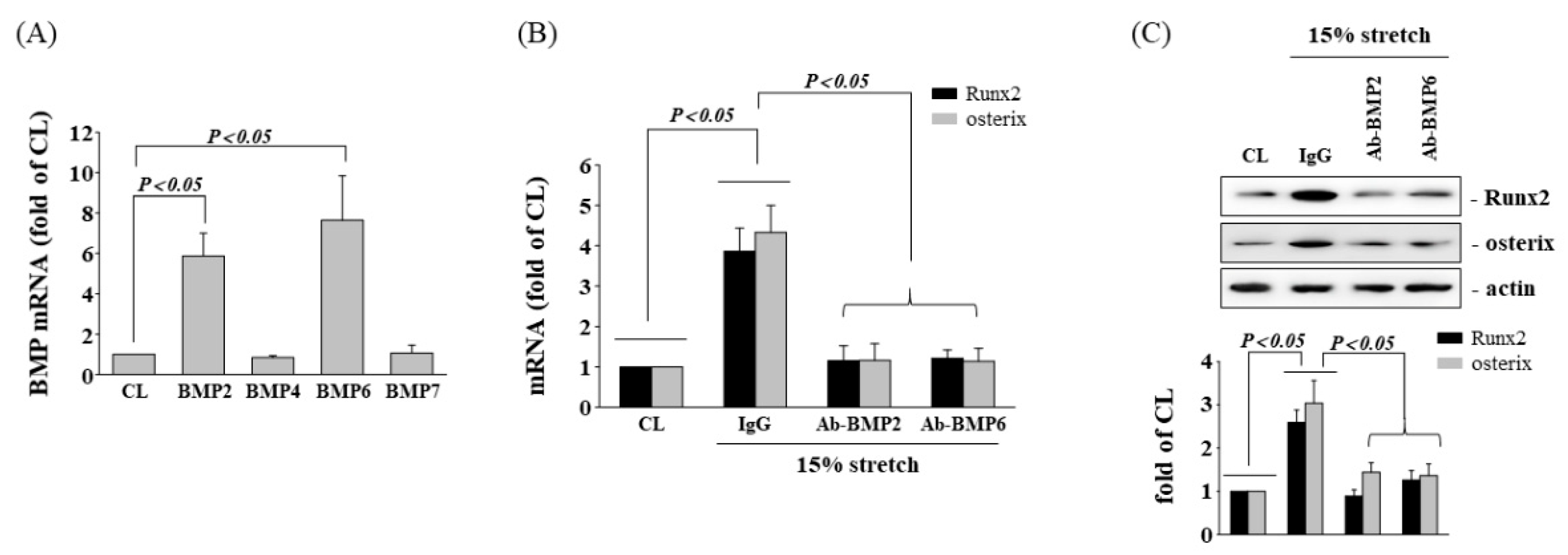
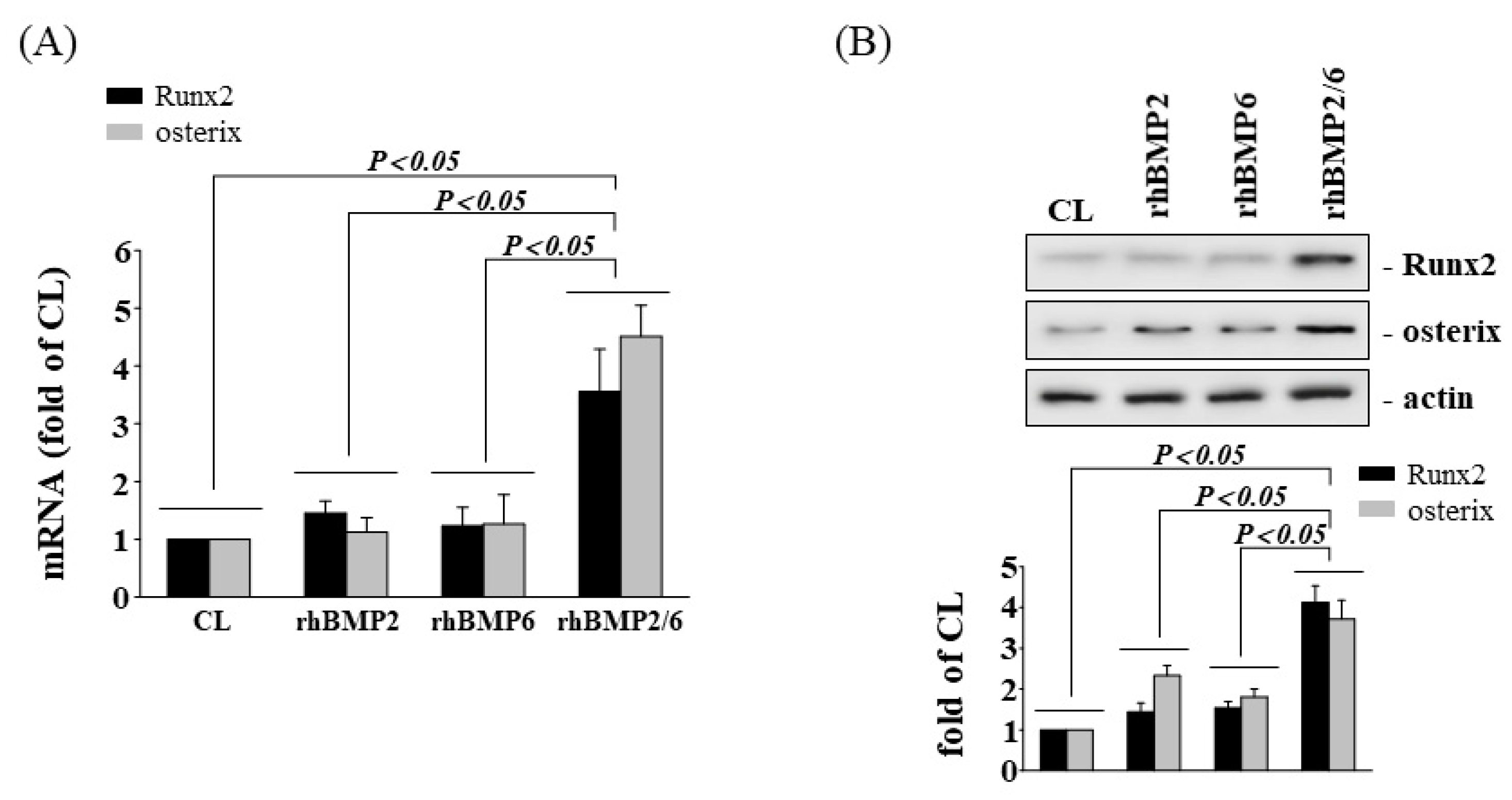
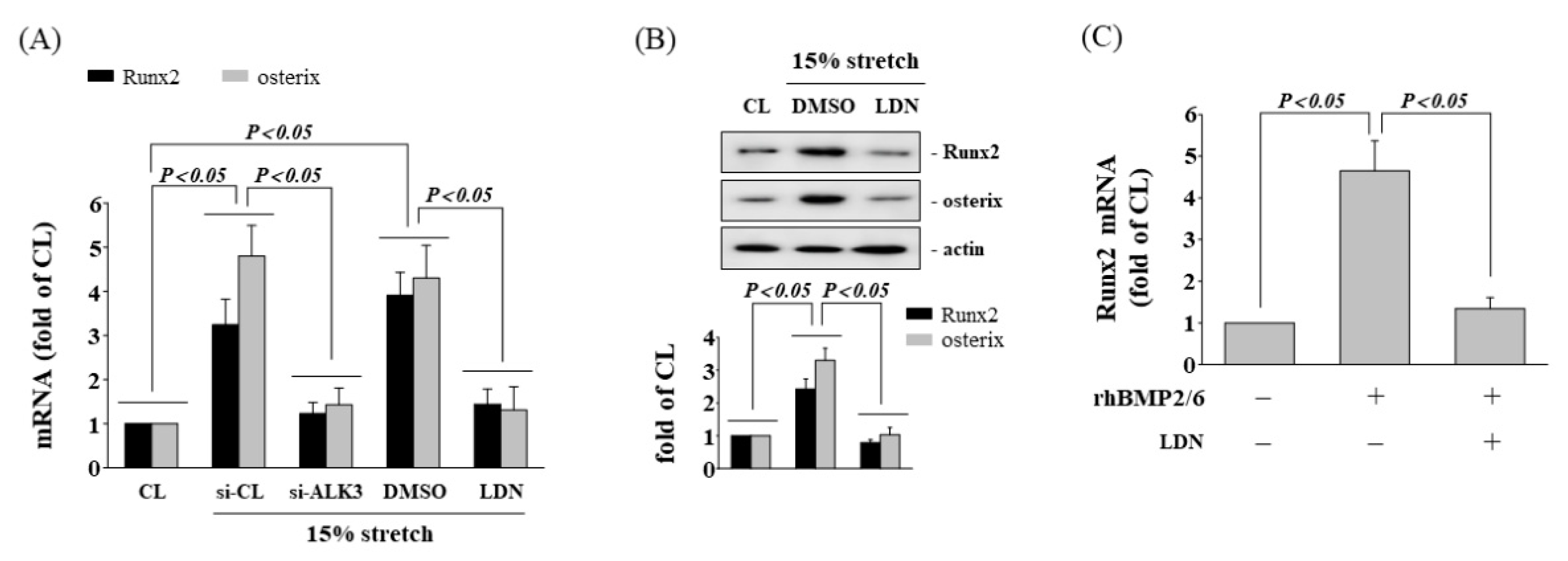
3.3. BMP-2/6 Heterodimer Mediates Stretch-Induced Osteogenic Gene Expression
3.4. HCS-Induced Runx2 Expression Is p38 MAPK Dependent
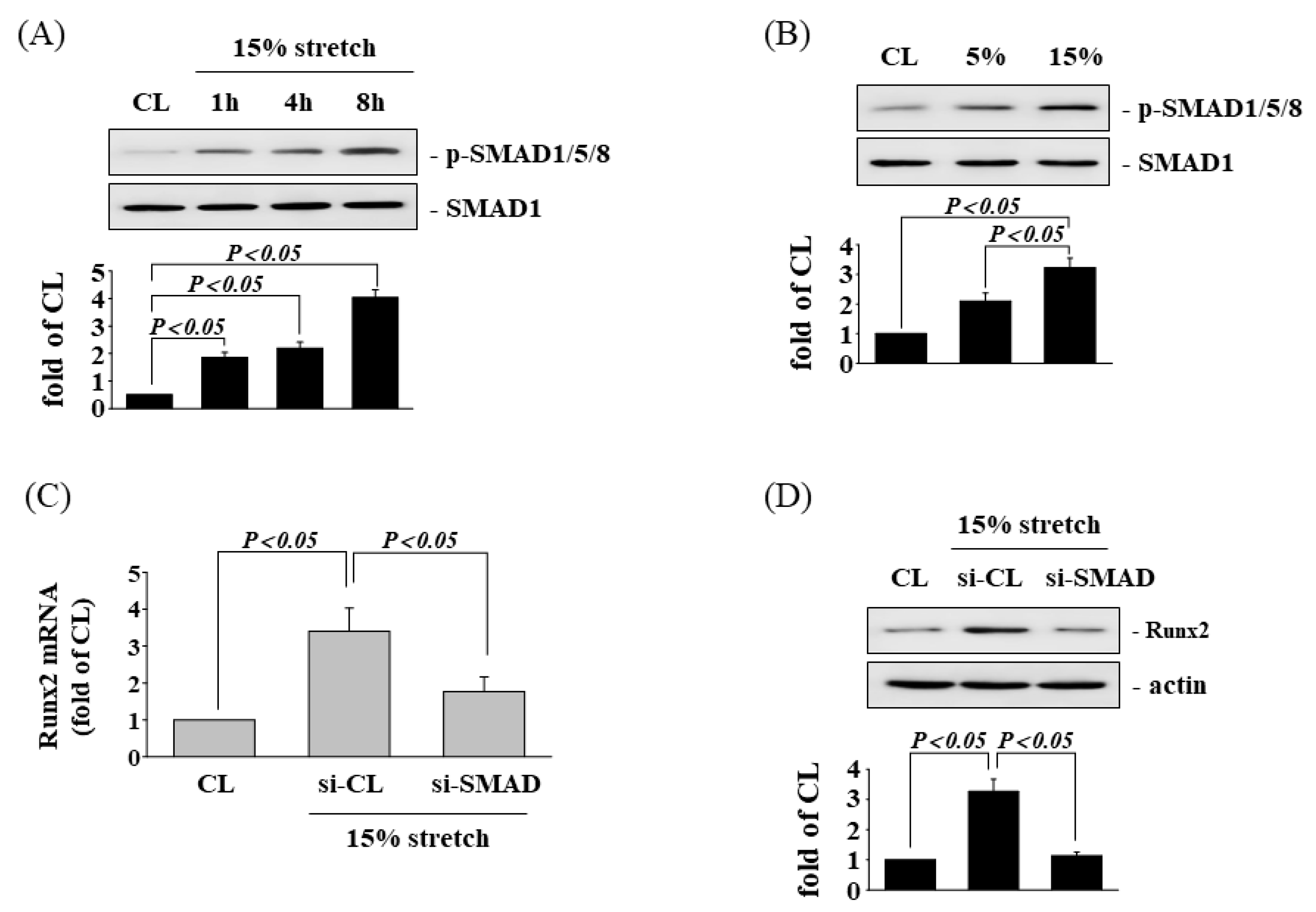
3.5. SAMD1/5/8 Is Involved in Tensile Stretch-Induced Runx2 Expression
3.6. Inhibition of ALK3 Receptor Decrease Tensile Stretch-Induced Osteogenic Gene Expression
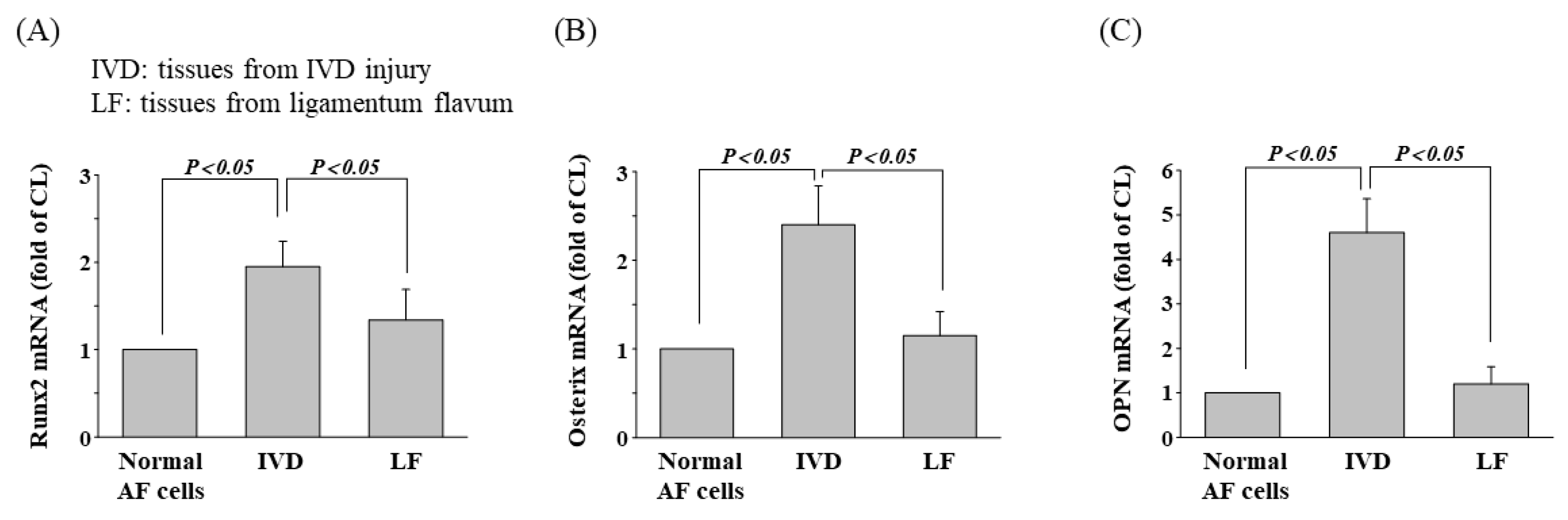
3.7. Osteogenic Gene Expression in Patients with IVD Injury
4. Discussion
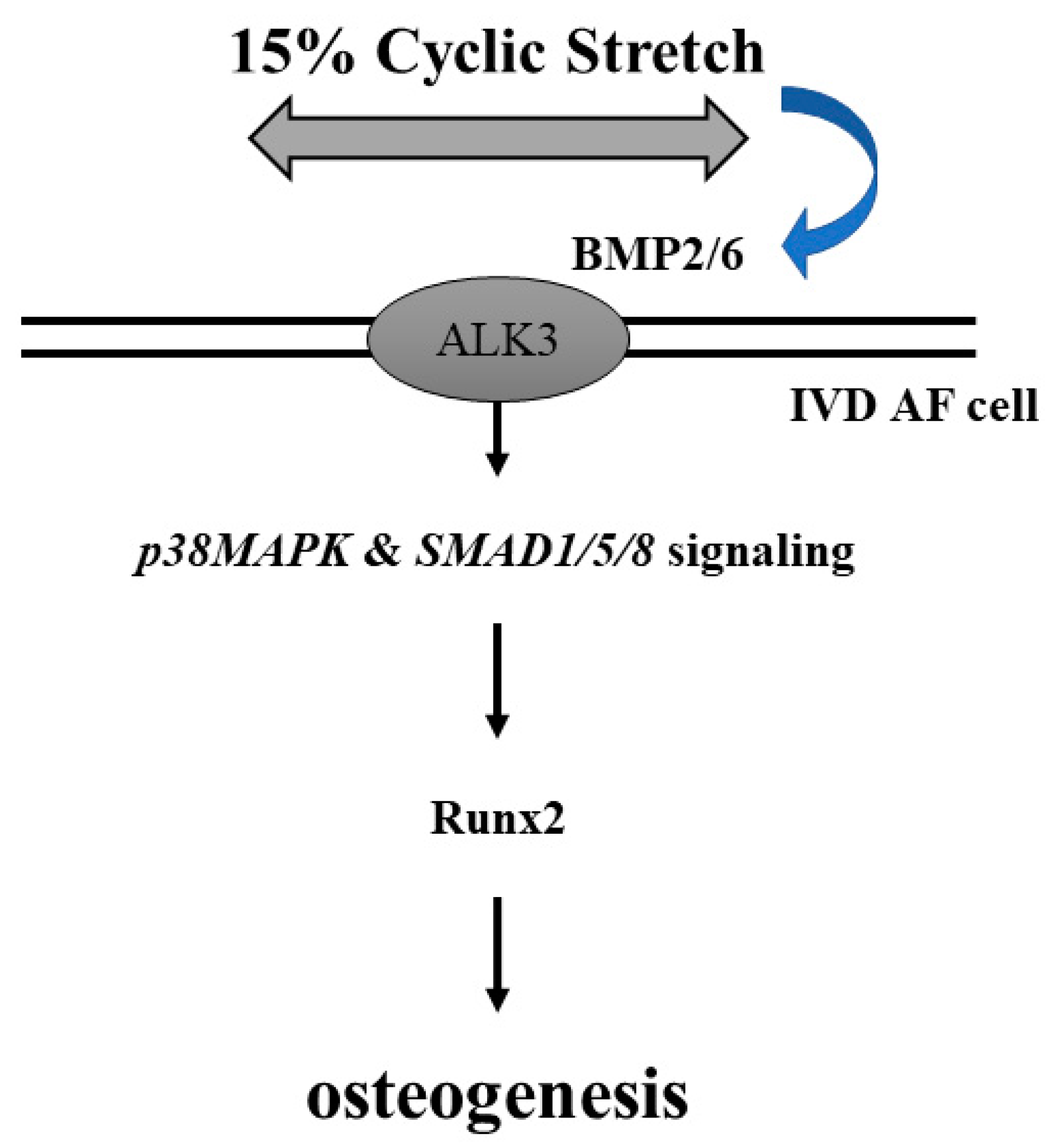
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Luoma, K.; Riihimaki, H.; Luukkonen, R.; Raininko, R.; Viikari-Juntura, E.; Lamminen, A. Low back pain in relation to lumbar disc degeneration. Spine 2000, 25, 487–492. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.G.; Guo, T.M.; Guo, X.; Wu, S.X. Clinical diagnosis for discogenic low back pain. Int. J. Biol. Sci. 2009, 5, 647–658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freemont, A.J. The cellular pathobiology of the degenerate intervertebral disc and discogenic back pain. Rheumatology 2009, 48, 5–10. [Google Scholar] [CrossRef] [Green Version]
- Medical Advisory, S. Artificial discs for lumbar and cervical degenerative disc disease -update: An evidence-based analysis. Ont. Health Technol. Assess. Ser. 2006, 6, 1–98. [Google Scholar]
- Katz, J.N. Lumbar disc disorders and low-back pain: Socioeconomic factors and consequences. J. Bone Joint Surg. Am. 2006, 88 (Suppl. 2), 21–24. [Google Scholar] [CrossRef]
- Urban, J.P.; Roberts, S. Degeneration of the intervertebral disc. Arthritis Res. Ther. 2003, 5, 120–130. [Google Scholar] [CrossRef] [Green Version]
- Vo, N.V.; Hartman, R.A.; Patil, P.R.; Risbud, M.V.; Kletsas, D.; Iatridis, J.C.; Hoyland, J.A.; Le Maitre, C.L.; Sowa, G.A.; Kang, J.D. Molecular mechanisms of biological aging in intervertebral discs. J. Orthop. Res. 2016, 34, 1289–1306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, Y.S. Pathophysiology of degenerative disc disease. Asian Spine J. 2009, 3, 39–44. [Google Scholar] [CrossRef]
- Urban, J.P.; Roberts, S. Development and degeneration of the intervertebral discs. Mol. Med. Today 1995, 1, 329–335. [Google Scholar] [CrossRef]
- Chan, D.; Song, Y.; Sham, P.; Cheung, K.M. Genetics of disc degeneration. Eur. Spine J. 2006, 15 (Suppl. 3), S317–S325. [Google Scholar] [CrossRef] [Green Version]
- Steelman, T.; Lewandowski, L.; Helgeson, M.; Wilson, K.; Olsen, C.; Gwinn, D. Population-based Risk Factors for the Development of Degenerative Disk Disease. Clin. Spine Surg. 2018, 31, E409–E412. [Google Scholar] [CrossRef] [PubMed]
- Gawri, R.; Rosenzweig, D.H.; Krock, E.; Ouellet, J.A.; Stone, L.S.; Quinn, T.M.; Haglund, L. High mechanical strain of primary intervertebral disc cells promotes secretion of inflammatory factors associated with disc degeneration and pain. Arthritis Res. Ther. 2014, 16, R21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gilbert, H.T.J.; Hodson, N.; Baird, P.; Richardson, S.M.; Hoyland, J.A. Acidic pH promotes intervertebral disc degeneration: Acid-sensing ion channel -3 as a potential therapeutic target. Sci. Rep. 2016, 6, 37360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osti, O.L.; Vernon-Roberts, B.; Moore, R.; Fraser, R.D. Annular tears and disc degeneration in the lumbar spine. A post-mortem study of 135 discs. J. Bone Joint Surg. Br. 1992, 74, 678–682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roberts, S.; Evans, H.; Trivedi, J.; Menage, J. Histology and pathology of the human intervertebral disc. J. Bone Joint Surg. Am. 2006, 88 (Suppl. 2), 10–14. [Google Scholar] [PubMed] [Green Version]
- Jaumard, N.V.; Welch, W.C.; Winkelstein, B.A. Spinal facet joint biomechanics and mechanotransduction in normal, injury and degenerative conditions. J. Biomech. Eng. 2011, 133, 071010. [Google Scholar] [CrossRef] [Green Version]
- Dowdell, J.; Erwin, M.; Choma, T.; Vaccaro, A.; Iatridis, J.; Cho, S.K. Intervertebral Disk Degeneration and Repair. Neurosurgery 2017, 80, S46–S54. [Google Scholar] [CrossRef]
- Jin, L.; Liu, Q.; Scott, P.; Zhang, D.; Shen, F.; Balian, G.; Li, X. Annulus fibrosus cell characteristics are a potential source of intervertebral disc pathogenesis. PLoS ONE 2014, 9, e96519. [Google Scholar] [CrossRef]
- Fearing, B.V.; Hernandez, P.A.; Setton, L.A.; Chahine, N.O. Mechanotransduction and cell biomechanics of the intervertebral disc. JOR Spine 2018, 1, e1026. [Google Scholar] [CrossRef]
- Zhang, W.; Wang, H.; Yuan, Z.; Chu, G.; Sun, H.; Yu, Z.; Liang, H.; Liu, T.; Zhou, F.; Li, B. Moderate mechanical stimulation rescues degenerative annulus fibrosus by suppressing caveolin-1 mediated pro-inflammatory signaling pathway. Int. J. Biol. Sci. 2021, 17, 1395–1412. [Google Scholar] [CrossRef]
- Walter, B.A.; Korecki, C.L.; Purmessur, D.; Roughley, P.J.; Michalek, A.J.; Iatridis, J.C. Complex loading affects intervertebral disc mechanics and biology. Osteoarthr. Cartil. 2011, 19, 1011–1018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berger-Roscher, N.; Casaroli, G.; Rasche, V.; Villa, T.; Galbusera, F.; Wilke, H.J. Influence of Complex Loading Conditions on Intervertebral Disc Failure. Spine 2017, 42, E78–E85. [Google Scholar] [CrossRef] [PubMed]
- Chu, G.; Shi, C.; Wang, H.; Zhang, W.; Yang, H.; Li, B. Strategies for Annulus Fibrosus Regeneration: From Biological Therapies to Tissue Engineering. Front. Bioeng. Biotechnol. 2018, 6, 90. [Google Scholar] [CrossRef] [PubMed]
- Kasamkattil, J.; Gryadunova, A.; Martin, I.; Barbero, A.; Scharen, S.; Krupkova, O.; Mehrkens, A. Spheroid-Based Tissue Engineering Strategies for Regeneration of the Intervertebral Disc. Int. J. Mol. Sci. 2022, 23, 2530. [Google Scholar] [CrossRef] [PubMed]
- Wuertz, K.; Haglund, L. Inflammatory mediators in intervertebral disk degeneration and discogenic pain. Glob. Spine J. 2013, 3, 175–184. [Google Scholar] [CrossRef] [Green Version]
- Alkhatib, B.; Rosenzweig, D.H.; Krock, E.; Roughley, P.J.; Beckman, L.; Steffen, T.; Weber, M.H.; Ouellet, J.A.; Haglund, L. Acute mechanical injury of the human intervertebral disc: Link to degeneration and pain. Eur. Cell Mater. 2014, 28, 98–110; discussion 110–111. [Google Scholar] [CrossRef]
- Brisby, H.; Papadimitriou, N.; Brantsing, C.; Bergh, P.; Lindahl, A.; Barreto Henriksson, H. The presence of local mesenchymal progenitor cells in human degenerated intervertebral discs and possibilities to influence these in vitro: A descriptive study in humans. Stem Cells Dev. 2013, 22, 804–814. [Google Scholar] [CrossRef]
- Liu, L.T.; Huang, B.; Li, C.Q.; Zhuang, Y.; Wang, J.; Zhou, Y. Characteristics of stem cells derived from the degenerated human intervertebral disc cartilage endplate. PLoS ONE 2011, 6, e26285. [Google Scholar] [CrossRef] [Green Version]
- Yourek, G.; McCormick, S.M.; Mao, J.J.; Reilly, G.C. Shear stress induces osteogenic differentiation of human mesenchymal stem cells. Regen. Med. 2010, 5, 713–724. [Google Scholar] [CrossRef] [Green Version]
- Schreivogel, S.; Kuchibhotla, V.; Knaus, P.; Duda, G.N.; Petersen, A. Load-induced osteogenic differentiation of mesenchymal stromal cells is caused by mechano-regulated autocrine signaling. J. Tissue Eng. Regen. Med. 2019, 13, 1992–2008. [Google Scholar] [CrossRef] [Green Version]
- Huang, K.Y.; Yan, J.J.; Hsieh, C.C.; Chang, M.S.; Lin, R.M. The in vivo biological effects of intradiscal recombinant human bone morphogenetic protein-2 on the injured intervertebral disc: An animal experiment. Spine 2007, 32, 1174–1180. [Google Scholar] [CrossRef] [PubMed]
- May, R.D.; Frauchiger, D.A.; Albers, C.E.; Tekari, A.; Benneker, L.M.; Klenke, F.M.; Hofstetter, W.; Gantenbein, B. Application of Cytokines of the Bone Morphogenetic Protein (BMP) Family in Spinal Fusion—Effects on the Bone, Intervertebral Disc and Mesenchymal Stromal Cells. Curr. Stem Cell Res. Ther. 2019, 14, 618–643. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.S.; Akhtar, N.; Jamil, H.M.; Banik, R.S.; Asaduzzaman, S.M. TGF-beta/BMP signaling and other molecular events: Regulation of osteoblastogenesis and bone formation. Bone Res. 2015, 3, 15005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kirker-Head, C.A. Potential applications and delivery strategies for bone morphogenetic proteins. Adv. Drug Deliv. Rev. 2000, 43, 65–92. [Google Scholar] [CrossRef]
- Kim, D.J.; Moon, S.H.; Kim, H.; Kwon, U.H.; Park, M.S.; Han, K.J.; Hahn, S.B.; Lee, H.M. Bone morphogenetic protein-2 facilitates expression of chondrogenic, not osteogenic, phenotype of human intervertebral disc cells. Spine 2003, 28, 2679–2684. [Google Scholar] [CrossRef]
- Gulati, T.; Chung, S.A.; Wei, A.Q.; Diwan, A.D. Localization of bone morphogenetic protein 13 in human intervertebral disc and its molecular and functional effects in vitro in 3D culture. J. Orthop. Res. 2015, 33, 1769–1775. [Google Scholar] [CrossRef]
- Carlisle, E.; Fischgrund, J.S. Bone morphogenetic proteins for spinal fusion. Spine J. 2005, 5 (Suppl. 6), 240S–249S. [Google Scholar] [CrossRef]
- Hecht, B.P.; Fischgrund, J.S.; Herkowitz, H.N.; Penman, L.; Toth, J.M.; Shirkhoda, A. The use of recombinant human bone morphogenetic protein 2 (rhBMP-2) to promote spinal fusion in a nonhuman primate anterior interbody fusion model. Spine 1999, 24, 629–636. [Google Scholar] [CrossRef] [Green Version]
- Welch, R.D.; Jones, A.L.; Bucholz, R.W.; Reinert, C.M.; Tjia, J.S.; Pierce, W.A.; Wozney, J.M.; Li, X.J. Effect of recombinant human bone morphogenetic protein-2 on fracture healing in a goat tibial fracture model. J. Bone Miner. Res. 1998, 13, 1483–1490. [Google Scholar] [CrossRef]
- Nakamura, Y.; Nakaya, H.; Saito, N.; Wakitani, S. Coordinate expression of BMP-2, BMP receptors and Noggin in normal mouse spine. J. Clin. Neurosci. 2006, 13, 250–256. [Google Scholar] [CrossRef]
- Takae, R.; Matsunaga, S.; Origuchi, N.; Yamamoto, T.; Morimoto, N.; Suzuki, S.; Sakou, T. Immunolocalization of bone morphogenetic protein and its receptors in degeneration of intervertebral disc. Spine 1999, 24, 1397–1401. [Google Scholar] [CrossRef] [PubMed]
- Hsu, Y.H.; Lin, R.M.; Chiu, Y.S.; Liu, W.L.; Huang, K.Y. Effects of IL-1β, IL-20, and BMP-2 on Intervertebral Disc Inflammation under Hypoxia. J. Clin. Med. 2020, 9, 140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, K.Y.; Lin, R.M.; Chen, W.Y.; Lee, C.L.; Yan, J.J.; Chang, M.S. IL-20 may contribute to the pathogenesis of human intervertebral disc herniation. Spine 2008, 33, 2034–2040. [Google Scholar] [CrossRef] [PubMed]
- Huang, K.Y.; Hsu, Y.H.; Chen, W.Y.; Tsai, H.L.; Yan, J.J.; Wang, J.D.; Liu, W.L.; Lin, R.M. The roles of IL-19 and IL-20 in the inflammation of degenerative lumbar spondylolisthesis. J. Inflamm. 2018, 15, 19. [Google Scholar] [CrossRef] [Green Version]
- Valera, E.; Isaacs, M.J.; Kawakami, Y.; Izpisua Belmonte, J.C.; Choe, S. BMP-2/6 heterodimer is more effective than BMP-2 or BMP-6 homodimers as inductor of differentiation of human embryonic stem cells. PLoS ONE 2010, 5, e11167. [Google Scholar] [CrossRef] [Green Version]
- Chang, S.F.; Huang, K.C.; Chang, H.I.; Lee, K.C.; Su, Y.P.; Chen, C.N. 2 dyn/cm(2) shear force upregulates kruppel-like factor 4 expression in human chondrocytes to inhibit the interleukin-1beta-activated nuclear factor-kappaB. J. Cell Physiol. 2018, 234, 958–968. [Google Scholar] [CrossRef] [Green Version]
- Feng, G.; Yang, X.; Shang, H.; Marks, I.W.; Shen, F.H.; Katz, A.; Arlet, V.; Laurencin, C.T.; Li, X. Multipotential differentiation of human anulus fibrosus cells: An in vitro study. J. Bone Joint Surg. Am. 2010, 92, 675–685. [Google Scholar] [CrossRef] [PubMed]
- Stich, S.; Jagielski, M.; Fleischmann, A.; Meier, C.; Bussmann, P.; Kohl, B.; Schmidt, J.; Kruger, J.P.; Endres, M.; Cabraja, M.; et al. Degeneration of Lumbar Intervertebral Discs: Characterization of Anulus Fibrosus Tissue and Cells of Different Degeneration Grades. Int. J. Mol. Sci. 2020, 21, 2165. [Google Scholar] [CrossRef] [Green Version]
- Yeh, C.H.; Jin, L.; Shen, F.; Balian, G.; Li, X.J. miR-221 attenuates the osteogenic differentiation of human annulus fibrosus cells. Spine J. 2016, 16, 896–904. [Google Scholar] [CrossRef] [Green Version]
- Rannou, F.; Lee, T.S.; Zhou, R.H.; Chin, J.; Lotz, J.C.; Mayoux-Benhamou, M.A.; Barbet, J.P.; Chevrot, A.; Shyy, J.Y. Intervertebral disc degeneration: The role of the mitochondrial pathway in annulus fibrosus cell apoptosis induced by overload. Am. J. Pathol 2004, 164, 915–924. [Google Scholar] [CrossRef]
- Gilbert, H.T.; Hoyland, J.A.; Freemont, A.J.; Millward-Sadler, S.J. The involvement of interleukin-1 and interleukin-4 in the response of human annulus fibrosus cells to cyclic tensile strain: An altered mechanotransduction pathway with degeneration. Arthritis Res. Ther. 2011, 13, R8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pratsinis, H.; Papadopoulou, A.; Neidlinger-Wilke, C.; Brayda-Bruno, M.; Wilke, H.J.; Kletsas, D. Cyclic tensile stress of human annulus fibrosus cells induces MAPK activation: Involvement in proinflammatory gene expression. Osteoarthr. Cartil. 2016, 24, 679–687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.J.; Liao, H.Y.; Bai, D.Y.; Wang, Z.Q.; Xie, X.W. MAPK /ERK signaling pathway: A potential target for the treatment of intervertebral disc degeneration. Biomed. Pharmacother. 2021, 143, 112170. [Google Scholar] [CrossRef] [PubMed]
- Leung, V.Y.L.; Zhou, L.; Tam, W.K.; Sun, Y.; Lv, F.; Zhou, G.; Cheung, K.M.C. Bone morphogenetic protein-2 and -7 mediate the anabolic function of nucleus pulposus cells with discrete mechanisms. Connect. Tissue Res. 2017, 58, 573–585. [Google Scholar] [CrossRef]
- Chang, H.I.; Chen, C.N.; Huang, K.Y. Mechanical Stretch-Induced NLRP3 Inflammasome Expression on Human Annulus Fibrosus Cells Modulated by Endoplasmic Reticulum Stress. Int J. Mol. Sci 2022, 23, 7951. [Google Scholar] [CrossRef]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, C.-N.; Chang, H.-I.; Yen, C.-K.; Liu, W.-L.; Huang, K.-Y. Mechanical Stretch Induced Osteogenesis on Human Annulus Fibrosus Cells through Upregulation of BMP-2/6 Heterodimer and Activation of P38 and SMAD1/5/8 Signaling Pathways. Cells 2022, 11, 2600. https://doi.org/10.3390/cells11162600
Chen C-N, Chang H-I, Yen C-K, Liu W-L, Huang K-Y. Mechanical Stretch Induced Osteogenesis on Human Annulus Fibrosus Cells through Upregulation of BMP-2/6 Heterodimer and Activation of P38 and SMAD1/5/8 Signaling Pathways. Cells. 2022; 11(16):2600. https://doi.org/10.3390/cells11162600
Chicago/Turabian StyleChen, Cheng-Nan, Hsin-I Chang, Chia-Kung Yen, Wen-Lung Liu, and Kuo-Yuan Huang. 2022. "Mechanical Stretch Induced Osteogenesis on Human Annulus Fibrosus Cells through Upregulation of BMP-2/6 Heterodimer and Activation of P38 and SMAD1/5/8 Signaling Pathways" Cells 11, no. 16: 2600. https://doi.org/10.3390/cells11162600