
Transcriptome Profiling Reveals Differential Expression of Circadian Behavior Genes in Peripheral Blood of Monozygotic Twins Discordant for Parkinson’s Disease

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patients
2.2. Sequencing and RNA-Seq Data Analysis
2.3. RNA Isolation and Expression Analysis of Individual Candidate Genes
2.4. Statistical Processing and Bioinformatic Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- De Lau, L.M.; Breteler, M.M. Epidemiology of Parkinson’s disease. Lancet Neurol. 2006, 5, 525–535. [Google Scholar] [CrossRef]
- Kalia, L.V.; Lang, A.E. Parkinson’s disease. Lancet 2015, 386, 896–912. [Google Scholar] [CrossRef]
- Surguchov, A. Biomarkers in Parkinson’s Disease. In Neurodegenerative Diseases Biomarkers: Towards Translating Research to Clinical Practice; Peplow, P.V., Martinez, B., Gennarelli, T.A., Eds.; Springer: New York, NY, USA, 2022; pp. 155–180. [Google Scholar] [CrossRef]
- Zeng, X.S.; Geng, W.S.; Jia, J.J.; Chen, L.; Zhang, P.P. Cellular and Molecular Basis of Neurodegeneration in Parkinson Disease. Front. Aging Neurosci. 2018, 10, 109. [Google Scholar] [CrossRef] [Green Version]
- Schapira, A.H.V.; Chaudhuri, K.R.; Jenner, P. Non-motor features of Parkinson disease. Nat. Rev. Neurosci. 2017, 18, 435–450. [Google Scholar] [CrossRef] [PubMed]
- Shan, L.; Dauvilliers, Y.; Siegel, J.M. Interactions of the histamine and hypocretin systems in CNS disorders. Nat. Rev. Neurol. 2015, 11, 401–413. [Google Scholar] [CrossRef]
- Caronti, B.; Antonini, G.; Calderaro, C.; Ruggieri, S.; Palladini, G.; Pontieri, F.E.; Colosimo, C. Dopamine transporter immunoreactivity in peripheral blood lymphocytes in Parkinson’s disease. J. Neural Transm. 2001, 108, 803–807. [Google Scholar] [CrossRef]
- Buttarelli, F.R.; Capriotti, G.; Pellicano, C.; Prosperi, D.; Circella, A.; Festa, A.; Giovannelli, M.; Tofani, A.; Pontieri, F.E.; Scopinaro, F. Central and peripheral dopamine transporter reduction in Parkinson’s disease. Neurol. Res. 2009, 31, 687–691. [Google Scholar] [CrossRef]
- Barbanti, P.; Fabbrini, G.; Ricci, A.; Cerbo, R.; Bronzetti, E.; Caronti, B.; Calderaro, C.; Felici, L.; Stocchi, F.; Meco, G.; et al. Increased expression of dopamine receptors on lymphocytes in Parkinson’s disease. Mov. Disord. 1999, 14, 764–771. [Google Scholar] [CrossRef]
- Karlsson, M.K.; Sharma, P.; Aasly, J.; Toft, M.; Skogar, O.; Saebo, S.; Lonneborg, A. Found in transcription: Accurate Parkinson’s disease classification in peripheral blood. J. Park. Dis. 2013, 3, 19–29. [Google Scholar] [CrossRef] [Green Version]
- Shamir, R.; Klein, C.; Amar, D.; Vollstedt, E.J.; Bonin, M.; Usenovic, M.; Wong, Y.C.; Maver, A.; Poths, S.; Safer, H.; et al. Analysis of blood-based gene expression in idiopathic Parkinson disease. Neurology 2017, 89, 1676–1683. [Google Scholar] [CrossRef]
- Calligaris, R.; Banica, M.; Roncaglia, P.; Robotti, E.; Finaurini, S.; Vlachouli, C.; Antonutti, L.; Iorio, F.; Carissimo, A.; Cattaruzza, T.; et al. Blood transcriptomics of drug-naive sporadic Parkinson’s disease patients. BMC Genom. 2015, 16, 876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, Y.; Xiao, S. Progression Rate Associated Peripheral Blood Biomarkers of Parkinson’s Disease. J. Mol. Neurosci. 2018, 65, 312–318. [Google Scholar] [CrossRef] [PubMed]
- Alieva, A.; Shadrina, M.I.; Filatova, E.V.; Karabanov, A.V.; Illarioshkin, S.N.; Limborska, S.A.; Slominsky, P.A. Involvement of endocytosis and alternative splicing in the formation of the pathological process in the early stages of Parkinson’s disease. Biomed. Res. Int. 2014, 2014, 718732. [Google Scholar] [CrossRef] [PubMed]
- Chi, L.M.; Wang, L.P.; Jiao, D. Identification of Differentially Expressed Genes and Long Noncoding RNAs Associated with Parkinson’s Disease. Park. Dis. 2019, 2019, 6078251. [Google Scholar] [CrossRef] [Green Version]
- Santiago, J.A.; Potashkin, J.A. Blood Transcriptomic Meta-analysis Identifies Dysregulation of Hemoglobin and Iron Metabolism in Parkinson’ Disease. Front. Aging Neurosci. 2017, 9, 73. [Google Scholar] [CrossRef] [Green Version]
- Jiang, F.; Wu, Q.; Sun, S.; Bi, G.; Guo, L. Identification of potential diagnostic biomarkers for Parkinson’s disease. FEBS Open Bio 2019, 9, 1460–1468. [Google Scholar] [CrossRef] [Green Version]
- Scherzer, C.R.; Eklund, A.C.; Morse, L.J.; Liao, Z.; Locascio, J.J.; Fefer, D.; Schwarzschild, M.A.; Schlossmacher, M.G.; Hauser, M.A.; Vance, J.M.; et al. Molecular markers of early Parkinson’s disease based on gene expression in blood. Proc. Natl. Acad. Sci. USA 2007, 104, 955–960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pinho, R.; Guedes, L.C.; Soreq, L.; Lobo, P.P.; Mestre, T.; Coelho, M.; Rosa, M.M.; Goncalves, N.; Wales, P.; Mendes, T.; et al. Gene Expression Differences in Peripheral Blood of Parkinson’s Disease Patients with Distinct Progression Profiles. PLoS ONE 2016, 11, e0157852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alieva, A.; Filatova, E.V.; Karabanov, A.V.; Illarioshkin, S.N.; Slominsky, P.A.; Shadrina, M.I. Potential Biomarkers of the Earliest Clinical Stages of Parkinson’s Disease. Park. Dis. 2015, 2015, 294396. [Google Scholar] [CrossRef] [Green Version]
- Castillo-Fernandez, J.E.; Spector, T.D.; Bell, J.T. Epigenetics of discordant monozygotic twins: Implications for disease. Genome Med. 2014, 6, 60. [Google Scholar] [CrossRef]
- Woodard, C.M.; Campos, B.A.; Kuo, S.H.; Nirenberg, M.J.; Nestor, M.W.; Zimmer, M.; Mosharov, E.V.; Sulzer, D.; Zhou, H.; Paull, D.; et al. iPSC-derived dopamine neurons reveal differences between monozygotic twins discordant for Parkinson’s disease. Cell Rep. 2014, 9, 1173–1182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaut, O.; Schmitt, I.; Tost, J.; Busato, F.; Liu, Y.; Hofmann, P.; Witt, S.H.; Rietschel, M.; Frohlich, H.; Wullner, U. Epigenome-wide DNA methylation analysis in siblings and monozygotic twins discordant for sporadic Parkinson’s disease revealed different epigenetic patterns in peripheral blood mononuclear cells. Neurogenetics 2017, 18, 7–22. [Google Scholar] [CrossRef] [PubMed]
- Mazzetti, S.; Basellini, M.J.; Ferri, V.; Cassani, E.; Cereda, E.; Paolini, M.; Calogero, A.M.; Bolliri, C.; De Leonardis, M.; Sacilotto, G.; et al. alpha-Synuclein oligomers in skin biopsy of idiopathic and monozygotic twin patients with Parkinson’s disease. Brain 2020, 143, 920–931. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dulovic-Mahlow, M.; Konig, I.R.; Trinh, J.; Diaw, S.H.; Urban, P.P.; Knappe, E.; Kuhnke, N.; Ingwersen, L.C.; Hinrichs, F.; Weber, J.; et al. Discordant Monozygotic Parkinson Disease Twins: Role of Mitochondrial Integrity. Ann. Neurol. 2021, 89, 158–164. [Google Scholar] [CrossRef]
- Alieva, A.K.; Rudenok, M.M.; Novosadova, E.V.; Vlasov, I.N.; Arsenyeva, E.L.; Rosinskaya, A.V.; Grivennikov, I.A.; Slominsky, P.A.; Shadrina, M.I. Whole-Transcriptome Analysis of Dermal Fibroblasts, Derived from Three Pairs of Monozygotic Twins, Discordant for Parkinson’s Disease. J. Mol. Neurosci. 2020, 70, 284–293. [Google Scholar] [CrossRef]
- Vlasov, I.N.; Alieva, A.K.; Novosadova, E.V.; Arsenyeva, E.L.; Rosinskaya, A.V.; Partevian, S.A.; Grivennikov, I.A.; Shadrina, M.I. Transcriptome Analysis of Induced Pluripotent Stem Cells and Neuronal Progenitor Cells, Derived from Discordant Monozygotic Twins with Parkinson’s Disease. Cells 2021, 10, 3478. [Google Scholar] [CrossRef]
- Suslov, O.; Steindler, D.A. PCR inhibition by reverse transcriptase leads to an overestimation of amplification efficiency. Nucleic Acids Res. 2005, 33, e181. [Google Scholar] [CrossRef]
- Gene Ontology Consortium. The Gene Ontology Resource: 20 years and still GOing strong. Nucleic Acids Res. 2019, 47, D330–D338. [Google Scholar] [CrossRef] [Green Version]
- Bindea, G.; Mlecnik, B.; Hackl, H.; Charoentong, P.; Tosolini, M.; Kirilovsky, A.; Fridman, W.H.; Pages, F.; Trajanoski, Z.; Galon, J. ClueGO: A Cytoscape plug-in to decipher functionally grouped gene ontology and pathway annotation networks. Bioinformatics 2009, 25, 1091–1093. [Google Scholar] [CrossRef] [Green Version]
- Bindea, G.; Galon, J.; Mlecnik, B. CluePedia Cytoscape plugin: Pathway insights using integrated experimental and in silico data. Bioinformatics 2013, 29, 661–663. [Google Scholar] [CrossRef]
- Wheeler, D.L.; Church, D.M.; Federhen, S.; Lash, A.E.; Madden, T.L.; Pontius, J.U.; Schuler, G.D.; Schriml, L.M.; Sequeira, E.; Tatusova, T.A.; et al. Database resources of the National Center for Biotechnology. Nucleic Acids Res. 2003, 31, 28–33. [Google Scholar] [CrossRef] [PubMed]
- Nassan, M.; Videnovic, A. Circadian rhythms in neurodegenerative disorders. Nat. Rev. Neurol. 2022, 18, 7–24. [Google Scholar] [CrossRef] [PubMed]
- Videnovic, A.; Golombek, D. Circadian Dysregulation in Parkinson’s Disease. Neurobiol. Sleep Circadian Rhythm. 2017, 2, 53–58. [Google Scholar] [CrossRef] [PubMed]
- Hunt, J.; Coulson, E.J.; Rajnarayanan, R.; Oster, H.; Videnovic, A.; Rawashdeh, O. Sleep and circadian rhythms in Parkinson’s disease and preclinical models. Mol. Neurodegener 2022, 17, 2. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Wang, Y.; Wang, F.; Hu, L.F.; Liu, C.F. A New Perspective for Parkinson’s Disease: Circadian Rhythm. Neurosci. Bull. 2017, 33, 62–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, Y.; Liu, S.; Sothern, R.B.; Xu, S.; Chan, P. Expression of clock genes Per1 and Bmal1 in total leukocytes in health and Parkinson’s disease. Eur. J. Neurol. 2010, 17, 550–554. [Google Scholar] [CrossRef]
- Ding, H.; Liu, S.; Yuan, Y.; Lin, Q.; Chan, P.; Cai, Y. Decreased expression of Bmal2 in patients with Parkinson’s disease. Neurosci. Lett. 2011, 499, 186–188. [Google Scholar] [CrossRef] [PubMed]
- Breen, D.P.; Vuono, R.; Nawarathna, U.; Fisher, K.; Shneerson, J.M.; Reddy, A.B.; Barker, R.A. Sleep and circadian rhythm regulation in early Parkinson disease. JAMA Neurol. 2014, 71, 589–595. [Google Scholar] [CrossRef]
- Fifel, K.; Videnovic, A. Circadian and Sleep Dysfunctions in Neurodegenerative Disorders-An Update. Front. Neurosci. 2020, 14, 627330. [Google Scholar] [CrossRef]
- Li, T.; Cheng, C.; Jia, C.; Leng, Y.; Qian, J.; Yu, H.; Liu, Y.; Wang, N.; Yang, Y.; Al-Nusaif, M.; et al. Peripheral Clock System Abnormalities in Patients With Parkinson’s Disease. Front. Aging Neurosci. 2021, 13, 736026. [Google Scholar] [CrossRef]
- Urade, Y.; Eguchi, N. Lipocalin-type and hematopoietic prostaglandin D synthases as a novel example of functional convergence. Prostaglandins Other Lipid Mediat. 2002, 68–69, 375–382. [Google Scholar] [CrossRef]
- Ahmad, A.S.; Ottallah, H.; Maciel, C.B.; Strickland, M.; Dore, S. Role of the L-PGDS-PGD2-DP1 receptor axis in sleep regulation and neurologic outcomes. Sleep 2019, 42, zsz073. [Google Scholar] [CrossRef]
- Pinzar, E.; Kanaoka, Y.; Inui, T.; Eguchi, N.; Urade, Y.; Hayaishi, O. Prostaglandin D synthase gene is involved in the regulation of non-rapid eye movement sleep. Proc. Natl. Acad. Sci. USA 2000, 97, 4903–4907. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Onoe, H.; Ueno, R.; Fujita, I.; Nishino, H.; Oomura, Y.; Hayaishi, O. Prostaglandin D2, a cerebral sleep-inducing substance in monkeys. Proc. Natl. Acad. Sci. USA 1988, 85, 4082–4086. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mizoguchi, A.; Eguchi, N.; Kimura, K.; Kiyohara, Y.; Qu, W.M.; Huang, Z.L.; Mochizuki, T.; Lazarus, M.; Kobayashi, T.; Kaneko, T.; et al. Dominant localization of prostaglandin D receptors on arachnoid trabecular cells in mouse basal forebrain and their involvement in the regulation of non-rapid eye movement sleep. Proc. Natl. Acad. Sci. USA 2001, 98, 11674–11679. [Google Scholar] [CrossRef] [Green Version]
- Wang, P.; Li, Q.; Dong, X.; An, H.; Li, J.; Zhao, L.; Yan, H.; Aritake, K.; Huang, Z.; Strohl, K.P.; et al. Lipocalin-type prostaglandin D synthase levels increase in patients with narcolepsy and idiopathic hypersomnia. Sleep 2021, 44, zsaa234. [Google Scholar] [CrossRef] [PubMed]
- Fujimori, K.; Fukuhara, A.; Inui, T.; Allhorn, M. Prevention of paraquat-induced apoptosis in human neuronal SH-SY5Y cells by lipocalin-type prostaglandin D synthase. J. Neurochem. 2012, 120, 279–291. [Google Scholar] [CrossRef] [PubMed]
- Fukuhara, A.; Yamada, M.; Fujimori, K.; Miyamoto, Y.; Kusumoto, T.; Nakajima, H.; Inui, T. Lipocalin-type prostaglandin D synthase protects against oxidative stress-induced neuronal cell death. Biochem. J. 2012, 443, 75–84. [Google Scholar] [CrossRef] [Green Version]
- Choi, D.J.; An, J.; Jou, I.; Park, S.M.; Joe, E.H. A Parkinson’s disease gene, DJ-1, regulates anti-inflammatory roles of astrocytes through prostaglandin D2 synthase expression. Neurobiol. Dis. 2019, 127, 482–491. [Google Scholar] [CrossRef]
- Poewe, W.; Seppi, K.; Tanner, C.M.; Halliday, G.M.; Brundin, P.; Volkmann, J.; Schrag, A.E.; Lang, A.E. Parkinson disease. Nat. Rev. Dis. Primers 2017, 3, 17013. [Google Scholar] [CrossRef]
- Satoh, S.; Matsumura, H.; Suzuki, F.; Hayaishi, O. Promotion of sleep mediated by the A2a-adenosine receptor and possible involvement of this receptor in the sleep induced by prostaglandin D2 in rats. Proc. Natl. Acad. Sci. USA 1996, 93, 5980–5984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, B.J.; Huang, Z.L.; Chen, J.F.; Urade, Y.; Qu, W.M. Adenosine A2A receptor deficiency attenuates the somnogenic effect of prostaglandin D2 in mice. Acta Pharmacol. Sin. 2017, 38, 469–476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lazarus, M.; Chen, J.F.; Huang, Z.L.; Urade, Y.; Fredholm, B.B. Adenosine and Sleep. Handb. Exp. Pharm. 2019, 253, 359–381. [Google Scholar] [CrossRef]
- Negida, A.; Elfil, M.; Attia; Farahat, E.; Gabr, M.; Essam, A.; Attia, D.; Ahmed, H. Caffeine; the Forgotten Potential for Parkinson’s Disease. CNS Neurol. Disord. Drug Targets 2017, 16, 652–657. [Google Scholar] [CrossRef] [PubMed]
- Ren, X.; Chen, J.F. Caffeine and Parkinson’s Disease: Multiple Benefits and Emerging Mechanisms. Front. Neurosci. 2020, 14, 602697. [Google Scholar] [CrossRef] [PubMed]
- Borroto-Escuela, D.O.; Hinz, S.; Navarro, G.; Franco, R.; Muller, C.E.; Fuxe, K. Understanding the Role of Adenosine A2AR Heteroreceptor Complexes in Neurodegeneration and Neuroinflammation. Front. Neurosci. 2018, 12, 43. [Google Scholar] [CrossRef] [Green Version]
- Hu, Q.; Ren, X.; Liu, Y.; Li, Z.; Zhang, L.; Chen, X.; He, C.; Chen, J.F. Aberrant adenosine A2A receptor signaling contributes to neurodegeneration and cognitive impairments in a mouse model of synucleinopathy. Exp. Neurol. 2016, 283, 213–223. [Google Scholar] [CrossRef]
- Ferreira, D.G.; Batalha, V.L.; Vicente Miranda, H.; Coelho, J.E.; Gomes, R.; Goncalves, F.Q.; Real, J.I.; Rino, J.; Albino-Teixeira, A.; Cunha, R.A.; et al. Adenosine A2A Receptors Modulate alpha-Synuclein Aggregation and Toxicity. Cereb. Cortex 2017, 27, 718–730. [Google Scholar] [CrossRef] [Green Version]
- Pinna, A.; Serra, M.; Morelli, M.; Simola, N. Role of adenosine A2A receptors in motor control: Relevance to Parkinson’s disease and dyskinesia. J. Neural. Transm. 2018, 125, 1273–1286. [Google Scholar] [CrossRef]
- Fuxe, K.; Marcellino, D.; Borroto-Escuela, D.O.; Guescini, M.; Fernandez-Duenas, V.; Tanganelli, S.; Rivera, A.; Ciruela, F.; Agnati, L.F. Adenosine-dopamine interactions in the pathophysiology and treatment of CNS disorders. CNS Neurosci. Ther. 2010, 16, e18–e42. [Google Scholar] [CrossRef]
- Varani, K.; Vincenzi, F.; Tosi, A.; Gessi, S.; Casetta, I.; Granieri, G.; Fazio, P.; Leung, E.; MacLennan, S.; Granieri, E.; et al. A2A adenosine receptor overexpression and functionality, as well as TNF-alpha levels, correlate with motor symptoms in Parkinson’s disease. FASEB J. 2010, 24, 587–598. [Google Scholar] [CrossRef] [PubMed]
- Calon, F.; Dridi, M.; Hornykiewicz, O.; Bedard, P.J.; Rajput, A.H.; Di Paolo, T. Increased adenosine A2A receptors in the brain of Parkinson’s disease patients with dyskinesias. Brain 2004, 127, 1075–1084. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casetta, I.; Vincenzi, F.; Bencivelli, D.; Corciulo, C.; Gentile, M.; Granieri, E.; Borea, P.A.; Varani, K. A(2A) adenosine receptors and Parkinson’s disease severity. Acta Neurol. Scand. 2014, 129, 276–281. [Google Scholar] [CrossRef] [PubMed]
- Sen, N.; Gui, B.; Kumar, R. Physiological functions of MTA family of proteins. Cancer Metastasis Rev. 2014, 33, 869–877. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, D.Q.; Pakala, S.B.; Reddy, S.D.; Peng, S.; Balasenthil, S.; Deng, C.X.; Lee, C.C.; Rea, M.A.; Kumar, R. Metastasis-associated protein 1 is an integral component of the circadian molecular machinery. Nat. Commun. 2013, 4, 2545. [Google Scholar] [CrossRef] [Green Version]
- Reddy, S.D.; Rayala, S.K.; Ohshiro, K.; Pakala, S.B.; Kobori, N.; Dash, P.; Yun, S.; Qin, J.; O’Malley, B.W.; Kumar, R. Multiple coregulatory control of tyrosine hydroxylase gene transcription. Proc. Natl. Acad. Sci. USA 2011, 108, 4200–4205. [Google Scholar] [CrossRef] [Green Version]
- Kumar, A.S.; Jagadeeshan, S.; Subramanian, A.; Chidambaram, S.B.; Surabhi, R.P.; Singhal, M.; Bhoopalan, H.; Sekar, S.; Pitani, R.S.; Duvuru, P.; et al. Molecular Mechanism of Regulation of MTA1 Expression by Granulocyte Colony-stimulating Factor. J. Biol. Chem. 2016, 291, 12310–12321. [Google Scholar] [CrossRef] [Green Version]


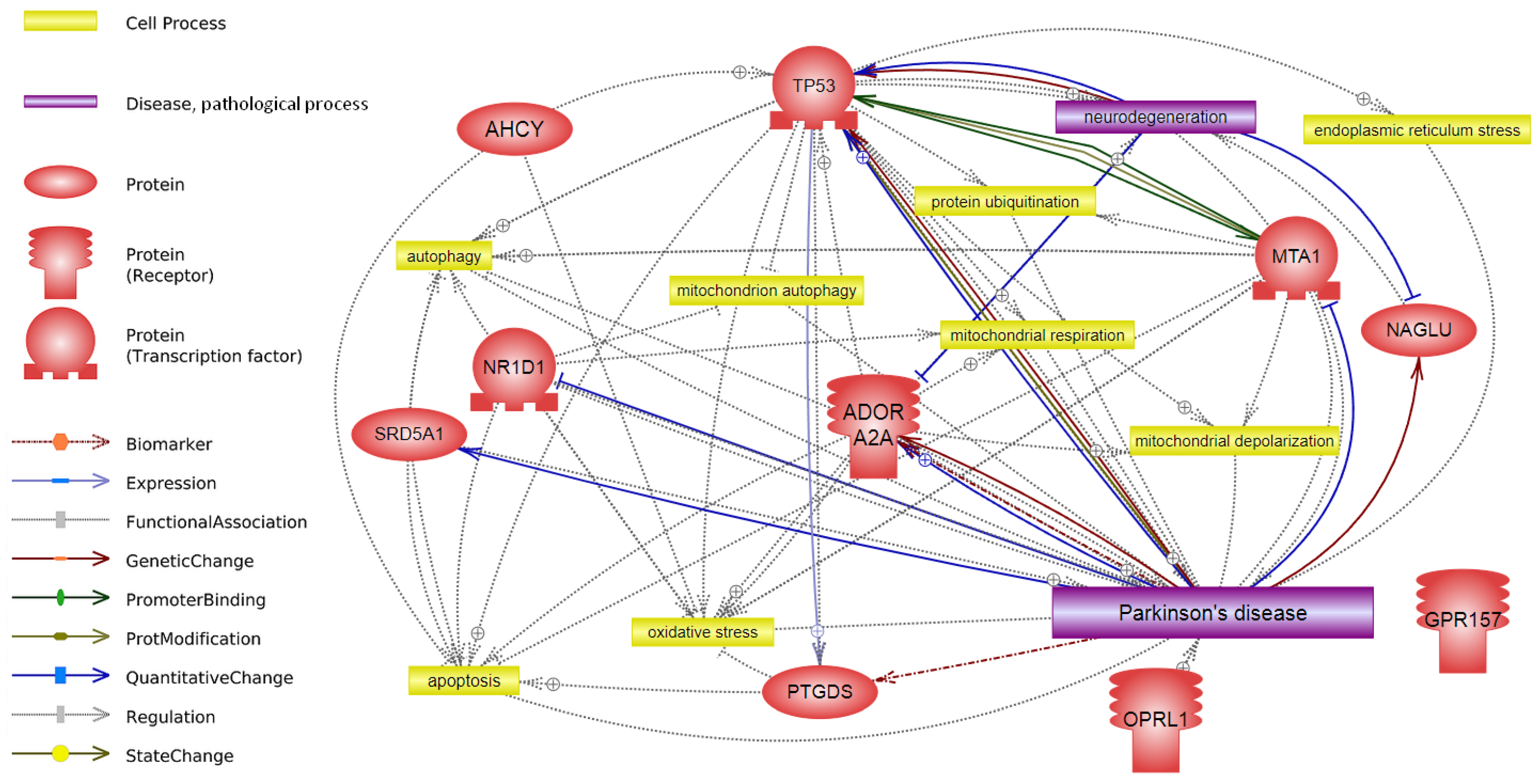{kind=link}
| Patient No. 1 | Patient No. 2 | Patient No. 3 | |
|---|---|---|---|
| Sex | Female | Female | Female |
| Year of birth | 1956 | 1947 | 1949 |
| Disease onset date | 2002 | 2007 | 2011 |
| PD stage (Hoehn–Yahr scale) | 2 | 4 | 3 |
| Motor symptoms: | |||
| Tremor | Yes | Yes | Yes |
| Rigidity | Yes | Yes | Yes |
| Bradykinesia | Yes | Yes | Yes |
| Postural instability | Yes | Yes | Yes |
| Mimic disorders | Hypomimia | Hypomimia | Hypomimia |
| Handwriting changes | Micrographia | - | - |
| Speech problems | Yes | - | - |
| Non-motor symptoms: | |||
| Olfactory dysfunction | Hyposmia | - | - |
| Sleep disturbances | - | Insomnia | Insomnia, nightmares |
| Emotional disturbances | - | Anxiety, depression | Anxiety |
| Cognitive deficits | - | Memory impairment, hallucinations | Memory impairment, hallucinations |
| Pain | Yes | Yes | - |
| Constipation | - | - | Yes |
| Urinary dysfunction | - | - | Yes |
| Gene | Nucleotide Sequence |
|---|---|
| SARS1 | Probe: 5′-VIC-TCGCCACTCGCTGTCTGCCTTCACCA-BHQ2-3′ |
| (Seryl-TRNA Synthetase 1) | Forward primer: 5′-CCCAGCCCTCATCCGAGAG-3′ |
| NM_001330669.1 * | Reverse primer: 5′-TGTTCAAGTTGTCTGCCCGAAATC-3′ |
| PSMD6 | Probe: 5′-VIC-AGGCGGTTTCTCCTGTCCCAGTCTCCTC-BHQ2-3′ |
| (Proteasome 26S Subunit, Non-ATPase 6) | Forward primer: 5′-AACACAGAAAAGGCCAAAAGCTTAAT-3′ |
| NM_001271779.1 * | Reverse primer: 5′-AATAGCCACACAATAAAGACCCTGAT-3′ |
| MTA1 | Probe: 5′-VIC-ATTTCCCCTTCCTCGCCGTTGTCCG-BHQ2-3′ |
| (Metastasis Associated 1) | Forward primer: 5′-ACATCTCCAGCACCCTCATCG-3′ |
| NM_004689.4 * | Reverse primer: 5′-TCGGGCAGGTCCACCATTT-3′ |
| TP53 | Probe: 5′-VIC-GTGTGGTGGTGCCCTATGAGCCG-BHQ2-3′ |
| (Tumor Protein P53) | Forward primer: 5′-GCGTGTGGAGTATTTGGATGAC-3′ |
| NM_001126114.2 * | Reverse primer: 5′-ATGTAGTTGTAGTGGATGGTGGTA-3′ |
| ADORA2A | Probe: 5′-VIC-AATGATGCCCTTAGCCCTCGTGCCG-BHQ2-3′ |
| (Adenosine A2a Receptor) | Forward primer: 5′-CATCGCCATTGACCGCTACA-3′ |
| NM_001278497.1 * | Reverse primer: 5′-GTTCCAACCTAGCATGGGAGTC-3′ |
| NR1D1 | Probe: 5′-VIC-GTGATGACGCCACCTGTGTTGTTGTTG-BHQ2-3′ |
| (Nuclear Receptor Subfamily 1 Group D Member 1) | Forward primer: 5′-CCAGTTTGAATGACCGCTCTCA-3′ |
| NM_021724.5 * | Reverse primer: 5′-GCTGCCATTGGAGTTGTCACTA-3′ |
| OPRL1 | Probe: 5′-VIC-GCTCCTGGGGAACTGCCTTGTCA-BHQ2-3′ |
| (Opioid Related Nociceptin Receptor 1) | Forward primer: 5′-CATCGTGGGGCTCTACCTG-3′ |
| NM_001318853.2 * | Reverse primer: 5′-ATTGGTGGCTGTCTTCATTTTGG-3′ |
| PTGDS | Probe: 5′-VIC-TTCACAGAGGATACCATTGTCTTCCTGCC-BHQ2-3′ |
| (Prostaglandin D2 Synthase) | Forward primer: 5′-GGAGAAATTCACCGCCTTCTG-3′ |
| NM_000954.6 * | Reverse primer: 5′-AGCCCTGGGGAGTCCTATT-3′ |
| AHCY | Probe: 5′-VIC-CATTGTGTGGATGCTGAAACTGAACCC-BHQ2-3′ |
| (Adenosylhomocysteinase) | Forward primer: 5′-TAGTTCATCAAGTTGCTACCAGAGT-3′ |
| NM_001322086.2 * | Reverse primer: 5′-TACCGCTCCCGCATACG-3′ |
| GPR157 | Probe: 5′-VIC-GCCTCGCACAGATCGCCTG-BHQ2-3′ |
| (G Protein-Coupled Receptor 157) | Forward primer: 5′-CTCTACTTGTACCTCAGCATCG-3′ |
| NM_024980.5 * | Reverse primer: 5′-GCGTCATAGCCAATCTTCTTCA-3′ |
| NAGLU | Probe: 5′-VIC-CGCTCCTTCGGCATGACCCCA-BHQ2-3′ |
| (N-Acetyl-Alpha-Glucosaminidase) | Forward primer: 5′-CCCCTCCTGGCACATCAAG-3′ |
| NM_000263.4 * | Reverse primer: 5′-GCCCATCTTCGTGACATTGAC-3′ |
| SRD5A1 | Probe: 5′-VIC-TTCCTCCTCGCATCAGAAATGGGT-BHQ2-3′ |
| (Steroid 5 Alpha-Reductase 1) | Forward primer: 5′-ATGGTCAGAATGGAAACAAATAACAAG-3′ |
| NM_001324323.2 * | Reverse primer: 5′-GCCGTTACAGGTACAGAACATAA-3′ |
| GO Term (GO ID) | Benjamini-Hochberg Adjustment p-Value of Hypergeometric Test for Enrichment | GO Group 1 | Number Of DEG, Associated with GO Group | Group p-Value Corrected with Benjamini-Hochberg | Percentage of Genes in Group, Associated with PD 2 |
|---|---|---|---|---|---|
| circadian behavior (GO:0048512) | 7.05 × 10−3 | I | 10 | 2.38 × 10−3 | 80.00 |
| negative regulation of glucose catabolic process to lactate via pyruvate (GO:1904024) | 5.15 × 10−3 | II | 6 | 2.10 × 10−3 | 50.00 |
| glucose catabolic process to lactate via pyruvate (GO:0019661) | 7.82 × 10−3 | ||||
| negative regulation of mitophagy (GO:1901525) | 7.82 × 10−3 | ||||
| lactate metabolic process (GO:0006089) | 8.56 × 10−3 | ||||
| response to testosterone (GO:0033574) | 5.13 × 10−3 | III | 12 | 4.33 × 10−3 | 41.66 |
| cellular response to testosterone stimulus (GO:0071394) | 9.62 × 10−3 | ||||
| protein transmembrane import into intracellular organelle (GO:0044743) | 8.25 × 10−3 | IV | 8 | 3.31 × 10−3 | 25.00 |
| positive regulation of ligase activity (GO:0051351) | 8.83 × 10−3 | V | 4 | 3.31 × 10−3 | 25.00 |
| regulation of triglyceride biosynthetic process (GO:0010866) | 7.91 × 10−3 | VI | 6 | 2.60 × 10−3 | 16.66 |
| regulation of ATP biosynthetic process (GO:2001169) | 8.61 × 10−3 | VII | 6 | 3.33 × 10−3 | 16.66 |
| cytoplasmic sequestering of protein (GO:0051220) | 5.19 × 10−3 | VIII | 7 | 1.95 × 10−3 | 14.29 |
| cytoplasmic sequestering of transcription factor (GO:0042994) | 8.56 × 10−3 | ||||
| branched-chain amino acid metabolic process (GO:0009081) | 7.39 × 10−3 | IX | 8 | 2.53 × 10−3 | 0 |
| leucine metabolic process (GO:0006551) | 7.46 × 10−3 |
| Genes | Peripheral Blood |
|---|---|
| ADORA2A | 1.811 1.34–2.162 |
| MTA1 | 1.54 1.05–1.86 |
| NR1D1 | 0.77 0.36–1.2 |
| OPRL1 | 1.19 1.08–2.08 |
| TP53 | 0.91 0.69–1.23 |
| PTGDS | 2.67 2.25–2.98 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Semenova, E.I.; Vlasov, I.N.; Partevian, S.A.; Rosinskaya, A.V.; Rybolovlev, I.N.; Slominsky, P.A.; Shadrina, M.I.; Alieva, A.K. Transcriptome Profiling Reveals Differential Expression of Circadian Behavior Genes in Peripheral Blood of Monozygotic Twins Discordant for Parkinson’s Disease. Cells 2022, 11, 2599. https://doi.org/10.3390/cells11162599
Semenova EI, Vlasov IN, Partevian SA, Rosinskaya AV, Rybolovlev IN, Slominsky PA, Shadrina MI, Alieva AK. Transcriptome Profiling Reveals Differential Expression of Circadian Behavior Genes in Peripheral Blood of Monozygotic Twins Discordant for Parkinson’s Disease. Cells. 2022; 11(16):2599. https://doi.org/10.3390/cells11162599
Chicago/Turabian StyleSemenova, Ekaterina I., Ivan N. Vlasov, Suzanna A. Partevian, Anna V. Rosinskaya, Ivan N. Rybolovlev, Petr A. Slominsky, Maria I. Shadrina, and Anelya Kh. Alieva. 2022. "Transcriptome Profiling Reveals Differential Expression of Circadian Behavior Genes in Peripheral Blood of Monozygotic Twins Discordant for Parkinson’s Disease" Cells 11, no. 16: 2599. https://doi.org/10.3390/cells11162599