3.1. Calculating the Required Dimensions for the Bioreactor Growth Chamber That Would Provide Sufficient Oxygen for Optimal Kidney Growth and Maintenance
Being able to fit the bioreactor unit into a tissue-culture incubator would eliminate the need for a media warmer, water-jacketed tubing and a second pump. Therefore, the diameter and depth measurement of the reservoir had practical limitations. The diameter and length of the reservoir collection tube used to draw the medium up and into circulation had to be considered so it would not disturb the air:liquid interface and generate air bubbles that could damage the kidney. The depth of the medium, the amount of exposed surface area and the time the medium is exposed to atmosphere air were important factors to consider. We empirically calculated the amount of medium required based on the minimal medium required to supply sufficient nutrients to the kidney over a 24-h period. This time period was chosen as it is optimal to change the media every 24 h. A mouse kidney contains approximately 100 million cells [
13]. Empirical data were achieved by using tissue-culture studies with adult primary renal epithelial cells (REC) grown in Corning cell stacks. One stack with a surface area of 636 cm
2 supported 10
9 cells and used 500 mL of medium for 24 h. We assumed a linear relationship between the cell number at confluence and volume of medium. Therefore, 100 million cells should be maintained with 50 mL medium/24 h. Based on this volume, we chose a bioreactor radius of 5 cm. This allows for a bioreactor that is small enough to easily handle and move in/out of an incubator and also provides for a large medium surface area to volume ratio.
To calculate the [O
2] at bottom of the reservoir we must consider that an oxygen gradient forms as oxygen diffuses to the base of the reservoir. Previous studies have assessed these internal gradients within cell culture media and often use Fick’s first law (Equation (1)) or derivations of Fick’s first law to describe oxygen consumption.
Fick’s first law describes the flux of oxygen and is based upon the presence of an oxygen source and an oxygen sink. In the referenced study, the [O2] in the gas acted as the oxygen source while cellular consumption of oxygen acted as the sink. These two variables were the driving force for the oxygen gradient. Due to the nature of our experimental setup, cellular consumption of oxygen was not considered a sink for the oxygen gradient in the reservoir as the organ or cells are not present in the reservoir. In addition, vascular perfusion of the whole kidney could not be equated to a monolayer of a single cell type. It was assumed that the only significant oxygen gradient would be in the acellular media reservoir and that a constant concentration of oxygen would be delivered to the kidney through the tubing. Following cellular consumption, the media would then have oxygen reinfused in the media reservoir. The recycled media in the reservoir created a transient system, and therefore the simplified version of Fick’s first law could not be applied to model the gradient.
Instead, a complimentary error function was derived to approximate oxygen flux (Equation (2)). Further information on this derivation can be reviewed elsewhere [
14]. We want to solve for
ykidney, the concentration of oxygen in the kidney.
Oxygen Concentration Approximation:
y
liquid is the molar concentration of oxygen at the liquid-gas interface. This is calculated from Henry’s law (Equation (3)), which states that the partial pressure of oxygen in the gas phase is proportional to the dissolved oxygen in the liquid medium (
yliquid) immediately below the interface.
Henry’s constant for oxygen diffusion into medium at standard incubator conditions (37 °C and 18.6% Oxygen) was previously determined [
15]. Since Henry’s constant (
H) is unaffected by air pressure differences less than 506 kPa [
14,
15] and the atmospheric pressure between our conditions and the study’s conditions are similar, a Henry’s constant of H = 771.65 mmHg/mM was used. To obtain
the following were used: Incubator conditions =18.6% oxygen, the average atmospheric pressure in Toronto, Canada = 753.66 mmHg, therefore
(partial pressure of oxygen (pO
2)) in the incubator air = 753.66 mmHg × 0.186= 140.18 mmHg. Therefore, using Henry’s law, the dissolved gas (y
liquid) just below the liquid interface = 140.18 mmHg/771.65 mmHg/mM = 0.181 mM = 5.8 ppm.
The height of the 50 mL volume was calculated from: Height of a Cylinder: .
This results in a total height of the medium of 0.667 cm. The collection tube is 0.267 cm from the bottom, therefore it is collecting medium from a depth of 0.40 cm.
x (medium depth) = 0.40 cm.
D (Diffusion Coefficient) =2.84 × 10−5 cm2/s.
H (Henry’s Constant) = Toronto Atmospheric Pressure: P = 100.48 kPa = 753.66 mmHg.
Incubator Oxygen Content (18.6%): pO2 = 132 mmHg @ 37 °C.
; = 132 mmHg/771.65 mmHg/mM = 122.4 mmHg oxygen.
The oxygenation at depth was calculated for two medium renewal rates. Medium renewal is the time it takes for all of medium in the reservoir to flow through the system once. ∆t = 2730 s is close to physiological renal flow rates for mouse and ∆t = 9091 s was calculated to determine if giving the medium more time in the reservoir increased oxygenation.
(a) Δt = 2730 s. and (b) ∆t = 9090 s.
Oxygen Concentration Approximation
)
- (a)
)
)
= 0.171 × 0.3086 = 0.0528 mM = 40 mmHg = 1.69 ppm
- (b)
)
)
= 0.171 × 0.3631 = 0.0621 mM = 47 mmHg = 1.98 ppm
Under physiologic conditions an oxygen gradient is formed between the blood and the tissue. The oxygen content in the blood is maintained at 0.130 mM, and this value then drops to 0.052 mM (40 mmHg = 1.69 ppm) at tissue level [
15]. The theoretical oxygen diffusion achieved by the bioreactor reservoir for both medium renewal rates falls within the range of physiological conditions at tissue level.
3.2. Bioreactor Assembly
The bioreactor consists of four primary components—the main chamber, the base lid, the kidney suspension plate, and the alignment cap (
Figure 1A–E). The main chamber is a flanged glass jar (
Figure 1(C1)) that houses the suspended kidney during long-term cell culture. The chamber has an inner diameter of 10 cm and a height of 10 cm. The lid of the chamber is assembled from three separate steel components. The first component contains the functional ports and is the base for the lid assembly (
Figure 1(C2)). The base lid includes a long tube to collect the recycled media from the reservoir, an opening for the kidney suspension plate and additional ports. Integrating a steel media tube prevents tube movement in the bioreactor, which prevents damage to the kidney. Previous iterations of the design involved a silicone collection tube within the chamber [
16]; however, the natural twisting of the tubing often damaged the kidney during assembly.
Each opening has embedded metal male luer locks so that the bioreactor is compatible with 3/32” tubing. The luer locks also act as a port site for media analysis and replenishment. The second component of the lid assembly is the circular kidney suspension plate (
Figure 1(C3)), a separate plate that only attaches to the kidney and allows for unobstructed addition/removal of the kidney. The suspension plate is 45 mm in diameter and has two sets of connected male luer locks. The top of the plate has two standard male luer locks that are 1.8 cm apart and then these standard locks connect to two custom swivel male luer ports below to create two ducts through the platform. The custom ports attach to the cannulas of the renal artery and vein or ureter. The swivel luer locks prevent twisting of the cannula and reduce damage to the kidney during handling. This suspension plate sits within a central ledge engraved in the base lid. The third component is the alignment piece (
Figure 1(C4)). It sits above both the base and suspension plate and aligns the components with the main glass chamber. The alignment piece seals the gap between the suspension plate and the base and provides a platform for clamping. The alignment plate is in two halves, which allows for removal without having to disconnect any of the tubing attached to the kidney. At each junction, an O-ring is inserted to create an air-tight seal. All components are clamped together with a ring clamp (
Figure 1E) to maintain an isolated system. The final setup, with the pump, is then placed into the incubator (
Figure 1F,G). The assembly of the bioreactor is performed in a biological safety cabinet to maintain sterile conditions.
3.3. Validation of the Mathematical Model
To confirm that the bioreactor could support an isolated kidney we attached cannulated mouse kidneys to the bioreactor and measured artery and venous oxygen, urine composition directly from the ureter and histo-pathology using H&E and IF. The higher protein content of the 10% FBS/DMEM medium was more suitable to measuring protein reabsorption than the REC medium, so 10% FBS/DMEM medium was used for all oxygen measurements and urine experiments. Histo-pathology was done on kidneys grown in both media. Media did not contain hemoglobin or oxygen carriers, so the oxygen calculations are applicable to both media. In order to obtain experimental values for oxygenation, reading of oxygen levels in the medium were taken with a Presens oxygen meter. To determine if the bioreactor could re-oxygenate the venous medium sufficiently, the renal artery of the ex vivo adult mouse kidney was cannulated and connected to the bioreactor with the oxygen sensor situated just before the renal artery-cannula connection.
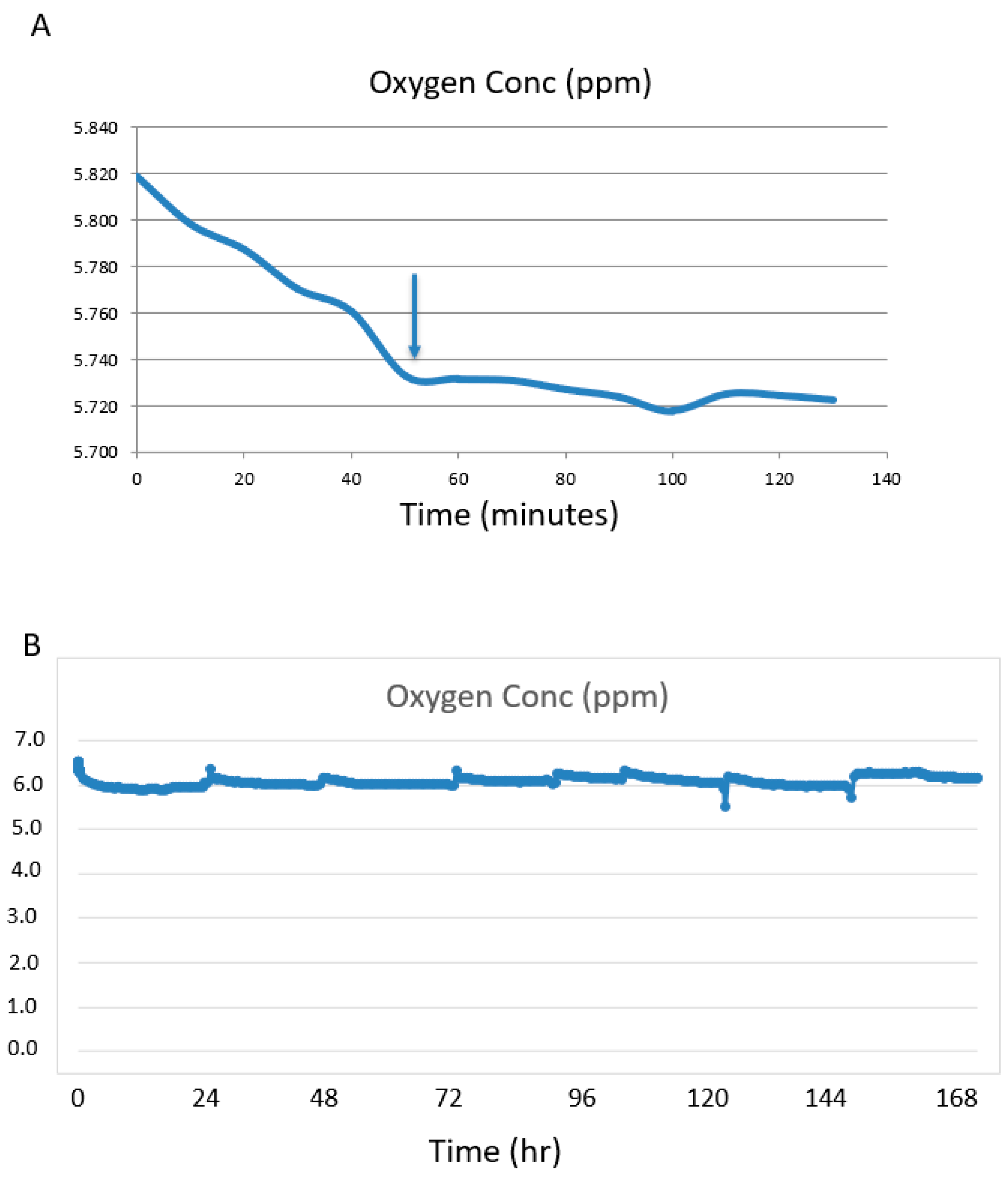
The flow rate was set at 1.1 mL/minute. The system was first run without a kidney attached to determine the time for the [O
2] to stabilize. The medium in the bottle stored at 4 °C was oxygenated as the oxygen level at time = 0 was 5.82 ppm (
Figure 2A). During the cycling period through the bioreactor at 37 °C the [O
2] stabilized a little lower at 5.74 ppm at ~50 min (
Figure 2A). This oxygen concentration was higher than the calculated amount of 2.0 ppm.
Silicone tubing is porous to oxygen and can act as an oxygenator. Increasing the length of the tubing between the pump and the bioreactor will increase the oxygen in the medium if saturation has not been reached and will decrease the time to saturation. In a series of non-kidney experiments we tested 1 m and 6 m of silicone tubing and compared this to 6 m of Tygon tubing. Tygon tubing was used as a control as it is not permeable to oxygen. There was no difference in [O2] for all parameters tested. The [O2] for silicone tubing was the same for 1 m and 6 m lengths and for the Tygon tubing (6 m), indicating oxygen saturation of the medium was achieved through diffusion of medium in the reservoir. For all tubing types and lengths, the medium oxygenation levels were ~5.7 ppm, so 1 m of silicone tubing was used for all experiments.
When the mouse kidney was attached to the system, the medium oxygen concentration held at a narrow range of between 5.7 ppm and 6.3 ppm for the 7 days (
Figure 2B). Fluctuations were observed during media changes or if the bioreactor was repositioned, as the sensor is sensitive to motion. These are seen as spikes at 24 h intervals—the times the media was changed.
To measure the oxygen consumption of the kidney, the venous medium was sampled and measured. The venous O2 was 4.4 ppm (4.2–4.6 ppm) for the duration of the experiments. Oxygen consumption was calculated and adjusted to ppm/mL/gram of kidney.
(OxygenArtery − OxygenVein)/mL/gram of kidney weight:
OxygenA: 5.73 ppm
OxygenV: 4.40 ppm
Flow rate (mL/min): 1.1
weight of kidney: 150 mg
=1.18 ppm/mL/0.150 g (per kidney)
=6.77 ppm/mL/gram (adjusted to gram of tissue).
The calculations determined that the bioreactor built to our calculated specifications and run near the physiological flow rate (1.1 mL/min) would provide sufficient medium oxygenation to meet the physiological tissue requirement of 40 mmHg = 1.7 ppm. Interestingly, the actual reading of [O2] in the medium near the bottom of the reservoir where the intake tube draws up the reservoir medium, at a flow rate of 1.1 mL/minute, was 5.73 ppm. This was about the same as the calculated amount for the [O2] at the medium surface (5.8 ppm). This could be explained by an unaccounted mixing of the medium at the bottom with surface medium as it is drawn up into the collection tube causing increased oxygenation.
Our goal was to achieve physiological [O2] levels in the medium. The experimental arterial reading was 5.73 ppm and exceeded the physiological oxygen levels of arterial blood at the tissue level in mice of 40 mmHg = 1.7 ppm. Importantly, our reading at the vein cannula was 4.2–4.6 ppm, which is above the minimum physiological requirement, indicating the kidney was well oxygenated.
3.4. Urine Analysis
Because the kidney is cultured ex vivo without the influence from other tissues or organs, the urine production is in the form of ultrafiltrate and does not contain all the components of normal urine. For example, there is no muscle in the system, so creatinine is not produced and therefore does not appear in the ultrafiltrate. We used 10%FBS/DMEM-high glucose as the medium for these experiments. This medium contains high levels of total protein, albumin and glucose, and thus we could test the efficiency of the kidney to reabsorb proteins and glucose better than with REC medium, which contains lower amounts of each of these components.
If the ex vivo perfused kidneys were functioning properly, it was expected that there would be a decrease in the amount of protein and glucose from the fresh medium (t = 0) compared to perfusion medium at 20 h. Fresh 10%FBS/DMEM/high glucose medium contained 386.82 mg/dL of total protein and 3.23 mg/dL of albumin. The reservoir medium after 20 h of circulation through the kidney contained 305.55 mg/dL of total protein and 2.87 mg/dL of albumin, indicating the kidney consumed nutrients as expected.
Furthermore, if the ex vivo kidney was functioning properly the functioning glomeruli would filter out the protein followed by reabsorption of the proteins by the proximal tubules. This would result in a low protein concentration in the urine compared to the medium. It is also expected that the high glucose in the medium would result in the movement of glucose from the perfusion medium to the ultrafiltrate being produced by the glomeruli, but a properly functioning kidney should then reabsorb the glucose so the excreted urine glucose levels would remain low. For urine analysis the ureter was cannulated and 0.15 mL of fluid was collected at day 7 of ex vivo perfusion. The perfusion medium, acting as the ‘blood’ (control 2), is considered input and was collected at the same time urine was collected from the ureter cannula. The medium was changed every 24 h, and the input perfusion medium and ureter ultrafiltrate were collected ~20 h post medium change at day 7 (
Table 1). The reservoir medium at the time of urine collection (t = 20 h) had 305.55 mg/dL of total protein and 2.87 mg/dL of albumin while the total protein concentration in the ultrafiltrate collected from the ureter at the same time contained 18.76 mg/dL of total protein (range 9.68–27.84) and 0.06 mg/dL of albumin (range 0.01–0.11), indicating that the majority of the protein was reabsorbed as expected for a functioning kidney (
Table 1). Glucose levels were measured at the same time protein was measured. Glucose is freely filtered by the glomeruli but is reabsorbed in the tubules by SGLT1 and SGLT2 [
17]. At twenty hours post medium renewal on day 7, the circulating medium contained 399.78 mg/dL of glucose. In the ultrafiltrate the glucose was 4.08 mg/dL, about 1% of the levels in the perfusion medium, indicating the kidney was functioning correctly and reabsorbed the excess glucose accumulating in the ultrafiltrate.
3.5. Physical Characterization of the Bioreactor Grown Kidneys
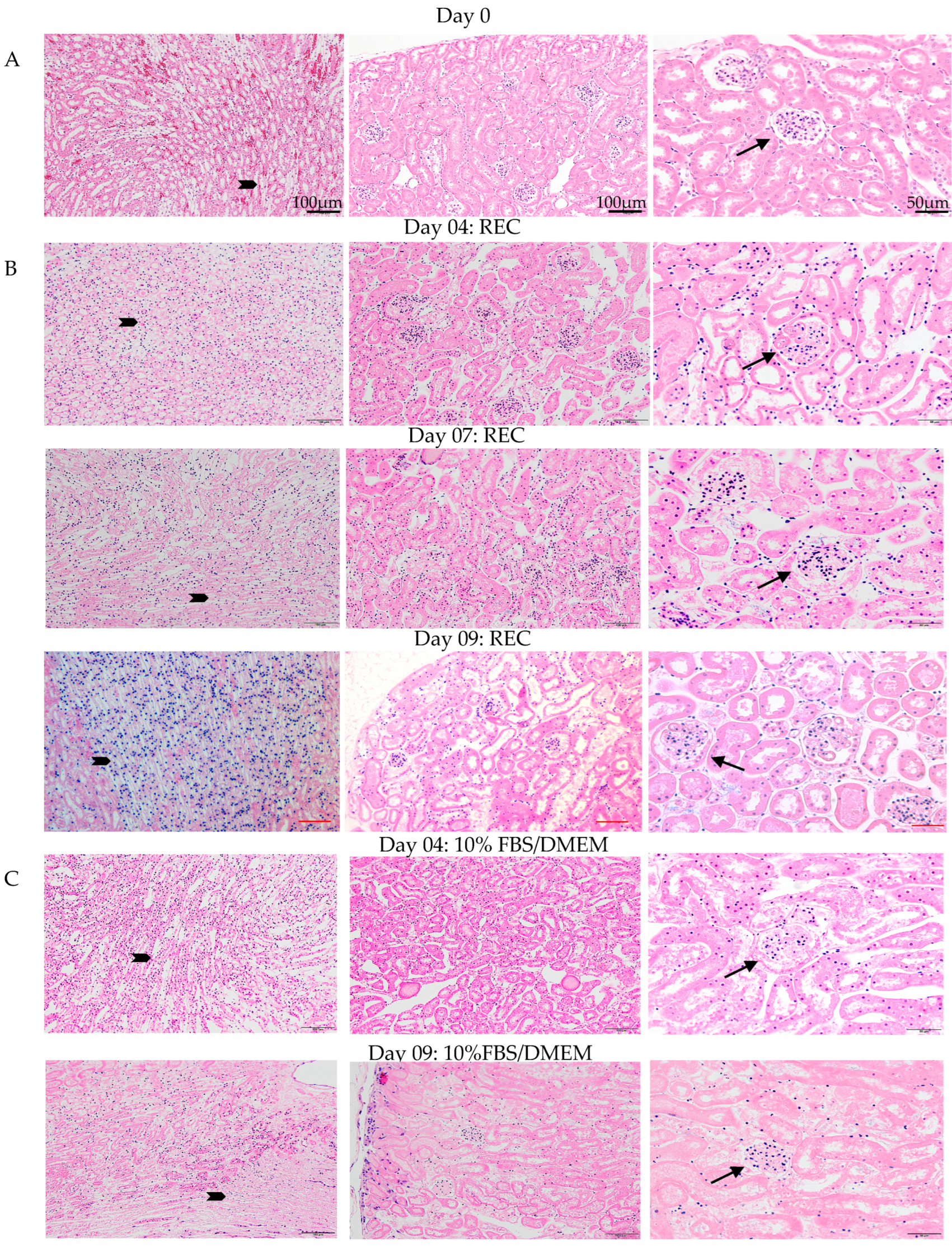
Oxygenation levels of the medium during ex vivo perfusion exceeded physiological levels, and urine analysis indicated that the kidney is healthy and functioning. Histological examination of the kidneys by H&E and immunofluorescence (IF) was carried out to confirm the physical integrity of the kidney post ex vivo perfusion. Ex vivo perfusion used either 10%FBS/DMEM or specialized renal epithelial cell (REC) media. H&E staining was done to assess kidney morphology (
Figure 3). Day 0 control kidneys were fixed and processed directly after cannulation. All three time points for kidneys grown in REC medium (day 4, 7, 9) have a similar morphology to that of the day 0 controls. The cells look healthy with healthy nuclei and no signs of oedema or necrosis. The day 4 10%FBS/DMEM grown kidney also resembled the day 0 control kidney. The kidney grown for 9 days in 10%FBS/DMEM had good morphology but was showing signs that tissue degradation was beginning as fewer healthy nuclei were observed in the tubules. Representative magnification bars are in the first panel for each column (
Figure 3).
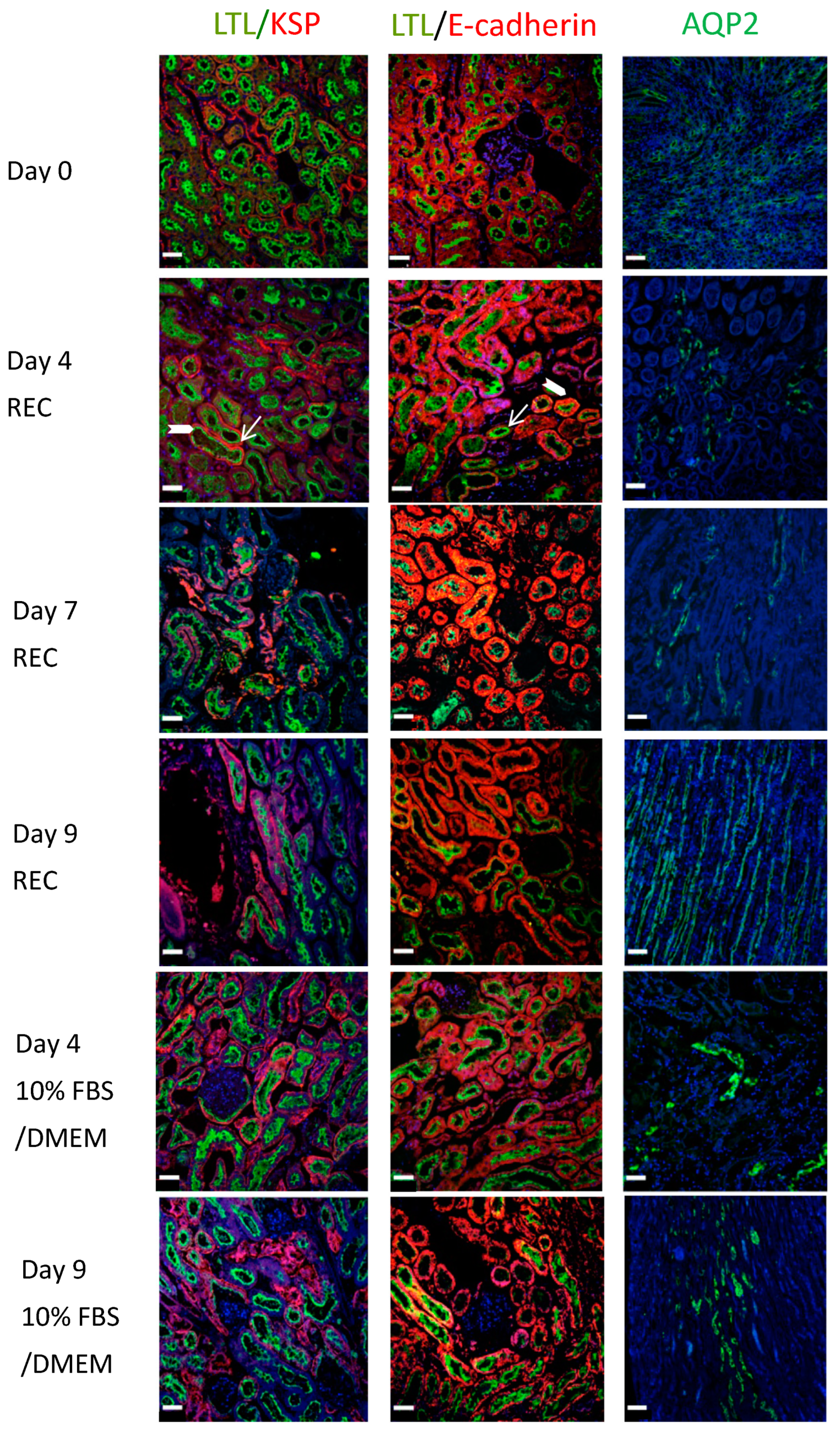
Renal sections were also co-stained for LTL (green) and E-cadherin (red). E-cadherin surrounds the tubule cells (
Figure 4, block arrow: Day 4 REC: LTL/E-cadherin) while LTL is mainly on the apical side, seen here located as an inner ring in cross-sections of tubules (
Figure 4, arrow: Day 4 REC: LTL/E-cadherin). E-cadherin is abundant in the distal renal tubules and its expression is low in the proximal renal tubules. Its co-expression with LTL depicts proximal tubule cells. AQP2 (green) highlights collecting ducts. The ducts are more numerous in the medullary region (
Figure 4: Day 9 REC: AQP2) compared to the cortical regions (
Figure 4: Day 4 DMEM: AQP2 and Day 7 REC: AQP2). All data points demonstrate strong staining and excellent tissue morphology. Despite the onset of tissue decline observed with H&E staining of the day 09 kidney perfused with 10%FBS/DMEM. The IF staining looked similar to the day 0 controls for all proteins.
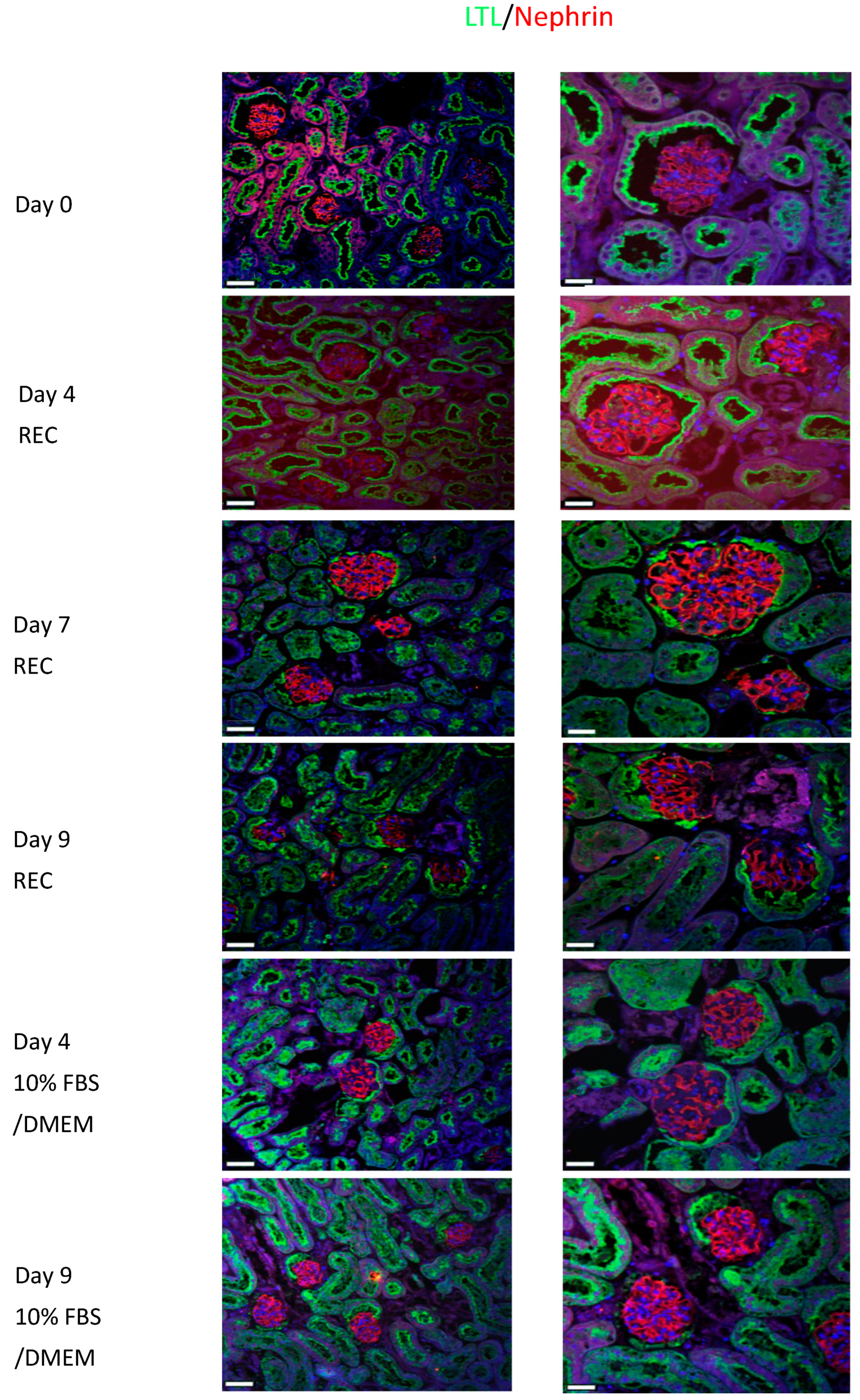
Nephrin (red) is localized to the podocytes and has a distinct staining pattern that can be clearly observed in the glomerulus of the day 0 control kidneys (
Figure 5). All REC and 10%FBS/DMEM cultured kidneys show similar staining to the control kidney. LTL (green) is used as a counter stain. Nephrin is important for maintaining the slit diaphragm and it is important for proper kidney function.

 ,
, 





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}