
Fine-Mapping Analysis of the Genes Associated with Pre-Harvest Sprouting Tolerance in Rice (Oryza sativa L.)
 and
and
Abstract
:1. Introduction
2. Materials and Methods
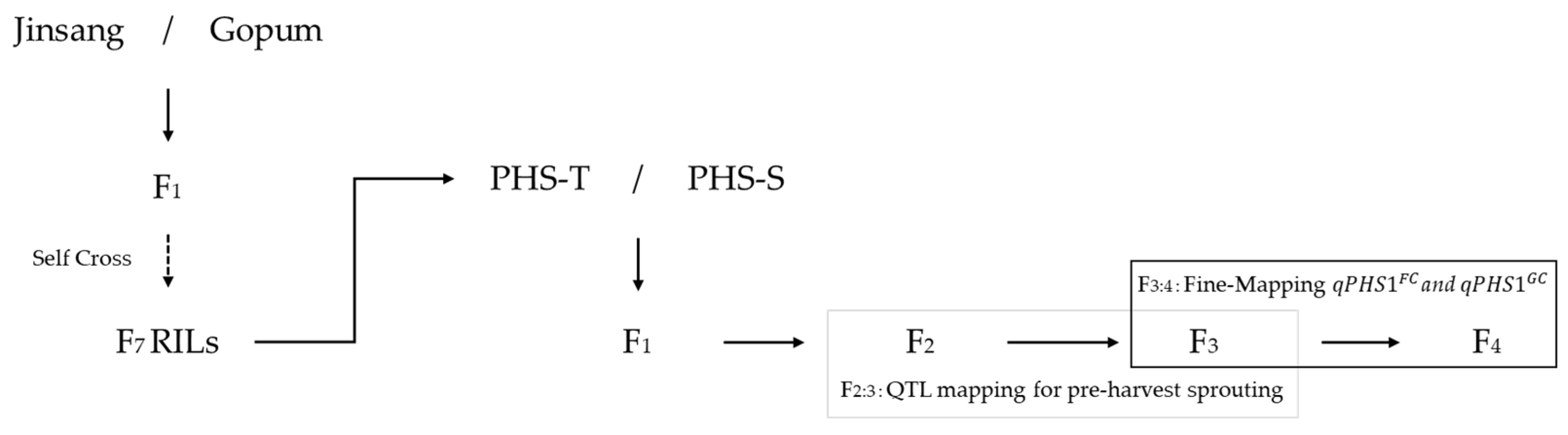
2.1. Plant Materials
2.2. Phenotypic Evaluation of Pre-Harvest Sprouting
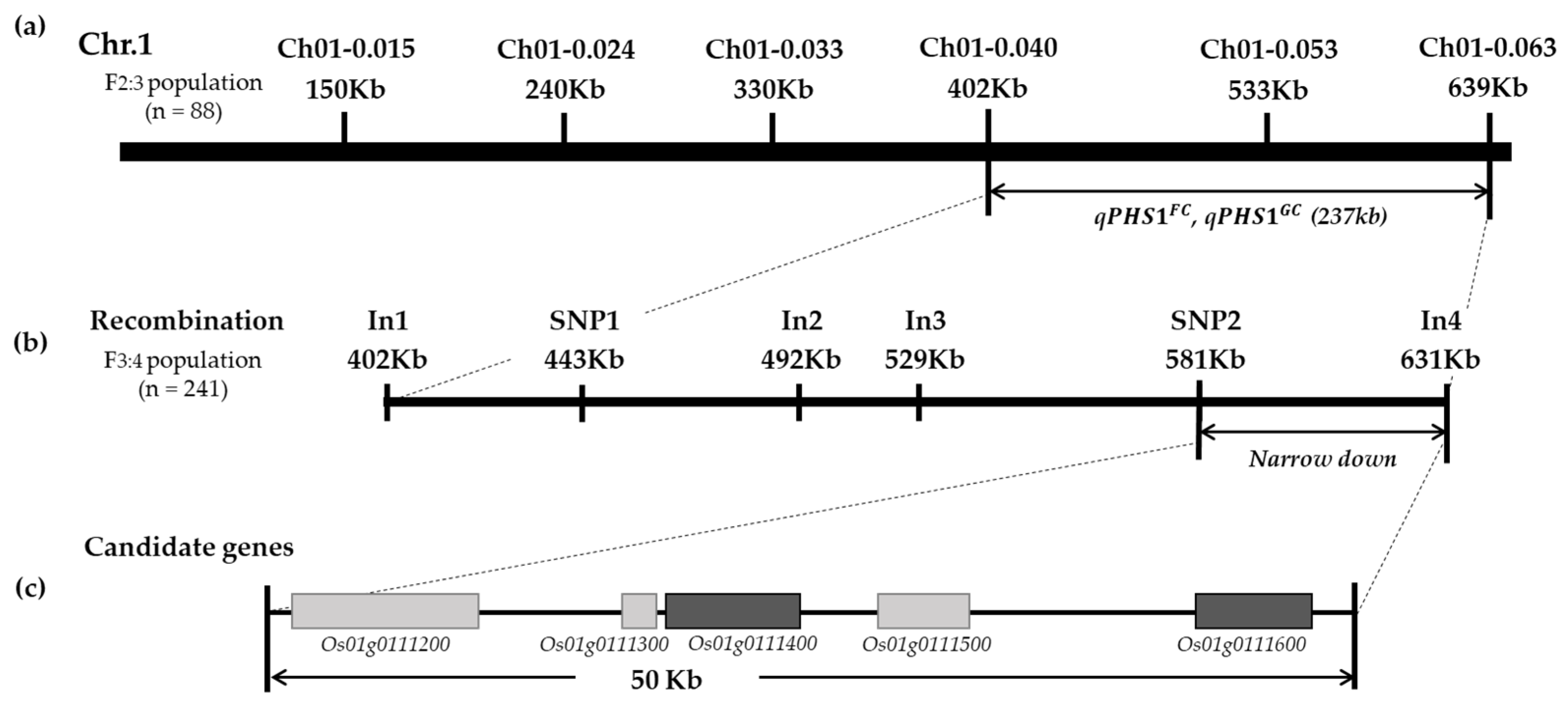
2.3. Fine-Mapping of qPHS1GC and qPHS1FC
2.4. RNA Extraction and qRT-PCR
2.5. Genome Sequence Data for Korean-Bred Rice Varieties
2.6. Haplotype Analysis for Korean-Bred Varieties
2.7. Seed Germination Assay and ABA Treatment
3. Results
3.1. Fine-Mapping of qPHS1FC and qPHS1GC
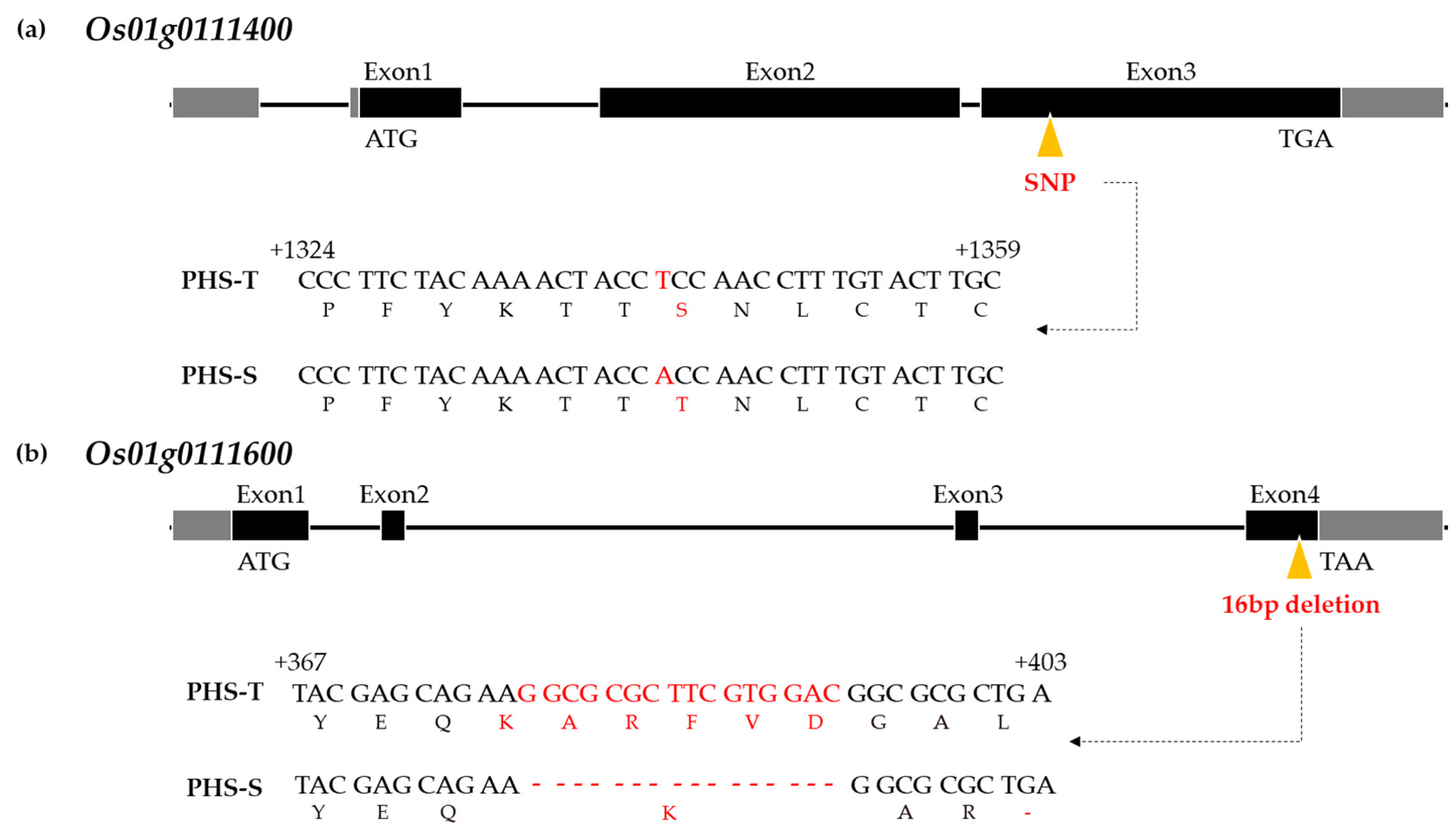
3.2. qPHS1FC and qPHS1GC Candidate Gene Prediction
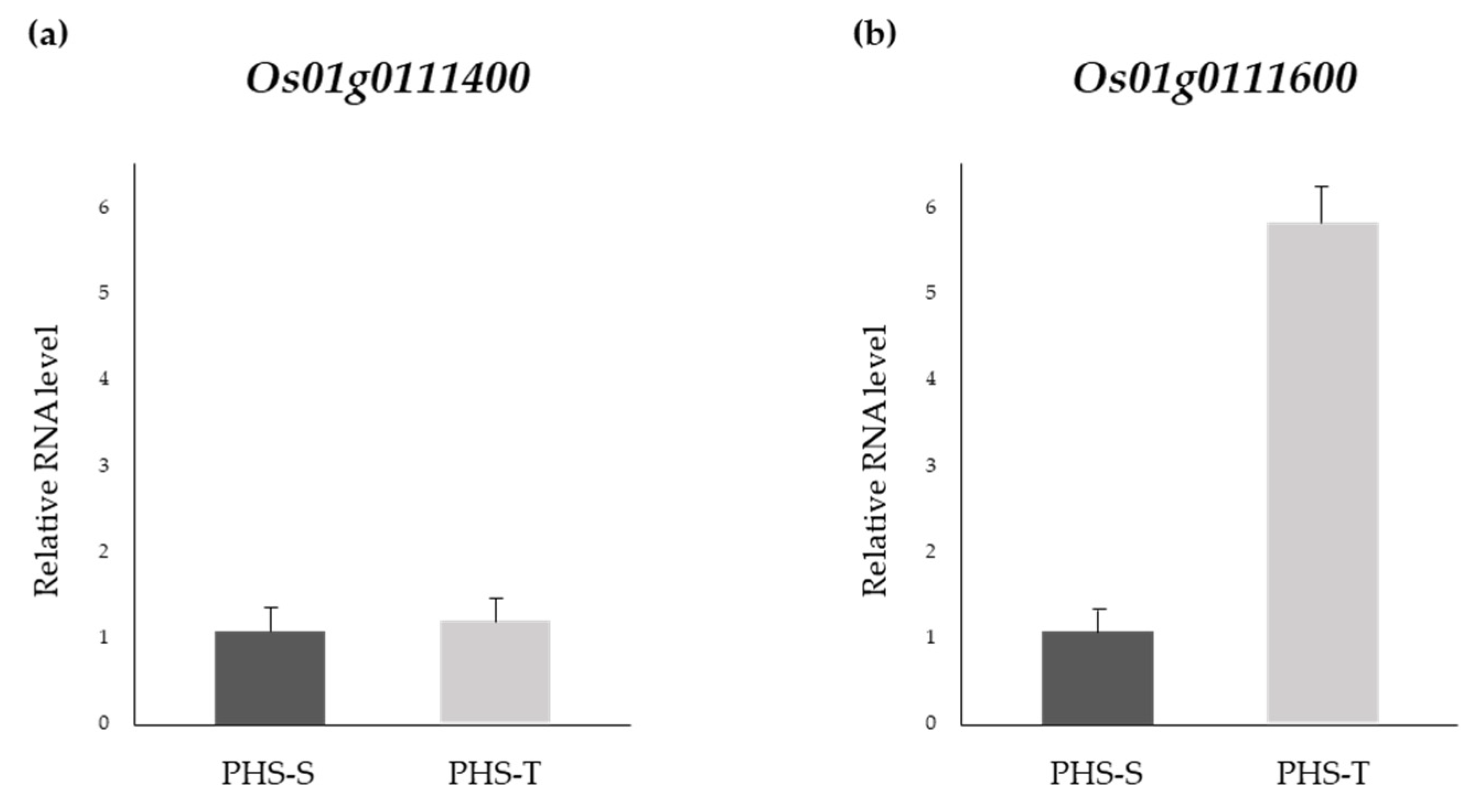
3.3. Expression Patterns of Os01g0111400 and Os01g0111600
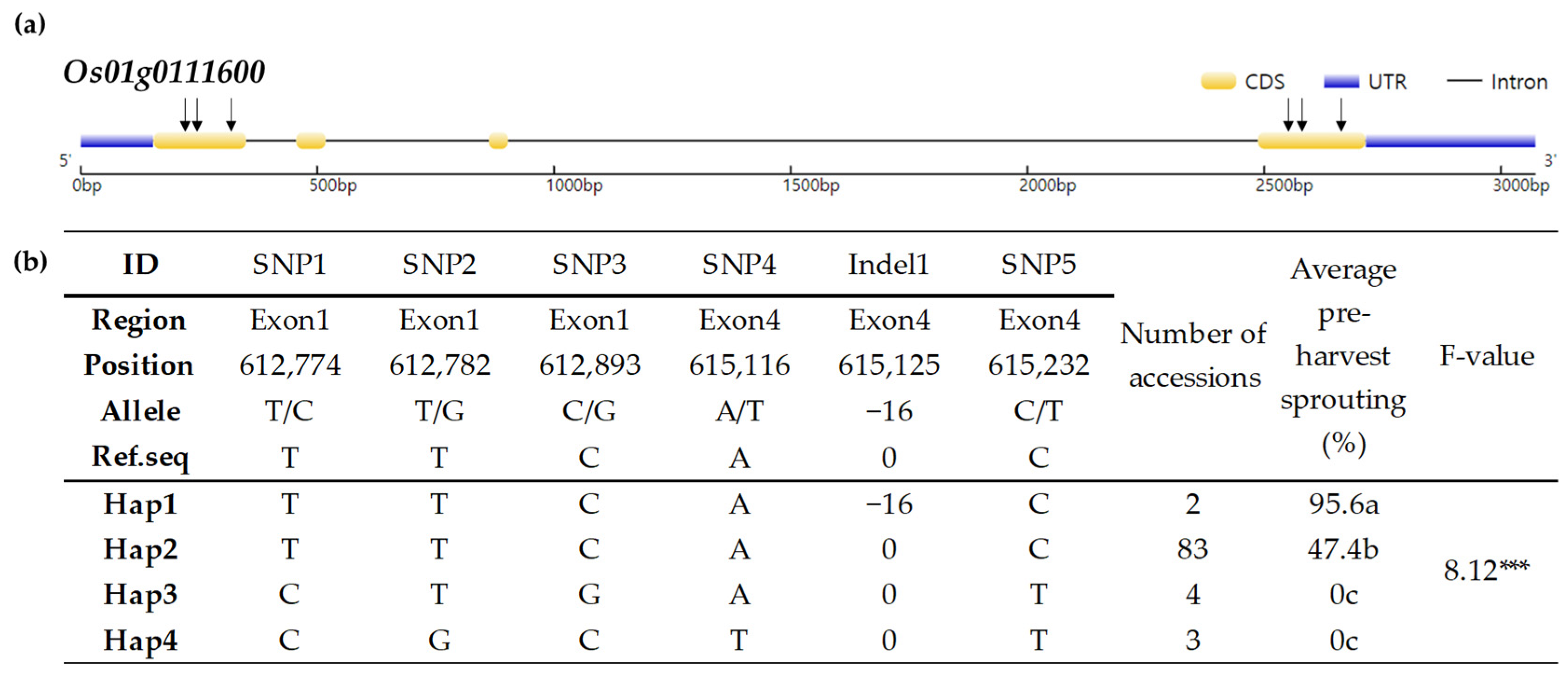
3.4. Haplotype Analysis of Os01g0111600 in Korean-Bred Varieties
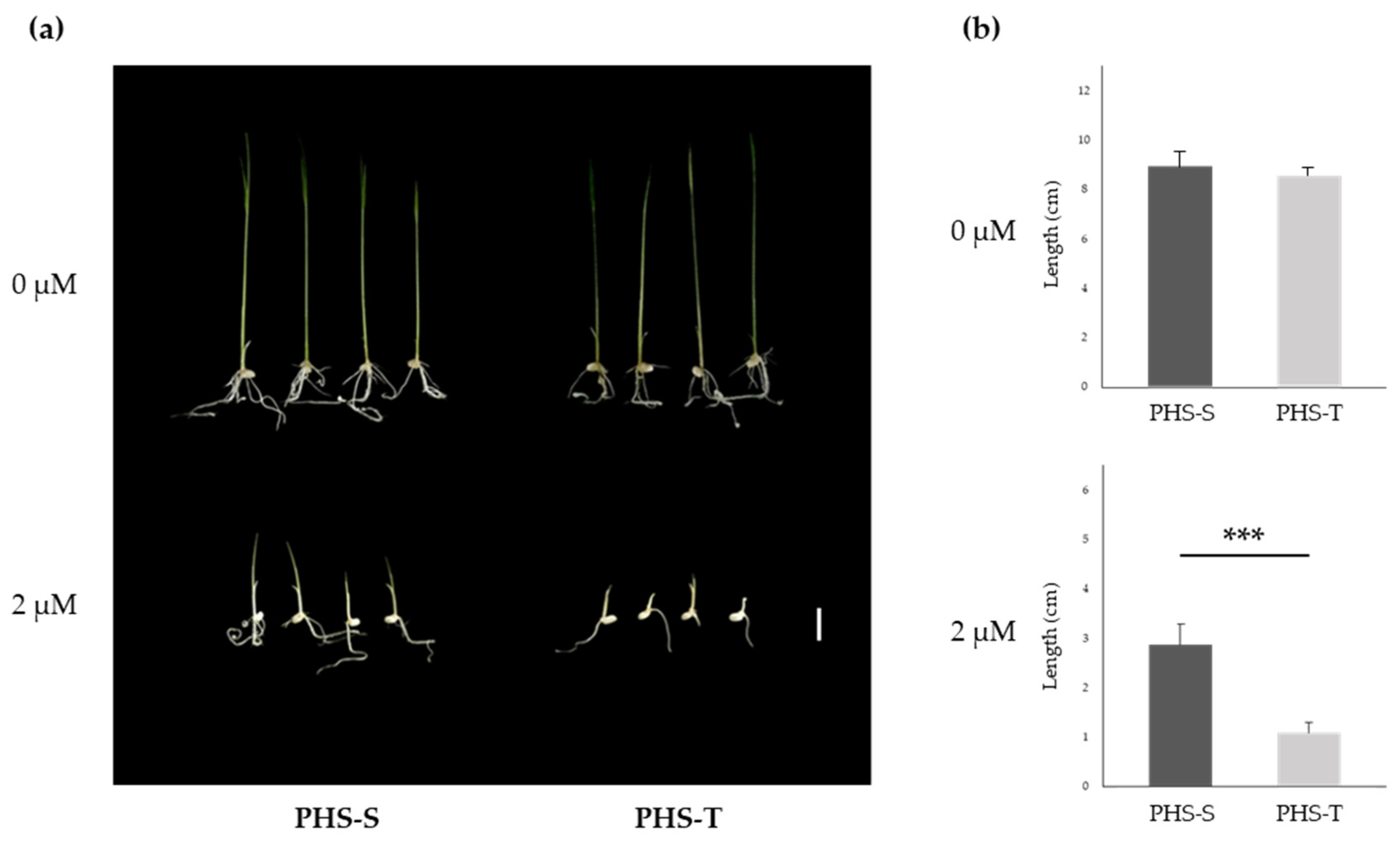
3.5. Response to ABA Treatment of PHS-S and PHS-T
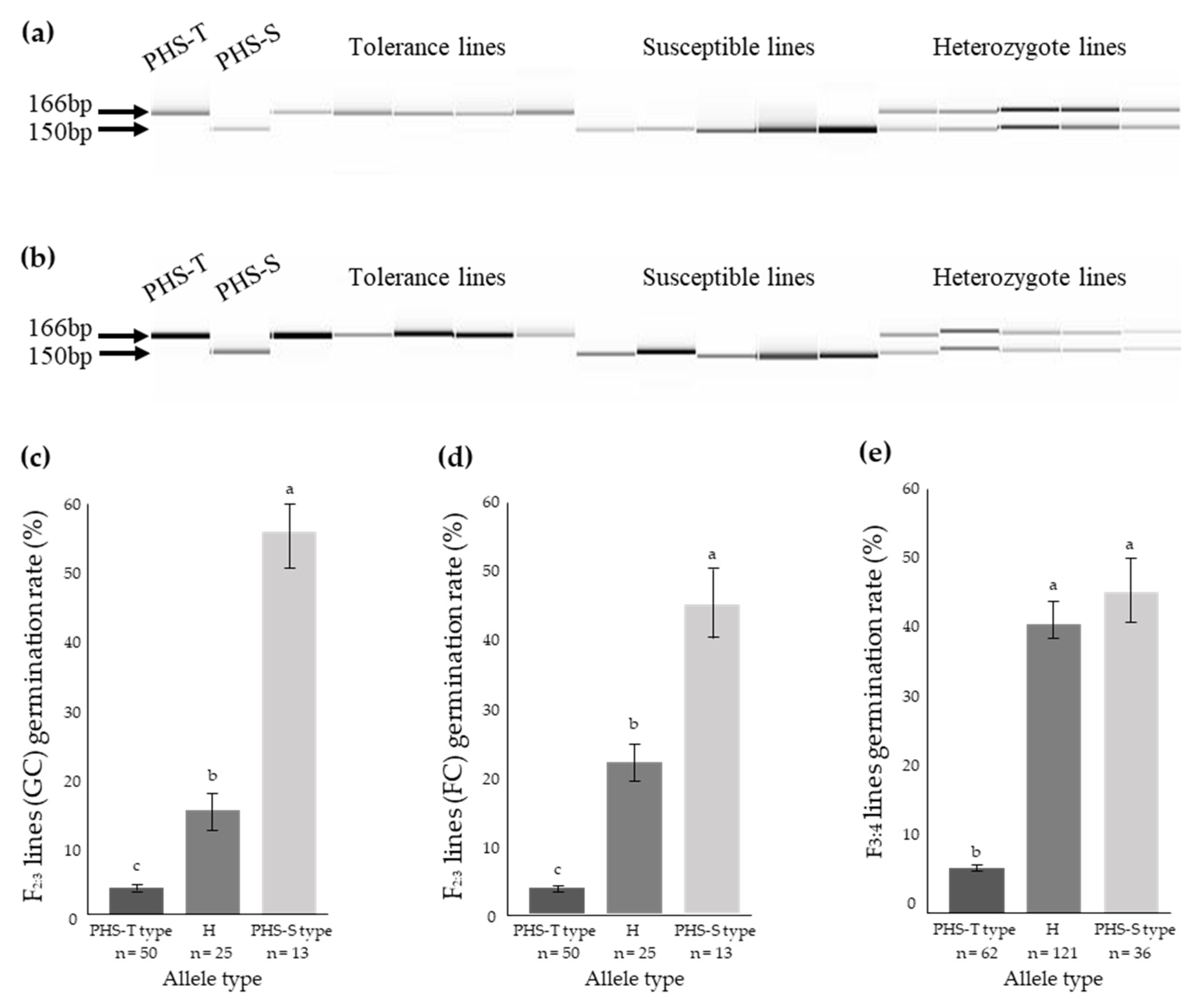
3.6. Co-Segregation between Indel Markers of Os01g0111600 and the PHS Trait
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gnanamanickam, S.S. Rice and its importance to human life. In Biological Control of Rice Diseases; Progress in Biological Control; Springer Science & Business Media: New York, NY, USA, 2009; Volume 8, pp. 1–11. [Google Scholar]
- Lenaerts, B.; Collard, B.C.; Mey, Y.; Demont, M. Global survey data on rice breeders’ characteristics and willingness to adopt alternative breeding methods. Data Brief 2019, 23, 103782. [Google Scholar]
- Miao, X.L.; Zhang, Y.J.; Xia, X.C.; He, Z.H.; Zhang, Y.; Yan, J.; Chen, X.M. Mapping quantitative trait loci for pre-harvest sprouting resistance in white-grained winter wheat line CA 0431. Crop Pasture Sci. 2013, 64, 573–579. [Google Scholar]
- Fang, J.; Chu, C. Abscisic acid and the pre-harvest sprouting in cereals. Plant Signal. Behav. 2008, 3, 1046–1048. [Google Scholar] [PubMed] [Green Version]
- Gubler, F.; Millar, A.A.; Jacobsen, J.V. Dormancy release ABA and pre-harvest sprouting. Curr. Opin. Plant Biol. 2005, 8, 183–187. [Google Scholar] [PubMed]
- Chono, M.; Honda, I.; Shinoda, S.; Kushiro, T.; Kamiya, Y.; Nambara, E.; Kawakami, N.; Kaneko, S.; Watanabe, Y. Field studies on the regulation of abscisic acid content and germinability during grain development of barley: Molecular and chemical analysis of pre-harvest sprouting. J. Exp. Bot. 2006, 57, 2421–2434. [Google Scholar] [CrossRef]
- Wan, J.M.; Jiang, L.; Tang, J.Y.; Wang, C.M.; Hou, M.Y.; Jing, W.; Zhang, L.X. Genetic dissection of the seed dormancy trait in cultivated rice (Oryza sativa L.). Plant Sci. 2006, 170, 786–792. [Google Scholar]
- Tejakhod, S.; Ellis, R.H. Effect of simulated flooding during rice seed development and maturation on subsequent seed quality. Seed Sci. Res. 2008, 28, 72–81. [Google Scholar]
- Xu, F.; Tang, J.; Gao, S.; Cheng, X.; Du, L.; Chu, C. Control of rice pre-harvest sprouting by glutaredoxin-mediated abscisic acid signaling. Plant J. 2019, 100, 1036–1051. [Google Scholar]
- Sohn, S.I.; Pandian, S.; Kumar, T.S.; Zoclanclounon, Y.A.B.; Muthuramalingam, P.; Shilpha, J.; Satish, L.; Ramesh, M. Seed Dormancy and Pre-Harvest Sprouting in Rice—An Updated Overview. Int. J. Mol. Sci. 2021, 22, 11804. [Google Scholar]
- Wu, M.; Wu, J.; Gan, Y. The new insight of auxin functions: Transition from seed dormancy to germination and floral opening in plants. Plant Growth Regul. 2020, 91, 169–174. [Google Scholar]
- Du, L.; Xu, F.; Fang, J.; Gao, S.P.; Tang, J.Y.; Fang, S.; Wang, H.; Tong, H.; Zhang, F.; Chu, J.; et al. Endosperm sugar accumulation caused by mutation of PHS8/ISA1 leads to pre-harvest sprouting in rice. Plant J. 2018, 95, 545–556. [Google Scholar] [PubMed] [Green Version]
- Debeaujon, I.; Koornneef, M. Gibberellin requirement for Arabidopsis seed germination is determined both by testa characteristics and embryonic abscisic acid. Plant Physiol. 2000, 122, 415–424. [Google Scholar] [PubMed] [Green Version]
- Li, B.; Foley, M.E. Genetic and molecular control of seed dormancy. Trends Plant Sci. 1997, 2, 384–389. [Google Scholar]
- Gu, X.Y.; Chen, Z.X.; Foley, M.E. Inheritance of seed dormancy in weedy rice. Crop Sci. 2003, 43, 835–843. [Google Scholar]
- Chen, C.X.; Cai, S.B.; Bai, G.H. A major QTL controlling seed dormancy and pre-harvest sprouting resistance on chromosome 4A in a Chinese wheat landrace. Mol. Breed. 2008, 21, 351–358. [Google Scholar]
- Gu, X.Y.; Foley, M.E.; Horvath, D.P.; Anderson, J.V.; Feng, J.; Zhang, L.; Mowry, C.R.; Ye, H.; Suttle, J.C.; Kadowaki, K.I.; et al. Association between seed dormancy and pericarp color is controlled by a pleiotropic gene that regulates abscisic acid and flavonoid synthesis in weedy red rice. Genetics 2011, 189, 1515–1524. [Google Scholar]
- Graeber, K.; Nakabayashi, K.; Miatton, E.; Leubner-Metzger, G.; Soppe, W.J. Molecular mechanisms of seed dormancy. Plant Cell Environ. 2012, 35, 1769–1786. [Google Scholar]
- Gao, F.Y.; Ren, G.J.; Lu, X.J.; Sun, S.X.; Li, H.J.; Gao, Y.M.; Luo, H.; Yan, W.G.; Zhang, Y.Z. QTL analysis for resistance to preharvest sprouting in rice (Oryza sativa). Plant Breed. 2008, 127, 268–273. [Google Scholar]
- Hori, K.; Sugimoto, K.; Nonoue, Y.; Ono, N.; Matsubara, K.; Yamanouchi, U.; Abe, A.; Takeuchi, Y.; Yano, M. Detection of quantitative trait loci controlling pre-harvest sprouting resistance by using backcrossed populations of japonica rice cultivars. Theor. Appl. Genet. 2010, 120, 1547–1557. [Google Scholar]
- Li, W.; Xu, L.; Bai, X.; Xing, Y. Quantitative trait loci for seed dormancy in rice. Euphytica 2011, 178, 427–435. [Google Scholar]
- Cai, H.W.; Morishima, H. Genomic regions affecting seed shattering and seed dormancy in rice. Theor. Appl. Genet. 2000, 100, 840–846. [Google Scholar]
- Gu, X.Y.; Kianian, S.F.; Foley, M.E. Multiple loci and epistases control genetic variation for seed dormancy in weedy rice (Oryza sativa). Genetics 2004, 66, 1503–1516. [Google Scholar]
- Cheon, K.S.; Won, Y.J.; Jeong, Y.M.; Lee, Y.Y.; Kang, D.Y.; Oh, J.; Oh, H.; Kim, S.L.; Kim, N.; Lee, E.; et al. QTL mapping for pre-harvest sprouting resistance in japonica rice varieties utilizing genome resequencing. Mol Genet Genom. 2020, 295, 1129–1140. [Google Scholar]
- Mizuno, Y.; Yamanouchi, U.; Hoshino, T.; Nonoue, Y.; Nagata, K.; Fukuoka, S.; Ando, T.; Yano, M.; Sugimoto, K. Genetic dissection of pre-harvest sprouting resistance in an upland rice cultivar. Breed Sci. 2018, 68, 200–209. [Google Scholar] [PubMed] [Green Version]
- Sugimoto, K.; Takeuchi, Y.; Ebana, K.; Miyao, A.; Hirochika, H.; Hara, N.; Ishiyama, K.; Kobayashi, M.; Ban, Y.; Hattori, T.; et al. Molecular cloning of Sdr4 a regulator involved in seed dormancy and domestication of rice. Proc Natl Acad Sci. USA 2010, 107, 5792–5797. [Google Scholar]
- Ye, H.; Feng, J.; Zhang, L.; Zhang, J.; Mispan, M.S.; Cao, Z.; Beighley, D.H.; Yang, J.; Gu, X.Y. Map-based cloning of seed dormancy1-2 identified a gibberellin synthesis gene regulating the development of endosperm-imposed dormancy in rice. Plant Physiol. 2015, 169, 2152–2165. [Google Scholar]
- Lee, J.S.; Chebotarov, D.; McNally, K.L.; Pede, V.; Setiyono, T.D.; Raquid, R.; Hyun, W.J.; Jeung, J.U.; Kohil, A.; Mo, Y. Novel sources of pre-harvest sprouting resistance for japonica rice improvement. Plants 2021, 10, 1709. [Google Scholar]
- Zhang, Y.; Miao, X.; Xia, X.; He, Z. Cloning of seed dormancy genes (TaSdr) associated with tolerance to pre-harvest sprouting in common wheat and development of a functional marker. Theor. Appl. Genet. 2014, 127, 855–866. [Google Scholar]
- Hong, W.J.; Yoon, M.Y.; Ra, W.H.; Park, J.W.; Min, S.K.; Kwon, S.W.; Cho, Y.H.; Park, Y.A. New variety ‘Jinsang’ with lodging tolerant and low amylose content. Korean J. Breed. Sci. 2014, 46, 103–107. [Google Scholar]
- Kang, S.; Shon, J.; Kim, H.S.; Kim, S.J.; Choi, J.S.; Park, J.H.; Yun, Y.H.; Sim, J.; Yang, W. Analysis of genetic variation in pre-harvest sprouting at different cumulative temperatures after heading of rice. Korean J. Crop Sci. 2018, 63, 8–17. [Google Scholar]
- Jang, S.G.; Zhang, H.; Lee, A.R.; Lee, J.H.; Kim, N.E.; Park, S.Y.; Lee, J.; Ham, T.H.; Kwon, S.W. Detection of Whole-Genome Resequencing-Based QTLs Associated with Pre-Harvest Sprouting in Rice (Oryza sativa L). Plant Breed. Biotechnol. 2020, 8, 396–404. [Google Scholar] [CrossRef]
- Porebski, S.; Bailey, L.G.; Baum, B.R. Modification of a CTAB DNA extraction protocol for plants containing high polysaccharide and polyphenol components. Plant. Mol. Biol. Rep. 1997, 15, 8–15. [Google Scholar] [CrossRef]
- Untergasser, A.; Cutcutache, I.; Koressaar, T.; Ye, J.; Faircloth, B.C.; Remm, M.; Rozen, S.G. Primer3—New capabilities and interfaces. Nucleic Acids Res. 2012, 40, 115. [Google Scholar]
- Purcell, S.; Neale, B.; Todd-Brown, K.; Thomas, L.; Ferreira, M.A.R.; Bender, D.; Maller, J.; Sklar, P.; Bakker, P.I.W.D.; Daly, M.J.; et al. PLINK: A tool set for whole-genome association and population-based linkage analyses. Am. J. Hum. Genet. 2007, 81, 559–575. [Google Scholar] [PubMed] [Green Version]
- Hu, B.; Jin, J.P.; Guo, A.Y.; Zhang, H.; Luo, J.C.; Gao, G. GSDS 20: An upgraded gene feature visualization server. Bioinformatics 2015, 31, 1296–1297. [Google Scholar] [PubMed] [Green Version]
- Song, S.; Wang, G.; Wu, H.; Fan, X.; Liang, L.; Zhao, H.; Li, S.; Hu, Y.; Liu, H.; Ayaad, M.; et al. OsMFT2 is involved in the regulation of ABA signaling-mediated seed germination through interacting with OsbZIP23/66/72 in rice. Plant J. 2020, 103, 532–546. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, M.; Barrero, J.M.; Corbineau, F.; Gubler, F.; BenechArnold, R.L. Dormancy in cereals (not too much not so little): About the mechanisms behind this trait. Seed Sci. Res. 2015, 25, 99–119. [Google Scholar]
- Kushiro, T.; Okamoto, M.; Nakabayashi, K.; Yamagishi, K.; Kitamura, S.; Asami, T.; Hirai, N.; Koshiba, T.; Kamiya, Y.; Nambara, E. The Arabidopsis cytochrome P450 CYP707A encodes ABA 8’-hydroxylases: Key enzymes in ABA catabolism. EMBO J. 2004, 23, 1647–1656. [Google Scholar] [CrossRef] [Green Version]
- Frey, A.; Effroy, D.; Lefebvre, V.; Seo, M.; Perreau, F.; Berger, A.; Sechet, J.; To, A.; North, H.M.; Marion-Poll, A. Epoxycarotenoid cleavage by NCED5 fine-tunes ABA accumulation and affects seed dormancy and drought tolerance with other NCED family members. Plant J. 2012, 70, 501–512. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Length of CDS (bp) | Putative Function | Reported Gene | Sequence Variation |
|---|---|---|---|---|
| Os01g0111200 | 1863 | Expressed protein | No variation | |
| Os01g0111300 | 162 | Expressed protein | No variation | |
| Os01g0111400 | 2106 | Transposon protein | 1 SNP: T → A caused Ser → Thr | |
| Os01g0111500 | 888 | Regulation of root hair development | OsbHLH125 OsRSL1 | No variation |
| Os01g0111600 | 525 | Regulation of ABA signaling-mediated seed germination | OsMFT2 | 1 InDel: 16 bp deletion caused stop codon |
| Marker Name | Type | Forward Primer (5′-3′) | Reverse Primer (5′-3′) | PHS-T-Type (bp) | PHS-S-Type (bp) |
|---|---|---|---|---|---|
| Os01g0111600-16bp | InDel | GGTGGGGATACACAGGTACG | CCTCTGGGAGTTGAAGTGGA | 166 | 150 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jang, S.-G.; Kim, B.; Choi, I.; Lee, J.; Ham, T.-H.; Kwon, S.-W. Fine-Mapping Analysis of the Genes Associated with Pre-Harvest Sprouting Tolerance in Rice (Oryza sativa L.). Agronomy 2023, 13, 818. https://doi.org/10.3390/agronomy13030818
Jang S-G, Kim B, Choi I, Lee J, Ham T-H, Kwon S-W. Fine-Mapping Analysis of the Genes Associated with Pre-Harvest Sprouting Tolerance in Rice (Oryza sativa L.). Agronomy. 2023; 13(3):818. https://doi.org/10.3390/agronomy13030818
Chicago/Turabian StyleJang, Seong-Gyu, Backki Kim, Insoo Choi, Joohyun Lee, Tae-Ho Ham, and Soon-Wook Kwon. 2023. "Fine-Mapping Analysis of the Genes Associated with Pre-Harvest Sprouting Tolerance in Rice (Oryza sativa L.)" Agronomy 13, no. 3: 818. https://doi.org/10.3390/agronomy13030818