
Genome-Wide Identification of Phytophthora sojae-Associated microRNAs and Network in a Resistant and a Susceptible Soybean Germplasm

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and P. sojae Isolate
2.2. Inoculation Method
2.3. RNA Extraction, Library Construction, and Illumina Sequencing
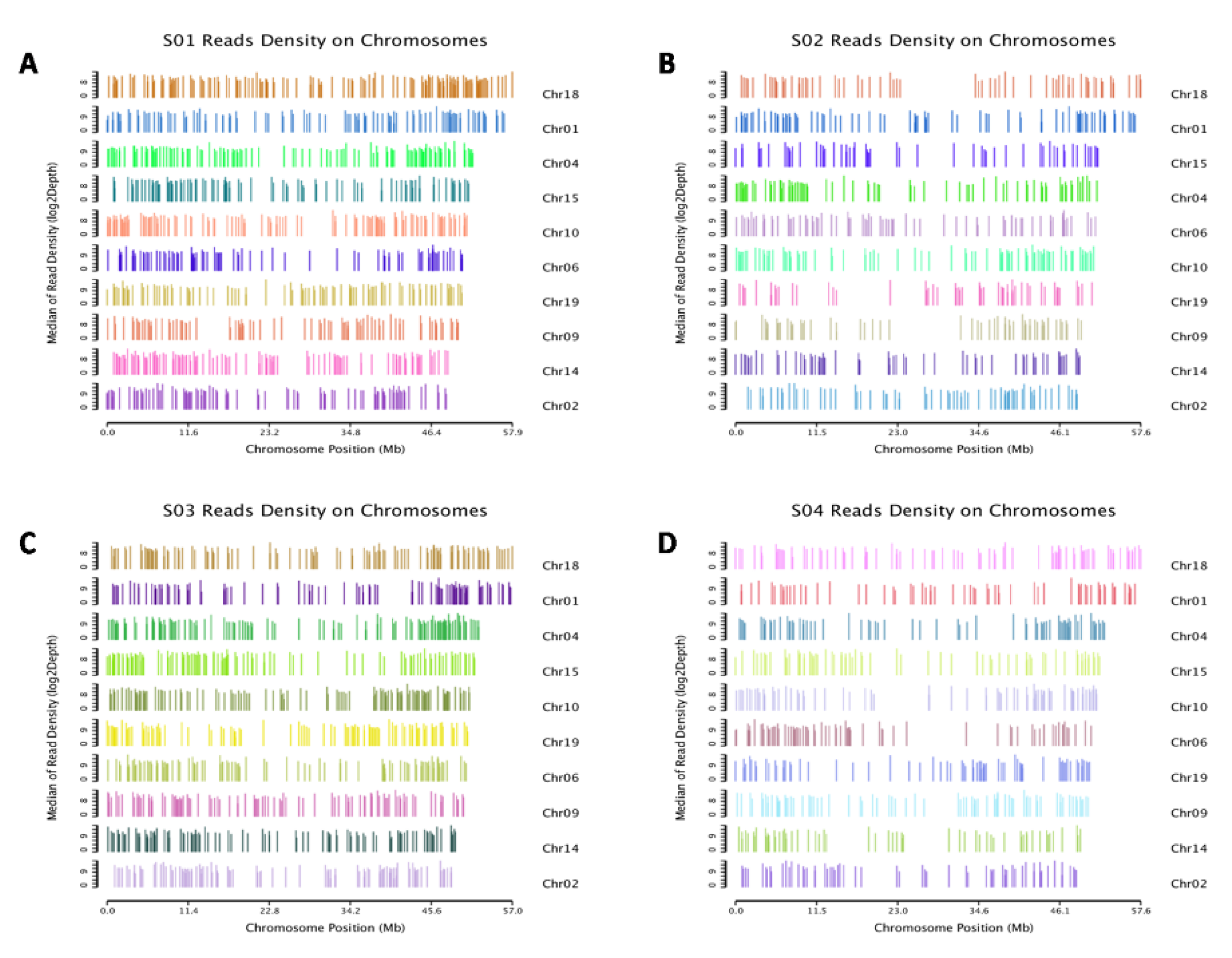
2.4. Alignment and Mapping of the Small-RNA Sequence Data
2.5. Identification of Known and Novel miRNAs
2.6. microRNA Expression Analysis
2.7. Validation of Differentially Expressed microRNA by Quantitative Real-Time PCR
2.8. Prediction and Confirmation of microRNA Target Genes
2.9. Functional Annotations of the Predicted Targets of the Differentially Expressed miRNA
3. Results
3.1. Resistance Response of Two Soybean Germplasms to P. sojae
3.2. High-Throughput Sequencing and Small-RNA Data Analysis
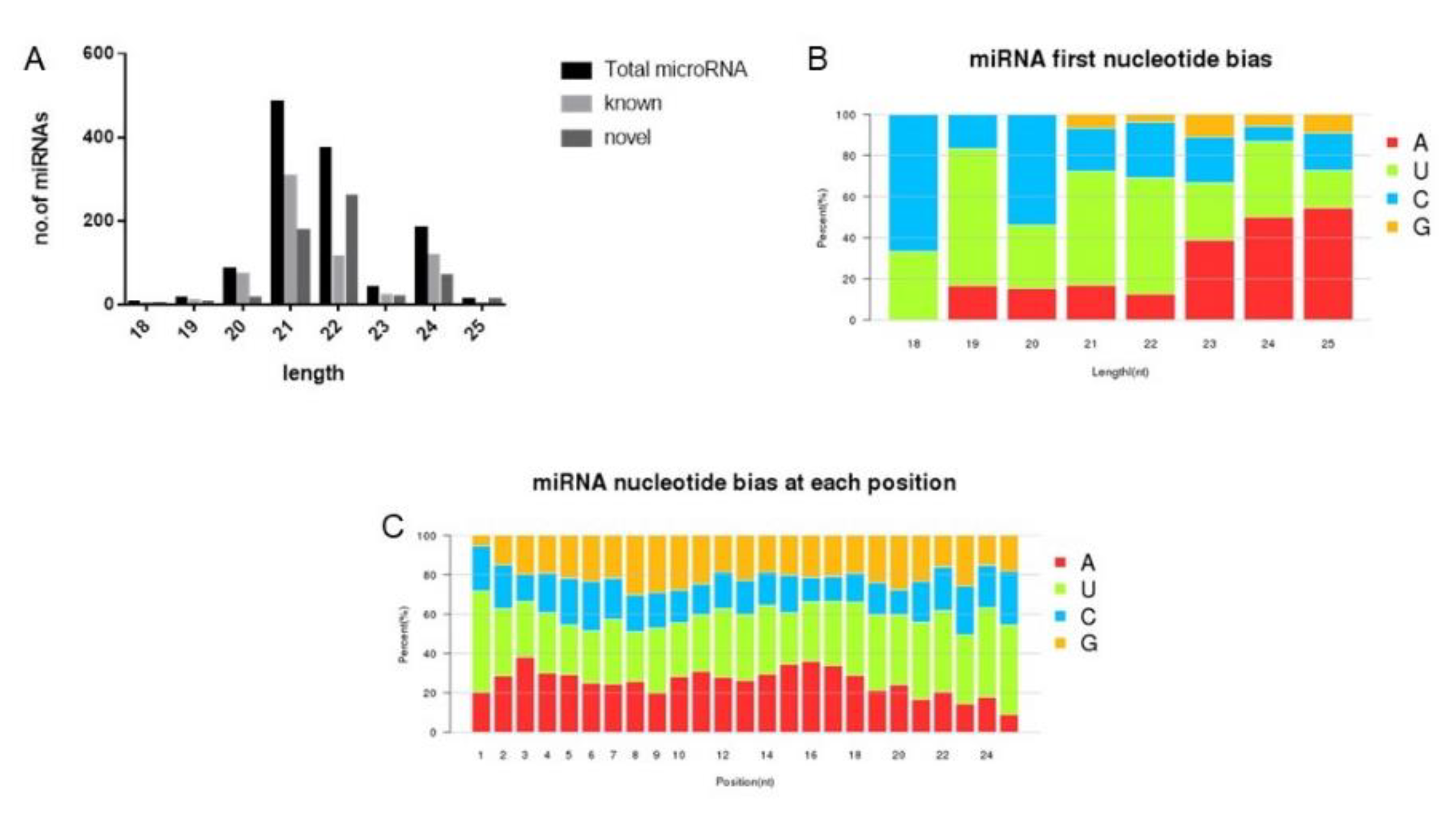
3.3. Identification of Conserved and Novel miRNA
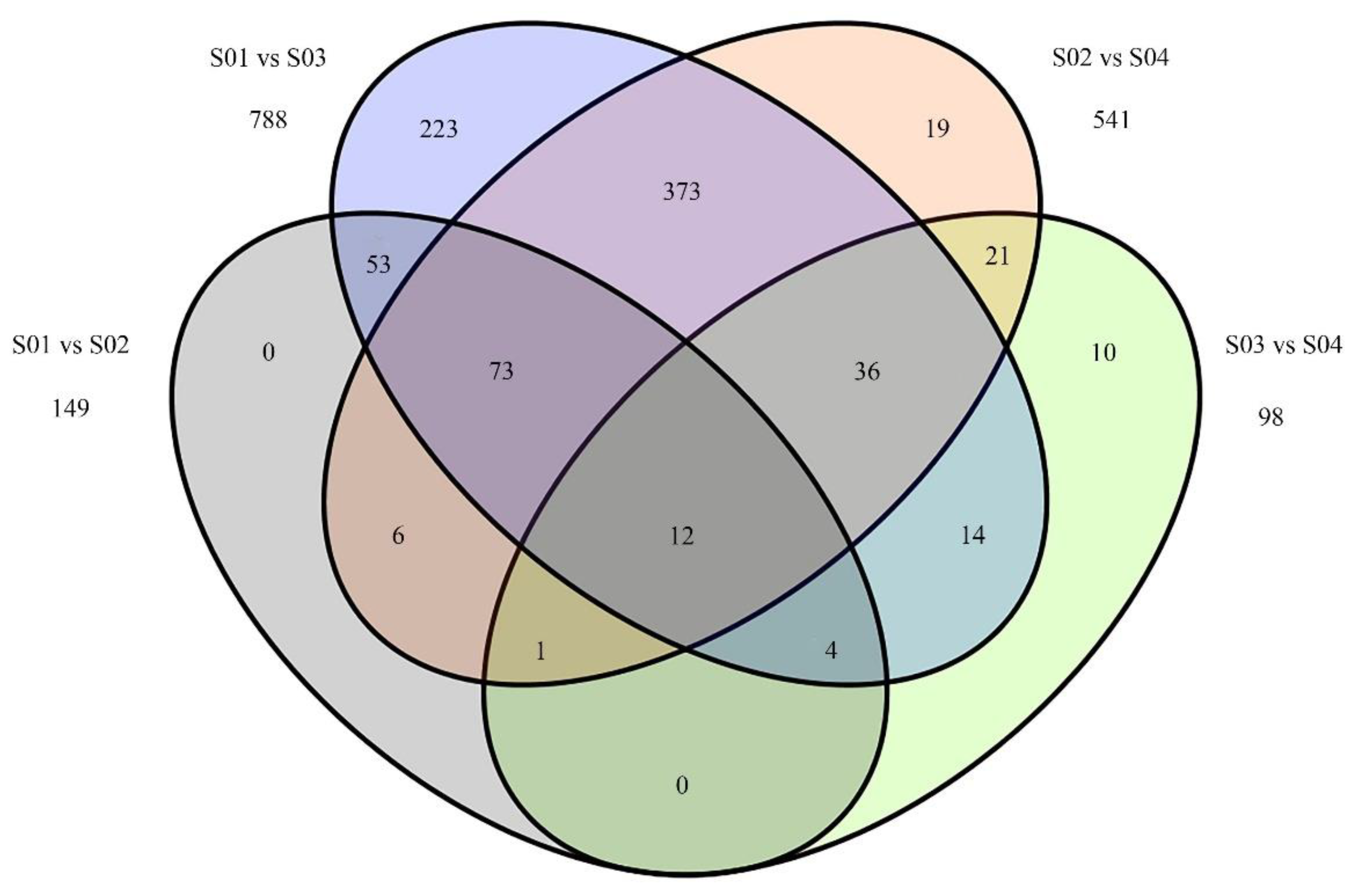
3.4. Identification of Differentially Expressed miRNAs upon P. sojae Infection
3.5. Validation of Sequencing Data by Stem-Loop qRT-PCR
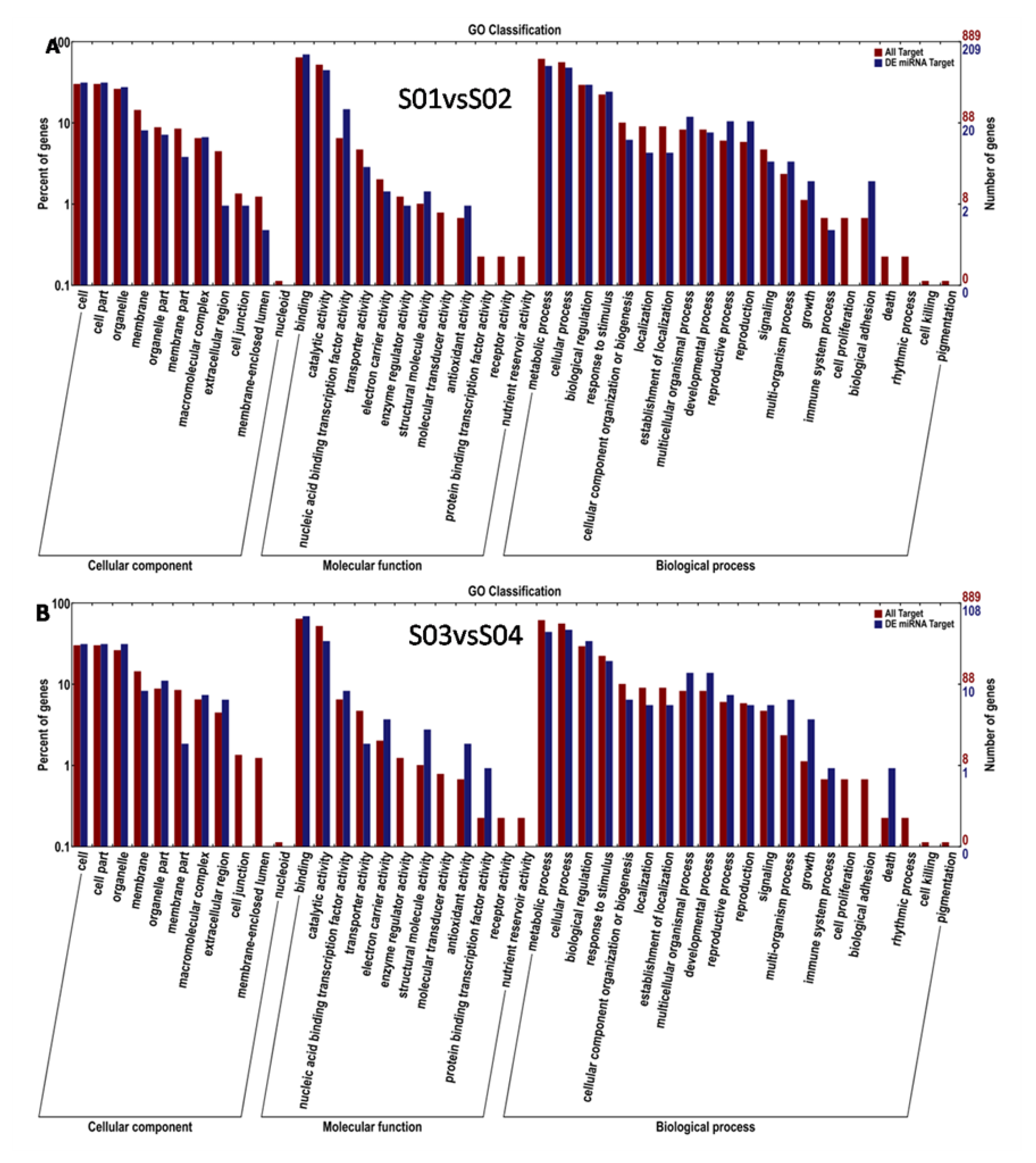
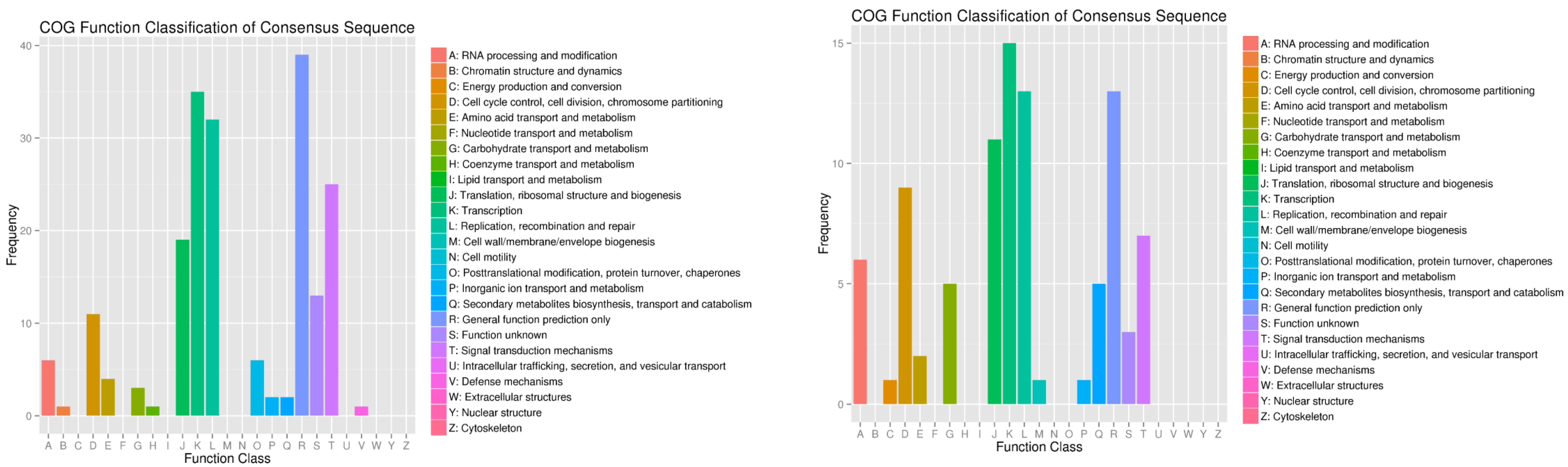
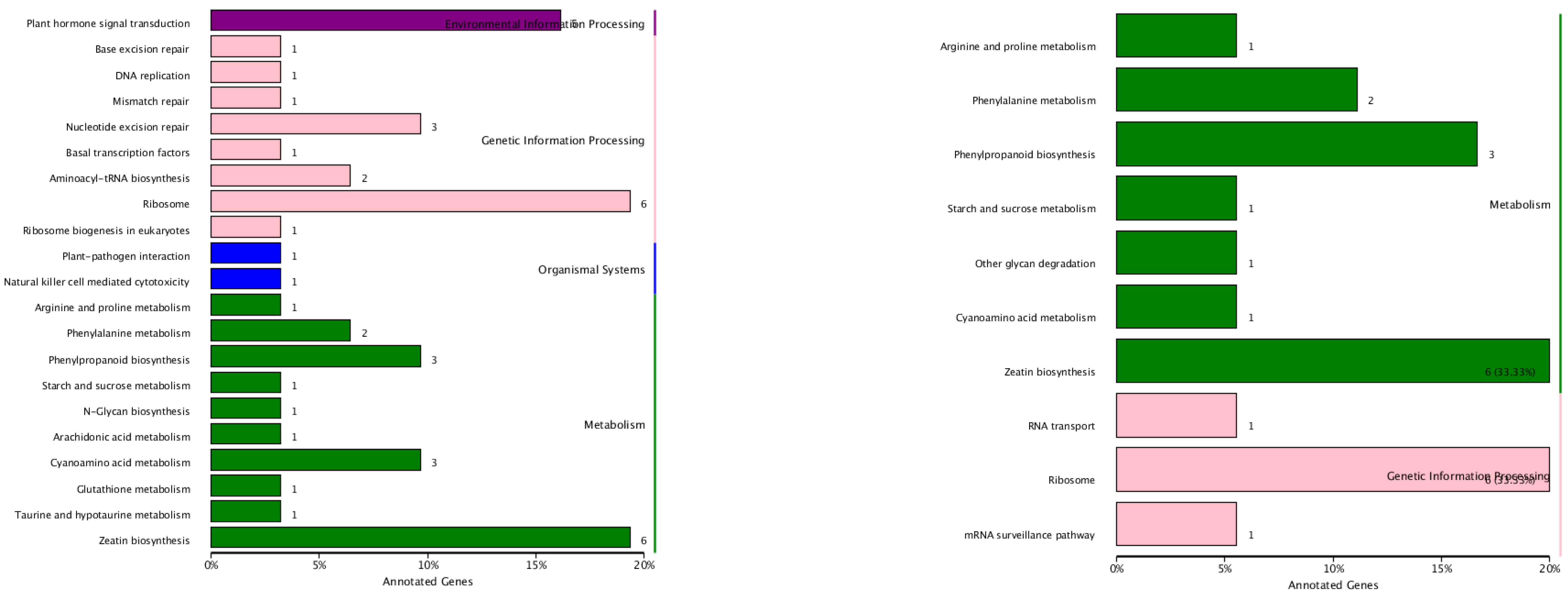
3.6. Target-Gene Prediction and Functional Annotations of the Predicted Targets of P. sojae-Responsive miRNAs in Resistant and Susceptible Soybean Germplasms
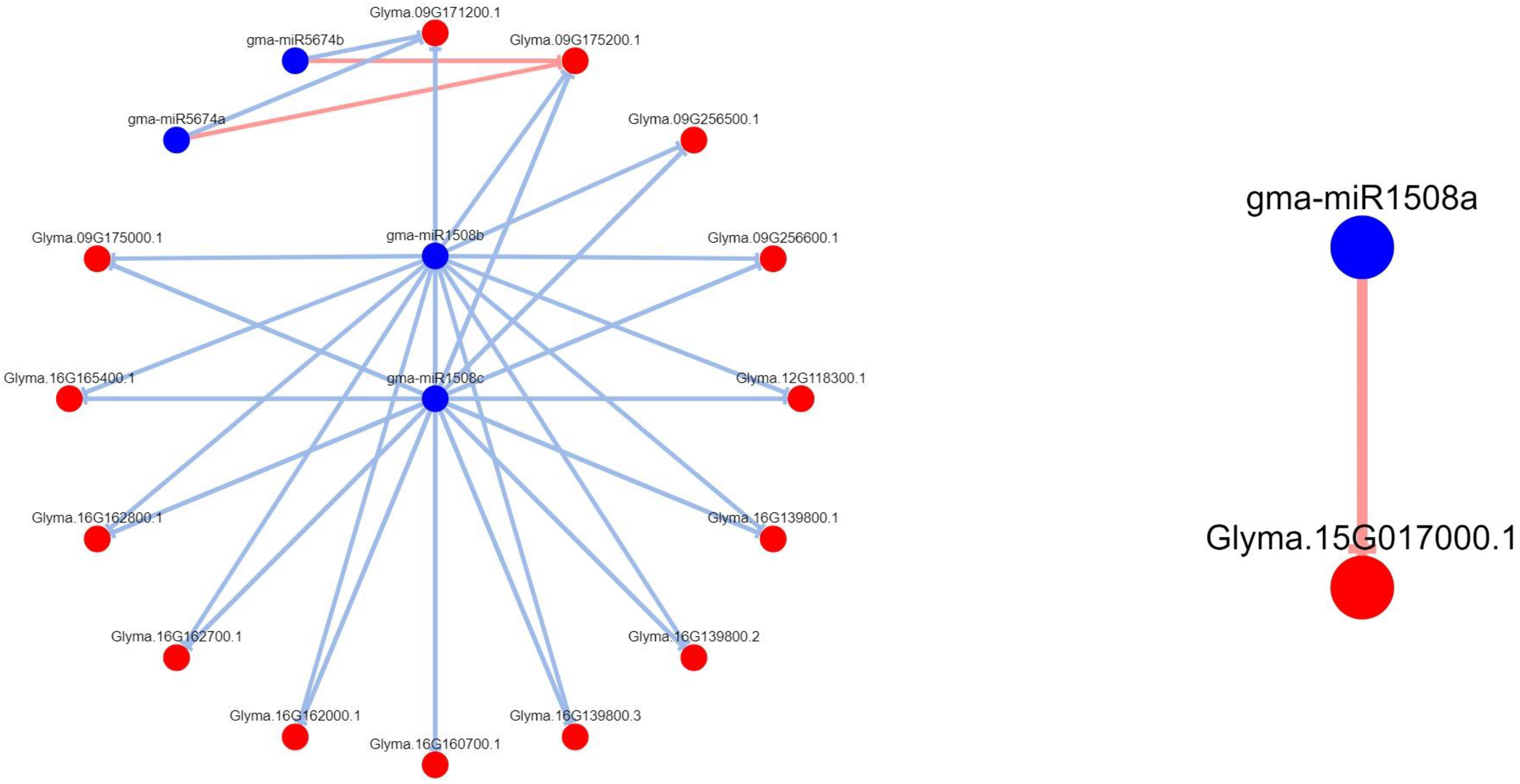
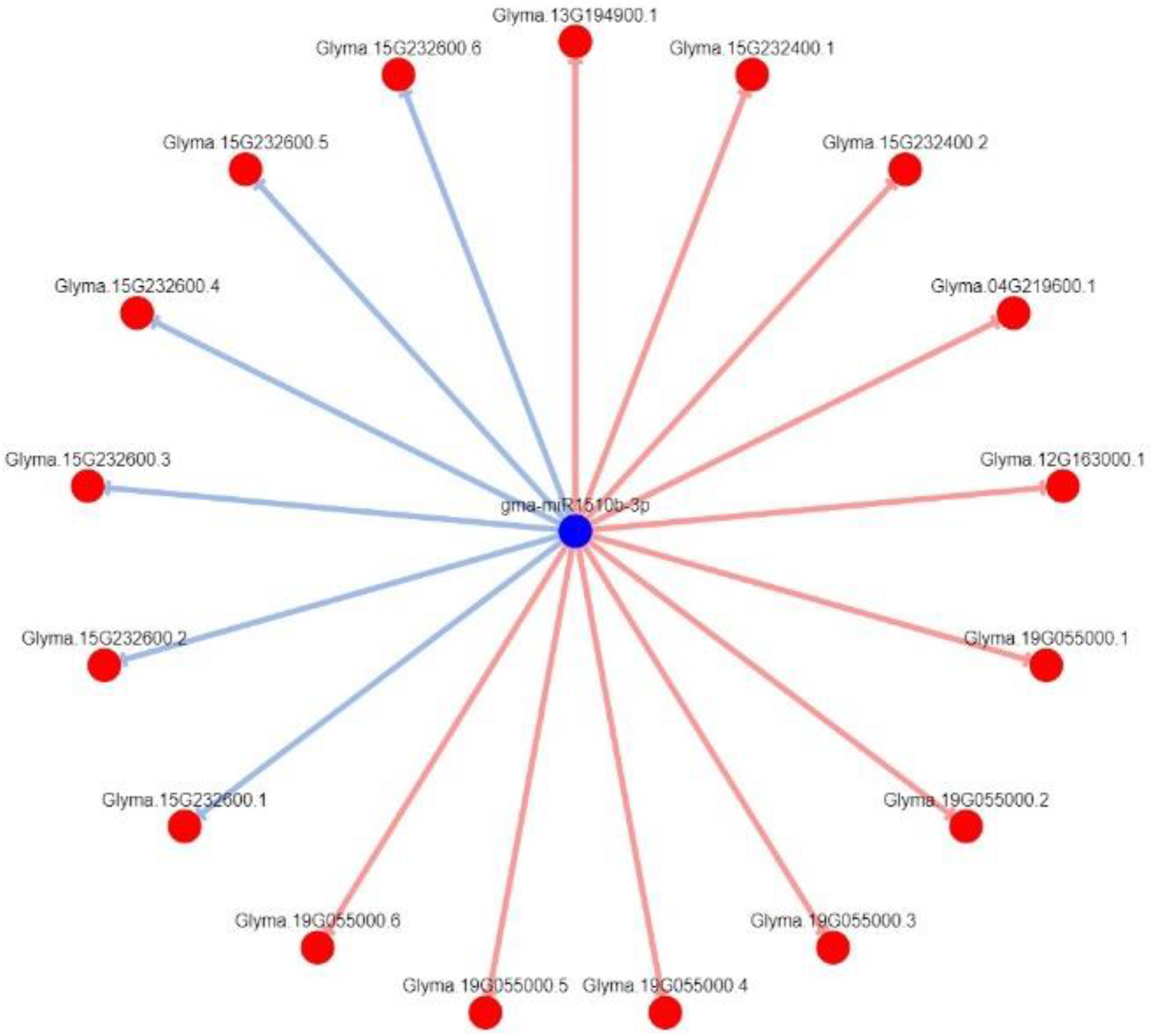
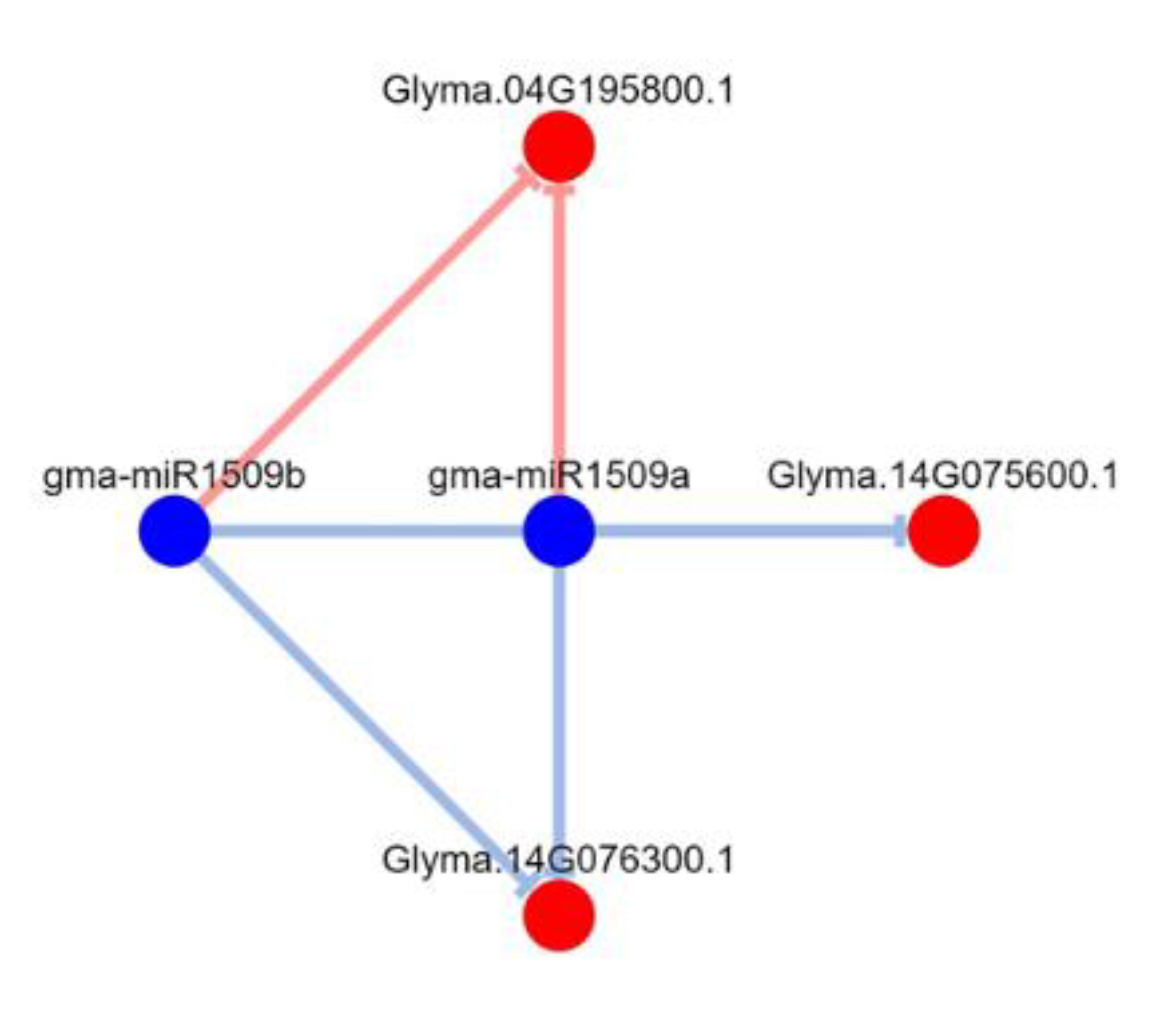
3.7. Regulatory Networks of Leguminous-Specific miRNA Targets
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Kamoun, S. Molecular genetics of pathogenic oomycetes. Eukaryot. Cell 2003, 2, 191–199. [Google Scholar] [CrossRef] [Green Version]
- Mattick, J.S. Challenging the dogma: The hidden layer of non-protein-coding RNAs in complex organisms. Bioessays 2003, 25, 930–939. [Google Scholar] [CrossRef] [PubMed]
- Reinhart, B.J.; Weinstein, E.G.; Rhoades, M.W.; Bartel, B.; Bartel, D.P. MicroRNAs in plants. Genes Dev. 2002, 16, 1616–1626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Röther, S.; Meister, G. Small RNAs derived from longer non-coding RNAs. Biochimie 2011, 93, 1905–1915. [Google Scholar] [CrossRef] [PubMed]
- Borges, F.; Martienssen, R.A. The expanding world of small RNAs in plants. Nat. Rev. Mol. Cell Biol. 2015, 16, 727–741. [Google Scholar] [CrossRef] [Green Version]
- Nazarov, P.V.; Kreis, S. Integrative approaches for analysis of mRNA and microRNA high-throughput data. Comput. Struct. Biotechnol. J. 2021, 19, 1154–1162. [Google Scholar] [CrossRef]
- Zhu, E.; Zhao, F.; Xu, G.; Hou, H.; Zhou, L.; Li, X.; Sun, Z.; Wu, J. mirTools: microRNA profiling and discovery based on high-throughput sequencing. Nucleic Acids Res. 2010, 38 (Suppl. S2), W392–W397. [Google Scholar] [CrossRef]
- Jones-Rhoades, M.W.; Bartel, D.P.; Bartel, B. MicroRNAs and their regulatory roles in plants. Annu. Rev. Plant Biol. 2006, 57, 19–53. [Google Scholar] [CrossRef]
- Guo, N.; Ye, W.-W.; Wu, X.-L.; Shen, D.-Y.; Wang, Y.-C.; Xing, H.; Dou, D.-L. Microarray profiling reveals microRNAs involving soybean resistance to Phytophthora sojae. Genome 2011, 54, 954–958. [Google Scholar] [CrossRef]
- Cui, X.; Yan, Q.; Gan, S.; Xue, D.; Dou, D.; Guo, N.; Xing, H. Overexpression of gma-miR1510a/b suppresses the expression of a NB-LRR domain gene and reduces resistance to Phytophthora sojae. Gene 2017, 621, 32–39. [Google Scholar] [CrossRef]
- Chen, L.; Luan, Y.; Zhai, J. Sp-miR396a-5p acts as a stress-responsive genes regulator by conferring tolerance to abiotic stresses and susceptibility to Phytophthora nicotianae infection in transgenic tobacco. Plant Cell Rep. 2015, 34, 2013–2025. [Google Scholar] [CrossRef] [PubMed]
- Robert-Seilaniantz, A.; MacLean, D.; Jikumaru, Y.; Hill, L.; Yamaguchi, S.; Kamiya, Y.; Jones, J.D.G. The microRNA miR393 re-directs secondary metabolite biosynthesis away from camalexin and towards glucosinolates. Plant J. 2011, 67, 218–231. [Google Scholar] [CrossRef] [PubMed]
- Schreiber, K. Chemical genomic analyses of plant-pathogen interactions. Ph.D. Thesis, University of Toronto, Toronto, ON, Canada, 2012. [Google Scholar]
- Hou, Y.; Zhai, Y.; Feng, L.; Karimi, H.Z.; Rutter, B.D.; Zeng, L.; Choi, D.S.; Zhang, B.; Gu, W.; Chen, X.; et al. A Phytophthora effector suppresses trans-kingdom RNAi to promote disease susceptibility. Cell Host Microbe 2019, 25, 153–165.e5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, J.; Gao, L.; Yang, Y.; Zhai, J.; Arikit, S.; Yu, Y.; Duan, S.; Chan, V.; Xiong, Q.; Yan, J.; et al. Roles of small RNA s in soybean defense against Phytophthora sojae infection. Plant J. 2014, 79, 928–940. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carter, T.E., Jr.; Nelson, R.L.; Sneller, C.H.; Cui, Z. Genetic diversity in soybean. Soybeans Improv. Prod. Uses 2004, 16, 303–416. [Google Scholar]
- Giachero, M.L.; Declerck, S.; Marquez, N. Phytophthora root rot: Importance of the disease, current and novel methods of control. Agronomy 2022, 12, 610. [Google Scholar] [CrossRef]
- Sun, J.; Li, L.; Zhao, J.; Huang, J.; Yan, Q.; Xing, H.; Guo, N. Genetic analysis and fine mapping of RpsJS, a novel resistance gene to Phytophthora sojae in soybean [Glycine max (L.) Merr.]. Theor. Appl. Genet. 2014, 127, 913–919. [Google Scholar] [CrossRef]
- Burnham, K.D.; Dorrance, A.E.; VanToai, T.T.; Martin, S.K.S. Quantitative trait loci for partial resistance to Phytophthora sojae in soybean. Crop. Sci. 2003, 43, 1610–1617. [Google Scholar] [CrossRef] [Green Version]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2012, 41, D590–D596. [Google Scholar] [CrossRef]
- Chan, P.P.; Lowe, T.M. GtRNAdb: A database of transfer RNA genes detected in genomic sequence. Nucleic Acids Res. 2009, 37 (Suppl. S1), D93–D97. [Google Scholar] [CrossRef]
- Kalvari, I.; Nawrocki, E.P.; Ontiveros-Palacios, N.; Argasinska, J.; Lamkiewicz, K.; Marz, M.; Griffiths-Jones, S.; Toffano-Nioche, C.; Gautheret, D.; Weinberg, Z.; et al. Rfam 14: Expanded coverage of metagenomic, viral and microRNA families. Nucleic Acids Res. 2021, 49, D192–D200. [Google Scholar] [CrossRef] [PubMed]
- Bao, W.; Kojima, K.K.; Kohany, O. Repbase Update, a database of repetitive elements in eukaryotic genomes. Mob. DNA 2015, 6, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Q.; Liu, G.; Bao, Y.; Wu, Y.; You, Q. Evaluation and application of tools for the identification of known microRNAs in plants. Appl. Plant Sci. 2021, 9, e11414. [Google Scholar] [CrossRef] [PubMed]
- Friedländer, M.R.; Mackowiak, S.D.; Li, N.; Chen, W.; Rajewsky, N. miRDeep2 accurately identifies known and hundreds of novel microRNA genes in seven animal clades. Nucleic Acids Res. 2012, 40, 37–52. [Google Scholar] [CrossRef] [Green Version]
- Kozomara, A.; Birgaoanu, M.; Griffiths-Jones, S. miRBase: From microRNA sequences to function. Nucleic Acids Res. 2018, 47, D155–D162. [Google Scholar] [CrossRef] [PubMed]
- Axtell, M.J.; Meyers, B.C. Revisiting criteria for plant microRNA annotation in the era of big data. Plant Cell 2018, 30, 272–284. [Google Scholar] [CrossRef] [Green Version]
- Zuker, M. Mfold web server for nucleic acid folding and hybridization prediction. Nucleic Acids Res. 2003, 31, 3406–3415. [Google Scholar] [CrossRef]
- Zhao, S.; Ye, Z.; Stanton, R. Misuse of RPKM or TPM normalization when comparing across samples and sequencing protocols. Rna 2020, 26, 903–909. [Google Scholar] [CrossRef] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative C(T) method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]
- Bo, X.; Wang, S. TargetFinder: A software for antisense oligonucleotide target site selection based on MAST and secondary structures of target mRNA. Bioinformatics 2004, 21, 1401–1402. [Google Scholar] [CrossRef]
- Mistry, J.; Chuguransky, S.; Williams, L.; Qureshi, M.; Salazar, G.A.; Sonnhammer, E.L.L.; Tosatto, S.C.; Paladin, L.; Raj, S.; Richardson, L.J.; et al. Pfam: The protein families database in 2021. Nucleic Acids Res. 2020, 49, D412–D419. [Google Scholar] [CrossRef] [PubMed]
- Fei, Y.; Wang, R.; Li, H.; Liu, S.; Zhang, H.; Huang, J. DPMIND: Degradome-based plant miRNA–target interaction and network database. Bioinformatics 2017, 34, 1618–1620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moy, P.; Qutob, D.; Chapman, B.P.; Atkinson, I.; Gijzen, M. Patterns of gene expression upon infection of soybean plants by Phytophthora sojae. Mol. Plant-Microbe Interact. 2004, 17, 1051–1062. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, B.; Wang, S.; Todd, T.C.; Johnson, C.D.; Tang, G.; Trick, H.N. Genome-wide identification of soybean microRNA responsive to soybean cyst nematodes infection by deep sequencing. BMC Genom. 2017, 18, 572. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Z.; Wang, Z.; Li, W.; Fang, C.; Shen, Y.; Li, C.; Wu, Y.; Tian, Z. Comprehensive analyses of microRNA gene evolution in paleopolyploid soybean genome. Plant J. 2013, 76, 332–344. [Google Scholar] [CrossRef] [PubMed]
- Yan, Z.; Hossain, M.S.; Arikit, S.; Valdés-López, O.; Zhai, J.; Wang, J.; Libault, M.; Ji, T.; Qiu, L.; Meyers, B.C.; et al. Identification of microRNAs and their mRNA targets during soybean nodule development: Functional analysis of the role of miR393j-3p in soybean nodulation. New Phytol. 2015, 207, 748–759. [Google Scholar] [CrossRef]
- Tian, F.; Yang, D.-C.; Meng, Y.-Q.; Jin, J.; Gao, G. PlantRegMap: Charting functional regulatory maps in plants. Nucleic Acids Res. 2020, 48, D1104–D1113. [Google Scholar] [CrossRef]
- Kanehisa, M.; Goto, S. KEGG: Kyoto encyclopedia of genes and genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef]
- Zhang, B.; Pan, X.; Stellwag, E.J. Identification of soybean microRNAs and their targets. Planta 2008, 229, 161–182. [Google Scholar] [CrossRef]
- Kulcheski, F.R.; De Oliveira, L.F.; Molina, L.G.; Almerão, M.P.; A Rodrigues, F.; Marcolino, J.; Barbosa, J.F.; Stolf-Moreira, R.; Nepomuceno, A.L.; Marcelino-Guimarães, F.C.; et al. Identification of novel soybean microRNAs involved in abiotic and biotic stresses. BMC Genom. 2011, 12, 307–317. [Google Scholar] [CrossRef]
- Turner, M.; Yu, O.; Subramanian, S. Genome organization and characteristics of soybean microRNAs. BMC Genom. 2012, 13, 169. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Liu, C.Y.; Zhang, L.W.; Wang, J.L.; Hu, G.H.; Ding, J.J.; Chen, Q.S. MicroRNAs involved in the pathogenesis of phytophthora root rot of soybean (Glycine max). Agric. Sci. China 2011, 10, 1159–1167. [Google Scholar]
- Zhou, L.; Deng, S.; Xuan, H.; Fan, X.; Sun, R.; Zhao, J.; Wang, H.; Guo, N.; Xing, H. A novel TIR-NBS-LRR gene regulates immune response to Phytophthora root rot in soybean. Crop. J. 2022. [Google Scholar] [CrossRef]
- Dodds, P.; Rathjen, J. Plant immunity: Towards an integrated view of plant-pathogen interactions. Nat. Rev. Genet. 2010, 11, 539–548. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Pignatta, D.; Bendix, C.; Brunkard, J.O.; Cohn, M.M.; Tung, J.; Sun, H.; Kumar, P.; Baker, B. MicroRNA regulation of plant innate immune receptors. Proc. Natl. Acad. Sci. USA 2012, 109, 1790–1795. [Google Scholar] [CrossRef] [Green Version]
- Zhai, J.; Jeong, D.H.; De Paoli, E.; Park, S.; Rosen, B.D.; Li, Y.; González, A.J.; Yan, Z.; Kitto, S.L.; Grusak, M.A.; et al. MicroRNAs as master regulators of the plant NB-LRR defense gene family via the production of phased, trans-acting siRNAs. Genes Dev. 2011, 25, 2540–2553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shivaprasad, P.V.; Chen, H.-M.; Patel, K.; Bond, D.; Santos, B.; Baulcombe, D.C. A microRNA superfamily regulates nucleotide binding site–leucine-rich repeats and other mRNAs. Plant Cell 2012, 24, 859–874. [Google Scholar] [CrossRef] [Green Version]
- Lam, H.-M.; Xu, X.; Liu, X.; Chen, W.; Yang, G.; Wong, F.-L.; Li, M.-W.; He, W.; Qin, N.; Wang, B.; et al. Resequencing of 31 wild and cultivated soybean genomes identifies patterns of genetic diversity and selection. Nat. Genet. 2010, 42, 1053–1059. [Google Scholar] [CrossRef]
- Cuperus, J.T.; Carbonell, A.; Fahlgren, N.; Garcia-Ruiz, H.; Burke, R.T.; Takeda, A.; Sullivan, C.M.; Gilbert, S.D.; Montgomery, T.A.; Carrington, J.C. Unique functionality of 22-nt miRNAs in triggering RDR6-dependent siRNA biogenesis from target transcripts in Arabidopsis. Nat. Struct. Mol. Biol. 2010, 17, 997–1003. [Google Scholar] [CrossRef]
- Wang, L.; Gu, X.; Xu, D.; Wang, W.; Wang, H.; Zeng, M.; Chang, Z.; Huang, H.; Cui, X. miR396-targeted AtGRF transcription factors are required for coordination of cell division and differentiation during leaf development in Arabidopsis. J. Exp. Bot. 2011, 62, 761–773. [Google Scholar] [CrossRef] [Green Version]
- Akdogan, G.; Tufekci, E.D.; Uranbey, S.; Unver, T. miRNA-based drought regulation in wheat. Funct. Integr. Genom. 2016, 16, 221–233. [Google Scholar] [CrossRef] [PubMed]
- Chen, X. MicroRNA biogenesis and function in plants. FEBS Lett. 2005, 579, 5923–5931. [Google Scholar] [CrossRef] [Green Version]
- Chen, X. A microRNA as a translational repressor of APETALA2 in Arabidopsis flower development. Science 2004, 303, 2022–2025. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hansen, C.C.; Sørensen, M. Reconfigured cyanogenic glucoside biosynthesis in eucalyptus cladocalyx involves a cytochrome P450 CYP706C55. Plant Physiol. 2018, 178, 1081–1095. [Google Scholar] [CrossRef] [Green Version]
- Liu, D.; Song, Y.; Chen, Z.; Yu, D. Ectopic expression of miR396 suppresses GRF target gene expression and alters leaf growth in Arabidopsis. Physiol. Plant. 2009, 136, 223–236. [Google Scholar] [CrossRef]
- Yu, X.-Z.; Trapp, S.; Zhou, P.-H.; Chen, L. Effect of temperature on the uptake and metabolism of cyanide by weeping willows. Int. J. Phytoremediat. 2007, 9, 243–255. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples | Raw Reads | Low-Quality Reads | Containing ‘N’ Reads | <18 nt Reads | >30 nt Reads | Clean Reads | Q30(%) |
|---|---|---|---|---|---|---|---|
| S01 | 22,082,654 | 0 | 9865 | 1,083,291 | 6,606,261 | 14,383,237 | 92.45 |
| S02 | 22,573,647 | 0 | 9143 | 2,029,533 | 4,360,662 | 16,174,309 | 90.77 |
| S03 | 26,694,260 | 0 | 10,648 | 3,134,974 | 3,947,560 | 19,601,078 | 91.21 |
| S04 | 26,574,283 | 0 | 20,491 | 1,296,545 | 6,090,230 | 19,167,017 | 92.37 |
| Sample | Total (%) | Genome (%) | rRNA (%) | snRNA (%) | snoRNA (%) | tRNA (%) | Repbase (%) | Unannotated (%) |
|---|---|---|---|---|---|---|---|---|
| S01 | 14,383,237 (100.00) | 2,817,311 (19.59) | 6,433,872 (44.73) | 8672 (0.06) | 1619 (0.01) | 762,461 (5.30) | 99,096 (0.69) | 7,077,517 (49.21) |
| S02 | 16,174,309 (100.00) | 1,957,192 (12.10) | 10,286,919 (63.60) | 8778 (0.05) | 1071 (0.01) | 435,164 (2.69) | 64,788 (0.40) | 5,377,589 (33.25) |
| S03 | 19,601,078 (100.00) | 4,877,586 (24.88) | 8,649,047 (44.13) | 11,775 (0.06) | 3298 (0.02) | 728,214 (3.72) | 104,090 (0.53) | 10,104,654 (51.55) |
| S04 | 19,167,017 (100.0) | 2,607,881 (13.61) | 12,063,854 (62.94) | 14,216 (0.07) | 1397 (0.01) | 511,338 (2.67) | 59,459 (0.31) | 6,516,753 (34.00) |
| Type | No. | Up | Down | ||
|---|---|---|---|---|---|
| Novel | Known | Novel | Known | ||
| S01 vs. S02 | 149 | 6 | 12 | 69 | 62 |
| S01 vs. S03 | 788 | 6 | 17 | 416 | 349 |
| S02 vs. S04 | 541 | 1 | 6 | 247 | 287 |
| S03 vs. S04 | 98 | 39 | 9 | 4 | 46 |
| miRNA | Germplasm | Fold Change | Consistency | |
|---|---|---|---|---|
| RNA-seq (log2FC) | RT-PCR (2-△△Ct) | |||
| gma-miR1508a | Nannong 10-1 | −0.98 | −3.49 | Y |
| gma-miR1508c | Nannong 10-1 | −0.95 | −0.12 | Y |
| gma-miR1510a-5p | Nannong 10-1 | −1.19 | −0.32 | Y |
| gma-miR396b-5p | Nannong 10-1 | −0.80 | −0.47 | Y |
| gma-miR396c | Nannong 10-1 | −0.80 | −0.47 | Y |
| gma-miR396k-5p | Nannong 10-1 | −0.80 | −0.47 | Y |
| gma-miR4413b | Nannong 10-1 | −1.19 | −0.76 | Y |
| gma-miR482a-3p | Nannong 10-1 | −0.67 | −0.39 | Y |
| gma-miR5374-5p | Nannong 10-1 | −0.72 | −0.10 | Y |
| gma-miR5376 | Nannong 10-1 | −0.83 | −0.63 | Y |
| gma-miR1535b | 06-070583 | 0.78 | 1.33 | Y |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, N.; Tahir, A.; Cui, X.; Xu, J.; Sun, J.; Zhang, N.; Sun, R.; Deng, S.; Xing, H.; Zhao, J. Genome-Wide Identification of Phytophthora sojae-Associated microRNAs and Network in a Resistant and a Susceptible Soybean Germplasm. Agronomy 2022, 12, 2922. https://doi.org/10.3390/agronomy12122922
Guo N, Tahir A, Cui X, Xu J, Sun J, Zhang N, Sun R, Deng S, Xing H, Zhao J. Genome-Wide Identification of Phytophthora sojae-Associated microRNAs and Network in a Resistant and a Susceptible Soybean Germplasm. Agronomy. 2022; 12(12):2922. https://doi.org/10.3390/agronomy12122922
Chicago/Turabian StyleGuo, Na, Ammara Tahir, Xiaoxia Cui, Jianyu Xu, Jutao Sun, Nannan Zhang, Ruidong Sun, Sushuang Deng, Han Xing, and Jinming Zhao. 2022. "Genome-Wide Identification of Phytophthora sojae-Associated microRNAs and Network in a Resistant and a Susceptible Soybean Germplasm" Agronomy 12, no. 12: 2922. https://doi.org/10.3390/agronomy12122922