
Palynological Study of Weed Flora from Potohar Plateau


 ,
,  ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Specimen Collection and Plant Identification
2.2. Study of Pollen under an Optical Microscope
2.3. Study of Pollen under a Scanning Electron Microscope
2.4. Data Analysis
2.5. Percentages of Sterility and Fertility among Pollen
2.6. Taxonomic Key
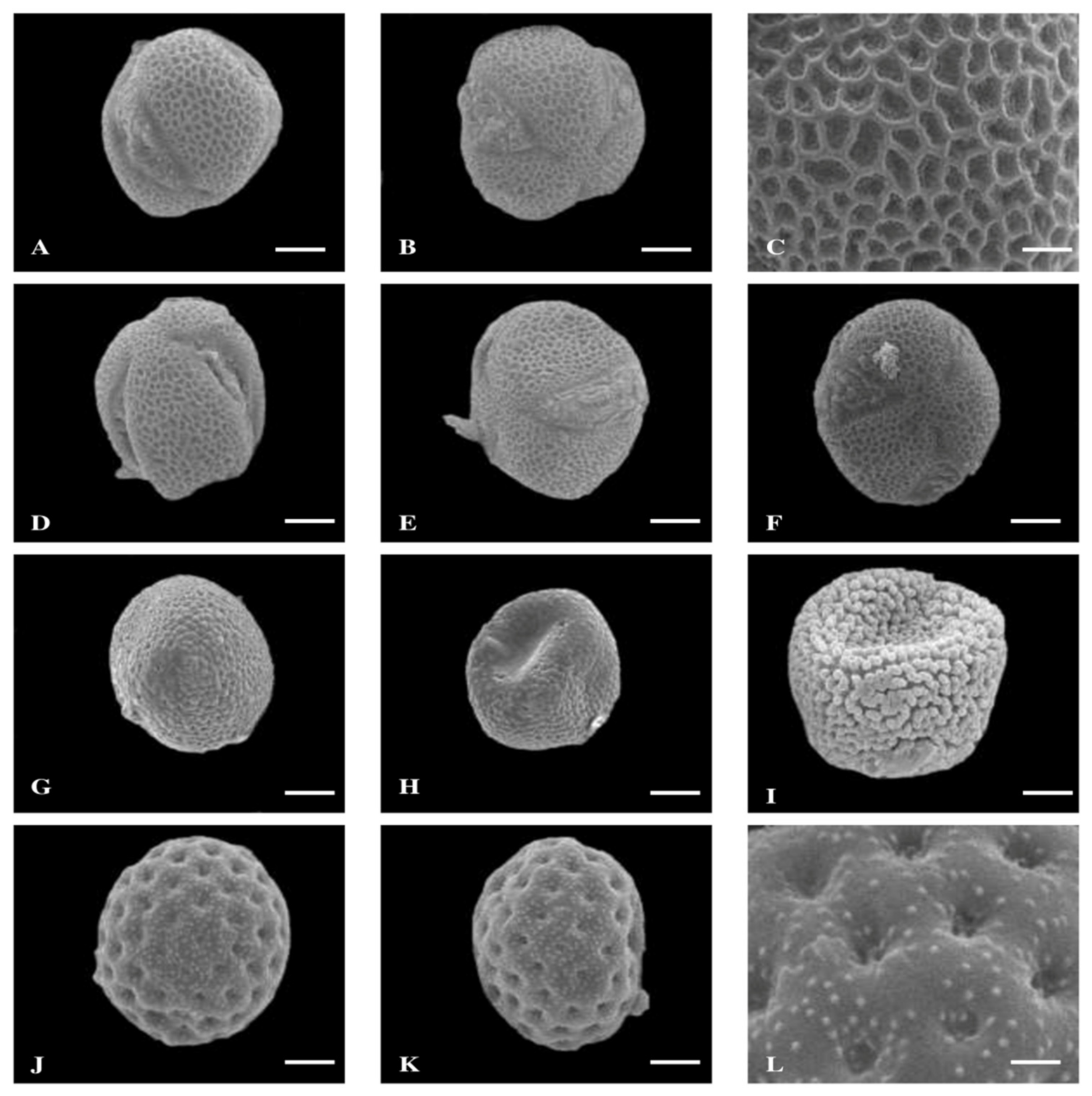
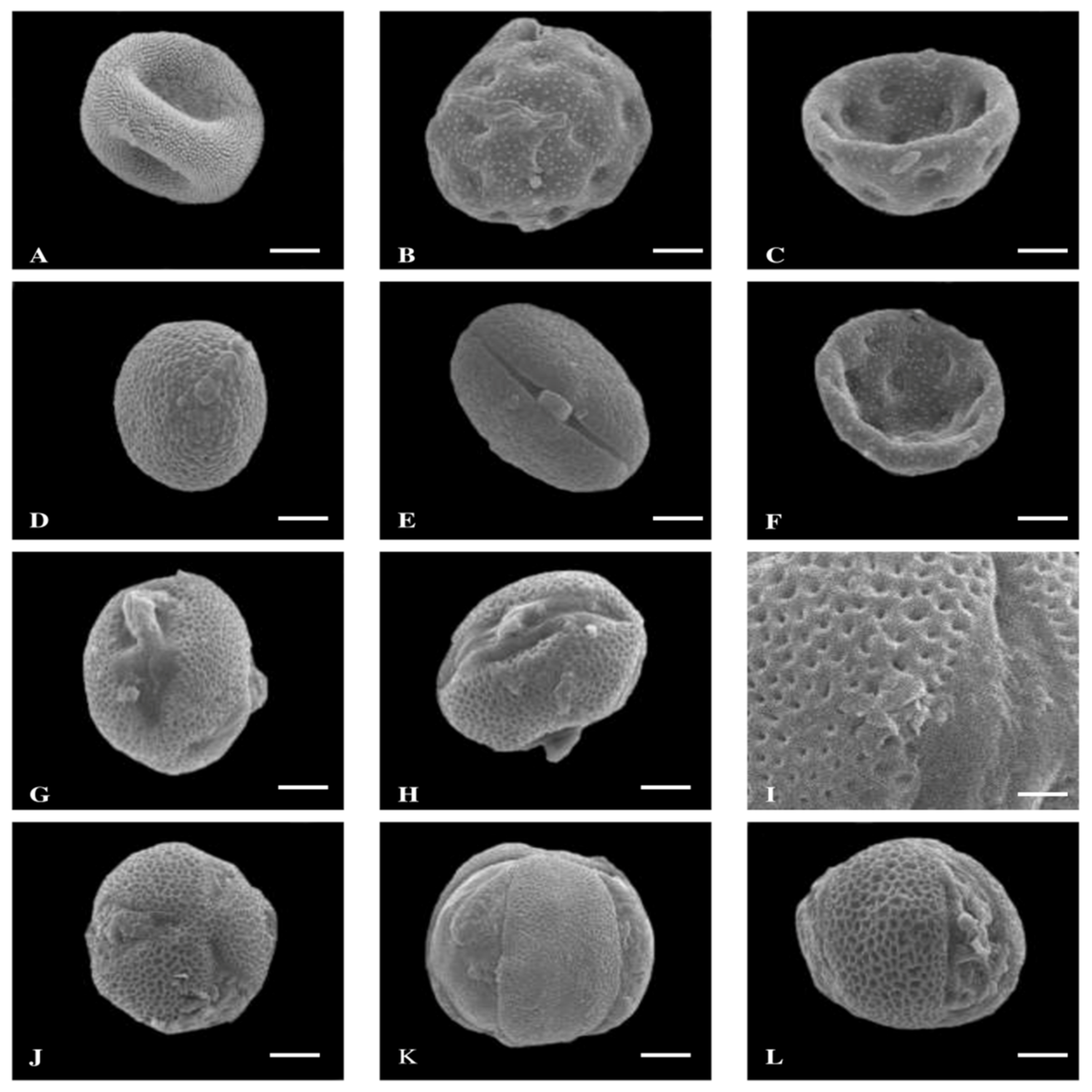
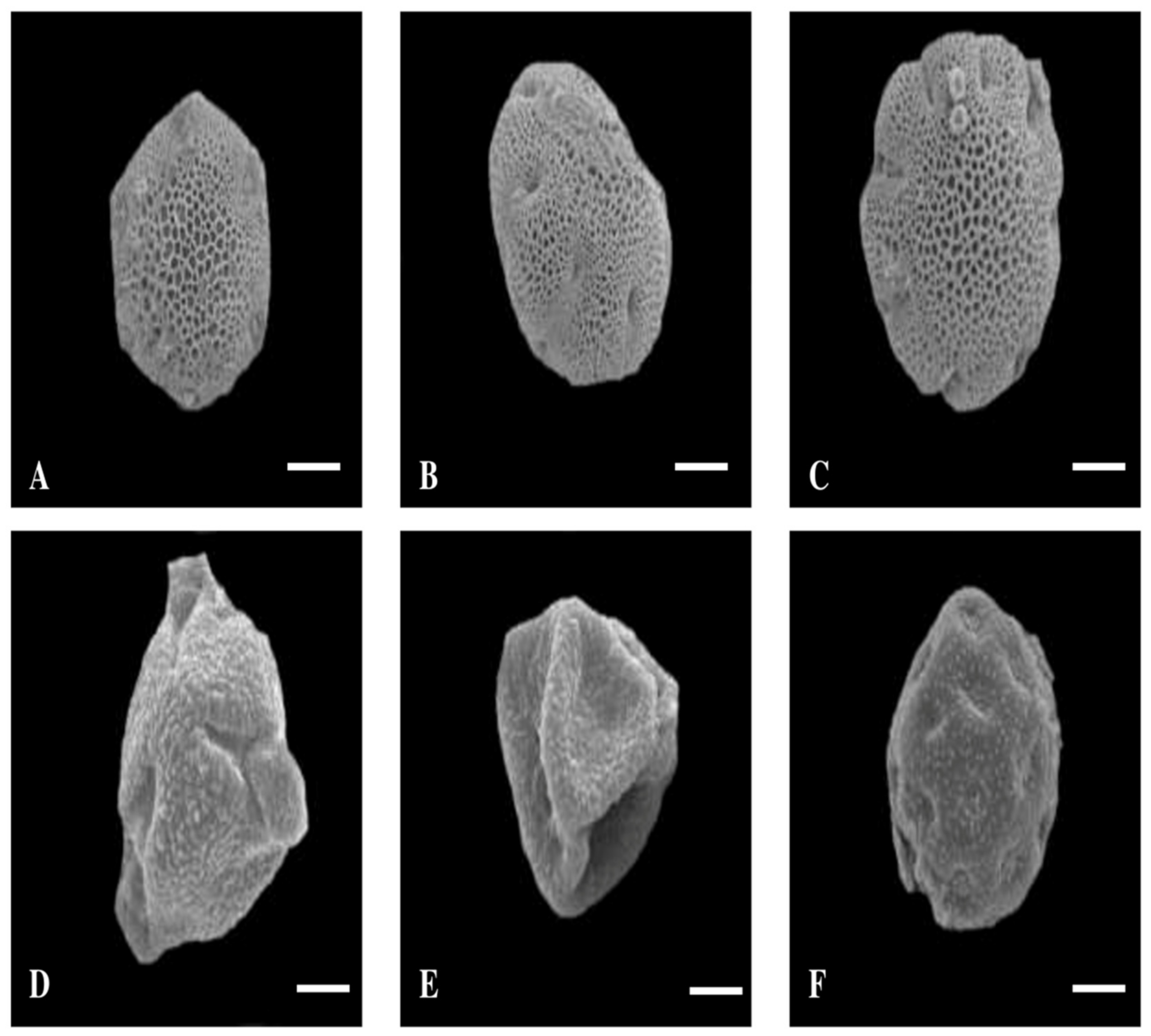
3. Results
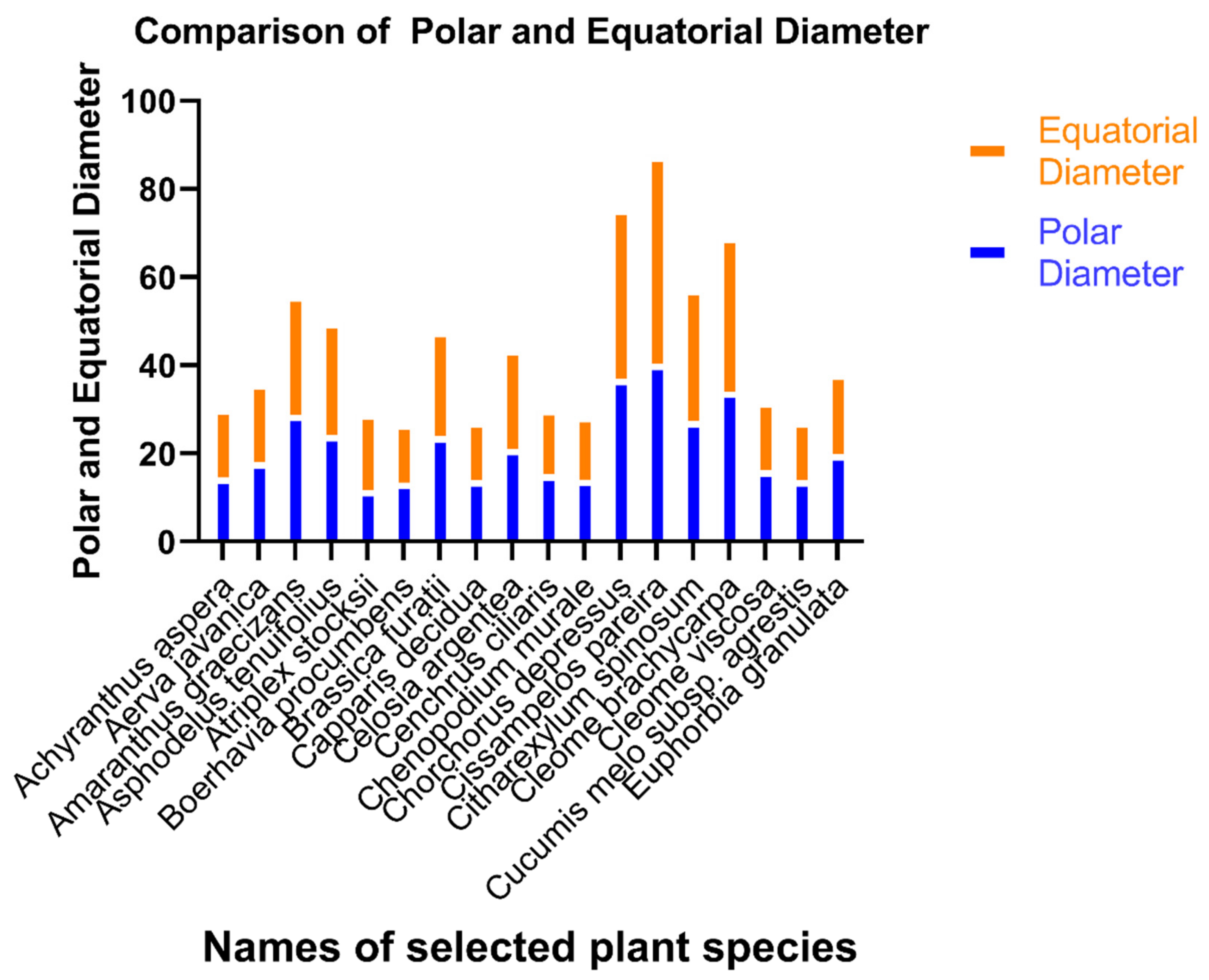
3.1. Pollen Size, Shape, and P/E Ratio
3.2. Quantitative and Qualitative Characters of Colpi
3.3. Exine Thickness
3.4. Exine Sculpturing
3.5. Fertility and Sterility Percentages
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Oerke, E.C.; Dehne, H.W. Safeguarding production—Losses in major crops and the role of crop protection. Crop Prot. 2004, 23, 275–285. [Google Scholar] [CrossRef]
- Khan, F.; Muhammad, Z.; Khan, K.; Ahmad, S.; Khan, M.J.; Bakht, T.; Kamal, A. Palynological investigation of allergenic and invasive weeds plants for biodiversity in district Lakki Marwat using scanning electron microscopy. Pak. J. Weed Sci. Res. 2020, 26, 349–365. [Google Scholar] [CrossRef]
- Qureshi, R.; Waheed, A.; Arshad, M. Weed communities of wheat crop in district Toba Tek Singh, Pakistan. Pak. J. Bot 2009, 41, 239–245. [Google Scholar]
- Neve, P.; Barney, J.N.; Buckley, Y.; Cousens, R.D.; Graham, S.; Jordan, N.R.; Lawton-Rauh, A.; Liebman, M.; Mesgaran, M.B.; Schut, M. Reviewing research priorities in weed ecology, evolution and management: A horizon scan. Weed Res. 2018, 58, 250–258. [Google Scholar] [CrossRef] [Green Version]
- Ullah, F.; Zafar, M.; Ahmad, M.; Dilbar, S.; Shah, S.N.; Sohail, A.; Zaman, W.; Iqbal, M.; Bahadur, S.; Tariq, A. Pollen morphology of subfamily Caryophylloideae (Caryophyllaceae) and its taxonomic significance. Microsc. Res. Tech. 2018, 81, 704–715. [Google Scholar] [CrossRef] [PubMed]
- Mamoona, M.; Mir, A.K.; Mushtaq, A.; Nighat, S.; Sidra, N.A.; Kanwal, T.; Saira, T.; Tehmeena, M.; Madhia, A.; Shazia, B. Foliar epidermal anatomy of some ethnobotanically important species of wild edible fruits of northern Pakistan. J. Med. Plants Res. 2011, 5, 5873–5880. [Google Scholar]
- Jogi, Q.; Hajano, G.A.; Kandharo, M.N.; Shah, A.N.; Soomro, A.A.; Abbasi, Z.A.; Burioro, M.; Baloch, S.K.; Kalwar, Z.A.; Abbasi, N.A. Examine different weed management techniques in sugarcane (Saccharum officinarum L.). Pure Appl. Biol. (PAB) 2019, 8, 151–159. [Google Scholar]
- Mehsud, A.; Mehmood, S.; Muhammad, A.; Khan, R.U.; Khan, S.U.; Khan, H.U.; Wazir, R.; Hussain, Z. Morphology and anatomy of some weeds from flora of district Bannu, Pakistan. Pak. J. Weed Sci. Res. 2013, 19, 437–445. [Google Scholar]
- Amarger, N. Rhizobia in the field. Adv. Agron. 2001, 23, 741–748. [Google Scholar]
- Marwat, K. A checklist and key to parasitic weeds of Pakistan. Pak. J. Weed Sci. Res. 1933, 6, 1–15. [Google Scholar]
- Naz, J.; Ahmad, M.; Sultana, S.; Zafar, M.; Naz, S.; Zaman, W.; Ullah, F. Invasive alien species of Potohar Plateau, Northern Pakistan. Ann. Bot. 2021, 11, 171–179. [Google Scholar]
- Rashid, K.; Rasul, G. Rainfall variability and maize production over the Potohar Plateau of Pakistan. Pak. J. Meteorol. 2011, 8, 63–74. [Google Scholar]
- Muhammad, Z.; Khan, M.; Mushtaq, A.; Shazia, S. Palynological and taxonomic studies of some weeds from flora of Rawalpindi. Pak. J. Weed Sci. Res. 2006, 12, 99–109. [Google Scholar]
- Hameed, A.; Zafar, M.; Ahmad, M.; Sultana, S.; Akhter, M.S.; Zaman, W.; Saqib, S.; Ullah, F. Micromorphology, phytochemical and pharmacological evaluation of Isodon rugosus (wall. Ex benth.) Codd. J. Anim. Plant Sci. 2022, 32, 736–745. [Google Scholar]
- Jehanzeb, S.; Zafar, M.; Ahmad, M.; Sultana, S.; Zaman, W.; Ullah, F. Comparative petioler anatomy of tribe Mentheae subfamily Nepetoideae, Lamiaceae from Pakistan. Feddes Repert. 2020, 131, 163–174. [Google Scholar] [CrossRef]
- Shah, S.N.; Ahmad, M.; Zafar, M.; Ullah, F.; Zaman, W.; Malik, K.; Rashid, N.; Gul, S. Taxonomic importance of spore morphology in Thelypteridaceae from Northern Pakistan. Microsc. Res. Tech. 2019, 82, 1326–1333. [Google Scholar] [CrossRef]
- Ullah, F.; Ayaz, A.; Saqib, S.; Parmar, G.; Bahadur, S.; Zaman, W. Taxonomic implication of leaf epidermal anatomy of selected taxa of Scrophulariaceae from Pakistan. Microsc. Res. Tech. 2020, 84, 521–530. [Google Scholar] [CrossRef]
- Usma, A.; Ahmad, M.; Zafar, M.; Sultana, S.; Lubna; Kalsoom, N.; Zaman, W.; Ullah, F. Micromorphological variations and taxonomic implications of caryopses of some grasses from Pakistan. Wulfenia 2020, 27, 86–96. [Google Scholar]
- Zaman, W.; Ullah, F.; Parmar, G.; Saqib, S.; Ayaz, A.; Park, S. Foliar micromorphology of selected medicinal Lamiaceae taxa and their taxonomic implication using scanning electron microscopy. Microsc. Res. Tech. 2022, 2022, 1–20. [Google Scholar] [CrossRef]
- Meo, A.; Khan, M. Palynological studies of some weeds of asteraceae from Pakistan. Pak. J. Weed Sci. Res. (Pak.) 2004, 10, 87–92. [Google Scholar]
- Ronald, O.K. Pollen and Spores, 2nd ed.; American Association of Stratographic Palynologists Foundation: College Station, TX, USA, 2000; pp. 13–21. [Google Scholar]
- Ashfaq, S.; Ahmad, M.; Zafar, M.; Sultana, S.; Nazish, M.; Khan, A.N. Systematics of medicinally important weeds of genus Convolvulus: Convolvulaceae. Planta Daninha 2020, 38. [Google Scholar] [CrossRef]
- Erdtmann, G. Pollen morphology and plant taxonomy III, Morina L. with an addition on pollen morphological terminology. Sven. Bot. Tidskr. 1952, 39, 187–191. [Google Scholar]
- Khan, M.A.; Stace, C.A. Breeding relationships in the genus Brachypodium (Poaceae: Pooideae). Nord. J. Bot. 1999, 19, 257–269. [Google Scholar] [CrossRef]
- Ahmad, S.; Zafar, M.; Ahmad, M.; Yaseen, G.; Sultana, S. Microscopic investigation of palyno-morphological features of melliferous flora of Lakki Marwat district, Khyber Pakhtunkhwa, Pakistan. Microsc. Res. Tech. 2019, 82, 720–730. [Google Scholar] [CrossRef]
- Ahmad, S.; Zafar, M.; Ahmad, M.; Ozdemir, F.; Yaseen, G.; Sultana, S.; Kutlu, M. Palynological studies of winter weeds melliferous flora of district Bannu, Khyber Pakhtunkhwa, Pakistan. Ann. Bot. 2020, 10, 77–86. [Google Scholar]
- Raees, K.; Abidin, S.Z.U.; Mumtaz, A.S.; Jamsheed, S.; Ullah, H. Comparative leaf and pollen micromorphology on some Grasses taxa (Poaceae) distributed in Pakistan. Int. J. Nat. Life Sci. 2017, 1, 72–82. [Google Scholar]
- Nazish, M.; Zafar, M.; Ahmad, M.; Sultana, S.; Ullah, R.; Alqahtani, A.S.; Ullah, F.; Ahmad, S.; Ashfaq, S.; Ullah, F. Palyno-morphological investigations of halophytic taxa of Amaranthaceae through SEM from Salt range of Northern Punjab, Pakistan. Microsc. Res. Tech. 2019, 82, 304–316. [Google Scholar] [CrossRef]
- Amina, H.; Ahmad, M.; Bhatti, G.R.; Zafar, M.; Sultana, S.; Butt, M.A.; Bahadur, S.; Haq, I.U.; Ghufran, M.A.; Ahmad, S. Microscopic investigation of pollen morphology of Brassicaceae from Central Punjab-Pakistan. Microsc. Res. Tech. 2020, 83, 446–454. [Google Scholar] [CrossRef]
- Perveen, A.; Qaiser, M. Pollen flora of Pakistan-LXIX. Poaceae. Pak. J. Bot. 2012, 44, 747–756. [Google Scholar]
- Khan, K.; Ahmad, M.; Zafar, M.; Malik, K.; Sultana, S.; Ahmad, S.; Khan, F.; Ullah, K. Palyno-Morphological Study of Weedy Melliferous (Bee Visited) Plants Using Light Microscopic Techniques From Southern Khyber Pakhtunkhwa, Pakistan. Pak. J. Weed Sci. Res. 2021, 27, 163–172. [Google Scholar] [CrossRef]
- Butt, M.A.; Zafar, M.; Ahmad, M.; Kayani, S.; Bahadur, S.; Ullah, F.; Khatoon, S. The use of taxonomic studies to the identification of wetlands weeds. Adv. Weed Sci. 2021, 39, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Bolick, M.R. Taxonomic, evolutionary, and functional considerations of Compositae pollen ultrastructure and sculpture. Plant Syst. Evol. 1978, 130, 209–218. [Google Scholar] [CrossRef]
- Aftab, R.; Perveen, A. A palynological study of some cultivated trees from Karachi. Pak. J. Bot. 2006, 38, 15–28. [Google Scholar]
- Usma, A.; Ahmad, M.; Ramadan, M.F.; Khan, A.M.; Zafar, M.; Hamza, M.; Sultana, S.; Yaseen, G. Micro-morphological diversity of pollen among Asteraceous taxa from Potohar Plateau-Pakistan. Microsc. Res. Tech. 2022, 85, 2467–2485. [Google Scholar] [CrossRef] [PubMed]
- Ullah, F.; Ullah, A.; Sohail, A.; Khan, R. Diversity, distribution and ecological importance of weeds in the maize crop at Maidan Valley, Dir (l), Khyber Pakhtunkhwa, Pakistan. Pak. J. Weed Sci. Res. 2015, 21, 543–553. [Google Scholar]
- Meo, A.; Hafiz, H.; Baig, I. Pollen grain description of 13 non allergic and four allergic Graminaceous species. J. Pure Appl. Sci. 1989, 8, 19–26. [Google Scholar]
- Perveen, A.; Qaiser, M.; Khan, R. Pollen Flora of Pakistan–XLII. Brassicaceae. Pak. J. Bot. 2004, 36, 683–700. [Google Scholar]
- Appel, O.; Al-Shehbaz, I. Cruciferae. In Flowering Plants Dicotyledons; Springer: Berlin/Heidelberg, Germany, 2003; pp. 75–174. [Google Scholar]
- Moore, P.D.; Webb, J.A.; Collison, M.E. Pollen Analysis; Blackwell Scientific Publications: Hoboken, NJ, USA, 1991. [Google Scholar]
- Brochmann, C. Polyploid evolution in arctic-alpine (Brassicaceae). Sommerfeltia 1992, 4, 1–44. [Google Scholar] [CrossRef]
- Al-Shehbaz, I.; Beilstein, M.; Kellogg, E. Systematics and phylogeny of the Brassicaceae (Cruciferae): An overview. Plant Syst. Evol. 2006, 259, 89–120. [Google Scholar] [CrossRef]
- Khan, M.; Kakar, S.; Marwat, K.; Khan, I. Differential response of Zea mays L. in relation to weed control and different macronutrient combinations. Sains Malays. 2013, 42, 1405–1411. [Google Scholar]
- Williams, N.M.; Kremen, C. Resource distributions among habitats determine solitary bee offspring production in a mosaic landscape. Ecol. Appl. 2007, 17, 910–921. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, M.; Khan, S.M.; Khan, M.A.; Ahmad, Z.; Abbas, Z.; Khan, S.M.; Khan, M.S. Distribution pattern and species richness of natural weeds of wheat in varying habitat conditions of district Malakand, Pakistan. Pak. J. Bot. 2017, 49, 2371–2382. [Google Scholar]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sr. No | Plant Taxa | Family | Locality | Voucher Specimen Number |
|---|---|---|---|---|
| 1. | Achyranthus aspera L. | Amaranthaceae | Salt Range Chakwal | AU-107 |
| 2. | Aerva javanica (Burm.f.) Juss. ex Schult. | Amaranthaceae | Miani Village Talagang | AU-113 |
| 3. | Amaranthus graecizans L. | Amaranthaceae | Khewra mines | AU-115 |
| 4. | Asphodelus tenuifolius Cav. | Asphodelaceae | Uchali Lake | AU-99 |
| 5. | Atriplex stocksii Boiss. | Amaranthaceae | Uchali Lake | AU-77 |
| 6. | Boerhavia procumbens Banks ex Roxb. | Nyctaginaceae | Rawalpindi | AU-65 |
| 7. | Brassica furatii Mouterde | Brassicaceae | Musa khel Mianwali | AU-39 |
| 8. | Capparis decidua (Forssk.) Edgew. | Capparaceae | Miani Village | AU-55 |
| 9. | Celosia argentea L. | Amaranthaceae | Kabkbi Lake | AU-110 |
| 10. | Cenchrus ciliaris L. | Poaceae | Islamabad | AU-205 |
| 11. | Chenopodium murale L. | Chenopodiaceae | Salt Range | AU-209 |
| 12. | Corchorus depressus (L.) Stocks | Tiliaceae | Attock | AU-211 |
| 13. | Cissampelos pareira L. | Menispermaceae | Salt Range | AU-215 |
| 14. | Citharexylum spinosum L. | Verbenaceae | Islamabad | AU-217 |
| 15. | Cleome brachycarpa (Forssk.) Vahl ex DC | Capparaceae | Miani village Talagang | AU-31 |
| 16. | Cleome viscosa L. | Capparaceae | Miani village Talagang | AU-45 |
| 17. | Cucumis melo subsp. agrestis (Naudin) Pangalo. | Cucurbitaceae | Miani village Talagang | AU-53 |
| 18. | Euphorbia granulata Forssk. | Euphorbiaceae | Thoha Bahadur Chakwal | AU-69 |
| Sr. No | Taxa | Shape | Aperture Condition | Mesocolpium | Exine Ornamentation | Colpi Orientation |
|---|---|---|---|---|---|---|
| 1. | Achyranthus aspera | Oblate-spheroidal | Tricolporate | Scabrate | Scabrate | Sunken, angular |
| 2. | Aerva javanica | Prolate-spheroidal | Tricolporate | Aerolate | Reticulate | Sunken, angular, margins distinct, and ends tapering |
| 3. | Amaranthus graecizans | Prolate-spheroidal | Trizonocolporate | Reticulate | Scabrate-reticulate | Prominent and rounded at ends |
| 4. | Asphodelus tenuifolius | Oblate-spheroidal | Tricolporate | Scabrate-reticulate | Aerolate | Sunken, slit-like margins, and end tapering |
| 5. | Atriplex stocksii | Oblate | Tricolporate | Scabrate | Reticulate-Perforate | Prominent and rounded at ends |
| 6. | Boerhavia procumbens | Oblate-spheroidal | Tricolporate | Reticulate | Scabrate | Sunken, angular, margins distinct |
| 7. | Brassica furatii | Spherical | Tricolporate | Scabrate | Echinate | Sunken, further divided into three slit-like portions, ends are tapering |
| 8. | Capparis decidua | Oblate-spheroidal | - | Aerolate | Faveolate | Sunken, slit-like, margins wavy and slightly pointed at ends |
| 9. | Celosia argentea | Oblate-spheroidal | Tricolporate | Perforate | Scabrate | Sunken, slit-like, margins distinct, and pointed at ends |
| 10. | Cenchrus ciliaris | Prolate-spheroidal | - | Scabrate | Reticulate | Prominent margins |
| 11. | Chenopodium murale | Oblate-spheroidal | Tricolporate | Reticulate | Aerolate | Sunken, long and slit-like |
| 12. | Corchorus depressus | Oblate-spheroidal | Not visible | Scabrate | Reticulate-scabrate | Prominent and tapering ends |
| 13. | Cissampelos pareira | Sub-oblate | Tricolporate | Reticulate | Perforate | Sunken, angular, margins distinct |
| 14. | Citharexylum spinosum | Oblate-spheroidal | Tricolporate | Aerolate-Perforate | Scabrate | Long and slit-like, tapering ends |
| 15. | Cleome brachycarpa | Oblate-spheroidal | Tricolporate | Reticulate | Reticulate | Sunken and tapering ends |
| 16. | Cleome viscosa | Prolate-spheroidal | Tricolporate | Scabrate | Scabrate-reticulate | Long and slit-like margins |
| 17. | Cucumis melo subsp. agrestis | Oblate-spheroidal | Not Visible | Perforate | Perforate | Sunken, slit-like, wavy margins |
| 18. | Euphorbia granulata | Prolate-spheroidal | Tricolporate | Scabrate-reticulate | Aerolate | Sunken, wavy, and margins distinct |
| Plant Name | Exine Thickness Mean (Min-Max) S.E µm | Polar Diameter Mean (Min-Max) S.E µm | Equatorial Diameter Mean (Min-Max) S.E µm | P/E Ratio | No of Pores | Colpi Length Mean (Min-Max) S.E µm | Colpi Width Mean (Min-Max) S.E µm | Pores Length Mean (Min-Max) S.E µm | Pores Width Mean (Min-Max) S.E µm |
|---|---|---|---|---|---|---|---|---|---|
| Achyranthus aspera | 1.56 (1.35–1.95) ± 0.11 | 14.5 (12.1–15.6) ± 0.63 | 15.6 (12.3–18.0) ± 0.96 | 0.93 | – | 3.81 (1.65–7.95) ± 3.81 | 2.55 (1.50–4.50) ± 0.53 | – | – |
| Aerva javanica | 1.80 (1.35–2.55) ± 0.23 | 18.03 (16.8–18.6) ± 0.34 | 17.8 (15.3–19.6) ± 0.84 | 1.01 | – | 6.12 (1.65–10.20) ± 1.40 | 3.45 (1.95–7.20) ± 0.97 | – | – |
| Amaranthus graecizans | 1.50 (1.20–1.80) ± 0.11 | 28.8 (25.5–33.15) ± 1.65 | 27.0 (25.6–30.3) ± 1.2 | 1.07 | – | 10.05 (4.05–17.55) ± 2.17 | 6.24 (2.85–13.20) ± 1.92 | – | – |
| Asphodelus tenuifolius | 6.12 (1.65–10.20) ± 1.40 | 3.45 (1.95–7.20) ± 0.97 | 6.12 (1.6–10.20) ± 1.40 | 0.97 | – | 12.2 (10.0–15.1) ± 2.44 | 5.79 (3.00–8.55) ± 2.31 | – | – |
| Atriplex stocksii | 2.01 (1.80–2.10) ± 0.10 | 11.7 (11.4–11.9) ± 0.15 | 17.3 (17.2–17.5) ± 0.09 | 0.67 | – | 12.30 (9.30–15.10) ± 1.80 | 7.20 (5.70–9.20) ± 1.03 | – | – |
| Boerhavia procumbens | 0.6 (0.5–0.75) ± 0.06 | 13.3 (12.2–15.25) ± 0.5 | 13.5 (12.2–15.5) ± 0.54 | 0.98 | – | 4.2 (3.2–5.00) ± 0.52 | 5.4 (4.9–6.3) ± 0.45 | – | – |
| Brassica furatii | 1.55 (1.0–2.0) ± 0.16 | 23.9 (22.75–25.5) ± 0.52 | 23.9 (22.2–25.5) ± 0.72 | 1.00 | – | 5.5 (4.6–6.7) ± 0.62 | 2.9 (2.3–3.5) ± 0.34 | – | – |
| Capparis decidua | 1.05 (0.75–1.25) ± 0.09 | 13.9 (12.0–15.50) ± 0.63 | 13.4 (12.0–15.5) ± 0.70 | 0.99 | – | 5.2 (4.2–6.1) ± 0.15 | 6.4 (5.7–7.3) ± 0.15 | – | – |
| Celosia argentea | 2.5 (2.25–2.75) ± 0.11 | 21.0 (15.5–24.75) ± 1.63 | 22.6 (18.0–25.2) ± 1.3 | 0.92 | – | 7.8 (6.8–9.1) ± 0.9 | 4.1 (3.4–5.1) ± 0.4 | – | – |
| Cenchrus ciliaris | 2.65 (2.25–3.0) ± 0.12 | 15.3 (14.7–15.75) ± 10.16 | 14.8 (14.0–15.7) ± 0.3 | 1.03 | 1 | 4.8 (4.4–5.4) ± 0.29 | 2.7 (2.4–3.1) ± 0.2 | 7.00 (5.25–8.50) ± 0.54 | 4.05 (3.00–5.00) ± 0.40 |
| Chenopodium murale | 1.9 (1.0–2.75) ± 0.34 | 14.0 (13.0–14.7) ± 0.32 | 14.5 (13.5–15.2) ± 0.3 | 0.96 | – | 6.7 (5.8–8.1) ± 0.6 | 3.1 (2.4–4.00) ± 0.4 | – | – |
| Corchorus depressus | 2.95 (2.75–3.25) ± 0.09 | 36.9 (33.7–43.00) ± 1.74 | 38.7 (37.2–40.5) ± 0.65 | 0.95 | – | 7.8 (5.8–9.5) ± 1.07 | 4.3 (3.7–5.00) ± 0.3 | – | – |
| Cissampelos pareira | 2.80 (2.25–3.25) ± 0.16 | 40.4 (35.7–45.50) ± 1.68 | 47.2 (45.2–48.5) ± 0.65 | 0.85 | – | 1.86 (1.35–2.40) ± 0.17 | 1.23 (.75–1.95) ± 0.21 | – | – |
| Citharexylum spinosum | 0.78 (0.6–1.1) ± 0.09 | 27.3 (22.7–32.7) ± 1.65 | 30.0 (24.5–33.5) ± 1.77 | 0.91 | – | 1.65 (1.05–2.40) ± 0.22 | 1.68 (1.05–2.10) ± 0.20 | – | – |
| Cleome brachycarpa | 2.5 (2.25–2.75) ± 0.11 | 34.05 (29.0–39.0) ± 1.69 | 35.1 (32.2–39.2) ± 1.32 | 0.96 | – | 2.25 (0.75–3.45) ± 0.47 | 2.64 (2.10–3.30) ± 0.20 | – | – |
| Cleome viscosa | 2.15 (1.5–3.0) ± 0.3 | 16.2 (13.5–18.0) ± 0.85 | 15.6 (13.2–18.5) ± 1.0 | 1.03 | – | 2.70 (1.95–3.30) ± 0.22 | 3.03 (2.40–3.60) ± 0.20 | – | – |
| Cucumis melo subsp. agrestis | 0.95 (0.75–1.25) ± 0.09 | 13.9 (12.0–15.5) ± 0.63 | 13.4 (12.0–15.5) ± 0.70 | 0.99 | – | 1.14 (0.75–1.50) ± 0.13 | 1.77 (1.50–2.10) ± 0.12 | – | – |
| Euphorbia granulata | 2.0 (1.50–2.25) ± 0.13 | 19.8 (17.2–21.25) ± 0.69 | 18.2 (16.2–20.5) ± 0.69 | 1.08 | 1 | – | – |
| Sr. No | Taxa | No. of Fertile Pollen | No. of Sterile Pollen | Fertility % | Sterility % |
|---|---|---|---|---|---|
| 1. | Achyranthus aspera | 70 | 15 | 82 | 17 |
| 2. | Aerva javanica | 90 | 40 | 69 | 30 |
| 3. | Amaranthus graecizans | 77 | 10 | 88 | 11 |
| 4. | Asphodelus tenuifolius | 99 | 12 | 89 | 10 |
| 5. | Atriplex stocksii | 88 | 22 | 80 | 20 |
| 6. | Boerhavia procumbens | 50 | 9 | 84 | 15 |
| 7. | Brassica furatii | 80 | 2 | 97 | 2 |
| 8. | Capparis decidua | 80 | 7 | 91 | 8 |
| 9. | Celosia argentea | 75 | 10 | 88 | 11 |
| 10. | Cenchrus ciliaris | 90 | 11 | 89 | 10 |
| 11. | Chenopodium murale | 118 | 7 | 94 | 5 |
| 12. | Corchorus depressus | 102 | 10 | 91 | 8 |
| 13. | Cissampelos pareira | 90 | 5 | 95 | 5 |
| 14. | Citharexylum spinosum | 110 | 14 | 89 | 11 |
| 15. | Cleome brachycarpa | 86 | 11 | 89 | 11 |
| 16. | Cleome viscosa | 99 | 12 | 89 | 10 |
| 17. | Cucumis melosubsp. agrestis | 143 | 9 | 94 | 5 |
| 18. | Euphorbia granulata | 81 | 14 | 85 | 14 |
| Link Characters | Present (+) /Absent (−) | Diagnostic Characters | Species Name |
|---|---|---|---|
| 1 | + | pollen shape oblate-spheroidal, mesocolpium scabrate, colpi orientation sunken, angular | Achyranthus aspera |
| − | pollen shape prolate-spheroidal, mesocolpium aerolate, colpi orientation Sunken, angular, margins distinct, and ends tapering | 2 | |
| 2 | + | aperture tricolporate, exine reticulate | Aerva javanica |
| − | aperture trizonocolporate, exine scabrate-reticulate | 3 | |
| 3 | + | The shape of pollen prolate-spheroidal, mescolopium reticulate | Amaranthus graecizans |
| − | pollen shape oblate-spheroidal, mescolopium scabrate-reticulate | 4 | |
| 4 | + | aperture tricolporate, colpi sunken, slit like margins, and end tapering | Asphodelus tenuifolius |
| − | aperture tricolporate, colpi prominent, and rounded at ends | 5 | |
| 5 | + | shape oblate, mescolopium scabrate | Atriplex stocksii |
| − | Shape oblate-spheroidal, mescolopium reticulate | 6 | |
| 6 | + | exine scabrate, colpi Sunken, angular, margins distinct | Boerhavia procumbens |
| − | exine echinate, colpi Sunken, further divided into three slit-like portions, ends are tapering | 7 | |
| 7 | + | pollen shape spherical, mescolopium scabrate | Brassica furatii |
| − | pollen shape oblate spheroidal, mescolpium aerolate | 8 | |
| 8 | + | exine faveolate, colpi sunken slit-like, margins wavy, and slightly pointed at the end | Capparis decidua |
| − | pollen shape oblate spheroidal, exine scabrate | 9 | |
| 9 | + | aperture tricolporate, mesocolpium perforate | Celosia argentea |
| − | aperture not clear, mescolpium scabrate | 10 | |
| 10 | + | pollen shape prolate-spheroidal, exine reticulate | Cenchrus ciliaris |
| − | pollen shape oblate-spheroidal, exine aerolate | 11 | |
| 11 | + | aperture tricolporate, colpi sunken, long and slit like | Chenopodium murale |
| − | aperture not visible, colpi prominent, and tapering | 12 | |
| 12 | + | pollen shape oblate-spheroidal, mesocolpium scabrate | Corchorus depressus |
| − | pollen shape sub-oblate, mescolpium perforate | 13 | |
| 13 | + | aperture tricolporate, exine sunken, angular, margins distinct | Cissampelos pareira |
| − | aperture tricolporate, exine long and slit like, tapering ends | 14 | |
| 14 | + | pollen shape oblate-spheroidal, mesocolpium aerolate-perforrate | Citharexylum spinosum |
| − | pollen shape oblate-spheroidal, mescolpium reticulate | 15 | |
| 15 | + | exine reticulate, colpi sunken, and tapering ends | Cleome brachycarpa |
| − | exine scabrate-reticulate, colpi long and slit-like margins | 16 | |
| 16 | + | pollen shape prolate-spheroidal, aperture tricolporate | Cleome viscosa |
| − | pollen shape oblate-spheroidal, aperture not visible | 17 | |
| 17 | + | mesocolpium perforate, exine perforate | Cucumis melo subsp. agrestis |
| − | mescolpium scabrate-reticulate, exine aerolate | Euphorbia granulata |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Usma, A.; Ahmad, M.; Zafar, M.; Sultana, S.; Ullah, F.; Saqib, S.; Ayaz, A.; Zaman, W. Palynological Study of Weed Flora from Potohar Plateau. Agronomy 2022, 12, 2500. https://doi.org/10.3390/agronomy12102500
Usma A, Ahmad M, Zafar M, Sultana S, Ullah F, Saqib S, Ayaz A, Zaman W. Palynological Study of Weed Flora from Potohar Plateau. Agronomy. 2022; 12(10):2500. https://doi.org/10.3390/agronomy12102500
Chicago/Turabian StyleUsma, Anwer, Mushtaq Ahmad, Muhammad Zafar, Shazia Sultana, Fazal Ullah, Saddam Saqib, Asma Ayaz, and Wajid Zaman. 2022. "Palynological Study of Weed Flora from Potohar Plateau" Agronomy 12, no. 10: 2500. https://doi.org/10.3390/agronomy12102500