
24-Epibrassnolide Alleviates the Adverse Effect of Salinity on Rice Grain Yield through Enhanced Antioxidant Enzyme and Improved K+/Na+ Homeostasis
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Experimental Design
2.2. Determination of Germination Indicators
2.3. Determination of LAI and Leaf Area Decay Rate
2.4. Leaf Na+ and K+ Content
2.5. Leaf Antioxidant Enzyme Activities, MDA, and Proline Content
2.6. Leaf Photosynthetic Potential, Crop Growth Rate, and Net Assimilation Rate
2.7. Rice Grain Yield and Yield Components
2.8. Statistical Analysis
3. Results
3.1. Rice Seed Germination Indicators (Germination Experiment)
3.2. Rice Seedling Morphological Traits (Seedling Experiment)
3.3. Rice Plant Height (Pot Experiment)
3.4. Rice Leaf Area Index, Efficient LAI, and Leaf Area Decay Rate
3.5. Rice Plant Dry Weight at Heading and Full-Ripening Stage
3.6. The Photosynthetic Potential, Crop Growth Rate, and Net Assimilation Rate of Rice
3.7. Rice Grain Yield, Yield Components, and Panicle Traits
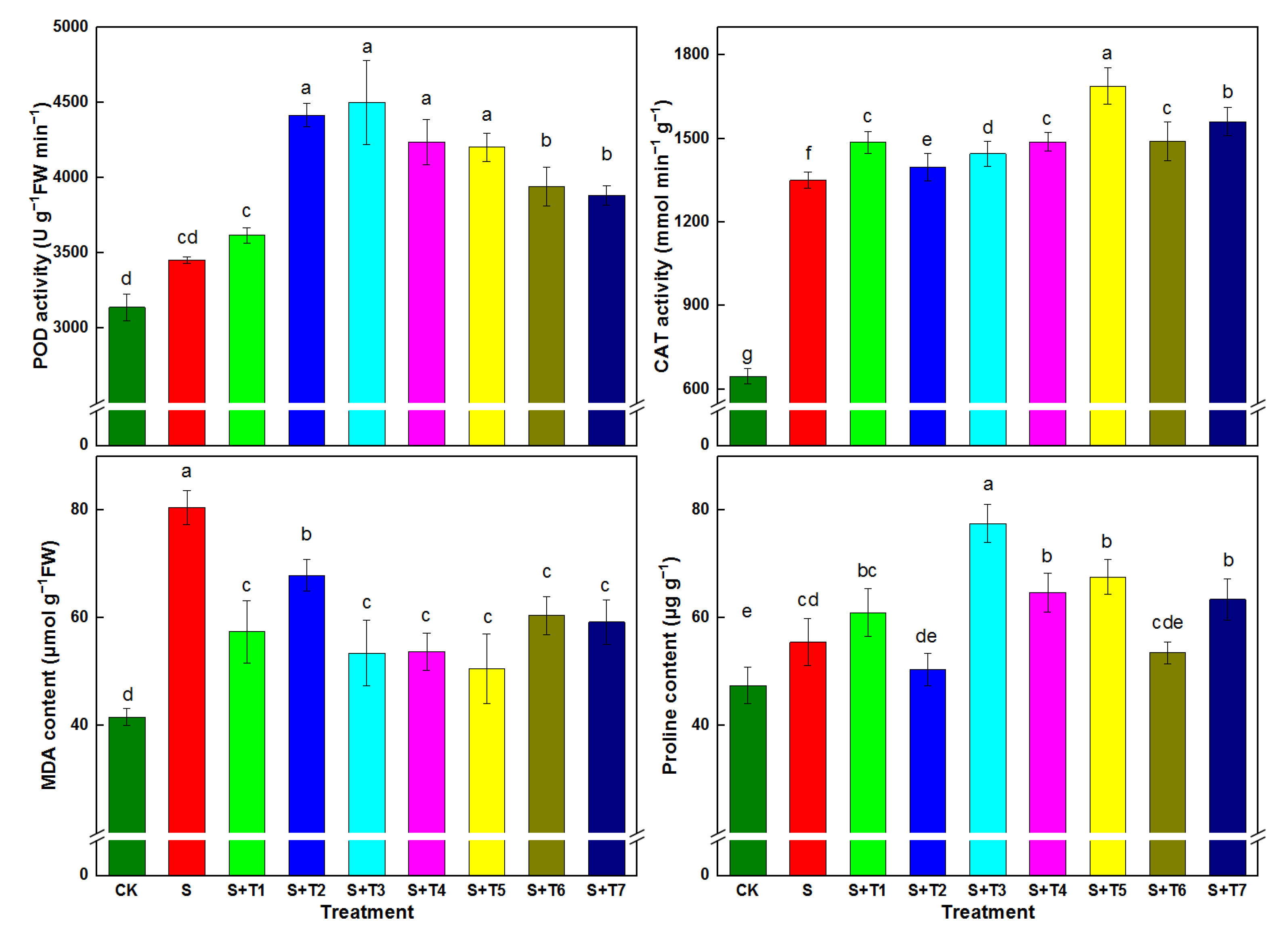
3.8. Leaf Antioxidant Enzyme Activities, MDA, and Proline Content
3.9. Na+ and K+ Content in Rice Organs
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hossain, M.S. Present Scenario of Global Salt Affected Soils, its Management and Importance of Salinity Research. Int. J. Biol. Sci. 2019, 1, 1–3. [Google Scholar]
- Chen, Y.; Liu, Y.; Ge, J.; Li, R.; Zhang, R.; Zhang, Y.; Huo, Z.; Xu, K.; Wei, H.; Dai, Q. Improved physiological and morphological traits of root synergistically enhanced salinity tolerance in rice under appropriate nitrogen application rate. Front. Plant Sci. 2022, 13, 982637. [Google Scholar] [CrossRef]
- Munns, R.; James, R.A.; Luchli, A. Approaches to increasing the salt tolerance of wheat and other cereals. J. Exp. Bot. 2006, 57, 1025–1043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prittesh, P.; Avnika, P.; Kinjal, P.; Jinal, H.N.; Amaresan, N. Amelioration effect of salt-tolerant plant growth-promoting bacteria on growth and physiological properties of rice (Oryza sativa) under salt-stressed conditions. Arch. Microbiol. 2020, 202, 2419–2428. [Google Scholar] [CrossRef] [PubMed]
- Soltabayeva, A.; Ongaltay, A.; Omondi, J.O.; Srivastava, S. Morphological, Physiological and Molecular Markers for Salt-Stressed Plants. Plants 2021, 10, 243. [Google Scholar] [CrossRef]
- Hussain, S.; Chu, Z.; Bai, Z.; Cao, X.; Zhu, L.; Hussain, A.; Zhu, C.; Fahad, S.; James, A.B.; Zhang, J. Effects of 1-Methylcyclopropene on Rice Growth Characteristics and Superior and Inferior Spikelet Development Under Salt Stress. J. Plant Growth Regul. 2018, 37, 1368–1384. [Google Scholar] [CrossRef]
- Grove, M.D.; Spencer, G.F.; Rohwedder, W.K.; Mandava, N.; Worley, J.F.; Warthen, J.D.; Steffens, G.L.; Flippen-Anderson, J.L.; Cook, J.C. Brassinolide, a plant growth-promoting steroid isolated from Brassica napus pollen. Nature 1979, 281, 216–217. [Google Scholar] [CrossRef]
- Sairam, R.K. Effects of homobrassinolide application on plant metabolism and grain yield under irrigated and moisture-stress conditions of two wheat varieties. Plant Growth Regul. 1994, 14, 173–181. [Google Scholar] [CrossRef]
- Bajguz, A. Effect of brassinosteroids on nucleic acids and protein content in cultured cells of Chlorella vulgaris. Plant Physiol. Biochem. 2000, 38, 209–215. [Google Scholar] [CrossRef]
- Tanveer, M.; Shahzad, B.; Sharma, A.; Khan, E.A. 24-Epibrassinolide application in plants: An implication for improving drought stress tolerance in plants. Plant Physiol. Biochem. 2019, 135, 295–303. [Google Scholar] [CrossRef]
- Shahzad, B.; Tanveer, M.; Che, Z.; Rehman, A.; Cheema, S.A.; Sharma, A.; Song, H.; Rehman, S.U.; Zhaorong, D. Role of 24-epibrassinolide (EBL) in mediating heavy metal and pesticide induced oxidative stress in plants: A review. Ecotoxicol. Environ. Saf. 2018, 147, 935–944. [Google Scholar] [CrossRef] [PubMed]
- da Silva Cunha, L.F.; de Oliveira, V.P.; do Nascimento, A.W.S.; da Silva, B.R.S.; Batista, B.L.; Alsahli, A.A.; Lobato, A. Leaf application of 24-epibrassinolide mitigates cadmium toxicity in young Eucalyptus urophylla plants by modulating leaf anatomy and gas exchange. Physiol. Plant 2021, 173, 67–87. [Google Scholar] [CrossRef] [PubMed]
- Anwar, A.; Bai, L.; Miao, L.; Liu, Y.; Li, S.; Yu, X.; Li, Y. 24-Epibrassinolide Ameliorates Endogenous Hormone Levels to Enhance Low-Temperature Stress Tolerance in Cucumber Seedlings. Int. J. Mol. Sci. 2018, 19, 2497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galal, A. 24-epibrassinolide application enhances growth and biochemical aspects of squash under salt stress conditions. Acta Biol. Hung. 2018, 69, 182–196. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Zhang, Q.; Ervin, E.H.; Yang, Z.; Zhang, X. Physiological Mechanism of Enhancing Salt Stress Tolerance of Perennial Ryegrass by 24-Epibrassinolide. Front. Plant Sci. 2017, 8, 1017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soliman, M.; Elkelish, A.; Souad, T.; Alhaithloul, H.; Farooq, M. Brassinosteroid seed priming with nitrogen supplementation improves salt tolerance in soybean. Physiol. Mol. Biol. Plants 2020, 26, 501–511. [Google Scholar] [CrossRef]
- Wani, A.S.; Tahir, I.; Ahmad, S.S.; Dar, R.A.; Nisar, S. Efficacy of 24-epibrassinolide in improving the nitrogen metabolism and antioxidant system in chickpea cultivars under cadmium and/or NaCl stress. Sci. Hortic. 2017, 225, 48–55. [Google Scholar] [CrossRef]
- Li, W.; Sun, J.; Zhang, X.; Ahmad, N.; Hou, L.; Zhao, C.; Pan, J.; Tian, R.; Wang, X.; Zhao, S. The Mechanisms Underlying Salt Resistance Mediated by Exogenous Application of 24-Epibrassinolide in Peanut. Int. J. Mol. Sci. 2022, 23, 6376. [Google Scholar] [CrossRef]
- Özdemir, F.; Bor, M.; Demiral, T.; Türkan, İ. Effects of 24-epibrassinolide on seed germination, seedling growth, lipid peroxidation, proline content and antioxidative system of rice (Oryza sativa L.) under salinity stress. Plant Growth Regul. 2004, 42, 203–211. [Google Scholar] [CrossRef]
- Anuradha, S.; Rao, S. Effect of brassinosteroids on salinity stress induced inhibition of seed germination and seedling growth of rice (Oryza sativa L.). Plant Growth Regul. 2001, 33, 151–153. [Google Scholar] [CrossRef]
- Sharma, I.; Ching, E.; Saini, S.; Bhardwaj, R.; Pati, P.K. Exogenous application of brassinosteroid offers tolerance to salinity by altering stress responses in rice variety Pusa Basmati-1. Plant Physiol. Biochem. 2013, 69, 17–26. [Google Scholar] [CrossRef] [PubMed]
- Yan, F.; Wei, H.; Li, W.; Liu, Z.; Tang, S.; Chen, L.; Ding, C.; Jiang, Y.; Ding, Y.; Li, G. Melatonin improves K+ and Na+ homeostasis in rice under salt stress by mediated nitric oxide. Ecotox. Environ. Saf. 2020, 206, 111358. [Google Scholar] [CrossRef] [PubMed]
- Jiang, D.; Hou, J.; Gao, W.; Tong, X.; Li, M.; Chu, X.; Chen, G. Exogenous spermidine alleviates the adverse effects of aluminum toxicity on photosystem II through improved antioxidant system and endogenous polyamine contents. Ecotox. Environ. Saf. 2021, 207, 111265. [Google Scholar] [CrossRef] [PubMed]
- Panda, A.; Rangani, J.; Kumari, A.; Parida, A.K. Efficient regulation of arsenic translocation to shoot tissue and modulation of phytochelatin levels and antioxidative defense system confers salinity and arsenic tolerance in the Halophyte Suaeda maritima. Environ. Exp. Bot. 2017, 143, 149–171. [Google Scholar] [CrossRef]
- Kramer, G.F.; Norman, H.A.; Krizek, D.T.; Mirecki, R.M. Influence of UV-B radiation on polyamines, lipid peroxidation and membrane lipids in cucumber. Phytochemistry 1991, 30, 2101–2108. [Google Scholar] [CrossRef]
- Kishor, P.; Hong, Z.; Miao, G.H.; Hu, C.; Verma, D. Overexpression of [delta]-Pyrroline-5-Carboxylate Synthetase Increases Proline Production and Confers Osmotolerance in Transgenic Plants. Plant Physiol. 1995, 108, 1387–1394. [Google Scholar] [CrossRef] [Green Version]
- Darini, M.T. Response, correlation of growth analysis and shallot yield under different nitrogen fertilizer dosage and rhizobacteria source. J. Agron. 2020, 19, 131–137. [Google Scholar]
- Larré, C.F.; Moraes, D.M.d.; Lopes, N.F. Qualidade fisiológica de sementes de arroz tratadas com solução salina e 24-epibrassinolídeo X1—Physiological quality of rice seeds treated with saline solution and 24-epibrassinolide. Rev. Bras. Sementes 2011, 33, 86–94. [Google Scholar] [CrossRef]
- Ganie, S.A.; Molla, K.A.; Henry, R.J.; Bhat, K.V.; Mondal, T.K. Advances in understanding salt tolerance in rice. Theor. Appl. Genet. 2019, 132, 851–870. [Google Scholar] [CrossRef]
- Hussain, M.; Ahmad, S.; Hussain, S.; Lal, R.; Ul-Allah, S.; Nawaz, A. Chapter Six—Rice in Saline Soils: Physiology, Biochemistry, Genetics, and Management. In Advances in Agronomy; Sparks, D.L., Ed.; Academic Press: Cambridge, MA, USA, 2018; Volume 148, pp. 231–287. [Google Scholar]
- Shahid, M.A.; Balal, R.M.; Pervez, M.A.; Abbas, T.; Aqeel, M.A.; Riaz, A.; Mattson, N.S. Exogenous 24-Epibrassinolide Elevates the Salt Tolerance Potential of Pea (Pisum sativum L.) by Improving Osmotic Adjustment Capacity and Leaf Water Relations. J. Plant Nutr. 2015, 38, 1050–1072. [Google Scholar] [CrossRef]
- Kolomeichuk, L.V.; Efimova, M.V.; Zlobin, I.E.; Kreslavski, V.D.; Murgan, O.K.; Kovtun, I.S.; Khripach, V.A.; Kuznetsov, V.V.; Allakhverdiev, S.I. 24-Epibrassinolide alleviates the toxic effects of NaCl on photosynthetic processes in potato plants. Photosynth. Res. 2020, 146, 151–163. [Google Scholar] [CrossRef] [PubMed]
- Saini, S.; Kaur, N.; Pati, P.K. Reactive oxygen species dynamics in roots of salt sensitive and salt tolerant cultivars of rice. Anal. Biochem. 2018, 550, S0003269718301982. [Google Scholar] [CrossRef] [PubMed]
- Singh, V.; Singh, A.P.; Bhadoria, J.; Giri, J.; Singh, J.; Vineeth, T.V.; Sharma, P.C. Differential expression of salt-responsive genes to salinity stress in salt-tolerant and salt-sensitive rice (Oryza sativa L.) at seedling stage. Protoplasma 2018, 255, 1667–1681. [Google Scholar] [CrossRef] [PubMed]
- Tanveer, M.; Shahzad, B.; Sharma, A.; Biju, S.; Bhardwaj, R. 24-Epibrassinolide; an active brassinolide and its role in salt stress tolerance in plants: A review. Plant Physiol. Biochem. 2018, 130, 69–79. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Li, R.; Ge, J.; Liu, J.; Wang, W.; Xu, M.; Zhang, R.; Hussain, S.; Wei, H.; Dai, Q. Exogenous melatonin confers enhanced salinity tolerance in rice by blocking the ROS burst and improving Na+/K+ homeostasis. Environ. Exp. Bot. 2021, 189, 104530. [Google Scholar] [CrossRef]
- Szabados, L.; Savouré, A. Proline: A multifunctional amino acid. Trends Plant Sci. 2010, 15, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Hayat, S.; Hayat, Q.; Alyemeni, M.N.; Wani, A.S.; Pichtel, J.; Ahmad, A. Role of proline under changing environments: A review. Plant Signal. Behav. 2012, 7, 1456–1466. [Google Scholar] [CrossRef] [Green Version]
- Hussain, M.; Khan, T.A.; Yusuf, M.; Fariduddin, Q. Silicon-mediated role of 24-epibrassinolide in wheat under high-temperature stress. Environ. Sci. Pollut. Res. Int. 2019, 26, 17163–17172. [Google Scholar] [CrossRef]
- López-Gómez, M.; Hidalgo-Castellanos, J.; Lluch, C.; Herrera-Cervera, J.A. 24-Epibrassinolide ameliorates salt stress effects in the symbiosis Medicago truncatula-Sinorhizobium meliloti and regulates the nodulation in cross-talk with polyamines. Plant Physiol. Biochem. 2016, 108, 212–221. [Google Scholar] [CrossRef]
- Wang, L.; Ruan, Y.-L. New insights into roles of cell wall invertase in early seed development revealed by comprehensive spatial and temporal expression patterns of GhCWIN1 in cotton. Plant Physiol. 2012, 160, 777–787. [Google Scholar] [CrossRef] [Green Version]
- Wu, H.; Zhu, M.; Shabala, L.; Zhou, M.; Shabala, S. K+ retention in leaf mesophyll, an overlooked component of salinity tolerance mechanism: A case study for barley. J. Integr. Plant Biol. 2015, 57, 171–185. [Google Scholar] [CrossRef] [PubMed]
- Qin, H.; Huang, R. The phytohormonal regulation of Na+/K+ and reactive oxygen species homeostasis in rice salt response. Mol. Breed. 2020, 40, 47. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Salt Concentration (%) | EBR Concentration (mg L−1) | EBR Spraying Period | Denotation |
|---|---|---|---|
| 0 | 0 | / | CK |
| 0.3 | 0 | / | S |
| 0.3 | 0.5 | Transplanting | S+T1 |
| 0.3 | 0.5 | Panicle initiation | S+T2 |
| 0.3 | 0.5 | Heading | S+T3 |
| 0.3 | 0.5 | Transplanting + panicle initiation | S+T4 |
| 0.3 | 0.5 | Transplanting + heading | S+T5 |
| 0.3 | 0.5 | Panicle initiation + heading | S+T6 |
| 0.3 | 0.5 | Transplanting + panicle initiation + heading | S+T7 |
| Salt Concentration (mmol L−1) | EBR Concentration (mg L−1) | GE (%) | Relative GE | GC (%) | Relative GC | GI |
|---|---|---|---|---|---|---|
| 0 | 0 | 72.5 b | 100.0 b | 94.6 a | 100.0 a | 21.5 c |
| 0.05 | 80.8 a | 111.4 a | 95.8 a | 101.3 a | 22.6 ab | |
| 0.1 | 78.3 a | 108.1 a | 92.9 ab | 98.3 ab | 22.1 ab | |
| 0.5 | 79.2 a | 109.2 a | 96.3 a | 101.8 a | 22.7 a | |
| 1.0 | 79.6 a | 109.9 a | 93.3 ab | 98.7 ab | 22.0 bc | |
| 100 | 0 | 23.3 f | 32.1 f | 77.5 f | 81.9 f | 14.0 g |
| 0.05 | 33.8 e | 46.6 e | 82.1 e | 86.8 e | 16.3 f | |
| 0.1 | 38.3 de | 53.0 d | 86.7 cd | 91.6 cd | 17.6 e | |
| 0.5 | 46.7 c | 64.4 c | 90.0 bc | 95.2 bc | 18.6 d | |
| 1.0 | 42.1 cd | 58.1 d | 88.3 cd | 93.4 cd | 17.7 e | |
| 150 | 0 | 7.9 h | 10.9 h | 66.3 h | 70.1 h | 10.9 i |
| 0.05 | 13.8 g | 19.0 g | 71.7 g | 75.8 g | 12.1 h | |
| 0.1 | 14.2 g | 19.6 g | 76.3 f | 80.6 f | 12.7 h | |
| 0.5 | 15.4 g | 21.3 g | 86.3 cd | 91.2 cd | 14.5 g | |
| 1.0 | 14.6 g | 20.2 g | 85.4 e | 90.3 de | 14.0 g |
| Salt Concentration (%) | EBR Concentration (mg L−1) | Plant Height (cm) | Root Length (cm) | Above-Ground Fresh Weight (mg plant−1) | Under-Ground Fresh Weight (mg plant−1) | Above-Ground Dry Weight (mg plant−1) | Under-Ground Dry Weight (mg plant−1) |
|---|---|---|---|---|---|---|---|
| 0 | 0 | 14.33 a | 13.90 abc | 71.0 a | 74.3 ab | 11.8 a | 7.9 abc |
| 0.1 | 13.30 b | 12.73 e | 63.1 b | 68.4 cd | 10.2 b | 7.8 abc | |
| 0.5 | 13.98 a | 14.53 a | 70.4 a | 74.4 ab | 12.0 a | 8.7 a | |
| 1 | 12.48 cd | 14.37 ab | 67.9 a | 77.5 a | 11.4 a | 8.3 ab | |
| 0.3 | 0 | 11.63 e | 13.37 cde | 45.7 e | 65.6 de | 8.2 c | 6.8 de |
| 0.1 | 12.48 cd | 12.73 e | 51.7 d | 67.8 cd | 9.4 b | 7.1 cde | |
| 0.5 | 12.80 bc | 13.60 bcd | 53.8 cd | 71.1 bc | 9.5 b | 7.5 bcd | |
| 1 | 12.32 cde | 13.37 cde | 52.9 d | 67.3 cd | 9.2 b | 7.3 bcd | |
| 0.5 | 0 | 10.87 f | 11.30 f | 32.2 g | 54.4 f | 7.5 c | 6.4 e |
| 0.1 | 10.9 f | 12.90 de | 36.9 f | 54.5 f | 7.6 c | 6.4 e | |
| 0.5 | 11.87 de | 13.10 cde | 47.5 e | 64.9 de | 9.7 b | 7.2 cde | |
| 1 | 10.95 f | 11.03 f | 38.5 f | 62.9 e | 7.8 c | 7.0 cde |
| Treatment | LAI (Heading Stage) | Efficient LAI (Heading Stage) | LAI (Full-Ripening Stage) | Leaf Area Decay Rate (%) |
|---|---|---|---|---|
| CK | 6.23 a | 4.15 a | 3.93 a | 36.45 f |
| S | 2.15 e | 1.57 f | 1.00 f | 52.63 bc |
| S+T1 | 2.39 e | 1.72 ef | 1.28 de | 47.55 d |
| S+T2 | 2.84 d | 1.95 de | 1.60 bc | 42.02 e |
| S+T3 | 2.43 e | 1.77 ef | 1.12 ef | 55.18 b |
| S+T4 | 3.37 bc | 2.30 bc | 1.33 d | 60.17 a |
| S+T5 | 3.39 b | 2.33 b | 1.72 b | 50.34 cd |
| S+T6 | 3.07 cd | 2.17 bcd | 1.44 cd | 52.39 bc |
| S+T7 | 3.11 bcd | 2.09 cd | 1.62 bc | 47.29 d |
| Treatment | Heading Stage | Full-Ripening Stage | ||||||
|---|---|---|---|---|---|---|---|---|
| Stem Dry Weight (g hill−1) | Leaf Dry Weight (g hill−1) | Spike Dry Weight (g hill−1) | Total Dry Weight (g hill−1) | Stem Dry Weight (g hill−1) | Leaf Dry Weight (g hill−1) | Spike Dry Weight (g hill−1) | Total Dry Weight (g hill−1) | |
| CK | 19.61 a | 7.95 a | 5.03 a | 32.59 a | 18.58 a | 8.06 a | 25.91 a | 52.05 a |
| S | 4.33 g | 2.41 f | 0.98 e | 7.72 f | 4.31 e | 1.76 e | 5.14 g | 11.21 f |
| S+T1 | 4.75 f | 2.78 def | 0.92 e | 8.44 e | 4.59 de | 2.07 de | 5.77 fg | 12.43 ef |
| S+T2 | 5.53 e | 3.04 cde | 1.30 d | 9.87 d | 4.77 de | 2.89 c | 6.97 d | 14.62 d |
| S+T3 | 4.70 f | 2.72 ef | 1.41 d | 8.83 e | 4.91 cde | 2.31 cde | 6.26 ef | 13.47 de |
| S+T4 | 6.42 b | 3.38 bc | 1.32 d | 11.11 c | 4.72 de | 2.15 de | 9.61 c | 16.47 c |
| S+T5 | 6.40 b | 3.59 b | 1.82 c | 11.81 b | 5.13 cd | 3.96 b | 10.38 b | 19.46 b |
| S+T6 | 6.06 c | 3.26 bcd | 2.00 c | 11.32 bc | 5.63 bc | 2.60 cd | 6.52 de | 14.74 d |
| S+T7 | 5.81 d | 3.15 bcde | 2.45 b | 11.40 bc | 6.11 b | 3.70 b | 9.41 c | 19.21 b |
| Treatment | Heading Stage—Full-Ripening Stage | ||
|---|---|---|---|
| Photosynthetic Potential (m2 dm−2) | Crop Growth Rate (g m−2 d−1) | Net Assimilation Rate (g m−2 d−1) | |
| CK | 243.54 a | 15.59 a | 3.13 a |
| S | 75.50 e | 2.79 d | 1.87 cde |
| S+T1 | 88.13 d | 3.19 cd | 1.81 cde |
| S+T2 | 106.44 c | 3.81 cd | 1.77 de |
| S+T3 | 84.95 d | 3.72 cd | 2.21 bcd |
| S+T4 | 112.82 c | 4.30 c | 1.96 cde |
| S+T5 | 122.64 b | 6.13 b | 2.50 abc |
| S+T6 | 107.92 c | 2.74 d | 1.28 e |
| S+T7 | 113.53 c | 6.25 b | 2.74 ab |
| Treatment | Panicle Number (pot−1) | Spikelets per Panicle | Filled Kernel Percentage (%) | 1000-Grain Weight (g) | Yield (g pot−1) |
|---|---|---|---|---|---|
| CK | 36.0 a | 148.14 a | 81.33 a | 27.41 a | 116.72 a |
| S | 25.2 d | 40.49 g | 73.84 c | 23.71 f | 16.08 g |
| S+T1 | 24.0 d | 63.56 de | 67.84 d | 23.48 g | 17.16 g |
| S+T2 | 28.0 c | 55.33 f | 82.02 a | 24.08 e | 23.42 e |
| S+T3 | 28.0 c | 53.81 f | 78.58 b | 22.76 i | 21.52 f |
| S+T4 | 32.0 b | 72.42 c | 75.15 c | 24.95 b | 28.74 c |
| S+T5 | 36.0 a | 79.41 b | 73.47 c | 24.58 c | 31.26 b |
| S+T6 | 29.2 c | 62.23 e | 63.85 e | 24.20 d | 26.44 d |
| S+T7 | 32.0 c | 66.04 d | 78.68 b | 23.36 h | 27.33 d |
| Treatment | Panicle Length (cm) | Number of Primary Branches | Grain Number of Primary Branch | Number of Secondary Branches | Grain Number of Secondary Branch |
|---|---|---|---|---|---|
| CK | 16.82 a | 11.59 a | 73.00 a | 25.00 a | 75.15 a |
| S | 10.19 e | 5.68 e | 30.29 f | 4.40 e | 11.26 e |
| S+T1 | 11.60 c | 7.00 cd | 41.66 cd | 8.00 cd | 21.89 cd |
| S+T2 | 11.10 d | 6.95 cd | 37.52 de | 6.86 d | 17.81 d |
| S+T3 | 11.09 d | 6.81 cd | 35.14 ef | 7.57 d | 18.66 d |
| S+T4 | 12.33 b | 7.42 bc | 43.21 bc | 10.79 b | 29.59 b |
| S+T5 | 12.28 b | 8.33 b | 47.93 b | 11.40 b | 31.85 b |
| S+T6 | 10.86 d | 6.33 de | 35.25 ef | 7.69 cd | 20.86 cd |
| S+T7 | 12.06 bc | 7.50 bc | 41.13 cd | 9.00 c | 24.92 c |
| Treatment | Root | Stem | Leaf | Spike | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| K+ (mg g−1) | Na+ (mg g−1) | K+/Na+ | K+ (mg g−1) | Na+ (mg g−1) | K+/Na+ | K+ (mg g−1) | Na+ (mg g−1) | K+/Na+ | K+ (mg g−1) | Na+ (mg g−1) | K+/Na+ | |
| CK | 9.59 a | 30.65 e | 0.31 a | 53.69 a | 25.35 e | 2.12 a | 36.63 e | 13.88 g | 2.64 a | 6.16 d | 8.29 d | 0.74 a |
| S | 1.73 ef | 50.15 bcd | 0.03 ef | 30.01 cd | 90.29 c | 0.33 cd | 17.97 g | 53.19 d | 0.34 c | 9.11 c | 18.84 ab | 0.48 c |
| S+T1 | 0.70 f | 44.33 d | 0.02 f | 26.82 de | 114.35 a | 0.24 f | 40.87 d | 91.79 a | 0.45 bc | 15.45 a | 23.29 a | 0.66 ab |
| S+T2 | 4.88 bc | 48.77 cd | 0.10 c | 27.40 de | 116.65 a | 0.24 f | 49.55 b | 88.34 ab | 0.56 bc | 11.85 b | 22.93 a | 0.52 bc |
| S+T3 | 2.48 de | 48.01 cd | 0.05 de | 32.13 bc | 107.58 b | 0.30 de | 57.89 a | 83.84 bc | 0.69 b | 10.36 bc | 21.92 a | 0.47 c |
| S+T4 | 2.53 de | 60.69 a | 0.04 ef | 33.97 b | 73.69 d | 0.46 b | 21.53 f | 43.03 f | 0.50 bc | 8.37 c | 13.59 c | 0.62 abc |
| S+T5 | 5.03 b | 34.90 e | 0.14 b | 30.11 cd | 103.94 b | 0.29 e | 44.92 c | 79.58 c | 0.57 bc | 10.22 bc | 21.82 a | 0.47 c |
| S+T6 | 3.68 cd | 53.86 bc | 0.07 d | 25.88 e | 89.12 c | 0.29 e | 12.53 h | 44.23 ef | 0.28 c | 9.03 c | 15.77 bc | 0.57 bc |
| S+T7 | 5.9 b | 55.18 ab | 0.11 c | 29.57 cd | 86.96 c | 0.34 c | 23.28 f | 50.33 de | 0.46 bc | 8.74 c | 14.34 c | 0.61 abc |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, Y.; Ge, J.; Liu, Y.; Li, R.; Zhang, R.; Li, K.; Huo, Z.; Xu, K.; Wei, H.; Dai, Q. 24-Epibrassnolide Alleviates the Adverse Effect of Salinity on Rice Grain Yield through Enhanced Antioxidant Enzyme and Improved K+/Na+ Homeostasis. Agronomy 2022, 12, 2499. https://doi.org/10.3390/agronomy12102499
Chen Y, Ge J, Liu Y, Li R, Zhang R, Li K, Huo Z, Xu K, Wei H, Dai Q. 24-Epibrassnolide Alleviates the Adverse Effect of Salinity on Rice Grain Yield through Enhanced Antioxidant Enzyme and Improved K+/Na+ Homeostasis. Agronomy. 2022; 12(10):2499. https://doi.org/10.3390/agronomy12102499
Chicago/Turabian StyleChen, Yinglong, Jianfei Ge, Yang Liu, Rongkai Li, Rui Zhang, Kangrui Li, Zhongyang Huo, Ke Xu, Huanhe Wei, and Qigen Dai. 2022. "24-Epibrassnolide Alleviates the Adverse Effect of Salinity on Rice Grain Yield through Enhanced Antioxidant Enzyme and Improved K+/Na+ Homeostasis" Agronomy 12, no. 10: 2499. https://doi.org/10.3390/agronomy12102499