
The Alterations and Roles of Glycosaminoglycans in Human Diseases

Abstract
:1. Introduction
2. GAGs in Atherosclerosis
3. GAGs in Cancers
4. GAGs in Diabetes Mellitus
5. GAGs in Neurodegenerative Disease
6. GAGs in Virus Infection
7. GAGs in Other Diseases
8. Prospects and Challenges
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
Abbreviations
References
- Li, L.; Ly, M.; Linhardt, R.J. Proteoglycan Sequence. Mol. Biosyst. 2012, 8, 1613–1625. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Zhang, F.; Linhardt, R.J. Analysis of the Glycosaminoglycan Chains of Proteoglycans. J. Histochem. Cytochem. 2021, 69, 121–135. [Google Scholar] [CrossRef] [PubMed]
- Shi, D.; Sheng, A.; Chi, L. Glycosaminoglycan-Protein Interactions and Their Roles in Human Disease. Front. Mol. Biosci. 2021, 8, 639666. [Google Scholar] [CrossRef] [PubMed]
- Buijsers, B.; Yanginlar, C.; Maciej-Hulme, M.L.; de Mast, Q.; van der Vlag, J. Beneficial Non-Anticoagulant Mechanisms Underlying Heparin Treatment of Covid-19 Patients. EBioMedicine 2020, 59, 102969. [Google Scholar] [CrossRef] [PubMed]
- De Pasquale, V.; Pavone, L.M. Heparan Sulfate Proteoglycans: The Sweet Side of Development Turns Sour in Mucopolysaccharidoses. Biochim. Biophys. Acta Mol. Basis Dis. 2019, 1865, 165539. [Google Scholar] [CrossRef] [PubMed]
- Aquino, R.S.; Lee, E.S.; Park, P.W. Diverse Functions of Glycosaminoglycans in Infectious Diseases. In Progress in Molecular Biology and Translational Science; Zhang, L., Ed.; Academic Press: New York, NY, USA, 2010; Volume 93, pp. 373–394. [Google Scholar]
- Sugahara, K.; Mikami, T.; Uyama, T.; Mizuguchi, S.; Nomura, K.; Kitagawa, H. Recent Advances in the Structural Biology of Chondroitin Sulfate and Dermatan Sulfate. Curr. Opin. Struct. Biol. 2003, 13, 612–620. [Google Scholar] [CrossRef]
- Malavaki, C.; Mizumoto, S.; Karamanos, N.; Sugahara, K. Recent Advances in the Structural Study of Functional Chondroitin Sulfate and Dermatan Sulfate in Health and Disease. Connect. Tissue Res. 2008, 49, 133–139. [Google Scholar] [CrossRef]
- Tang, F.; Lord, M.S.; Stallcup, W.B.; Whitelock, J.M. Cell Surface Chondroitin Sulphate Proteoglycan 4 (Cspg4) Binds to the Basement Membrane Heparan Sulphate Proteoglycan, Perlecan, and Is Involved in Cell Adhesion. J. Biochem. 2018, 163, 399–412. [Google Scholar] [CrossRef] [Green Version]
- Avram, S.; Shaposhnikov, S.; Buiu, C.; Mernea, M. Chondroitin Sulfate Proteoglycans: Structure-Function Relationship with Implication in Neural Development and Brain Disorders. Biomed Res. Int. 2014. [Google Scholar] [CrossRef] [Green Version]
- Sugahara, K.; Mikami, T. Chondroitin/Dermatan Sulfate in the Central Nervous System. Curr. Opin. Struct. Biol. 2007, 17, 536–545. [Google Scholar] [CrossRef]
- Du Souich, P.; Garcia, A.G.; Verges, J.; Montell, E. Immunomodulatory and Anti-Inflammatory Efects of Chondroitin Sulphate. J. Cell. Mol. Med. 2009, 13, 1451–1463. [Google Scholar] [CrossRef] [PubMed]
- Sodhi, H.; Panitch, A. Glycosaminoglycans in Tissue Engineering: A Review. Biomolecules 2020, 11, 29. [Google Scholar] [CrossRef] [PubMed]
- Caterson, B.; Melrose, J. Keratan Sulfate, a Complex Glycosaminoglycan with Unique Functional Capability. Glycobiology 2018, 28, 182–206. [Google Scholar] [CrossRef] [Green Version]
- Iozzo, R.V.; Schaefer, L. Proteoglycan Form and Function: A Comprehensive Nomenclature of Proteoglycans. Matrix Biol. 2015, 42, 11–55. [Google Scholar] [CrossRef]
- Necas, J.; Bartosikova, L.; Brauner, P.; Kolar, J. Hyaluronic Acid (Hyaluronan): A Review. Vet. Med. 2008, 53, 397–411. [Google Scholar] [CrossRef] [Green Version]
- Gesslbauer, B.; Rek, A.; Falsone, F.; Rajkovic, E.; Kungl, A.J. Proteoglycanomics: Tools to Unravel the Biological Function of Glycosaminoglycans. Proteomics 2007, 7, 2870–2880. [Google Scholar] [CrossRef]
- Bulow, H.E.; Hobert, O. The Molecular Diversity of Glycosaminoglycans Shapes Animal Development. Annu. Rev. Cell Dev. Biol. 2006, 22, 375–407. [Google Scholar] [CrossRef]
- Esko, J.D.; Selleck, S.B. Order out of Chaos: Assembly of Ligand Binding Sites in Heparan Sulfate. Annu. Rev. Biochem. 2002, 71, 435–471. [Google Scholar] [CrossRef] [PubMed]
- Capila, I.; Linhardt, R.J. Heparin-Protein Interactions. Angew. Chem. Int. Ed. 2002, 41, 390–412. [Google Scholar] [CrossRef]
- Couchman, J.R. Transmembrane Signaling Proteoglycans. Annu. Rev. Cell Dev. Biol. 2010, 26, 89–114. [Google Scholar] [CrossRef]
- Nakato, H.; Kimata, K. Heparan Sulfate Fine Structure and Specificity of Proteoglycan Functions. Biochim. Biophys. Acta 2002, 1573, 312–318. [Google Scholar] [CrossRef]
- Gallagher, J.T. Multiprotein Signalling Complexes: Regional Assembly on Heparan Sulphate. Biochem. Soc. Trans. 2006, 34, 438–441. [Google Scholar] [CrossRef] [PubMed]
- Hiroko, H.; Goichiro, M.; Ken, N.; Asato, K.; Yoichi, M.; Marion, K.G.; Osami, H.; Masayuki, T.; Koji, K. Biosynthesis of Heparan Sulphate with Diverse Structures and Functions Two Alternatively Spliced Forms of Human Heparan Sulphate 6-O-Sulphotransferase-2 Having Different Expression Patterns and Properties. Biochem. J. 2003, 371, 131–142. [Google Scholar]
- Dagenais, G.R.; Leong, D.P.; Rangarajan, S.; Lanas, F.; Lopez-Jaramillo, P.; Gupta, R.; Diaz, R.; Avezum, A.; Oliveira, G.B.F.; Wielgosz, A.; et al. Variations in Common Diseases, Hospital Admissions, and Deaths in Middle-Aged Adults in 21 Countries from Five Continents (Pure): A Prospective Cohort Study. Lancet 2020, 395, 785–794. [Google Scholar] [CrossRef]
- La Sala, L.; Prattichizzo, F.; Ceriello, A. The Link between Diabetes and Atherosclerosis. Eur. J. Prev. Cardiol. 2019, 26, 15–24. [Google Scholar] [CrossRef]
- Ye, J.; Li, L.; Wang, M.; Ma, Q.; Tian, Y.; Zhang, Q.; Liu, J.; Li, B.; Zhang, B.; Liu, H.; et al. Diabetes Mellitus Promotes the Development of Atherosclerosis: The Role of Nlrp3. Front. Immunol. 2022, 13, 900254. [Google Scholar] [CrossRef]
- Pennathur, S.; Heinecke, J.W. Oxidative Stress and Endothelial Dysfunction in Vascular Disease. Curr. Diabetes Rep. 2007, 7, 257–264. [Google Scholar] [CrossRef]
- Luscher, T.F. Inflammation: The New Cardiovascular Risk Factor. Eur. Heart J. 2018, 39, 3483–3487. [Google Scholar] [CrossRef]
- Lancellotti, P.; Marechal, P.; Donis, N.; Oury, C. Inflammation, Cardiovascular Disease, and Cancer: A Common Link with Far-Reaching Implications. Eur. Heart J. 2019, 40, 3910–3912. [Google Scholar] [CrossRef] [Green Version]
- Valdivielso, J.M.; Rodriguez-Puyol, D.; Pascual, J.; Barrios, C.; Bermudez-Lopez, M.; Sanchez-Nino, M.D.; Perez-Fernandez, M.; Ortiz, A. Atherosclerosis in Chronic Kidney Disease: More, Less, or Just Different? Arterioscler. Thromb. Vasc. Biol. 2019, 39, 1938–1966. [Google Scholar] [CrossRef]
- Choi, J.; Haan, J.D.; Sharma, A. Animal Models of Diabetes-Associated Vascular Diseases: An Update on Available Models and Experimental Analysis. Br. J. Pharmacol. 2022, 179, 748–769. [Google Scholar] [CrossRef] [PubMed]
- Drinkwater, J.J.; Davis, T.M.E.; Davis, W.A. The Relationship between Carotiddisease and Retinopathy in Diabetes: A Systematic Review. Cardiovasc. Diabetol. 2020, 19, 54. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.J.; Jung, S.K. Nutraceuticals for Prevention of Atherosclerosis: Targeting Monocyte Infiltration to the Vascular Endothelium. J. Food Biochem. 2020, 44, e13200. [Google Scholar] [CrossRef] [PubMed]
- Masola, V.; Zaza, G.; Arduini, A.; Onisto, M.; Gambaro, G. Endothelial Glycocalyx as a Regulator of Fibrotic Processes. Int. J. Mol. Sci. 2021, 22, 2996. [Google Scholar] [CrossRef]
- Ghadie, N.M.; St-Pierre, J.P.; Labrosse, M.R. The Contribution of Glycosaminoglycans/Proteoglycans to Aortic Mechanics in Health and Disease: A Critical Review. IEEE Trans. Biomed. Eng. 2021, 68, 3491–3500. [Google Scholar] [CrossRef]
- Naso, M.F.; Zimmermann, D.R.; Iozzo, R.V. Characterization of the Complete Genomic Structure of the Human Versican Gene and Functional Analysis of Its Promoter. J. Biol. Chem. 1994, 269, 32999–33008. [Google Scholar] [CrossRef]
- Zimmermann, D.R.; Ruoslahti, E. Multiple Domains of the Large Fibroblast Proteoglycan, Versican. EMBO J. 1989, 8, 2975–2981. [Google Scholar] [CrossRef]
- Jackson, R.L.; Busch, S.J.; Cardin, A.D. Glycosaminoglycans: Molecular Properties, Protein Interactions, and Role in Physiological Processes. Physiol. Rev. 1991, 71, 481–539. [Google Scholar] [CrossRef]
- Hultgardh-Nilsson, A.; Boren, J.; Chakravarti, S. The Small Leucine-Rich Repeat Proteoglycans in Tissue Repair and Atherosclerosis. J. Intern. Med. 2015, 278, 447–461. [Google Scholar] [CrossRef] [Green Version]
- Boren, J.; Olin, K.; Lee, I.; Chait, A.; Wight, T.N.; Innerarity, T.L. Identification of the Principal Proteoglycan-Binding Site in Ldl. A Single-Point Mutation in Apo-B100 Severely Affects Proteoglycan Interaction without Affecting Ldl Receptor Binding. J. Clin. Investig. 1998, 101, 2658–2664. [Google Scholar] [CrossRef] [Green Version]
- Olin, K.L.; Potter-Perigo, S.; Barrett, P.H.; Wight, T.N.; Chait, A. Lipoprotein Lipase Enhances the Binding of Native and Oxidized Low Density Lipoproteins to Versican and Biglycan Synthesized by Cultured Arterial Smooth Muscle Cells. J. Biol. Chem. 1999, 274, 34629–34636. [Google Scholar] [CrossRef] [PubMed]
- Pentikäinen, M.O.; Oörni, K.; Lassila, R.; Kovanen, P.T. The Proteoglycan Decorin Links Low Density Lipoproteins with Collagen Type I. J. Biol. Chem. 1997, 272, 7633–7638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wight, T.N.; Merrilees, M.J. Proteoglycans in Atherosclerosis and Restenosis: Key Roles for Versican. Circ. Res. 2004, 94, 1158–1167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vijayagopal, P.; Figueroa, J.E.; Fontenot, J.D.; Glancy, D.L. Isolation and Characterization of a Proteoglycan Variant from Human Aorta Exhibiting a Marked Affinity for Low Density Lipoprotein and Demonstration of Its Enhanced Expression in Atherosclerotic Plaques. Atherosclerosis 1996, 127, 195–203. [Google Scholar] [CrossRef]
- Hinek, A.; Mecham, R.P.; Keeley, F.; Rabinovitch, M. Impaired Elastin Fiber Assembly Related to Reduced 67-KD Elastin-Binding Protein in Fetal Lamb Ductus Arteriosus and in Cultured Aortic Smooth Muscle Cells Treated with Chondroitin Sulfate. J. Clin. Investig. 1991, 88, 2083–2094. [Google Scholar] [CrossRef]
- Wight, T.N. A Role for Proteoglycans in Vascular Disease. Matrix Biol. 2018, 71–72, 396–420. [Google Scholar] [CrossRef]
- Sambandam, T.; Baker, J.R.; Christner, J.E.; Eborg, S.L. Specificity of the Low Density Lipoproteins-Glycosaminoglycan Interaction. Arterioscler. Thromb. 1991, 11, 561–568. [Google Scholar] [CrossRef] [Green Version]
- Cardoso, L.E.; Mourão, P.A. Glycosaminoglycan Fractions from Human Arteries Presenting Diverse Susceptibilities to Atherosclerosis Have Different Binding Affinities to Plasma Ldl. Arterioscler. Thromb. 1994, 14, 115–124. [Google Scholar] [CrossRef] [Green Version]
- Theocharis, A.D.; Theocharis, D.A.; Luca, G.D.; Hjerpe, A.; Karamanos, N.K. Compositional and Structural Aterations of Chondroitin and Dermatan Sulfates During the Progression of Atherosclerosis and Aneurysmal Dilatation of the Human Abdominal Aorta. Biochimie 2002, 84, 667–674. [Google Scholar] [CrossRef]
- Little, P.J.; Tannock, L.; Olin, K.L.; Chait, A.; Wight, T.N. Proteoglycans Synthesized by Arterial Smooth Muscle Cells in the Presence of Transforming Growth Factor-Beta1 Exhibit Increased Binding to Ldls. Arterioscler. Thromb. Vasc. Biol. 2002, 22, 55–60. [Google Scholar] [CrossRef] [Green Version]
- Pillarisetti, S. Lipoprotein Modulation of Subendothelial Heparan Sulfate Proteoglycans (Perlecan) and Atherogenicity. Trends Cardiovasc. Med. 2000, 10, 60–65. [Google Scholar] [CrossRef]
- Tran, P.K.; Agardh, H.E.; Tran-Lundmark, K.; Ekstrand, J.; Roy, J.; Henderson, B.; Gabrielsen, A.; Hansson, G.K.; Swedenborg, J.; Paulsson-Berne, G.; et al. Reduced Perlecan Expression and Accumulation in Human Carotid Atherosclerotic Lesions. Atherosclerosis 2007, 190, 264–270. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, M.; Imaizumi, K.; Shigemi, U.; Nakashima, Y.; Kikuchi, Y. Relationship of Sulfated Glycosaminoglycans and Cholesterol Content in Normal and Atherosclerotic Human Aorta. Stroke 1976, 7, 594–598. [Google Scholar] [CrossRef] [Green Version]
- Völker, W.; Schmidt, A.; Oortmann, W.; Broszey, T.; Faber, V.; Buddecke, E. Mapping of Proteoglycans in Atherosclerotic Lesions. Eur. Heart J. 1990, 11, 29–40. [Google Scholar] [CrossRef] [PubMed]
- Pillarisetti, S.; Paka, L.; Obunike, J.C.; Berglund, L.; Goldberg, I.J. Subendothelial Retention of Lipoprotein (a). Evidence That Teduced Heparan Sulfate Promotes Lipoprotein Binding to Subendothelial Matrix. J. Clin. Investig. 1997, 100, 867–874. [Google Scholar] [CrossRef] [PubMed]
- Duan, W.; Paka, L.; Pillarisetti, S. Distinct Effects of Glucose and Glucosamine on Vascular Endothelial and Smooth Muscle Cells: Evidence for a Protective Role for Glucosamine in Atherosclerosis. Cardiovasc. Diabetol. 2005, 4, 16. [Google Scholar] [CrossRef] [Green Version]
- Asplund, A.; Ostergren-Lunden, G.; Camejo, G.; Stillemark-Billton, P.; Bondjers, G. Hypoxia Increases Macrophage Motility, Possibly by Decreasing the Heparan Sulfate Proteoglycan Biosynthesis. J. Leukoc. Biol. 2009, 86, 381–388. [Google Scholar] [CrossRef] [Green Version]
- Fischer, J.W. Role of Hyaluronan in Atherosclerosis: Current Knowledge and Open Questions. Matrix Biol. 2019, 78–79, 324–336. [Google Scholar] [CrossRef]
- Karangelis, D.E.; Kanakis, I.; Asimakopoulou, A.P.; Karousou, E.; Passi, A.; Theocharis, A.D.; Triposkiadis, F.; Tsilimingas, N.B.; Karamanos, N. Glycosaminoglycans as Key Molecules in Atherosclerosis: The Role of Versican and Hyaluronan. Curr. Med. Chem. 2010, 17, 4018–4026. [Google Scholar] [CrossRef]
- Vigetti, D.; Viola, M.; Karousou, E.; Rizzi, M.; Moretto, P.; Genasetti, A.; Clerici, M.; Hascall, V.C.; De Luca, G.; Passi, A. Hyaluronan-Cd44-Erk1/2 Regulate Human Aortic Smooth Muscle Cell Motility During Aging. J. Biol. Chem. 2008, 283, 4448–4458. [Google Scholar] [CrossRef] [Green Version]
- Seike, M.; Ikeda, M.; Matsumoto, M.; Hamada, R.; Takeya, M.; Kodama, H. Hyaluronan Forms Complexes with Low Density Lipoprotein While Also Inducing Foam Cell Infiltration in the Dermis. J. Dermatol. Sci. 2006, 41, 197–204. [Google Scholar] [CrossRef] [PubMed]
- Noble, P.W. Hyaluronan and Its Catabolic Products in Tissue Injury and Repair. Matrix Biol. 2002, 21, 25–29. [Google Scholar] [CrossRef]
- Grandoch, M.; Bollyky, P.L.; Fischer, J.W. Hyaluronan: A Master Switch between Vascular Homeostasis and Inflammation. Circ. Res. 2018, 122, 1341–1343. [Google Scholar] [CrossRef] [PubMed]
- Tabata, T.; Mine, S.; Okada, Y.; Tanaka, Y. Low Molecular Weight Hyaluronan Increases the Uptaking of Oxidized Ldl into Monocytes. Endocr. J. 2007, 54, 685–693. [Google Scholar] [CrossRef] [Green Version]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: Globocan Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Bergers, G.; Fendt, S.M. The Metabolism of Cancer Cells During Metastasis. Nat. Rev. Cancer 2021, 21, 162–180. [Google Scholar] [CrossRef]
- Lemaire, J.; Larrue, R.; Perrais, M.; Cauffiez, C.; Pottier, N. Fundamental Aspects of Oncogenesis. Bull. Cancer 2020, 107, 1148–1160. [Google Scholar]
- Poltavets, V.; Kochetkova, M.; Pitson, S.M.; Samuel, M.S. The Role of the Extracellular Matrix and Its Molecular and Cellular Regulators in Cancer Cell Plasticity. Front. Oncol. 2018, 8, 431. [Google Scholar] [CrossRef] [Green Version]
- Xiao, Y.; Yu, D. Tumor Microenvironment as a Therapeutic Target in Cancer. Pharmacol. Ther. 2021, 221, 107753. [Google Scholar] [CrossRef]
- Arneth, B. Tumor Microenvironment. Medicina 2020, 56, 15. [Google Scholar] [CrossRef] [Green Version]
- Walker, C.; Mojares, E.; Del Rio Hernandez, A. Role of Extracellular Matrix in Development and Cancer Progression. Int. J. Mol. Sci. 2018, 19, 3028. [Google Scholar] [CrossRef] [Green Version]
- Wei, J.; Hu, M.; Huang, K.; Lin, S.; Du, H. Roles of Proteoglycans and Glycosaminoglycans in Cancer Development and Progression. Int. J. Mol. Sci. 2020, 21, 5983. [Google Scholar] [CrossRef]
- Afratis, N.; Gialeli, C.; Nikitovic, D.; Tsegenidis, T.; Karousou, E.; Theocharis, A.D.; Pavao, M.S.; Tzanakakis, G.N.; Karamanos, N.K. Glycosaminoglycans: Key Players in Cancer Cell Biology and Treatment. FEBS J. 2012, 279, 1177–1197. [Google Scholar] [CrossRef]
- Berdiaki, A.; Neagu, M.; Giatagana, E.M.; Kuskov, A.; Tsatsakis, A.M.; Tzanakakis, G.N.; Nikitovic, D. Glycosaminoglycans: Carriers and Targets for Tailored Anti-Cancer Therapy. Biomolecules 2021, 11, 395. [Google Scholar] [CrossRef]
- Nagarajan, A.; Malvi, P.; Wajapeyee, N. Heparan Sulfate and Heparan Sulfate Proteoglycans in Cancer Initiation and Progression. Front. Endocrinol. 2018, 9, 483. [Google Scholar] [CrossRef] [Green Version]
- Soares da Costa, D.; Reis, R.L.; Pashkuleva, I. Sulfation of Glycosaminoglycans and Its Implications in Human Health and Disorders. Annu. Rev. Biomed. Eng. 2017, 19, 1–26. [Google Scholar] [CrossRef] [Green Version]
- Jansson, M.; Billing, O.; Herdenberg, C.; Lundin, C.; Tolockiene, E.; Nazemroaya, A.; Sund, M. Expression and Circulating Levels of Perlecan in Breast Cancer—Implications for Oestrogen Dependent Stromal Remodeling. J. Mammary Gland Biol. Neoplasia 2020, 25, 69–77. [Google Scholar] [CrossRef] [Green Version]
- Datta, S.; Pierce, M.; Datta, M.W. Perlecan Signaling: Helping Hedgehog Stimulate Prostate Cancer Growth. Int. J. Biochem. Cell Biol. 2006, 38, 1855–1861. [Google Scholar] [CrossRef]
- Garcia-Suarez, O.; Garcia, B.; Fernandez-Vega, I.; Astudillo, A.; Quiros, L.M. Neuroendocrine Tumors Show Altered Expression of Chondroitin Sulfate, Glypican 1, Glypican 5, and Syndecan 2 Depending on Their Differentiation Grade. Front. Oncol. 2014, 4, 15. [Google Scholar] [CrossRef] [Green Version]
- Fernández-Vega, I.; García, O.; Crespo, A.; Castañón, S.; Menéndez, P.; Astudillo, A.; Quirós, L.M. Specific Genes Involved in Synthesis and Editing of Heparan Sulfate Proteoglycans Show Altered Expression Patterns in Breast Cancer. BMC Cancer 2013, 17, 13–24. [Google Scholar] [CrossRef] [Green Version]
- Cooney, C.A.; Jousheghany, F.; Yao-Borengasser, A.Y.; Phanavanh, B.; Gomes, T.; Kieber-Emmons, A.M.; Siegel, E.R.; Suva, L.J.; Ferrone, S.; Kieber-Emmons, T.; et al. Chondroitin Sulfates Play a Major Role in Breast Cancer Metastasis: A Role for Cspg4 and Chst11 Gene Expression in Forming Surface P-Selectin Ligands in Aggressive Breast Cancer Cells. Breast Cancer Res. 2011, 13, R58. [Google Scholar] [CrossRef] [Green Version]
- Vynios, D.H.; Theocharis, D.A.; Papageorgakopoulou, N.; Papadas, T.A.; Mastronikolis, N.S.; Goumas, P.D.; Stylianou, M.; Skandalis, S.S. Biochemical Changes of Extracellular Proteoglycans in Squamous Cell Laryngeal Carcinoma. Connect. Tissue Res. 2008, 49, 239–243. [Google Scholar] [CrossRef]
- Backen, A.C.; Cole, C.L.; Lau, S.C.; Clamp, A.R.; McVey, R.; Gallagher, J.T.; Jayson, G.C. Heparan Sulphate Synthetic and Editing Enzymes in Ovarian Cancer. Br. J. Cancer 2007, 96, 1544–1548. [Google Scholar] [CrossRef] [Green Version]
- Vallen, M.J.; Massuger, L.F.; ten Dam, G.B.; Bulten, J.; van Kuppevelt, T.H. Highly Sulfated Chondroitin Sulfates, a Novel Class of Prognostic Biomarkers in Ovarian Cancer Tissue. Gynecol. Oncol. 2012, 127, 202–209. [Google Scholar] [CrossRef]
- Kalathas, D.; Theocharis, D.A.; Bounias, D.; Kyriakopoulou, D.; Papageorgakopoulou, N.; Stavropoulos, M.S.; Vynios, D.H. Alterations of Glycosaminoglycan Disaccharide Content and Composition in Colorectal Cancer: Structural and Expressional Studies. Oncol. Rep. 2009, 22, 369–375. [Google Scholar]
- Weyers, A.; Yang, B.; Yoon, D.S.; Park, J.H.; Zhang, F.; Lee, K.B.; Linhardt, R.J. A Structural Analysis of Glycosaminoglycans from Lethal and Nonlethal Breast Cancer Tissues: Toward a Novel Class of Theragnostics for Personalized Medicine in Oncology? OMICS 2012, 16, 79–89. [Google Scholar] [CrossRef] [Green Version]
- Faria-Ramos, I.; Poças, J.; Marques, C.; Santos-Antunes, J.; Macedo, G.; Reis, C.A.; Magalhães, A. Heparan Sulfate Glycosaminoglycans: (Un)Expected Allies in Cancer Clinical Management. Biomolecules 2021, 11, 136. [Google Scholar] [CrossRef]
- El Masri, R.; Seffouh, A.; Lortat-Jacob, H.; Vives, R.R. The “in and out” of Glucosamine 6-O-Sulfation: The 6th Sense of Heparan Sulfate. Glycoconj. J. 2017, 34, 285–298. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, D.; Zhang, T.; Xia, L. Fgf/Fgfr Signaling in Hepatocellular Carcinoma: From Carcinogenesis to Recent Therapeutic Intervention. Cancers 2021, 13, 1360. [Google Scholar] [CrossRef]
- Jomrich, G.; Hudec, X.; Harpain, F.; Winkler, D.; Timelthaler, G.; Mohr, T.; Marian, B.; Schoppmann, S.F. Expression of Fgf8, Fgf18, and Fgfr4 in Gastroesophageal Adenocarcinomas. Cells 2019, 8, 1092. [Google Scholar] [CrossRef] [Green Version]
- Stehbens, S.J.; Ju, R.J.; Adams, M.N.; Perry, S.R.; Haass, N.K.; Bryant, D.M.; Pollock, P.M. Fgfr2-Activating Mutations Disrupt Cell Polarity to Potentiate Migration and Invasion in Endometrial Cancer Cell Models. J. Cell Sci. 2018, 131, jcs213678. [Google Scholar] [CrossRef] [Green Version]
- Stauber, D.J.; DiGabriele, A.D.; Hendrickson, W.A. Structural Interactions of Fibroblast Growth Factor Receptor with Its Ligands. Proc. Natl. Acad. Sci. USA 2000, 97, 49–54. [Google Scholar] [CrossRef] [Green Version]
- Elgundi, Z.; Papanicolaou, M.; Major, G.; Cox, T.R.; Melrose, J.; Whitelock, J.M.; Farrugia, B.L. Cancer Metastasis: The Role of the Extracellular Matrix and the Heparan Sulfate Proteoglycan Perlecan. Front. Oncol. 2019, 9, 1482. [Google Scholar] [CrossRef]
- Lundin, L.; Larsson, H.; Kreuger, J.; Kanda, S.; Lindahl, U.; Salmivirta, M.; Claesson-Welsh, L. Selectively Desulfated Heparin Inhibits Fibroblast Growth Factor-Induced Mitogenicity and Angiogenesis. J. Biol. Chem. 2000, 275, 24653–24660. [Google Scholar] [CrossRef] [Green Version]
- Pye, D.A.; Vivès, R.R.; Hyde, P.; Gallagher, J.T. Regulation of Fgf-1 Mitogenic Activity by Heparan Sulfate Oligosaccharides Is Dependent on Specific Structural Features: Differential Requirements for the Modulation of Fgf-1 and Fgf-2. Glycobiology 2000, 10, 1183–1192. [Google Scholar] [CrossRef]
- Kreuger, J.; Salmivirta, M.; Sturiale, L.; Gimenez-Gallego, G.; Lindahl, U. Sequence Analysis of Heparan Sulfate Epitopes with Graded Affinities for Fibroblast Growth Factors 1 and 2. J. Biol. Chem. 2001, 276, 30744–30752. [Google Scholar] [CrossRef] [Green Version]
- Schultz, V.; Suflita, M.; Liu, X.; Zhang, X.; Yu, Y.; Li, L.; Green, D.E.; Xu, Y.; Zhang, F.; DeAngelis, P.L.; et al. Heparan Sulfate Domains Required for Fibroblast Growth Factor 1 and 2 Signaling through Fibroblast Growth Factor Receptor 1c. J. Biol. Chem. 2017, 292, 2495–2509. [Google Scholar] [CrossRef] [Green Version]
- Brunetti, J.; Depau, L.; Falciani, C.; Gentile, M.; Mandarini, E.; Riolo, G.; Lupetti, P.; Pini, A.; Bracci, L. Insights into the Role of Sulfated Glycans in Cancer Cell Adhesion and Migration through Use of Branched Peptide Probe. Sci. Rep. 2016, 6, 27174. [Google Scholar] [CrossRef] [Green Version]
- Falciani, C.; Brunetti, J.; Lelli, B.; Ravenni, N.; Lozzi, L.; Depau, L.; Scali, S.; Bernini, A.; Pini, A.; Bracci, L. Cancer Selectivity of Tetrabranched Neurotensin Peptides Is Generated by Simultaneous Binding to Sulfated Glycosaminoglycans and Protein Receptors. J. Med. Chem. 2013, 56, 5009–5018. [Google Scholar] [CrossRef]
- Falciani, C.; Fabbrini, M.; Pini, A.; Lozzi, L.; Lelli, B.; Pileri, S.; Brunetti, J.; Bindi, S.; Scali, S.; Bracci, L. Synthesis and Biological Activity of Stable Branched Neurotensin Peptides for Tumortargeting. Mol. Cancer Ther. 2007, 6, 2441–2448. [Google Scholar] [CrossRef] [Green Version]
- Falciani, C.; Brunetti, J.; Pagliuca, C.; Menichetti, S.; Vitellozzi, L.; Lelli, B.; Pini, A.; Bracci, L. Design and in Vitro Evaluation of Branched Peptide Conjugates: Turning Nonspecific Cytotoxic Drugs into Tumor-Selective Agents. ChemMedChem 2010, 5, 567–574. [Google Scholar] [CrossRef] [PubMed]
- Brunetti, J.; Falciani, C.; Lelli, B.; Minervini, A.; Ravenni, N.; Depau, L.; Siena, G.; Tenori, E.; Menichetti, S.; Pini, A.; et al. Neurotensin Branched Peptide as a Tumor-Targeting Agent for Human Bladder Cancer. Biomed Res. Int. 2015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brunetti, J.; Pillozzi, S.; Falciani, C.; Depau, L.; Tenori, E.; Scali, S.; Lozzi, L.; Pini, A.; Arcangeli, A.; Menichetti, S.; et al. Tumor-Selective Peptide-Carrier Delivery of Paclitaxel Increases in Vivo Activity of the Drug. Sci. Rep. 2015, 5, 17736. [Google Scholar] [CrossRef] [Green Version]
- Brunetti, J.; Riolo, G.; Gentile, M.; Bernini, A.; Paccagnini, E.; Falciani, C.; Lozzi, L.; Scali, S.; Depau, L.; Pini, A.; et al. Near-Infrared Quantum Dots Labelled with a Tumor Selective Tetrabranched Peptide for in Vivo Imaging. J. Nanobiotechnol. 2018, 16, 21. [Google Scholar] [CrossRef] [PubMed]
- Bracci, L.; Mandarini, E.; Brunetti, J.; Depau, L.; Pini, A.; Terzuoli, L.; Scali, S.; Falciani, C. The Gag-Specific Branched Peptide Nt4 Reduces Angiogenesis and Invasiveness of Tumor Cells. PLoS ONE 2018, 13, e0194744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brunetti, J.; Riolo, G.; Depau, L.; Mandarini, E.; Bernini, A.; Karousou, E.; Passi, A.; Pini, A.; Bracci, L.; Falciani, C. Unraveling Heparan Sulfate Proteoglycan Binding Motif for Cancer Cell Selectivity. Front. Oncol. 2019, 9, 843. [Google Scholar] [CrossRef]
- Liu, D.F.; Shriver, Z.; Venkataraman, G.; Shabrawi, Y.E.; Sasisekharan, R. Tumor Cell Surface Heparan Sulfate as Cryptic Promoters or Inhibitors of Tumor Growth and Metastasis. PNAS 2002, 99, 568–573. [Google Scholar] [CrossRef] [Green Version]
- Mikami, T.; Kitagawa, H. Biosynthesis and Function of Chondroitin Sulfate. Biochim. Biophys. Acta 2013, 1830, 4719–4733. [Google Scholar] [CrossRef]
- Batista, L.T.; Matos, L.L.; Machado, L.R.; Suarez, E.R.; Theodoro, T.R.; Martins, J.R.; Nader, H.B.; Pompeo, A.C.; Pinhal, M.A. Heparanase Expression and Glycosaminoglycans Profile in Renal Cell Carcinoma. Int. J. Urol. 2012, 19, 1036–1040. [Google Scholar] [CrossRef]
- Brauchle, E.; Kasper, J.; Daum, R.; Schierbaum, N.; Falch, C.; Kirschniak, A.; Schaffer, T.E.; Schenke-Layland, K. Biomechanical and Biomolecular Characterization of Extracellular Matrix Structures in Human Colon Carcinomas. Matrix Biol. 2018, 68–69, 180–193. [Google Scholar] [CrossRef]
- Rangel, M.P.; de Sa, V.K.; Prieto, T.; Martins, J.R.M.; Olivieri, E.R.; Carraro, D.; Takagaki, T.; Capelozzi, V.L. Biomolecular Analysis of Matrix Proteoglycans as Biomarkers in Non Small Cell Lung Cancer. Glycoconj. J. 2018, 35, 233–242. [Google Scholar] [CrossRef]
- Marolla, A.P.; Waisberg, J.; Saba, G.T.; Waisberg, D.R.; Margeotto, F.B.; Pinhal, M.A. Glycomics Expression Analysis of Sulfated Glycosaminoglycans of Human Colorectal Cancer Tissues and Non-Neoplastic Mucosa by Electrospray Ionization Mass Spectrometry. Einstein 2015, 13, 510–517. [Google Scholar] [CrossRef] [Green Version]
- Svensson, K.J.; Christianson, H.C.; Kucharzewska, P.; Fagerstrom, V.; Lundstedt, L.; Borgquist, S.; Jirstrom, K.; Belting, M. Chondroitin Sulfate Expression Predicts Poor Outcome in Breast Cancer. Int. J. Oncol. 2011, 39, 1421–1428. [Google Scholar] [PubMed]
- Tsidulko, A.Y.; Kazanskaya, G.M.; Volkov, A.M.; Suhovskih, A.V.; Kiselev, R.S.; Kobozev, V.V.; Gaytan, A.S.; Krivoshapkin, A.L.; Aidagulova, S.V.; Grigorieva, E.V. Chondroitin Sulfate Content and Decorin Expression in Glioblastoma Are Associated with Proliferative Activity of Glioma Cells and Disease Prognosis. Cell Tissue Res. 2020, 379, 147–155. [Google Scholar] [CrossRef] [PubMed]
- Ricciardelli, C.; Brooks, J.H.; Suwiwat, S.; Sakko, A.J.; Mayne, K.; Raymond, W.A.; Seshadri, R.; LeBaron, R.G.; Horsfall, D.J. Regulation of Stromal Versican Expression by Breast Cancer Cells and Importance to Relapse-Free Survival in Patients with Node-Negative Primary Breast Cancer. Clin. Cancer Res. 2002, 8, 1054–1060. [Google Scholar] [PubMed]
- Silver, D.J.; Silver, J. Contributions of Chondroitin Sulfate Proteoglycans to Neurodevelopment, Injury, and Cancer. Curr. Opin. Neurobiol. 2014, 27, 171–178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Kelly, G.; Zerillo, C.; Jaworski, D.M.; Hockfield, S. Expression of a Cleaved Brain-Specific Extracellular Matrix Protein Mediates Glioma Cell Invasion in Vivo. J. Neurosci. 1998, 18, 2370–2376. [Google Scholar] [CrossRef] [Green Version]
- Silver, D.J.; Siebzehnrubl, F.A.; Schildts, M.J.; Yachnis, A.T.; Smith, G.M.; Smith, A.A.; Scheffler, B.; Reynolds, B.A.; Silver, J.; Steindler, D.A. Chondroitin Sulfate Proteoglycans Potently Inhibit Invasion and Serve as a Central Organizer of the Brain Tumor Microenvironment. J. Neurosci. 2013, 33, 15603–15617. [Google Scholar] [CrossRef] [Green Version]
- Viapiano, M.S.; Bi, W.L.; Piepmeier, J.; Hockfield, S.; Matthews, R.T. Novel Tumor-Specific Isoforms of Behab/Brevican Identified in Human Malignant Gliomas. Cancer Res. 2005, 65, 6726–6733. [Google Scholar] [CrossRef] [Green Version]
- Muller, S.; Kunkel, P.; Lamszus, K.; Ulbricht, U.; Lorente, G.A.; Nelson, A.M.; von Schack, D.; Chin, D.J.; Lohr, S.C.; Westphal, M.; et al. A Role for Receptor Tyrosine Phosphatase Zeta in Glioma Cell Migration. Oncogene 2003, 22, 6661–6668. [Google Scholar] [CrossRef] [Green Version]
- Arslan, F.; Bosserhoff, A.K.; Nickl-Jockschat, T.; Doerfelt, A.; Bogdahn, U.; Hau, P. The Role of Versican Isoforms V0/V1 in Glioma Migration Mediated by Transforming Growth Factor-Β2. Br. J. Cancer 2007, 96, 1560–1568. [Google Scholar] [CrossRef]
- Viapiano, M.S.; Hockfield, S.; Matthews, R.T. Behab/Brevican Requires Adamts-Mediated Proteolytic Cleavage to Promote Glioma Invasion. J. Neurooncol. 2008, 88, 261–272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kovalszky, I.; Pogany, G.; Molnar, G.; Jeney, A.; Lapis, K.; Karacsonyi, S.; Szecseny, A.; Iozzo, R.V. Altered Glycosaminoglycan Composition in Reactive and Neoplastic Human Liver. Biochem. Biophys. Res. Commun. 1990, 167, 883–890. [Google Scholar] [CrossRef]
- Lan, Y.; Li, X.; Liu, Y.; He, Y.; Hao, C.; Wang, H.; Jin, L.; Zhang, G.; Zhang, S.; Zhou, A.; et al. Pingyangmycin Inhibits Glycosaminoglycan Sulphation in Both Cancer Cells and Tumour Tissues. J. Cell. Mol. Med. 2020, 24, 3419–3430. [Google Scholar] [CrossRef] [Green Version]
- Sobue, M.; Takeuchi, J.; Miura, K.; Kawase, K.; Mizuno, F.; Sato, E. Glycosaminoglycan Content and Synthesis in Gastric Carcinoma. Br. J. Cancer 1980, 42, 79–84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Theocharis, A.D.; Tsara, M.E.; Papageorgacopoulou, N.; Karavias, D.D.; Theocharis, D.A. Pancreatic Carcinoma Is Characterized by Elevated Content of Hyaluronan and Chondroitin Sulfate with Altered Disaccharide Composition. Biochim. Biophys. Acta 2000, 1502, 201–206. [Google Scholar] [CrossRef] [Green Version]
- Pudelko, A.; Wisowski, G.; Olczyk, K.; Kozma, E.M. The Dual Role of the Glycosaminoglycan Chondroitin-6-Sulfate in the Development, Progression and Metastasis of Cancer. FEBS J. 2019, 286, 1815–1837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morla, S. Glycosaminoglycans and Glycosaminoglycan Mimetics in Cancer and Inflammation. Int. J. Mol. Sci. 2019, 20, 1963. [Google Scholar] [CrossRef] [Green Version]
- Skandalis, S.S.; Kletsas, D.; Kyriakopoulou, D.; Stavropoulos, M.; Theocharis, D.A. The Greatly Increased Amounts of Accumulated Versican and Decorin with Specific Post-Translational Modifications May Be Closely Associated with the Malignant Phenotype of Pancreatic Cancer. Biochim. Biophys. Acta 2006, 1760, 1217–1225. [Google Scholar] [CrossRef]
- Al-Nakouzi, N.; Wang, C.K.; Oo, H.Z.; Nelepcu, I.; Lallous, N.; Spliid, C.B.; Khazamipour, N.; Lo, J.; Truong, S.; Collins, C.; et al. Reformation of the Chondroitin Sulfate Glycocalyx Enables Progression of Ar-Independent Prostate Cancer. Nat. Commun. 2022, 13, 4760. [Google Scholar] [CrossRef]
- Theocharis, A.D.; Vynios, D.H.; Papageorgakopoulou, N.; Skandalis, S.S.; Theocharis, D.A. Altered Content Composition and Structure of Glycosaminoglycans and Proteoglycans in Gastric Carcinoma. Int. J. Biochem. Cell Biol. 2003, 35, 376–390. [Google Scholar] [CrossRef]
- Ten Dam, G.B.; van de Westerlo, E.M.; Purushothaman, A.; Stan, R.V.; Bulten, J.; Sweep, F.C.; Massuger, L.F.; Sugahara, K.; van Kuppevelt, T.H. Antibody Gd3g7 Selected against Embryonic Glycosaminoglycans Defines Chondroitin Sulfate-E Domains Highly up-Regulated in Ovarian Cancer and Involved in Vascular Endothelial Growth Factor Binding. Am. J. Pathol. 2007, 171, 1324–1333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, F.; Ten Dam, G.B.; Murugan, S.; Yamada, S.; Hashiguchi, T.; Mizumoto, S.; Oguri, K.; Okayama, M.; van Kuppevelt, T.H.; Sugahara, K. Involvement of Highly Sulfated Chondroitin Sulfate in the Metastasis of the Lewis Lung Carcinoma Cells. J. Biol. Chem. 2008, 283, 34294–34304. [Google Scholar] [CrossRef] [PubMed]
- Seidler, D.G.; Breuer, E.; Grande-Allen, K.J.; Hascall, V.C.; Kresse, H. Core Protein Dependence of Epimerization of Glucuronosyl Residues in Galactosaminoglycans. J. Biol. Chem. 2002, 277, 42409–42416. [Google Scholar] [CrossRef] [Green Version]
- Momose, T.; Yoshimura, Y.; Harumiya, S.; Isobe, K.; Kito, M.; Fukushima, M.; Kato, H.; Nakayama, J. Chondroitin Sulfate Synthase 1 Expression Is Associated with Malignant Potential of Soft Tissue Sarcomas with Myxoid Substance. Hum. Pathol. 2016, 50, 15–23. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.H.; Lan, C.T.; Chou, J.F.; Tseng, T.J.; Liao, W.C. Chsy1 Promotes Aggressive Phenotypes of Hepatocellular Carcinoma Cells Via Activation of the Hedgehog Signaling Pathway. Cancer Lett. 2017, 403, 280–288. [Google Scholar] [CrossRef]
- Honda, T.; Kaneiwa, T.; Mizumoto, S.; Sugahara, K.; Yamada, S. Hyaluronidases Have Strong Hydrolytic Activity toward Chondroitin 4-Sulfate Comparable to That for Hyaluronan. Biomolecules 2012, 2, 549–563. [Google Scholar] [CrossRef] [Green Version]
- Nikitovic, D.; Assouti, M.; Sifaki, M.; Katonis, P.; Krasagakis, K.; Karamanos, N.K.; Tzanakakis, G.N. Chondroitin Sulfate and Heparan Sulfate-Containing Proteoglycans Are Both Partners and Targets of Basic Fibroblast Growth Factor-Mediated Proliferation in Human Metastatic Melanoma Cell Lines. Int. J. Biochem. Cell Biol. 2008, 40, 72–83. [Google Scholar] [CrossRef]
- Nikitovic, D.; Zafiropoulos, A.; Tzanakakis, G.N.; Karamanos, N.K.; Tsatsakis, A.M. Effects of Glycosaminoglycans on Cell Proliferation of Normal Osteoblasts and Human Osteosarcoma Cells Depend on Their Type and Fine Chemical Compositions. Anticancer Res. 2005, 25, 2851–2856. [Google Scholar]
- Merle, B.; Durussel, L.; Delmas, P.D.; Clézardin, P. Decorin Inhibits Cell Migration through a Process Requiring Its Glycosaminoglycan Side Chain. J. Cell Biochem. 1999, 75, 538–546. [Google Scholar] [CrossRef]
- Thelin, M.A.; Svensson, K.J.; Shi, X.; Bagher, M.; Axelsson, J.; Isinger-Ekstrand, A.; van Kuppevelt, T.H.; Johansson, J.; Nilbert, M.; Zaia, J.; et al. Dermatan Sulfate Is Involved in the Tumorigenic Properties of Esophagus Squamous Cell Carcinoma. Cancer Res. 2012, 72, 1943–1952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Auvinen, P.; Tammi, R.; Parkkinen, J.; Tammi, M.; Ågren, U.; Johansson, R.; Hirvikoski, P.; Eskelinen, M.; Kosma, V.M. Hyaluronan in Peritumoral Stroma and Malignant Cells Associates with Breast Cancer Spreading and Predicts Survival. Am. J. Pathol. 2000, 156, 529–536. [Google Scholar] [CrossRef]
- Pirinen, R.; Tammi, R.; Tammi, M.; Hirvikoski, P.; Parkkinen, J.J.; Johansson, R.; Böhm, J.; Hollmén, S.; Kosma, V.M. Prognostic Value of Hyaluronan Expression in Non-Small-Cell Lung Cancer: Increased Stromal Expression Indicates Unfavorable Outcome in Patients with Adenocarcinoma. Int. J. Cancer 2001, 95, 12–17. [Google Scholar] [CrossRef]
- Anttila, M.; Tammi, R.; Tammi, M.; Syrjänen, K.J.; Saarikoski, S.V.; Kosma, V.M. High Levels of Stromal Hyaluronan Predict Poor Disease Outcome in Epithelial Ovarian Cancer. Cancer Res. 2000, 60, 150–155. [Google Scholar]
- Spinelli, F.M.; Vitale, D.L.; Sevic, I.; Alaniz, L. Hyaluronan in the Tumor Microenvironment. Adv. Exp. Med. Biol. 2020, 1245, 67–83. [Google Scholar]
- Itano, N.; Kimata, K. Mammalian Hyaluronan Synthases. IUBMB Life 2002, 54, 195–199. [Google Scholar] [CrossRef]
- Tammi, R.H.; Passi, A.G.; Rilla, K.; Karousou, E.; Vigetti, D.; Makkonen, K.; Tammi, M.I. Transcriptional and Post-Translational Regulation of Hyaluronan Synthesis. FEBS J. 2011, 278, 1419–1428. [Google Scholar] [CrossRef]
- Bourguignon, L.Y.; Wong, G.; Earle, C.A.; Xia, W. Interaction of Low Molecular Weight Hyaluronan with Cd44 and Toll-Like Receptors Promotes the Actin Filament-Associated Protein 110-Actin Binding and Myd88-Nfκb Signaling Leading to Proinflammatory Cytokine/Chemokine Production and Breast Tumor Invasion. Cytoskeleton 2011, 68, 671–693. [Google Scholar] [CrossRef] [Green Version]
- Schwertfeger, K.L.; Cowman, M.K.; Telmer, P.G.; Turley, E.A.; McCarthy, J.B. Hyaluronan, Inflammation, and Breast Cancer Progression. Front. Immunol. 2015, 6, 236. [Google Scholar] [CrossRef]
- Misra, S.; Hascall, V.C.; Markwald, R.R.; Ghatak, S. Interactions between Hyaluronan and Its Receptors (Cd44, Rhamm) Regulate the Activities of Inflammation and Cancer. Front. Immunol. 2015, 6, 201. [Google Scholar] [CrossRef] [Green Version]
- Bourguignon, L.Y.; Singleton, P.A.; Zhu, H.; Diedrich, F. Hyaluronan-Mediated Cd44 Interaction with Rhogef and Rho Kinase Promotes Grb2-Associated Binder-1 Phosphorylation and Phosphatidylinositol 3-Kinase Signaling Leading to Cytokine (Macrophage-Colony Stimulating Factor) Production and Breast Tumor Progression. J. Biol. Chem. 2003, 278, 29420–29434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okamoto, I.; Tsuiki, H.; Kenyon, L.C.; Godwin, A.K.; Emlet, D.R.; Holgado-Madruga, M.; Lanham, I.S.; Joynes, C.J.; Vo, K.T.; Guha, A.; et al. Proteolytic Cleavage of the Cd44 Adhesion Molecule in Multiple Human Tumors. Am. J. Pathol. 2002, 160, 441–447. [Google Scholar] [CrossRef] [Green Version]
- Sugahara, K.N.; Murai, T.; Nishinakamura, H.; Kawashima, H.; Saya, H.; Miyasaka, M. Hyaluronan Oligosaccharides Induce Cd44 Cleavage and Promote Cell Migration in Cd44-Expressing Tumor Cells. J. Biol. Chem. 2003, 278, 32259–32265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kato, Y.; Hayatsu, N.; Kaneko, M.K.; Ogasawara, S.; Hamano, T.; Takahashi, S.; Nishikawa, R.; Matsutani, M.; Mishima, K.; Narimatsu, H. Increased Expression of Highly Sulfated Keratan Sulfate Synthesized in Malignant Astrocytic Tumors. Biochem. Biophys. Res. Commun. 2008, 369, 1041–1046. [Google Scholar] [CrossRef]
- Chhabra, M.; Doherty, G.G.; See, N.W.; Gandhi, N.S.; Ferro, V. From Cancer to COVID-19: A Perspective on Targeting Heparan Sulfate-Protein Interactions. Chem. Rec. 2021, 21, 1–16. [Google Scholar] [CrossRef]
- Cole, J.B.; Florez, J.C. Genetics of Diabetes Mellitus and Diabetes Complications. Nat. Rev. Nephrol. 2020, 16, 377–390. [Google Scholar] [CrossRef]
- Yang, Q.Q.; Sun, J.W.; Shao, D.; Zhang, H.H.; Bai, C.F.; Cao, F.L. The Association between Diabetes Complications, Diabetes Distress, and Depressive Symptoms in Patients with Type 2 Diabetes Mellitus. Clin. Nurs. Res. 2021, 30, 293–301. [Google Scholar] [CrossRef]
- Gowd, V.; Gurukar, A.; Chilkunda, N.D. Glycosaminoglycan Remodeling During Diabetes and the Role of Dietary Factors in Their Modulation. World J. Diabetes 2016, 7, 67–73. [Google Scholar] [CrossRef]
- Irving-Rodgers, H.F.; Ziolkowski, A.F.; Parish, C.R.; Sado, Y.; Ninomiya, Y.; Simeonovic, C.J.; Rodgers, R.J. Molecular Composition of the Peri-Islet Basement Membrane in Nod Mice: A Barrier against Destructive Insulitis. Diabetologia 2008, 51, 1680–1688. [Google Scholar] [CrossRef] [Green Version]
- Ziolkowski, A.F.; Popp, S.K.; Freeman, C.; Parish, C.R.; Simeonovic, C.J. Heparan Sulfate and Heparanase Play Key Roles in Mouse Beta Cell Survival and Autoimmune Diabetes. J. Clin. Investig. 2012, 122, 132–141. [Google Scholar] [CrossRef] [Green Version]
- Simeonovic, C.J.; Popp, S.K.; Starrs, L.M.; Brown, D.J.; Ziolkowski, A.F.; Ludwig, B.; Bornstein, S.R.; Wilson, J.D.; Pugliese, A.; Kay, T.W.H.; et al. Loss of Intra-Islet Heparan Sulfate Is a Highly Sensitive Marker of Type 1 Diabetes Progression in Humans. PLoS ONE 2018, 13, e0191360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Theodoraki, A.; Hu, Y.; Poopalasundaram, S.; Oosterhof, A.; Guimond, S.E.; Disterer, P.; Khoo, B.; Hauge-Evans, A.C.; Jones, P.M.; Turnbull, J.E.; et al. Distinct Patterns of Heparan Sulphate in Pancreatic Islets Suggest Novel Roles in Paracrine Islet Regulation. Mol. Cell. Endocrinol. 2015, 399, 296–310. [Google Scholar] [CrossRef] [PubMed]
- Simeonovic, C.J.; Ziolkowski, A.F.; Wu, Z.; Choong, F.J.; Freeman, C.; Parish, C.R. Heparanase and Autoimmune Diabetes. Front. Immunol. 2013, 4, 471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simeonovic, C.J.; Popp, S.K.; Brown, D.K.; Li, F.J.; Lafferty, A.R.A.; Freeman, C.; Parish, C.R. Heparanase and Type 1 Diabetes. Adv. Exp. Med. Biol. 2020, 1221, 607–630. [Google Scholar]
- Raats, C.J.; Bakker, M.A.; van den Born, J.; Berden, J.H. Hydroxyl Radicals Depolymerize Glomerular Heparan Sulfate in Vitro and in Experimental Nephrotic Syndrome. J. Biol. Chem. 1997, 272, 26734–26741. [Google Scholar] [CrossRef] [Green Version]
- Rota, C.; Liverani, L.; Spelta, F.; Mascellani, G.; Tomasi, A.; Iannone, A.; Vismara, E. Free Radical Generation During Chemical Depolymerization of Heparin. Anal. Biochem. 2005, 344, 193–203. [Google Scholar] [CrossRef]
- Dhounchak, S.; Popp, S.K.; Brown, D.J.; Laybutt, D.R.; Biden, T.J.; Bornstein, S.R.; Parish, C.R.; Simeonovic, C.J. Heparan Sulfate Proteoglycans in Beta Cells Provide a Critical Link between Endoplasmic Reticulum Stress, Oxidative Stress and Type 2 Diabetes. PLoS ONE 2021, 16, e0252607. [Google Scholar] [CrossRef]
- Liu, Z.; Lv, Y.; Zhao, N.; Guan, G.; Wang, J. Protein Kinase R-Like Er Kinase and Its Role in Endoplasmic Reticulum Stress-Decided Cell Fate. Cell Death Dis. 2015, 6, e1822. [Google Scholar] [CrossRef] [Green Version]
- Scheuner, D.; Kaufman, R. The Unfolded Protein Response: A Pathway That Links Insulin Demand with Beta-Cell Failure and Diabetes. Endocr. Rev. 2008, 29, 631. [Google Scholar] [CrossRef] [Green Version]
- Laybutt, D.R.; Preston, A.M.; Akerfeldt, M.C.; Kench, J.G.; Busch, A.K.; Biankin, A.V.; Biden, T.J. Endoplasmic Reticulum Stress Contributes to Beta Cell Apoptosis in Type 2 Diabetes. Diabetologia 2007, 50, 752–763. [Google Scholar] [CrossRef] [Green Version]
- Chan, J.Y.; Luzuriaga, J.; Bensellam, M.; Biden, T.J.; Laybutt, D.R. Failure of the Adaptive Unfolded Protein Response in Islets of Obese Mice Is Linked with Abnormalities in Β-Cell Gene Expression and Progression to Diabetes. Diabetes 2013, 62, 1557–1568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asthana, S.; Mallick, B.; Alexandrescu, A.T.; Jha, S. Iapp in Type Ii Diabetes: Basic Research on Structure, Molecular Interactions, and Disease Mechanisms Suggests Potential Intervention Strategies. Biochim. Biophys. Acta Biomembr. 2018, 1860, 1765–1782. [Google Scholar] [CrossRef] [PubMed]
- De Carufel, C.A.; Nguyen, P.T.; Sahnouni, S.; Bourgault, S. New Insights into the Roles of Sulfated Glycosaminoglycans in Islet Amyloid Polypeptide Amyloidogenesis and Cytotoxicity. Biopolymers 2013, 100, 645–655. [Google Scholar] [CrossRef] [PubMed]
- Castillo, G.M.; Cummings, J.A.; Yang, W.; Judge, M.E.; Sheardown, M.J.; Rimvall, K.; Hansen, J.B.; Snow, A.D. Sulfate Content and Specific Glycosaminoglycan Backbone of Perlecan Are Critical for Perlecan’s Enhancement of Islet Amyloid Polypeptide (Amylin) Fibril Formation. Diabetes 1998, 47, 612–620. [Google Scholar] [CrossRef]
- Quittot, N.; Fortier, M.; Babych, M.; Nguyen, P.T.; Sebastiao, M.; Bourgault, S. Cell Surface Glycosaminoglycans Exacerbate Plasma Membrane Perturbation Induced by the Islet Amyloid Polypeptide. FASEB J. 2021, 35, e21306. [Google Scholar] [CrossRef]
- Wang, H.; Raleigh, D.P. The Ability of Insulin to Inhibit the Formation of Amyloid by Pro-Islet Amyloid Polypeptide Processing Intermediates Is Significantly Reduced in the Presence of Sulfated Glycosaminoglycans. Biochemistry 2014, 53, 2605–2614. [Google Scholar] [CrossRef]
- Bogdani, M.; Johnson, P.Y.; Potter-Perigo, S.; Nagy, N.; Day, A.J.; Bollyky, P.L.; Wight, T.N. Ha-Hyaluronan and Hyaluronan-Binding Proteins Accumulate in Both Human Type 1 Diabetic Islets and Lymphoid Tissues and Associate with Inflammatory Cells in Insulitis. Diabetes 2014, 63, 2727–2743. [Google Scholar] [CrossRef]
- Bogdani, M.; Simeonovic, C.; Nagy, N.; Johnson, P.Y.; Chan, C.K.; Wight, T.N. The Detection of Glycosaminoglycans in Pancreatic Islets and Lymphoid Tissues. Methods Mol. Biol. 2015, 1229, 413–430. [Google Scholar]
- Poplawska-Kita, A.; Mierzejewska-Iwanowska, B.; Szelachowska, M.; Siewko, K.; Nikolajuk, A.; Kinalska, I.; Gorska, M. Glycosaminoglycans Urinary Excretion as a Marker of the Early Stages of Diabetic Nephropathy and the Disease Progression. Diabetes Metab. Res. Rev. 2008, 24, 310–317. [Google Scholar] [CrossRef]
- Parthasarathy, N.; Spiro, R.G. Effect of Diabetes on the Glycosaminoglycan Component of the Human Glomerular Basement Membrane. Diabetes 1982, 31, 738–741. [Google Scholar] [CrossRef]
- Wijnhoven, T.J.; van den Hoven, M.J.; Ding, H.; van Kuppevelt, T.H.; van der Vlag, J.; Berden, J.H.; Prinz, R.A.; Lewis, E.J.; Schwartz, M.; Xu, X. Heparanase Induces a Differential Loss of Heparan Sulphate Domains in Overt Diabetic Nephropathy. Diabetologia 2008, 51, 372–382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schumacher, V.A.; Schlotzer-Schrehardt, U.; Karumanchi, S.A.; Shi, X.; Zaia, J.; Jeruschke, S.; Zhang, D.; Pavenstadt, H.; Drenckhan, A.; Amann, K.; et al. Wt1-Dependent Sulfatase Expression Maintains the Normal Glomerular Filtration Barrier. J. Am. Soc. Nephrol. 2011, 22, 1286–1296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lauer, M.E.; Hascall, V.C.; Wang, A. Heparan Sulfate Analysis from Diabetic Rat Glomeruli. J. Biol. Chem. 2007, 282, 843–852. [Google Scholar] [CrossRef] [Green Version]
- Van den Born, J.; Pisa, B.; Bakker, M.A.; Celie, J.W.; Straatman, C.; Thomas, S.; Viberti, G.C.; Kjellen, L.; Berden, J.H. No Change in Glomerular Heparan Sulfate Structure in Early Human and Experimental Diabetic Nephropathy. J. Biol. Chem. 2006, 281, 29606–29613. [Google Scholar] [CrossRef] [Green Version]
- Reine, T.M.; Grondahl, F.; Jenssen, T.G.; Hadler-Olsen, E.; Prydz, K.; Kolset, S.O. Reduced Sulfation of Chondroitin Sulfate but Not Heparan Sulfate in Kidneys of Diabetic Db/Db Mice. J. Histochem. Cytochem. 2013, 61, 606–616. [Google Scholar] [CrossRef] [Green Version]
- Joladarashi, D.; Salimath, P.V.; Chilkunda, N.D. Diabetes Results in Structural Alteration of Chondroitin Sulfate/Dermatan Sulfate in the Rat Kidney:Effects on the Binding to Extracellular Matrix Components. Glycobiology 2011, 21, 960–972. [Google Scholar] [CrossRef] [Green Version]
- Jura-Poltorak, A.; Olczyk, P.; Chalas-Lipka, A.; Komosinska-Vassev, K.; Kuznik-Trocha, K.; Winsz-Szczotka, K.; Ivanova, D.; Kiselova-Kaneva, Y.; Krysik, K.; Telega, A.; et al. Urinary Sulphated Glycosaminoglycans Excretion in Obese Patients with Type 2 Diabetes Mellitus Treated with Metformin. Arch. Physiol. Biochem. 2022, 128, 507–513. [Google Scholar] [CrossRef] [PubMed]
- Zhao, T.; Lu, X.; Davies, N.; Gong, Y.; Guo, J.; Zhang, H.; Li, Z.; Hong, J.; Fu, G.; Li, P. Diabetes Results in Structural Alteration of Chondroitin Sulfate in the Urine. J. Pharm. Sci. 2013, 16, 486–493. [Google Scholar] [CrossRef] [PubMed]
- Srikanth, C.B.; Salimath, P.V.; Nandini, C.D. Erythrocytes Express Chondroitin Sulphate/Dermatan Sulphate, Which Undergoes Quantitative Changes During Diabetes and Mediate Erythrocyte Adhesion to Extracellular Matrix Components. Biochimie 2012, 94, 1347–1355. [Google Scholar] [CrossRef]
- Morofuji, Y.; Nakagawa, S. Drug Development for Central Nervous System Diseases Using in Vitro Blood-Brain Barrier Models and Drug Repositioning. Curr. Pharm. Des. 2020, 26, 1466–1485. [Google Scholar] [CrossRef]
- Cenini, G.; Voos, W. Mitochondria as Potential Targets in Alzheimer Disease Therapy: An Update. Front. Pharmacol. 2019, 10, 902. [Google Scholar] [CrossRef] [PubMed]
- Livingston, G.; Sommerlad, A.; Orgeta, V.; Costafreda, S.G.; Huntley, J.; Ames, D.; Ballard, C.; Banerjee, S.; Burns, A.; Cohen-Mansfield, J.; et al. Dementia Prevention, Intervention, and Care. Lancet 2017, 390, 2673–2734. [Google Scholar] [CrossRef] [Green Version]
- Hardy, J.A.; Higgins, G.A. Alzheimer’s Disease the Amyloid Cascade Hypothesis. Science 1992, 256, 184–185. [Google Scholar] [CrossRef] [PubMed]
- Krishnaswamy, V.R.; Benbenishty, A.; Blinder, P.; Sagi, I. Demystifying the Extracellular Matrix and Its Proteolytic Remodeling in the Brain: Structural and Functional Insights. Cell. Mol. Life Sci. 2019, 76, 3229–3248. [Google Scholar] [CrossRef]
- Bonneh-Barkay, D.; Wiley, C.A. Brain Extracellular Matrix in Neurodegeneration. Brain Pathol. 2009, 19, 573–585. [Google Scholar] [CrossRef]
- Bandtlow, C.E.; Zimmermann, D.R. Proteoglycans in the Developing Brain New Conceptual Insights for Old Proteins. Physiol. Rev. 2000, 80, 1267–1290. [Google Scholar] [CrossRef] [Green Version]
- Smith, P.D.; Coulson-Thomas, V.J.; Foscarin, S.; Kwok, J.C.; Fawcett, J.W. “Gag-Ing with the Neuron”: The Role of Glycosaminoglycan Patterning in the Central Nervous System. Exp. Neurol. 2015, 274, 100–114. [Google Scholar] [CrossRef]
- Soleman, S.; Filippov, M.A.; Dityatev, A.; Fawcett, J.W. Targeting the Neural Extracellular Matrix in Neurological Disorders. Neuroscience 2013, 253, 194–213. [Google Scholar] [CrossRef]
- Carulli, D.; Laabs, T.; Geller, H.M.; Fawcett, J.W. Chondroitin Sulfate Proteoglycans in Neural Development and Regeneration. Curr. Opin. Neurobiol. 2005, 15, 116–120. [Google Scholar] [CrossRef]
- Logsdon, A.F.; Francis, K.L.; Richardson, N.E.; Hu, S.J.; Faber, C.L.; Phan, B.A.; Nguyen, V.; Setthavongsack, N.; Banks, W.A.; Woltjer, R.L.; et al. Decoding Perineuronal Net Glycan Sulfation Patterns in the Alzheimer’s Disease Brain. Alzheimers Dement. 2022, 18, 942–954. [Google Scholar] [CrossRef]
- Bertolotto, A.; Rocca, G.; Schiffer, D. Chondroitin-4-Sulfate Proteoglycan Forms an Extracellular Network in Human and Rat Central Nervous System. J. Neurol. Sci. 1990, 100, 113–123. [Google Scholar] [CrossRef]
- Zhao, J.; Zhu, Y.; Song, X.; Xiao, Y.; Su, G.; Liu, X.; Wang, Z.; Xu, Y.; Liu, J.; Eliezer, D.; et al. 3-O-Sulfation of Heparan Sulfate Enhances Tau Interaction and Cellular Uptake. Angew. Chem. Int. Ed. Engl. 2020, 59, 1818–1827. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Katagiri, Y.; McCann, T.E.; Unsworth, E.; Goldsmith, P.; Yu, Z.X.; Tan, F.; Santiago, L.; Mills, E.M.; Wang, Y.; et al. Chondroitin-4-Sulfation Negatively Regulates Axonal Guidance and Growth. J. Cell Sci. 2008, 121, 3083–3091. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shioi, J.; Anderson, J.P.; Ripellino, J.A.; Robakis, N.K. Chondroitin Sulfate Proteoglycan Form of the Alzheimer’s Beta-Amyloid Precursor. J. Biol. Chem. 1992, 267, 13819–13822. [Google Scholar] [CrossRef]
- Mehra, S.; Ghosh, D.; Kumar, R.; Mondal, M.; Gadhe, L.G.; Das, S.; Anoop, A.; Jha, N.N.; Jacob, R.S.; Chatterjee, D.; et al. Glycosaminoglycans Have Variable Effects on Alpha-Synuclein Aggregation and Differentially Affect the Activities of the Resulting Amyloid Fibrils. J. Biol. Chem. 2018, 293, 12975–12991. [Google Scholar] [CrossRef] [Green Version]
- DeWitt, D.A.; Silver, J.; Canning, D.R.; Perry, G. Chondroitin Sulfate Proteoglycans Are Associated with the Lesions of Alzheimer’s Disease. Exp. Neurol. 1993, 121, 149–152. [Google Scholar] [CrossRef] [Green Version]
- Sobel, R.A.; Ahmed, A.S. White Matter Extracellular Matrix Chondroitin Sulfate/Dermatan Sulfate Proteoglycans in Multiple Sclerosis. J. Neuropathol. Exp. Neurol. 2001, 60, 1198–1207. [Google Scholar] [CrossRef] [Green Version]
- Stephenson, E.L.; Zhang, P.; Ghorbani, S.; Wang, A.; Gu, J.; Keough, M.B.; Rawji, K.S.; Silva, C.; Yong, V.W.; Ling, C.C. Targeting the Chondroitin Sulfate Proteoglycans: Evaluating Fluorinated Glucosamines and Xylosides in Screens Pertinent to Multiple Sclerosis. ACS Cent. Sci. 2019, 5, 1223–1234. [Google Scholar] [CrossRef]
- Keough, M.B.; Rogers, J.A.; Zhang, P.; Jensen, S.K.; Stephenson, E.L.; Chen, T.; Hurlbert, M.G.; Lau, L.W.; Rawji, K.S.; Plemel, J.R.; et al. An Inhibitor of Chondroitin Sulfate Proteoglycan Synthesis Promotes Central Nervous System Remyelination. Nat. Commun. 2016, 7, 11312. [Google Scholar] [CrossRef] [Green Version]
- Warford, J.R.; Lamport, A.C.; Clements, D.R.; Malone, A.; Kennedy, B.E.; Kim, Y.; Gujar, S.A.; Hoskin, D.W.; Easton, A.S. Surfen, a Proteoglycan Binding Agent, Reduces Inflammation but Inhibits Remyelination in Murine Models of Multiple Sclerosis. Acta Neuropathol. Commun. 2018, 6, 4. [Google Scholar] [CrossRef] [Green Version]
- Lau, L.W.; Cua, R.; Keough, M.B.; Haylock-Jacobs, S.; Yong, V.W. Pathophysiology of the Brain Extracellular Matrix: A New Target for Remyelination. Nat. Rev. Neurosci. 2013, 14, 722–729. [Google Scholar] [CrossRef] [PubMed]
- Pendleton, J.C.; Shamblott, M.J.; Gary, D.S.; Belegu, V.; Hurtado, A.; Malone, M.L.; McDonald, J.W. Chondroitin Sulfate Proteoglycans Inhibit Oligodendrocyte Myelination through Ptpsigma. Exp. Neurol. 2013, 247, 113–121. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, R.J.; McKeon, R.J.; Darr, A.; Calabro, A.; Hascall, V.C.; Bellamkonda, R.V. Cs-4,6 Is Differentiallyupregulated in Glial Scar and Is a Potent Inhibitor of Neurite Extension. Mol. Cell. Neurosci. 2005, 29, 545–558. [Google Scholar] [CrossRef]
- Maiza, A.; Chantepie, S.; Vera, C.; Fifre, A.; Huynh, M.B.; Stettler, O.; Ouidja, M.O.; Papy-Garcia, D. The Role of Heparan Sulfates in Protein Aggregation and Their Potential Impact on Neurodegeneration. FEBS Lett. 2018, 592, 3806–3818. [Google Scholar] [CrossRef] [Green Version]
- Yamazaki, T.; Iharass, Y. The Molecular Pathology of Alzheimer’s Disease. Neuropathology 1997, 17, 263–269. [Google Scholar] [CrossRef]
- Shimizu, H.; Ghazizadeh, M.; Sato, S.; Oguro, T.; Kawanami, O. Interaction between Beta-Amyloid Protein and Heparan Sulfate Proteoglycans from the Cerebral Capillary Basement Membrane in Alzheimer’s Disease. J. Clin. Neurosci. 2009, 16, 277–2782. [Google Scholar] [CrossRef] [PubMed]
- Bruinsma, I.B.; Te Riet, L.; Gevers, T.; ten Dam, G.B.; van Kuppevelt, T.H.; David, G.; Kusters, B.; de Waal, R.M.; Verbeek, M.M. Sulfation of Heparan Sulfate Associated with Amyloid-Beta Plaques in Patients with Alzheimer’s Disease. Acta Neuropathol. 2010, 119, 211–220. [Google Scholar] [CrossRef]
- Lindahl, B.; Westling, C.; Gimenez-Gallego, G.; Lindahl, U.; Salmivirta, M. Common Binding Sites for Beta-Amyloid Fibrils and Fibroblast Growth Factor-2 in Heparan Sulfate from Human Cerebral Cortex. J. Biol. Chem. 1999, 274, 30631–30635. [Google Scholar] [CrossRef]
- Zhou, X.; Wang, Y.; Zheng, W.; Deng, G.; Wang, F.; Jin, L. Characterizing Heparin Tetrasaccharides Binding to Amyloid-Beta Peptide. Front. Mol. Biosci. 2022, 9, 824146. [Google Scholar] [CrossRef]
- Konno, T.; Oiki, S.; Hasegawa, K.; Naiki, H. Anionic Contribution for Fibrous Maturation of Protofibrillar Assemblies of the Human Tau Repeat Domain in a Fluoroalcohol Solution. Biochemistry 2004, 43, 13613–13620. [Google Scholar] [CrossRef]
- Liu, I.H.; Uversky, V.N.; Munishkina, L.A.; Fink, A.L.; Halfter, W.; Cole, G.J. Agrin Binds Alpha-Synuclein and Modulates Alpha-Synuclein Fibrillation. Glycobiology 2005, 15, 1320–1331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tao, Y.; Sun, Y.; Lv, S.; Xia, W.; Zhao, K.; Xu, Q.; Zhao, Q.; He, L.; Le, W.; Wang, Y.; et al. Heparin Induces Alpha-Synuclein to Form New Fibril Polymorphs with Attenuated Neuropathology. Nat. Commun. 2022, 13, 4226. [Google Scholar] [CrossRef] [PubMed]
- Ihse, E.; Yamakado, H.; van Wijk, X.M.; Lawrence, R.; Esko, J.D.; Masliah, E. Cellular Internalization of Alpha-Synuclein Aggregates by Cell Surface Heparan Sulfate Depends on Aggregate Conformation and Cell Type. Sci. Rep. 2017, 7, 9008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DeWitt, D.A.; Richey, P.L.; Praprotnik, D.; Silver, J.; Perry, G. Chondroitin Sulfate Proteoglycans Are a Common Component of Neuronal Inclusions and Astrocytic Reaction in Neurodegenerative Diseases. Brain Res. 1994, 656, 205–209. [Google Scholar] [CrossRef] [Green Version]
- Ling, J.; Li, J.; Khan, A.; Lundkvist, A.; Li, J.P. Is Heparan Sulfate a Target for Inhibition of Rna Virus Infection? Am. J. Physiol. Cell Physiol. 2022, 322, C605–C613. [Google Scholar] [CrossRef]
- Shajahan, A.; Pepi, L.E.; Rouhani, D.S.; Heiss, C.; Azadi, P. Glycosylation of Sars-Cov-2: Structural and Functional Insights. Anal. Bioanal. Chem. 2021, 413, 7179–7193. [Google Scholar] [CrossRef]
- Sorin, M.N.; Kuhn, J.; Stasiak, A.C.; Stehle, T. Structural Insight into Non-Enveloped Virus Binding to Glycosaminoglycan Receptors: A Review. Viruses 2021, 13, 800. [Google Scholar] [CrossRef]
- Clausen, T.M.; Sandoval, D.R.; Spliid, C.B.; Pihl, J.; Perrett, H.R.; Painter, C.D.; Narayanan, A.; Majowicz, S.A.; Kwong, E.M.; McVicar, R.N.; et al. SARS-CoV-2 Infection Depends on Cellular Heparan Sulfate and Ace2. Cell 2020, 183, 1043–1057.e15. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.Y.; Jin, W.; Sood, A.; Montgomery, D.W.; Grant, O.C.; Fuster, M.M.; Fu, L.; Dordick, J.S.; Woods, R.J.; Zhang, F.; et al. Glycosaminoglycan Binding Motif at S1/S2 Proteolytic Cleavage Site on Spike Glycoprotein May Facilitate Novel Coronavirus (SARS-CoV-2) Host Cell Entry. Mol. Biol. 2020. [Google Scholar] [CrossRef] [Green Version]
- Kearns, F.L.; Sandoval, D.R.; Casalino, L.; Clausen, T.M.; Rosenfeld, M.A.; Spliid, C.B.; Amaro, R.E.; Esko, J.D. Spike-Heparan Sulfate Interactions in SARS-CoV-2 Infection. Curr. Opin. Struc. Biol. 2022, 76, 102439. [Google Scholar] [CrossRef]
- Avirutnan, P.; Zhang, L.; Punyadee, N.; Manuyakorn, A.; Puttikhunt, C.; Kasinrerk, W.; Malasit, P.; Atkinson, J.P.; Diamond, M.S. Secreted Ns1 of Dengue Virus Attaches to the Surface of Cells Via Interactions with Heparan Sulfate and Chondroitin Sulfate E. PLoS Pathog. 2007, 3, e183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dalrymple, N.; Mackow, E.R. Productive Dengue Virusinfection of Human Endothelial Cells Is Directed by Heparan Sulfate-Containing Proteoglycan Receptors. J. Virol. 2011, 85, 9478–9485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barth, H.; Schafer, C.; Adah, M.I.; Zhang, F.; Linhardt, R.J.; Toyoda, H.; Kinoshita-Toyoda, A.; Toida, T.; van Kuppevelt, T.H.; Depla, E.; et al. Cellular Binding of Hepatitis C Virus Envelope Glycoprotein E2 Requires Cell Surface Heparan Sulfate. J. Biol. Chem. 2003, 278, 41003–41012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uyama, T.; Ishida, M.; Izumikawa, T.; Trybala, E.; Tufaro, F.; Bergstrom, T.; Sugahara, K.; Kitagawa, H. Chondroitin 4-O-Sulfotransferase-1 Regulates E Disaccharide Expression of Chondroitin Sulfate Required for Herpes Simplex Virus Infectivity. J. Biol. Chem. 2006, 281, 38668–38674. [Google Scholar] [CrossRef]
- Abidine, Y.; Liu, L.; Wallen, O.; Trybala, E.; Olofsson, S.; Bergstrom, T.; Bally, M. Cellular Chondroitin Sulfate and the Mucin-Like Domain of Viral Glycoprotein C Promote Diffusion of Herpes Simplex Virus 1 While Heparan Sulfate Restricts Mobility. Viruses 2022, 14, 1836. [Google Scholar] [CrossRef] [PubMed]
- Kines, R.C.; Thompson, C.D.; Lowy, D.R.; Schiller, J.T.; Day, P.M. The Initial Steps Leading to Papillomavirus Infection Occur on the Basement Membrane Prior to Cell Surface Binding. Proc. Natl. Acad. Sci. USA 2009, 106, 20458–20463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alcorn, M.D.H.; Klimstra, W.B. Glycosaminoglycan Binding by Arboviruses: A Cautionary Tale. J. Gen. Virol. 2022, 103. [Google Scholar] [CrossRef]
- Klenk, K.; Roberts, S.R. Use of a Vesicular Stomatitis Virus Complementation System to Analyze Respiratory Syncytial Virus Binding. Virus Res. 2002, 90, 327–335. [Google Scholar] [CrossRef]
- Shi, D.; He, P.; Song, Y.; Cheng, S.; Linhardt, R.J.; Dordick, J.S.; Chi, L.; Zhang, F. Kinetic and Structural Aspects of Glycosaminoglycan-Monkeypox Virus Protein A29 Interactions Using Surface Plasmon Resonance. Molecules 2022, 27, 5898. [Google Scholar] [CrossRef]
- Kim, S.Y.; Jin, W.; Sood, A.; Montgomery, D.W.; Grant, O.C.; Fuster, M.M.; Fu, L.; Dordick, J.S.; Woods, R.J.; Zhang, F.; et al. Characterization of Heparin and Severe Acute Respiratory Syndrome-Related Coronavirus 2 (SARS-CoV-2) Spike Glycoprotein Binding Interactions. Antivir. Res. 2020, 181, 104873. [Google Scholar] [CrossRef]
- Tiwari, V.; Tandon, R.; Sankaranarayanan, N.V.; Beer, J.C.; Kohlmeir, E.K.; Swanson-Mungerson, M.; Desai, U.R. Preferential Recognition and Antagonism of Sars-Cov-2 Spike Glycoprotein Binding to 3-O-Sulfated Heparan Sulfate. bioRxiv 2020. [Google Scholar] [CrossRef]
- Du Preez, H.N.; Aldous, C.; Hayden, M.R.; Kruger, H.G.; Lin, J. Pathogenesis of Covid-19 Described through the Lens of an Undersulfated and Degraded Epithelial and Endothelial Glycocalyx. FASEB J. 2022, 36, e22052. [Google Scholar] [CrossRef] [PubMed]
- Andonegui-Elguera, S.; Taniguchi-Ponciano, K.; Gonzalez-Bonilla, C.R.; Torres, J.; Mayani, H.; Herrera, L.A.; Pena-Martinez, E.; Silva-Roman, G.; Vela-Patino, S.; Ferreira-Hermosillo, A.; et al. Molecular Alterations Prompted by SARS-CoV-2 Infection: Induction of Hyaluronan, Glycosaminoglycan and Mucopolysaccharide Metabolism. Arch. Med. Res. 2020, 51, 645–653. [Google Scholar] [CrossRef] [PubMed]
- Burgess, J.K.; Harmsen, M.C. Chronic Lung Diseases: Entangled in Extracellular Matrix. Eur. Respir. Rev. 2022, 31, 210202. [Google Scholar] [CrossRef]
- Ennemoser, M.; Pum, A.; Kungl, A. Disease-Specifc Glycosaminoglycan Patterns in the Extracellular Matrix of Human Lung and Brain. Carbohyd. Res. 2022, 511, 108480. [Google Scholar] [CrossRef]
- Westergren-Thorsson, G.; Hedstrom, U.; Nybom, A.; Tykesson, E.; Ahrman, E.; Hornfelt, M.; Maccarana, M.; van Kuppevelt, T.H.; Dellgren, G.; Wildt, M.; et al. Increased Deposition of Glycosaminoglycans and Altered Structure of Heparan Sulfate in Idiopathic Pulmonary Fibrosis. Int. J. Biochem. Cell Biol. 2017, 83, 27–38. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Olivenstein, R.; Taha, R.; Hamid, Q.; Ludwig, M. Enhanced Proteoglycan Deposition in the Airway Wall of Atopic Asthmatics. Am. J. Respir. Crit. Care Med. 1999, 160, 725–729. [Google Scholar] [CrossRef]
- Hallgren, O.; Alsafadi, H.N.; Nybom, A.; Zhou, X.; Bjermer, L.H.; Thorsson, G.W. Copd-Specific Chondroitin Sulfate Modifications Are Linked to Tgf-B1. Am. J. Respir. Crit. Care Med. 2019, 199, A5782. [Google Scholar]
- Reeves, E.P.; Williamson, M.; O’Neill, S.J.; Greally, P.; McElvaney, N.G. Nebulized Hypertonic Saline Decreases Il-8 in Sputum of Patients with Cystic Fibrosis. Am. J. Respir. Crit. Care Med. 2011, 183, 1517–1523. [Google Scholar] [CrossRef]
- Cheng, P.W.; Boat, T.F.; Cranfill, K.; Yankaskas, J.R.; BoucherR, C. Increased Sulfation of Glycoconjugates by Cultured Nasal Epithelial Cells from Patients with Cystic Fibrosis. J. Clin. Investig. 1989, 84, 68–72. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.S.; Werth, V.P. Identification of Specific Chondroitin Sulfate Species in Cutaneous Autoimmune Disease. J. Histochem. Cytochem. 2011, 59, 780–790. [Google Scholar] [CrossRef] [PubMed]
- Nikkilä, M.T. Urinary Glycosaminoglycan Excretion in Normal and Stone-Forming Subjects: Significant Disturbance in Recurrent Stone Formers. Urol. Int. 1989, 44, 157–159. [Google Scholar] [CrossRef]
- Verkoelen, C.F.; Verhulst, A. Proposed Mechanisms in Renal Tubular Crystal Retention. Kidney Int. 2007, 72, 13–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ustundağ, Y.; Huysal, K.; Guzelsoy, M.; Genim, C.E.; Yavuz, A. Urine and Serum Glycosaminoglycan Levels in the Diagnosis of Urological Diseases and Conditions: A Narrative Review of the Literature. Urol. J. 2021, 88, 103–109. [Google Scholar] [CrossRef] [PubMed]
- Shirane, Y.; Kurokawa, Y.; Miyashita, S.; Komatsu, H.; Kagawa, S. Study of Inhibition Mechanisms of Glycosaminoglycans on Calcium Oxalate Monohydrate Crystals by Atomic Force Microscopy. Urol. Res. 1999, 27, 426–431. [Google Scholar] [CrossRef]
- Suzuki, K.; Ryall, R.L. The Effect of Heparan Sulphate on the Crystallization of Calcium Oxalate in Undiluted, Ultrafiltered Human Urine. Br. J. Urol. 1996, 78, 15–21. [Google Scholar] [CrossRef]
- Gohel, M.D.I.; Shum, D.K.Y.; Tam, P.C. Electrophoretic Separation and Characterization of Urinary Glycosaminoglycans and Their Roles in Urolithiasis. Carbohyd. Res. 2007, 342, 79–86. [Google Scholar] [CrossRef]
- Dissayabutra, T.; Kalpongnukul, N.; Chindaphan, K.; Srisa-art, M.; Ungjaroenwathana, W.; Kaewwongse, M.; Lampenkhae, K.; Tosukhowong, P. Urinary Sulfated Glycosaminoglycan Insufficiency and Chondroitin Sulfate Supplement in Urolithiasis. PLoS ONE 2019, 14, e0213180. [Google Scholar] [CrossRef]
- Jappie, D.; Rodgers, A.; Ravenscroft, N.; Webber, D.; Gohel, M.D.I. Composition and Inhibitory Properties of Endogenous Urinary GAGs are Different in Subjects from Two Race Groups with Different Occurrence Rates of Kidney Stones: Pilot Studies Provide Unique Evidence in Support of an Inhibitory Role for This Group of Compounds. Clin. Chim. Acta 2022, 525, 84–90. [Google Scholar]
- Jappie, D.; Rodgers, A.; Webber, D.; Gohel, M.D.I. Seeking Consistency for the Role of Urinary Macromolecules and Glycosaminoglycans in Calcium Oxalate Crystallization Processes Pertaining to the Risk of Renal Stone Formation Using a Multi-Faceted Basic Science Approach. Clin. Chim. Acta 2021, 521, 76–84. [Google Scholar] [CrossRef]
- Pessentheiner, A.R.; Ducasa, G.M.; Gordts, P. Proteoglycans in Obesity-Associated Metabolic Dysfunction and Meta-Inflammation. Front. Immunol. 2020, 11, 769. [Google Scholar] [CrossRef] [PubMed]
- Hatano, S.; Watanabe, H. Regulation of Macrophage and Dendritic Cell Function by Chondroitin Sulfate in Innate to Antigen-Specific Adaptive Immunity. Front. Immunol. 2020, 11, 232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reeves, S.R.; Barrow, K.A.; Rich, L.M.; White, M.P.; Shubin, N.J.; Chan, C.K.; Kang, I.; Ziegler, S.F.; Piliponsky, A.M.; Wight, T.N.; et al. Respiratory Syncytial Virus Infection of Human Lung Fibroblasts Induces a Hyaluronan-Enriched Extracellular Matrix That Binds Mast Cells and Enhances Expression of Mast Cell Proteases. Front. Immunol. 2019, 10, 3159. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.; Arif, A.A.; Guo, J.; Ha, Z.; Lee-Sayer, S.S.M.; Poon, G.F.T.; Dosanjh, M.; Roskelley, C.D.; Huan, T.; Johnson, P. Cd44 Loss Disrupts Lung Lipid Surfactant Homeostasis and Exacerbates Oxidized Lipid-Induced Lung Inflammation. Front. Immunol. 2020, 11, 29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, B.; Chi, L. Chondroitin Sulfate/Dermatan Sulfate-Protein Interactions and Their Biological Functions in Human Diseases: Implications and Analytical Tools. Front. Cell Dev. Biol. 2021, 9, 693563. [Google Scholar] [CrossRef]
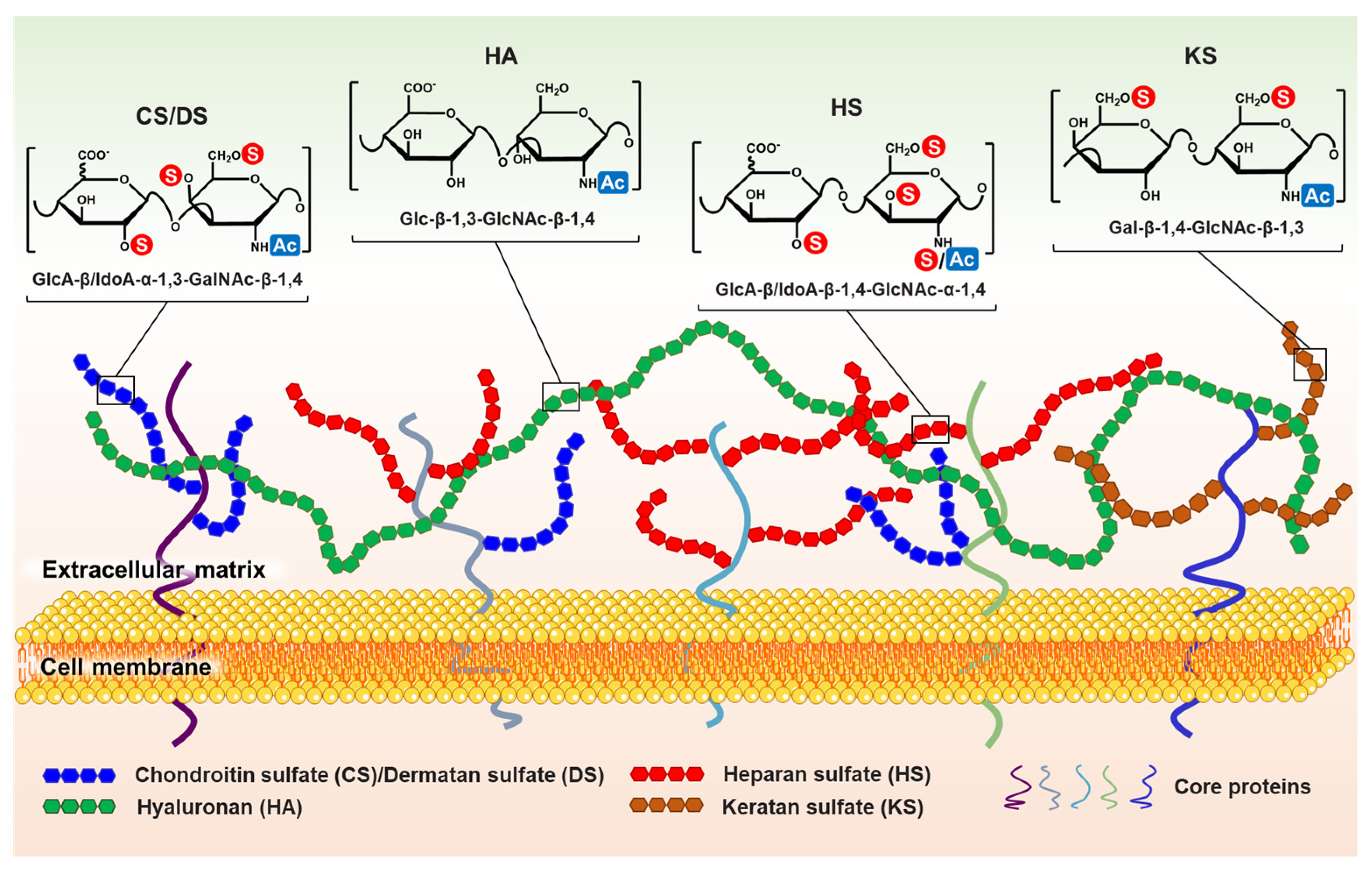
 ” and “
” and “ ” indicate that the hydroxyl can be substituted by a sulfate group and acetyl, respectively. It is worth noting that the structural formula represents the maximum degree of sulfation for each GAG type. “
” indicate that the hydroxyl can be substituted by a sulfate group and acetyl, respectively. It is worth noting that the structural formula represents the maximum degree of sulfation for each GAG type. “ ” only represents the possibility of being substituted by a sulfate group. Such as CS, they can be classified as CS-A (GlcA-GalNAc4S), CS-C (GlcA-GalNAc6S), CS-D (GlcA2S-GalNAc6S), CS-E (GlcA4S-GalNAc6S), etc. (Used with permission of Royal Society of Chemistry from ref. [1]; permission conveyed through Copyright Clearance Center, Inc).
” and “” indicate that the hydroxyl can be substituted by a sulfate group and acetyl, respectively. It is worth noting that the structural formula represents the maximum degree of sulfation for each GAG type. “” only represents the possibility of being substituted by a sulfate group. Such as CS, they can be classified as CS-A (GlcA-GalNAc4S), CS-C (GlcA-GalNAc6S), CS-D (GlcA2S-GalNAc6S), CS-E (GlcA4S-GalNAc6S), etc. (Used with permission of Royal Society of Chemistry from ref. [1]; permission conveyed through Copyright Clearance Center, Inc).
” only represents the possibility of being substituted by a sulfate group. Such as CS, they can be classified as CS-A (GlcA-GalNAc4S), CS-C (GlcA-GalNAc6S), CS-D (GlcA2S-GalNAc6S), CS-E (GlcA4S-GalNAc6S), etc. (Used with permission of Royal Society of Chemistry from ref. [1]; permission conveyed through Copyright Clearance Center, Inc).
” and “” indicate that the hydroxyl can be substituted by a sulfate group and acetyl, respectively. It is worth noting that the structural formula represents the maximum degree of sulfation for each GAG type. “” only represents the possibility of being substituted by a sulfate group. Such as CS, they can be classified as CS-A (GlcA-GalNAc4S), CS-C (GlcA-GalNAc6S), CS-D (GlcA2S-GalNAc6S), CS-E (GlcA4S-GalNAc6S), etc. (Used with permission of Royal Society of Chemistry from ref. [1]; permission conveyed through Copyright Clearance Center, Inc).


{kind=link}
{kind=link}
| Pathology Types | Study Objects (Cells/Tissues) | GAGs/PGs Alterations | References |
|---|---|---|---|
| Atherosclerotic type II | Aorta | Both the total CS content and the ratio of 6-O-S/4-O-S disaccharides in type II atherosclerosis arteries were increased | [50] |
| Atherosclerosis | Arterial smooth muscle cells | TGF-β prolongs CS chains in arterial smooth muscle cells and increases versican binding to LDL. | [51] |
| Atherosclerosis (symptomatic carotid stenosis) | Iliac arteries | The expression of perlecan gene decreased while versican gene remained unchanged. | [53] |
| Atherosclerosis and vascularrestenosis | Macrophages | Syndecan-1 protein level in macrophages was significantly decreased under hypoxia condition, and mRNA expression of key enzymes involved in HS biosynthesis in hypoxia cells was decreased. In addition, hypoxia also reduced the relative content of HS. | [58] |
| Atherosclerosis and vascularrestenosis | Aortic smooth muscle cells | HA and HA synthase are increased in senescent cells. HA accumulation promotes SMC metastasis via ERK1/2 modulation of the CD44 signaling pathway, resulting in intimal hyperplasia. | [61] |
| Atherosclerosis | Macrophages | LMW-HA induces macrophage/foam cell production and promotes atherosclerosis via the PKC pathway. LMW-HA also amplifies the migration of monocytes to inflammatory atherosclerotic plaques. | [65] |
| Breast cancer | Tumor tissue and plasma | Perlecan is absent in epithelial cell basement membrane while markedly upregulated in stroma. Furthermore, plasma perlecan level was significantly higher in estrogen receptor (ER)+ patients than ER- patients. | [78] |
| Prostate cancer | Perlecan expression is increased, which can the regulate sonic hedgehog signaling path. | [79] | |
| Neuroendocrine tumors (NETs) | Tumor tissue | Syndecan-2 is significantly increased in well-differentiated NETs and significantly decreased in poorly differentiated NETs. Glypican-5 was overexpressed in high-grade tumors with epithelial differentiation, but not in tumors with neuroendocrine phenotype. | [80] |
| Breast cancer | Tumor tissue | The GAG length increased by approximately 15% in tumor tissue compared to normal tissue. Both the 6-O-S CS and the total sulfation of HS increased. Compared to nonfatal breast cancers, the sulfation degree of HS, particularly 6-O-S, was decreased in fatal breast cancers, whereas the proportion of non-sulfated disaccharides was increased. | [87] |
| Glioblastoma multiforme (GBM) | Tumor tissue | 60–65% of GBM tumor samples showed increased levels of CS. A 1.5-fold increase in decorin, a 3-fold increase in biglycan and a 2-fold increase in serglycin. Only decorin levels were negatively associated with overall survival in GBM patients. | [115] |
| Pancreatic carcinoma | Tumor tissue | The total GAG level was increased by 4 times, HA increased 12 times, CS increased 22 times, DS increased 1.5 times. A significant increase in non-sulfate and 6-sulfate disaccharides of CS. | [127] |
| Pancreatic carcinoma | Tumor tissue | There are 27-fold and 7-fold increases in versican and decorin, respectively, compared with normal pancreases. The expression of 6-O-S and non-sulfated disaccharide units are enhanced. | [130] |
| Prostate cancer | Tumor tissue | Prostate cancer has an elevated 4-O-S CS content in the ECM, which may be due to inhibited androgen receptor (AR) signaling, thus resulting in increased 4-O-sulfotransferase CHST11 expression. | [131] |
| Gastric cancer | Tumor tissue | CS has a 10-fold increase in 6-O-S and non-disaccharide units, while 4-O-S disaccharides were correspondingly decreased. | [132] |
| Ovarian adenocarcinoma | Tumor tissue | High expression of CS-E in the ECM of ovarian adenocarcinoma enhances vascular endothelial growth factor (VEGF) mediation. | [133] |
| Lung cancer | Lewis lung carcinoma cells | The proportion of Δ4.5HexA-GalNAc-4, 6-O-disulfate was higher in highly metastatic lung cancer cell lines than in low metastatic cell lines. | [134] |
| Breast cancer | Tumor tissue | HA was significantly increased in 143 tumor tissue samples, indicating that HA is directly involved in breast cancer metastasis. | [143] |
| Breast cancer | Breast tumor cells (MDA-MB-231 cells) | LMW-HA activates actin filament-associated protein (AFAP-110) to bind to F-actin, resulting in nuclear translocation of myeloiddifferentiation factor (MyD88)/NF-xB and enhanced expression of pro-inflammatory cytokines IL-1β and IL-8. AFAP-110 binding with F-actin also promoted tumor cell metastasis. | [149] |
| Diabetes mellitus 1 type (T1D) | Pancreas | HSPGs such as Col18 and syndecan-1 showed significant loss in T1D human islets. | [162] |
| Diabetes mellitus | GAG (including perlecan) sulfation patterns affected amyloid fibril formation. The order of impact was heparin > N-desulfated acetylated heparin > fully desulfated N-sulfated heparin > fully desulfated N-acetylated heparin. | [175] | |
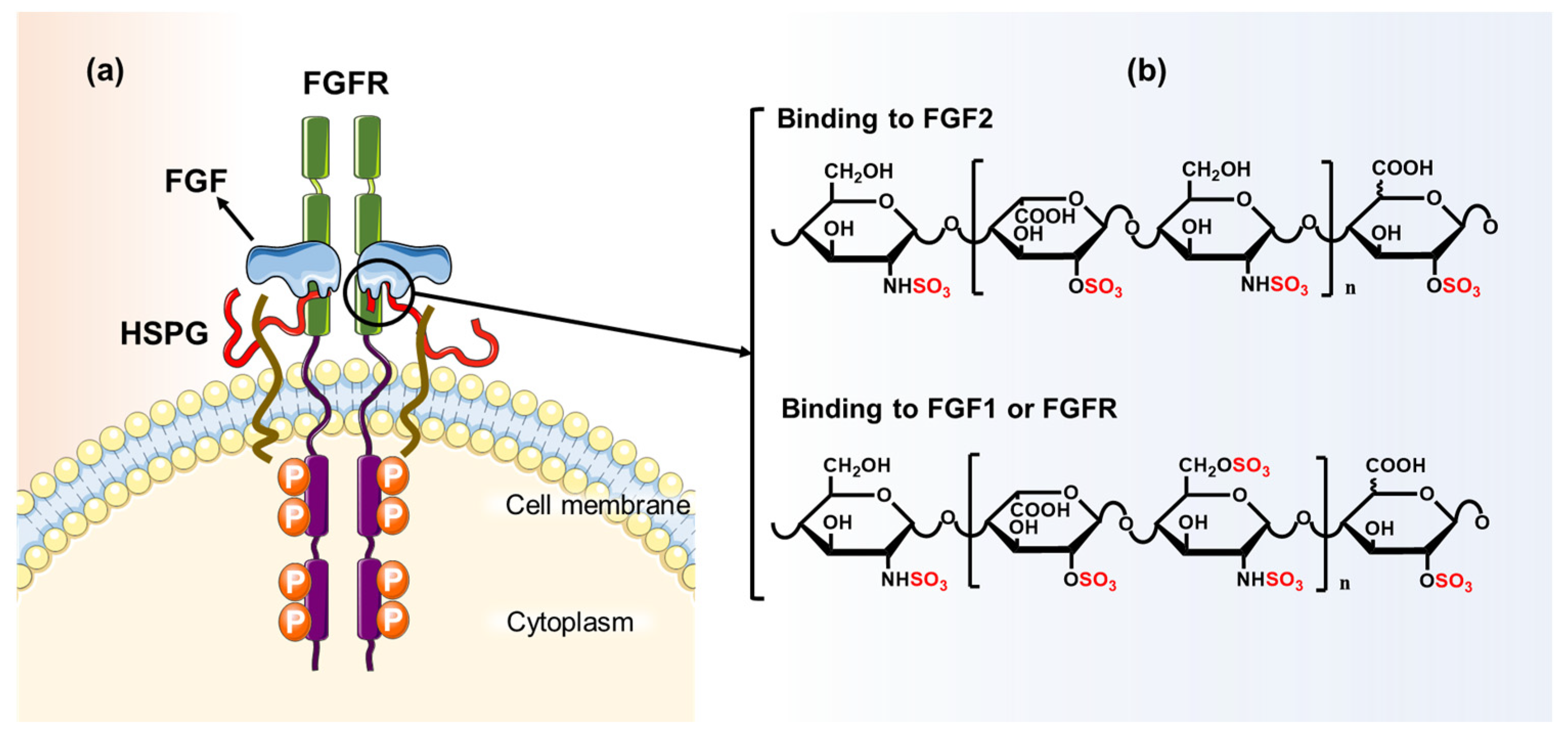
| Diabetic nephropathy | Glomerulus | Heparin sulfate 6-O-S plays an important role in extracellular matrix remodeling. Regulation of VEGFA and FGF2 signaling was achieved by increasing the expression of 6-O-endosulfatases Sulf1 and Sulf2 by the transcription factor Wilms’ Tumor 1 (WT1). | [183] |
| Diabetic nephropathy | Kidneys of rats | HS has less N-S in the GBM of diabetic rats compared to the normal group. | [184] |
| Diabetic nephropathy | Kidneys of rats | Renal CS/DS content decreased, accompanied by a decrease in the degree of sulfation, particularly 4, 6-O-sulfated GalN content. | [187] |
| Diabetic nephropathy | Renal cortex of diabetic db/db mice | 4-O-disaccharide sulfate significantly decreased from 65% to 40%, whereas 6-O-S disaccharide decreased from 11% to 6% and non-sulfated disaccharide increased from 21.5% to 51% in the renal cortex of diabetic db/db mice. | [186] |
| T2D | Urine | The contents of total GAGs, CS/DS, and HS in urine were significantly higher than those in healthy subjects. | [188] |
| Diabetes mellitus | Urine | 6-O-S and the 6-O-S/4-O-S ratio in the urine of diabetic patients with microalbuminuria were significantly increased compared with the healthy group. | [189] |
| Alzheimer’s disease (AD) | Brain | 4-O-S and 6-O-S CS are found in neurofibrillary tangles (NFTs) of AD patients, while only 4-O-S CS is found in senile plaques (SPs). | [207] |
| Multiple sclerosis | White matter | CSPGs (versican, neurocan, and aggrecan) and DSPGs were mainly located at the edge of active plaques, while the content of CSPGs in the active center of MS plaques was significantly decreased, possibly due to the internalization of PGs in PNNs by their foam macrophages together with myelin. | [208] |
| AD | Brain | PGs were more abundant in areas with amyloid plaques and neurofibrillary tangles. For example, relative to healthy individuals, the total PGs increased 1.6-fold in the AD hippocampus and 3.4-fold in the superior frontal gyrus (superior gyrus frontalis). Among them, HSPGs increased the most. | [217] |
| AD | Occipital neocortical and hippocampal tissue | Fibrillar Aβ plaques and nonfibrillar Aβ plaques contained high levels of N-sulfated HS, while N-sulfation was very low in nonfibrillar Aβ plaques. | [218] |
| AD | Cerebral cortex | The critical sites for binding of heparin sulfate to β-amyloid (Aβ) fibrils contain 2-O-S IdoA and N-S from the human cerebral cortex, whereas binding of Aβ monomers requires 6-O-S on GlcN residues. | [219] |
| Parkinson’s disease (PD) | Neuronal cells | The internalization of α-syn aggregates strongly depends on the cell surface HS and is associated with their total sulfation level. | [224] |
| COVID-19 infection | N-S, 2-O-S, and 6-O-S in HS and 6-O-S, 2-O-S, 3-O-S, and N-S in HP were critical for competitive binding to SARS-CoV-2 spike protein. | [241] | |
| COVID-19 infection | The presence of 3-O-S in HS contributes to the recognition and binding of SARS-CoV-2 spikes in vitro. | [242] | |
| COVID-19 infection | 3-O-sulfotransferase 3B overexpression and glycocalyx sulfation degree were too low to promote SARS-CoV-2 infection under pathological conditions. | [243] | |
| Idiopathic pulmonary fibrosis | Lung | CS/DS, HA, and the CS/DS ratio increased significantly. CS/DS increases in 4-O-S, 6-O-S, and 2-O-S disaccharide units and decreases in non-sulfated disaccharides, resulting in significant increases in sulfated levels. Similarly, significant increases were observed in NS, 2-O-S, and 6-O-S disaccharides of HS, particularly the UA2S–GlcNS6S unit. | [247] |
| Asthma | Endobronchial biopsy specimens | Significant increases in PG (versican, biglycan, and decorin) content at the lesion were observed in biopsy specimens from asthma cases. | [248] |
| Chronic obstructive pulmonary disease | Lung | HS increased significantly, and its sulfation pattern was related to the COPD stage, while CS/DS did not change significantly. The 2-O-S and NS of HS increased during the fourth phase of COPD, while 6-O-S did not change. | [249] |
| Cystic fibrosis | Lung | GAG expression is elevated in cystic fibrosis and abnormal sulfation of GAG can be found in bronchial epithelial cells of patients. | [250,251] |
| Familial urolithiasis | Urine | The total sulfated GAG, CS, and HS contents in urine all decreased, while the HA content was increased, and the proportion of HS in total sulfated GAGs was increased. | [259] |
| Kidney stones | Urine | Black South Africans were found to have higher CS levels in urine than whites (kidney stones were significantly more prevalent in whites than blacks in South Africans), suggesting that higher CS levels may inhibit kidney stone formation. | [260] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Q.; Chi, L. The Alterations and Roles of Glycosaminoglycans in Human Diseases. Polymers 2022, 14, 5014. https://doi.org/10.3390/polym14225014
Wang Q, Chi L. The Alterations and Roles of Glycosaminoglycans in Human Diseases. Polymers. 2022; 14(22):5014. https://doi.org/10.3390/polym14225014
Chicago/Turabian StyleWang, Qingchi, and Lianli Chi. 2022. "The Alterations and Roles of Glycosaminoglycans in Human Diseases" Polymers 14, no. 22: 5014. https://doi.org/10.3390/polym14225014