
The Effects of Time-Restricted Eating on Metabolism and Gut Microbiota: A Real-Life Study

 , , , , , ,
, , , , , , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Ethical Aspects
2.3. Design of the Study
2.4. Dietary Intervention
2.5. Monitoring
2.6. Measurements
2.7. DNA Extraction, Meta-Taxonomic Amplicon Sequencing, and Bioinformatic Analyses
2.8. Statistical Analyses
3. Results
3.1. Changes in Metabolic Variables
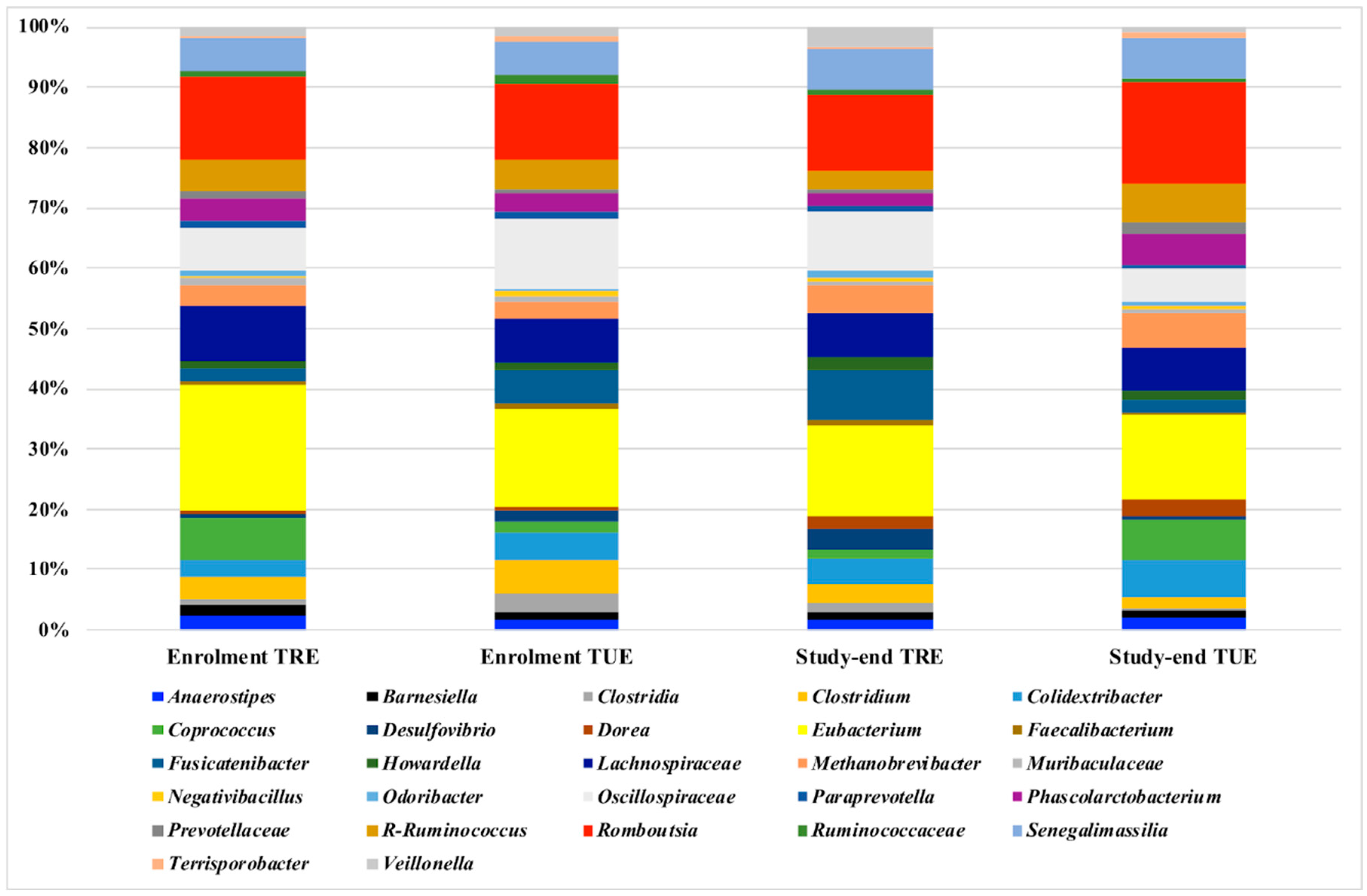
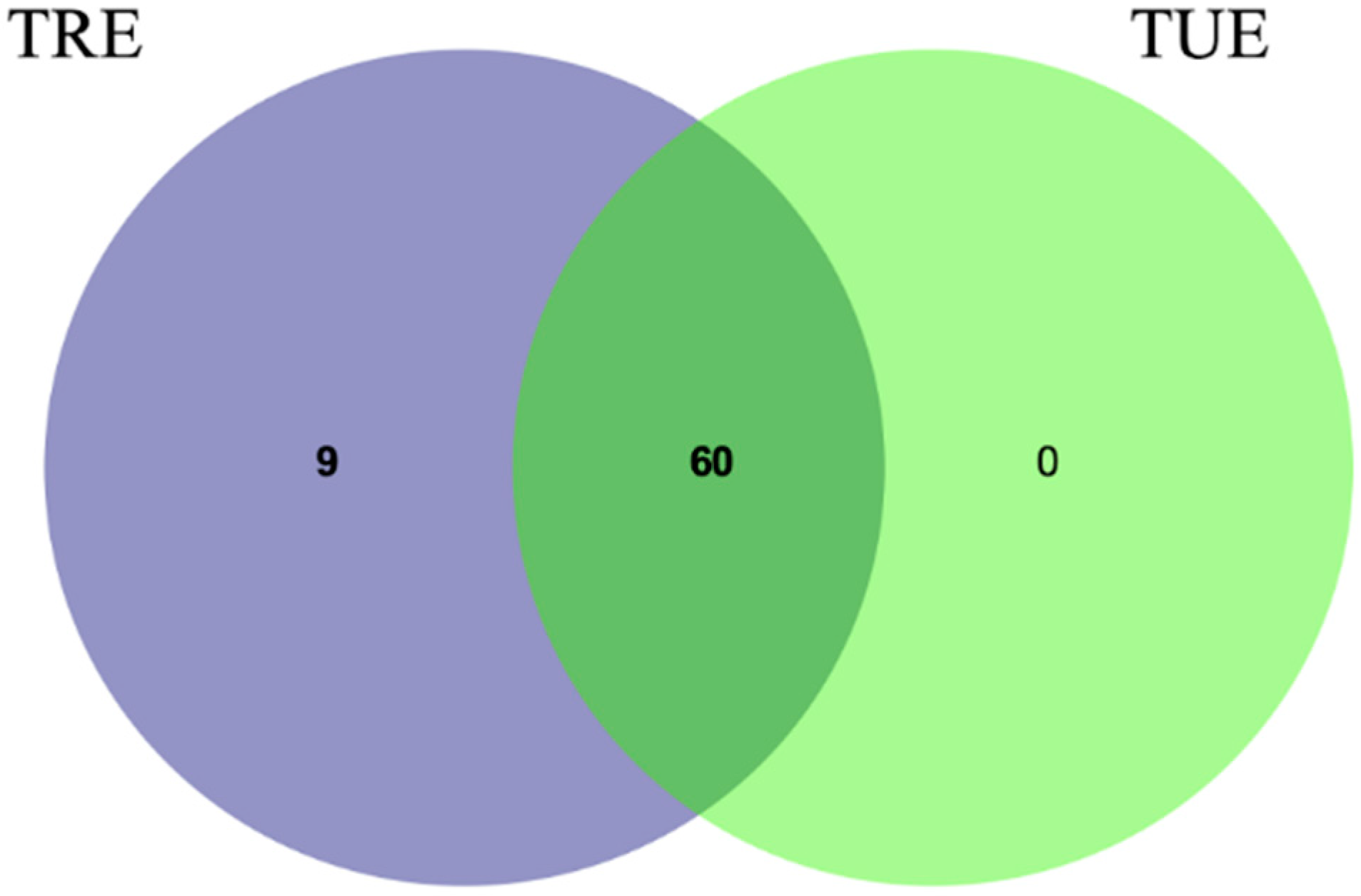
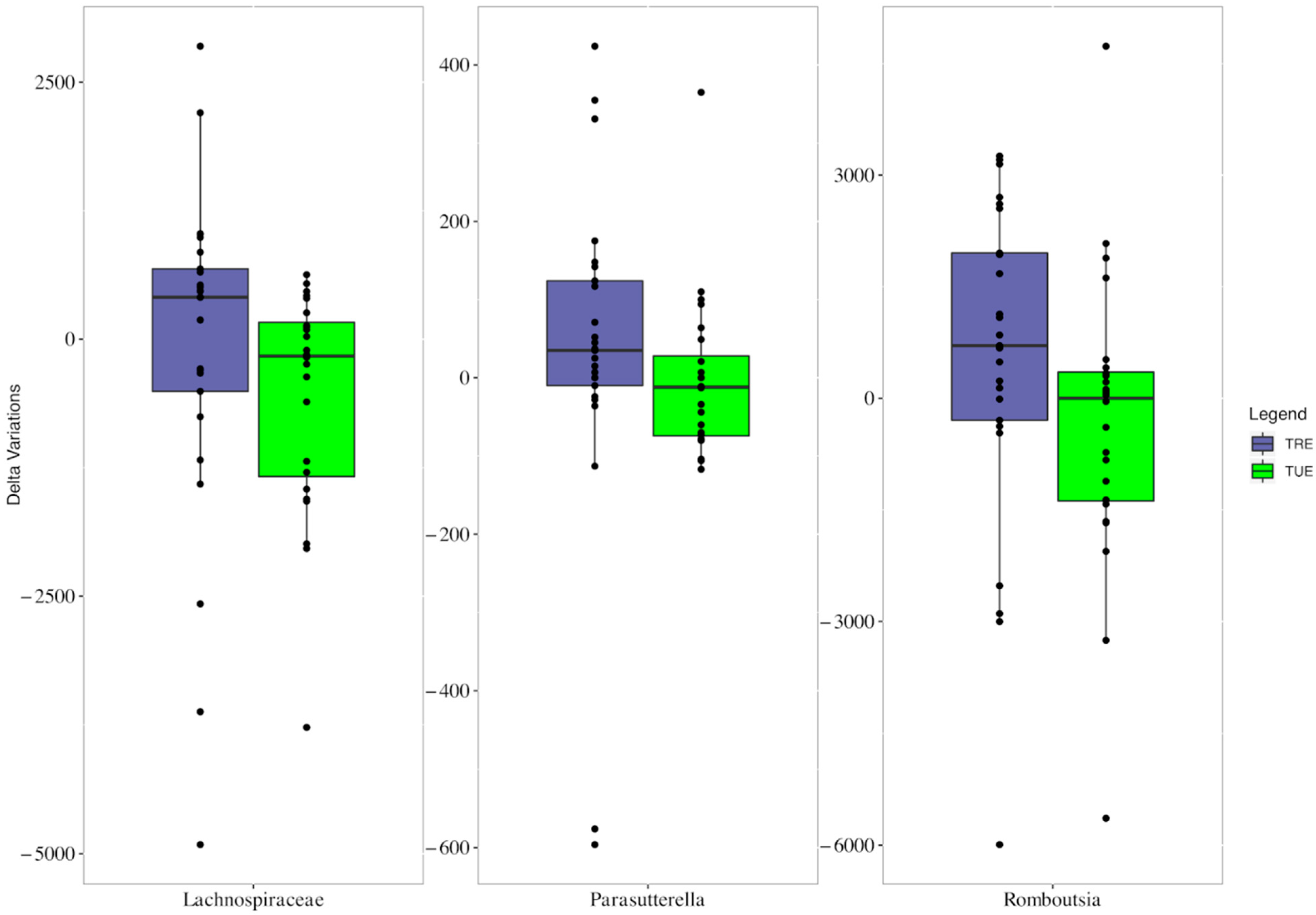
3.2. Microbiota Analysis
3.3. Correlations between Microbiota as well as Metabolic and Dietary Variables
4. Discussion
4.1. TRE and Metabolic Changes
4.2. TRE and Microbiota Changes
4.3. Limitations and Strengths
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chaix, A.; Manoogian, E.N.C.; Melkani, G.C.; Panda, S. Time-restricted eating to prevent and manage chronic metabolic diseases. Annu. Rev. Nutr. 2019, 39, 291–315. [Google Scholar] [CrossRef] [PubMed]
- Rynders, C.A.; Thomas, E.A.; Zaman, A.; Pan, Z.; Catenacci, V.A.; Melanson, E.L. Effectiveness of intermittent fasting and time-restricted feeding compared to continuous energy restriction for weight loss. Nutrients 2019, 11, 2442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pellegrini, M.; Cioffi, I.; Evangelista, A.; Ponzo, V.; Goitre, I.; Ciccone, G.; Ghigo, E.; Bo, S. Effects of time-restricted feeding on body weight and metabolism. a systematic review and meta-analysis. Rev. Endocr. Metab. Disord. 2019, 21, 17–33. [Google Scholar] [CrossRef] [PubMed]
- Moon, S.; Kang, J.; Kim, S.H.; Chung, H.S.; Kim, Y.J.; Yu, J.M.; Cho, S.T.; Oh, C.M.; Kim, T. Beneficial effects of time-restricted eating on metabolic diseases: A systemic review and meta-analysis. Nutrients 2020, 12, 1267. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira Maranhão Pureza, I.R.; de Lima Macena, M.; da Silva Junior, A.E.; Praxedes, D.R.S.; Vasconcelos, L.G.L.; Bueno, N.B. Effect of early time-restricted feeding on the metabolic profile of adults with excess weight: A systematic review with meta-analysis. Clin. Nutr. 2021, 40, 1788–1799. [Google Scholar] [CrossRef] [PubMed]
- Zeb, F.; Wu, X.; Chen, L.; Fatima, S.; Haq, I.; Chen, A.; Majeed, F.; Feng, Q.; Li, M. Effect of time-restricted feeding on metabolic risk and circadian rhythm associated with gut microbiome in healthy males. Br. J. Nutr. 2020, 123, 1216–1226. [Google Scholar] [CrossRef]
- Bo, S.; Musso, G.; Beccuti, G.; Fadda, M.; Fedele, D.; Gambino, R.; Gentile, L.; Durazzo, M.; Ghigo, E.; Cassader, M. Consuming more of daily caloric intake at dinner predisposes to obesity. A 6-year population-based prospective cohort study. PLoS ONE 2014, 9, e108467. [Google Scholar] [CrossRef] [PubMed]
- Bo, S.; Fadda, M.; Castiglione, A.; Ciccone, G.; De Francesco, A.; Fedele, D.; Guggino, A.; Parasiliti Caprino, M.; Ferrara, S.; Vezio Boggio, M.; et al. Is the timing of caloric intake associated with variation in diet-induced thermogenesis and in the metabolic pattern? A randomized cross-over study. Int. J. Obes. 2015, 39, 1689–1695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poggiogalle, E.; Jamshed, H.; Peterson, C.M. Circadian regulation of glucose, lipid, and energy metabolism in humans. Metab. Clin. Exp. 2018, 84, 11–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopez-Minguez, J.; Gómez-Abellán, P.; Garaulet, M. Timing of breakfast, lunch, and dinner. effects on obesity and metabolic risk. Nutrients 2019, 11, 2624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zarrinpar, A.; Chaix, A.; Yooseph, S.; Panda, S. Diet and feeding pattern affect the diurnal dynamics of the gut microbiome. Cell Metab. 2014, 20, 1006–1017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Özkul, C.; Yalınay, M.; Karakan, T. Islamic fasting leads to an increased abundance of akkermansia muciniphila and Bacteroides fragilis group: A preliminary study on intermittent fasting. Turk. J. Gastroenterol. 2019, 30, 1030–1035. [Google Scholar] [CrossRef] [PubMed]
- Zeb, F.; Wu, X.; Chen, L.; Fatima, S.; Ijaz-ul-Haq; Chen, A.; Xu, C.; Jianglei, R.; Feng, Q.; Li, M. Time-restricted feeding is associated with changes in human gut microbiota related to nutrient intake. Nutrition 2020, 78, 110797. [Google Scholar] [CrossRef] [PubMed]
- Xie, Z.; Sun, Y.; Ye, Y.; Hu, D.; Zhang, H.; He, Z.; Zhao, H.; Yang, H.; Mao, Y. Randomized controlled trial for time-restricted eating in healthy volunteers without obesity. Nat. Commun. 2022, 13, 1003. [Google Scholar] [CrossRef] [PubMed]
- Ozkul, C.; Yalinay, M.; Karakan, T. Structural changes in gut microbiome after Ramadan fasting: A pilot study. Benef. Microbes 2020, 11, 227–233. [Google Scholar] [CrossRef] [PubMed]
- Gabel, K.; Marcell, J.; Cares, K.; Kalam, F.; Cienfuegos, S.; Ezpeleta, M.; Varady, K.A. Effect of time restricted feeding on the gut microbiome in adults with obesity: A pilot study. Nutr. Health 2020, 26, 79–85. [Google Scholar] [CrossRef]
- Hu, D.; Ye, Y.; Mao, Y.; Liao, W.; Xu, W. Time-restricted feeding during childhood has persistent effects on mice commensal microbiota. Ann. Transl. Med. 2019, 7, 556. [Google Scholar] [CrossRef] [PubMed]
- Hu, D.; Mao, Y.; Xu, G.; Liao, W.; Ren, J.; Yang, H.; Yang, J.; Sun, L.; Chen, H.; Wang, W.; et al. Time-restricted feeding causes irreversible metabolic disorders and gut microbiota shift in pediatric mice. Pediatr. Res. 2019, 85, 518–526. [Google Scholar] [CrossRef]
- Ponzo, V.; Scumaci, E.; Goitre, I.; Beccuti, G.; Benso, A.; Belcastro, S.; Crespi, C.; De Michieli, F.; Pellegrini, M.; Scuntero, P.; et al. Predictors of attrition from a weight loss program. A study of adult patients with obesity in a community setting. Eat. Weight Disord. 2021, 26, 1729–1736. [Google Scholar] [CrossRef]
- Marzola, E.; Abbate-Daga, G.; Scumaci, E.; Ponzo, V.; Goitre, I.; Pellegrini, M.; D’eusebio, C.; Benso, A.; Belcastro, S.; De Michieli, F.; et al. In patients with obesity, are affective temperaments associated with attrition? An evaluation during and before the SARS-CoV-2 Pandemic. J. Clin. Med. 2022, 11, 862. [Google Scholar] [CrossRef] [PubMed]
- Bo, S.; Ponzo, V.; Ciccone, G.; Evangelista, A.; Saba, F.; Goitre, I.; Procopio, M.; Pagano, G.F.; Cassader, M.; Gambino, R. Six months of resveratrol supplementation has no measurable effect in type 2 diabetic patients. a randomized, double blind, placebo-controlled trial. Pharmacol. Res. 2016, 111, 896–905. [Google Scholar] [CrossRef]
- Ferrocino, I.; Ponzo, V.; Gambino, R.; Zarovska, A.; Leone, F.; Monzeglio, C.; Goitre, I.; Rosato, R.; Romano, A.; Grassi, G.; et al. Changes in the gut microbiota composition during pregnancy in patients with gestational diabetes mellitus (GDM). Sci. Rep. 2018, 8, 12216. [Google Scholar] [CrossRef]
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glöckner, F.O. Evaluation of General 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Res. 2013, 41, e1. [Google Scholar] [CrossRef]
- Callahan, B.J.; Mcmurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.; Holmes, S.P. DADA2: High resolution sample inference from amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [Green Version]
- Gabel, K.; Hoddy, K.K.; Haggerty, N.; Song, J.; Kroeger, C.M.; Trepanowski, J.F.; Panda, S.; Varady, K.A. Effects of 8-hour time restricted feeding on body weight and metabolic disease risk factors in obese adults: A pilot study. Nutr. Healthy Aging 2018, 4, 345–353. [Google Scholar] [CrossRef]
- Bo, S.; Ciccone, G.; Baldi, C.; Benini, L.; Dusio, F.; Forastiere, G.; Lucia, C.; Nuti, C.; Durazzo, M.; Cassader, M.; et al. Effectiveness of a lifestyle intervention on metabolic syndrome. A randomized controlled trial. J. Gen. Intern. Med. 2007, 22, 1695–1703. [Google Scholar] [CrossRef] [Green Version]
- Bo, S.; Rosato, R.; Ciccone, G.; Canil, S.; Gambino, R.; Poala, C.B.; Leone, F.; Valla, A.; Grassi, G.; Ghigo, E.; et al. Simple lifestyle recommendations and the outcomes of gestational diabetes. A 2 × 2 factorial randomized trial. Diabetes Obes. Metab. 2014, 16, 1032–1035. [Google Scholar] [CrossRef]
- Réda, A.; Wassil, M.; Mériem, M.; Alexia, P.; Abdelmalik, H.; Sabine, B.; Nassir, M. Food timing, circadian rhythm and chrononutrition: A systematic review of time-restricted eating’s effects on human health. Nutrients 2020, 12, 3770. [Google Scholar] [CrossRef]
- Liu, D.; Huang, Y.; Huang, C.; Yang, S.; Wei, X.; Zhang, P.; Guo, D.; Lin, J.; Xu, B.; Li, C.; et al. Calorie restriction with or without time-restricted eating in weight loss. N. Engl. J. Med. 2022, 386, 1495–1504. [Google Scholar] [CrossRef]
- Lowe, D.A.; Wu, N.; Rohdin-Bibby, L.; Holliston Moore, A.; Kelly, N.; En Liu, Y.; Philip, E.; Vittinghoff, E.; Heymsfield, S.B.; Olgin, J.E.; et al. Effects of time-restricted eating on weight loss and other metabolic parameters in women and men with overweight and obesity the TREAT randomized clinical trial. JAMA Intern. Med. 2020, 180, 1491–1499. [Google Scholar] [CrossRef]
- Carlson, O.; Martin, B.; Stote, K.S.; Golden, E.; Maudsley, S.; Najjar, S.S.; Ferrucci, L.; Ingram, D.K.; Longo, D.L.; Rumpler, W.V.; et al. Impact of reduced meal frequency without caloric restriction on glucose regulation in healthy, normal weight middle-aged men and women. Metabolism 2007, 56, 1729–1734. [Google Scholar] [CrossRef] [Green Version]
- Stote, K.S.; Baer, D.J.; Spears, K.; Paul, D.R.; Harris, G.K.; Rumpler, W.V.; Strycula, P.; Najjar, S.S.; Ferrucci, L.; Ingram, D.K.; et al. Controlled trial of reduced meal frequency without caloric restriction in healthy, normal-weight, middle-aged adults. Am. J. Clin. Nutr. 2007, 85, 981–988. [Google Scholar] [CrossRef]
- Lin, Y.J.; Wang, Y.T.; Chan, L.C.; Chu, N.F. Effect of time-restricted feeding on body composition and cardio-metabolic risk in middle-aged women in Taiwan. Nutrition 2022, 93, 111504. [Google Scholar] [CrossRef]
- Jamshed, H.; Beyl, R.A.; Manna, D.L.D.; Yang, E.S.; Ravussin, E.; Peterson, C.M. Early time-restricted feeding improves 24-hour glucose levels and affects markers of the circadian clock, aging, and autophagy in humans. Nutrients 2019, 11, 1234. [Google Scholar] [CrossRef] [Green Version]
- Manoogian, E.N.C.; Chow, L.S.; Taub, P.R.; Laferrère, B.; Panda, S. Time-restricted eating for the prevention and management of metabolic diseases. Endocr. Rev. 2022, 43, 405–436. [Google Scholar] [CrossRef]
- Jones, R.; Pabla, P.; Mallinson, J.; Nixon, A.; Taylor, T.; Bennett, A.; Tsintzas, K. Two weeks of early time-restricted feeding (ETRF) improves skeletal muscle insulin and anabolic sensitivity in healthy men. Am. J. Clin. Nutr. 2020, 112, 1015–1028. [Google Scholar] [CrossRef]
- Queiroz, J.N.; Macedo, R.C.O.; Tinsley, G.M.; Reischak-Oliveira, A. Time-restricted eating and circadian rhythms: The biological clock is ticking. Crit. Rev. Food Sci. Nutr. 2021, 61, 2863–2875. [Google Scholar] [CrossRef]
- Sutton, E.F.; Beyl, R.; Early, K.S.; Cefalu, W.T.; Ravussin, E.; Peterson, C.M. Early time-restricted feeding improves insulin sensitivity, blood pressure, and oxidative stress even without weight loss in men with prediabetes. Cell Metab. 2018, 27, 1212–1221. [Google Scholar] [CrossRef] [Green Version]
- Thomas, E.A.; Zaman, A.; Sloggett, K.J.; Steinke, S.; Grau, L.; Catenacci, V.A.; Cornier, M.-A.; Corey, M.A.; Rynders, A. Early time-restricted eating compared with daily caloric restriction: A randomized trial in adults with obesity. Obesity 2022, 30, 1027–1038. [Google Scholar] [CrossRef]
- Vyas, M.V.; Garg, A.X.; Iansavichus, A.V.; Costella, J.; Donner, A.; Laugsand, L.E.; Janszky, I.; Mrkobrada, M.; Parraga, G.; Hackam, D.G. Shift work and vascular events: Systematic review and meta-analysis. BMJ 2012, 345, e4800. [Google Scholar] [CrossRef] [Green Version]
- Dashti, H.S.; Gómez-Abellán, P.; Qian, J.; Esteban, A.; Morales, E.; Scheer, F.A.J.L.; Garaulet, M. Late eating is associated with cardiometabolic risk traits, obesogenic behaviors, and impaired weight loss. Am. J. Clin. Nutr. 2021, 113, 154–161. [Google Scholar] [CrossRef] [PubMed]
- Su, J.; Wang, Y.; Zhang, X.; Ma, M.; Xie, Z.; Pan, Q.; Ma, Z.; Peppelenbosch, M.P. Remodeling of the gut microbiome during ramadan-associated intermittent fasting. Am. J. Clin. Nutr. 2021, 113, 1332–1342. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Q.; Li, D.; He, Y.; Li, Y.; Yang, Z.; Zhao, X.; Liu, Y.; Wang, Y.; Sun, J.; Feng, X.; et al. Discrepant gut microbiota markers for the classification of obesity related metabolic abnormalities. Sci. Rep. 2019, 9, 13424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Testerman, T.; Li, Z.; Galuppo, B.; Graf, J.; Santoro, N. Insights from shotgun metagenomics into bacterial species and metabolic pathways associated with NAFLD in obese youth. Hepatol. Commun. 2022. [Google Scholar] [CrossRef]
- Henneke, L.; Schlicht, K.; Andreani, N.A.; Hollstein, T.; Demetrowitsch, T.; Knappe, C.; Hartmann, K.; Jensen-Kroll, J.; Rohmann, N.; Pohlschneider, D.; et al. Dietary carbohydrate–Gut Parasutterella–human fatty acid biosynthesis metabolic axis in obesity and type 2 diabetes. Gut Microbes 2022, 14, e2057778. [Google Scholar] [CrossRef]
- Maskarinec, G.; Raquinio, P.; Kristal, B.S.; Setiawan, V.W.; Wilkens, L.R.; Franke, A.A.; Lim, U.; Le Marchand, L.; Randolph, T.W.; Lampe, J.W.; et al. The Gut microbiome and type 2 diabetes status in the multiethnic cohort. PLoS ONE 2021, 16, e0250855. [Google Scholar] [CrossRef]
- Chávez-Carbajal, A.; Nirmalkar, K.; Pérez-Lizaur, A.; Hernández-Quiroz, F.; Ramírez-Del-Alto, S.; García-Mena, J.; Hernández-Guerrero, C. Gut microbiota and predicted metabolic pathways in a sample of mexican women affected by obesity and obesity plus metabolic syndrome. Int. J. Mol. Sci. 2019, 20, 438. [Google Scholar] [CrossRef] [Green Version]
- Dang, J.T.; Mocanu, V.; Park, H.; Laffin, M.; Hotte, N.; Karmali, S.; Birch, D.W.; Madsen, K.L. Roux-En-Y gastric bypass and sleeve gastrectomy induce substantial and persistent changes in microbial communities and metabolic pathways. Gut Microbes 2022, 14, e2050636. [Google Scholar] [CrossRef]
- Chen, Z.; Radjabzadeh, D.; Chen, L.; Kurilshikov, A.; Kavousi, M.; Ahmadizar, F.; Arfan Ikram, M.; Uitterlinden, A.G.; Zhernakova, A.; Fu, J.; et al. Association of insulin resistance and type 2 diabetes with gut microbial diversity a microbiome-wide analysis from population studies + supplemental content. JAMA Netw. Open 2021, 4, e2118811. [Google Scholar] [CrossRef]
- Biassoni, R.; Di Marco, E.; Squillario, M.; Barla, A.; Piccolo, G.; Ugolotti, E.; Gatti, C.; Minuto, N.; Patti, G.; Maghnie, M.; et al. Gut microbiota in T1DM-onset pediatric patients: Machine-learning algorithms to classify microorganisms as disease linked. J. Clin. Endocrinol. Metab. 2020, 105, e3114–e3126. [Google Scholar] [CrossRef]
- Companys, J.; Gosalbes, M.J.; Pla-Pagà, L.; Calderón-Pérez, L.; Llauradó, E.; Pedret, A.; Valls, R.M.; Jiménez-Hernández, N.; Sandoval-Ramirez, B.A.; Del Bas, M.J.; et al. Gut microbiota profile and its association with clinical variables and dietary intake in overweight/obese and lean subjects: A cross-sectional study. Nutrients 2021, 13, 2032. [Google Scholar] [CrossRef] [PubMed]
- Palomba, A.; Tanca, A.; Abbondio, M.; Sau, R.; Serra, M.; Marongiu, F.; Fraumene, C.; Pagnozzi, D.; Laconi, E.; Uzzau, S. Time-restricted feeding induces Lactobacillus and Akkermansia-specific functional changes in the rat fecal microbiota. NPJ Biofilms Microb. 2021, 7, 85. [Google Scholar] [CrossRef] [PubMed]
- Ye, Y.; Xu, H.; Xie, Z.; Wang, L.; Sun, Y.; Yang, H.; Hu, D.; Mao, Y. time-restricted feeding reduces the detrimental effects of a high-fat diet, possibly by modulating the circadian rhythm of hepatic lipid metabolism and gut microbiota. Front. Nutr. 2020, 7, 596285. [Google Scholar] [CrossRef] [PubMed]
- Pinto, F.C.S.; Silva, A.A.M.; Souza, S.L. Repercussions of intermittent fasting on the intestinal microbiota community and body composition: A systematic review. Nutr. Rev. 2022, 80, 613–628. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| All | TRE | TUE | p | |
|---|---|---|---|---|
| Number | 49 | 25 | 24 | |
| Age (years) | 56.9 ± 8.0 | 58.6 ± 7.9 | 55.1 ± 7.9 | 0.13 |
| Males (%) | 18.4 | 20.0 | 16.7 | 0.76 |
| Height (m) | 1.61 ± 0.08 | 1.60 ± 0.09 | 1.61 ± 0.08 | 0.72 |
| Weight (kg) | 91.3 ± 12.7 | 90.4 ± 11.8 | 92.2 ± 13.7 | 0.62 |
| BMI (kg/m2) | 35.3 ± 3.1 | 35.2 ± 3.0 | 35.4 ± 3.2 | 0.78 |
| Waist circumference (cm) | 114.9 ± 10.6 | 117.1 ± 9.5 | 112.7 ± 11.4 | 0.15 |
| Neck circumference (cm) | 38.6 ± 3.4 | 39.3 ± 3.2 | 37.9 ± 3.4 | 0.14 |
| Body-fat mass (kg) | 37.6 ± 8.3 | 37.1 ± 8.9 | 38.1 ± 7.7 | 0.69 |
| Body-lean mass (kg) | 53.2 ± 10.7 | 53.1 ± 12.4 | 53.3 ± 9.0 | 0.95 |
| Fat mass (%) | 41.3 ± 7.9 | 41.3 ± 9.7 | 41.2 ± 5.6 | 0.98 |
| Systolic blood pressure (mmHg) | 134.5 ± 13.4 | 135.4 ± 11.7 | 133.5 ± 15.2 | 0.63 |
| Diastolic blood pressure (mmHg) | 82.7 ± 7.6 | 82.8 ± 5.4 | 82.5 ± 9.4 | 0.89 |
| Calorimetric data | ||||
| RMR (kcal) | 1474.6 ± 269.0 | 1469.8 ± 265.4 | 1479.5 ± 278.3 | 0.90 |
| Respiratory quotient | 0.76 (0.06) | 0.76 (0.07) | 0.76 (0.08) | 0.36 * |
| Dietary intakes | ||||
| Total energy (kcal/day) | 2086.6 ± 399.7 | 2086.8 ± 429.7 | 2086.5 ± 375.3 | 0.99 |
| Carbohydrates (% total kcal) | 51.2 ± 6.2 | 49.9 ± 5.8 | 52.5 ± 6.4 | 0.14 |
| Proteins (% total kcal) | 14.0 ± 1.6 | 13.9 ± 1.8 | 14.1 ± 1.5 | 0.77 |
| Fats (% total kcal) | 31.7 ± 6.1 | 32.9 ± 5.1 | 30.5 ± 6.9 | 0.18 |
| Saturated fats (% total kcal) | 8.4 ± 1.7 | 8.7 ± 1.7 | 8.1 ± 1.6 | 0.16 |
| Monounsaturated fats (% total kcal) | 17.4 ± 3.9 | 18.1 ± 3.2 | 16.8 ± 4.5 | 0.24 |
| Polyunsaturated fats (% total kcal) | 4.7 ± 1.3 | 4.7 ± 1.3 | 4.6 ± 1.3 | 0.71 |
| Fiber (g/day) | 28.3 ± 5.9 | 27.3 ± 6.3 | 29.2 ± 5.4 | 0.27 |
| Laboratory variables | ||||
| Fasting glucose (mg/dL) | 102.5 ± 22.5 | 102.6 ± 18.2 | 102.5 ± 26.7 | 0.98 |
| Glycated hemoglobin (mmol/mol) | 40.4 ± 6.0 | 40.4 ± 4.3 | 40.5 ± 7.5 | 0.96 |
| Total cholesterol (mg/dL) | 204.7 ± 39.9 | 204.5 ± 39.5 | 204.9 ± 41.1 | 0.97 |
| HDL cholesterol (mg/dL) | 52.0 ± 12.2 | 51.2 ± 10.0 | 52.9 ± 14.4 | 0.62 |
| Triglycerides (mg/dL) | 133.3 ± 56.1 | 141.3 ± 54.3 | 125.0 ± 58.0 | 0.32 |
| Alanine aminotransferase (IU/L) | 27.0 (16.0) | 26.0 (30.0) | 28.5 (10.5) | 0.58 * |
| TRE | TUE | ||||
|---|---|---|---|---|---|
| After | Median Delta | After | Median Delta | p † | |
| Number | 25 | 24 | |||
| Weight (kg) | 86.0 ± 11.7 ** | −4.00 | 89.2 ± 13.6 ** | −2.20 | 0.049 |
| BMI (kg/m2) | 33.5 ± 3.3 ** | −1.60 | 34.1 ± 3.1 ** | −0.86 | 0.096 |
| Waist circumference (cm) | 110.2 ± 7.8 ** | −5.00 | 107.0 ± 8.8 ** | −4.50 | 0.670 |
| Neck circumference (cm) | 37.5 ± 2.9 ** | −1.50 | 36.5 ± 3.2 ** | −1.00 | 0.332 |
| Body fat mass (kg) | 33.6 ± 8.0 ** | −2.80 | 36.0 ± 7.7 * | −1.75 | 0.280 |
| Body lean mass (kg) | 52.2 ± 9.4 | −0.90 | 52.9 ± 8.8 | −0.30 | 0.490 |
| Fat mass (%) | 39.0 ± 7.5 * | −1.29 | 40.2 ± 5.3 | −0.61 | 0.447 |
| Systolic blood pressure (mmHg) | 126.8 ± 11.3 ** | −10.0 | 126.5 ± 18.1 ** | −2.05 | 0.430 |
| Diastolic blood pressure (mmHg) | 82.2 ± 6.9 | 0.00 | 80.8 ± 11.5 | 0.00 | 0.516 |
| Calorimetric data | |||||
| RMR (kcal) | 1493.8 ± 242.6 | +15.0 | 1480.2 ± 317.8 | +22.0 | 0.936 |
| Respiratory quotient | 0.77 (0.05) | 0.00 | 0.77 (0.08) | +0.02 | 0.306 |
| Dietary intakes | |||||
| Total energy (kcal/day) | 1521.5 ± 198.7 | −497.3 | 1564.6 ± 259.7 | −527.9 | 0.729 |
| Carbohydrates (% total kcal) | 50.9 ± 5.0 | +1.33 | 52.5 ± 5.6 | −0.54 | 0.631 |
| Proteins (% total kcal) | 15.5 ± 1.7 ** | +1.40 | 15.6 ± 1.9 ** | +1.59 | 0.984 |
| Fats (% total kcal) | 32.2 ± 4.5 | −1.08 | 30.3 ± 4.6 | −0.23 | 0.689 |
| Saturated fats (% total kcal) | 7.7 ± 1.5 ** | −0.50 | 7.1 ± 1.0 * | −0.30 | 0.317 |
| Monounsaturated fats (% total kcal) | 17.8 ± 2.6 | −0.29 | 16.8 ± 3.0 | −0.34 | 0.779 |
| Polyunsaturated fats (% total kcal) | 5.3 ± 1.3 * | +0.57 | 5.0 ± 1.1 | +0.26 | 0.575 |
| Fiber (g/day) | 29.1 ± 4.7 | +1.13 | 30.3 ± 4.1 | +0.93 | 0.984 |
| Laboratory variables | |||||
| Fasting glucose (mg/dL) | 85.0 ± 15.3 ** | −19.0 | 84.3 ± 30.0 ** | −10.5 | 0.764 |
| Glycated hemoglobin (mmol/mol) | 39.5 ± 4.6 | −1.00 | 39.2 ± 7.9 | −0.45 | 0.617 |
| Total cholesterol (mg/dL) | 191.7 ± 46.0 | 0.00 | 195.3 ± 43.7 | −3.50 | 0.412 |
| HDL cholesterol (mg/dL) | 53.8 ± 11.6 | +3.00 | 54.6 ± 12.3 | +1.00 | 0.225 |
| Triglycerides (mg/dL) | 120.4 ± 46.5 | −20.0 | 110.8 ± 51.8 | −7.50 | 0.589 |
| Alanine aminotransferase (IU/L) | 24.0 (11.0) $ | −2.00 | 22.5 (8.0) $ | −5.00 | 0.992 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ferrocino, I.; Pellegrini, M.; D’Eusebio, C.; Goitre, I.; Ponzo, V.; Fadda, M.; Rosato, R.; Mengozzi, G.; Beccuti, G.; Merlo, F.D.; et al. The Effects of Time-Restricted Eating on Metabolism and Gut Microbiota: A Real-Life Study. Nutrients 2022, 14, 2569. https://doi.org/10.3390/nu14132569
Ferrocino I, Pellegrini M, D’Eusebio C, Goitre I, Ponzo V, Fadda M, Rosato R, Mengozzi G, Beccuti G, Merlo FD, et al. The Effects of Time-Restricted Eating on Metabolism and Gut Microbiota: A Real-Life Study. Nutrients. 2022; 14(13):2569. https://doi.org/10.3390/nu14132569
Chicago/Turabian StyleFerrocino, Ilario, Marianna Pellegrini, Chiara D’Eusebio, Ilaria Goitre, Valentina Ponzo, Maurizio Fadda, Rosalba Rosato, Giulio Mengozzi, Guglielmo Beccuti, Fabio Dario Merlo, and et al. 2022. "The Effects of Time-Restricted Eating on Metabolism and Gut Microbiota: A Real-Life Study" Nutrients 14, no. 13: 2569. https://doi.org/10.3390/nu14132569