
Selenium Status in Paediatric Patients with Neurodevelopmental Diseases

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design
2.2. Trace Element Analysis
2.3. Quantification of Protein Biomarkers of Se and Cu Status
2.4. Statistical Analysis
3. Results
3.1. Patient Characteristics
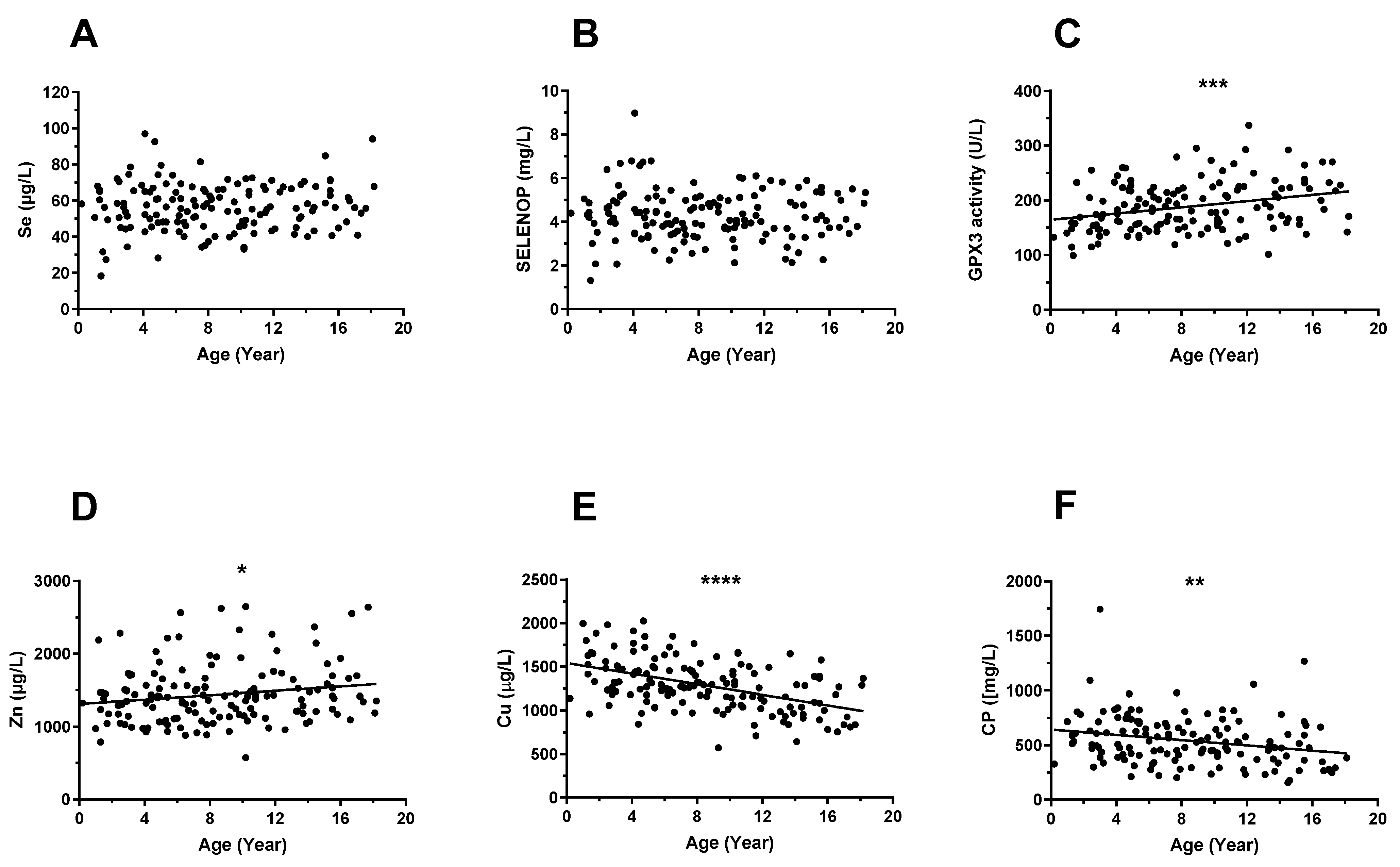
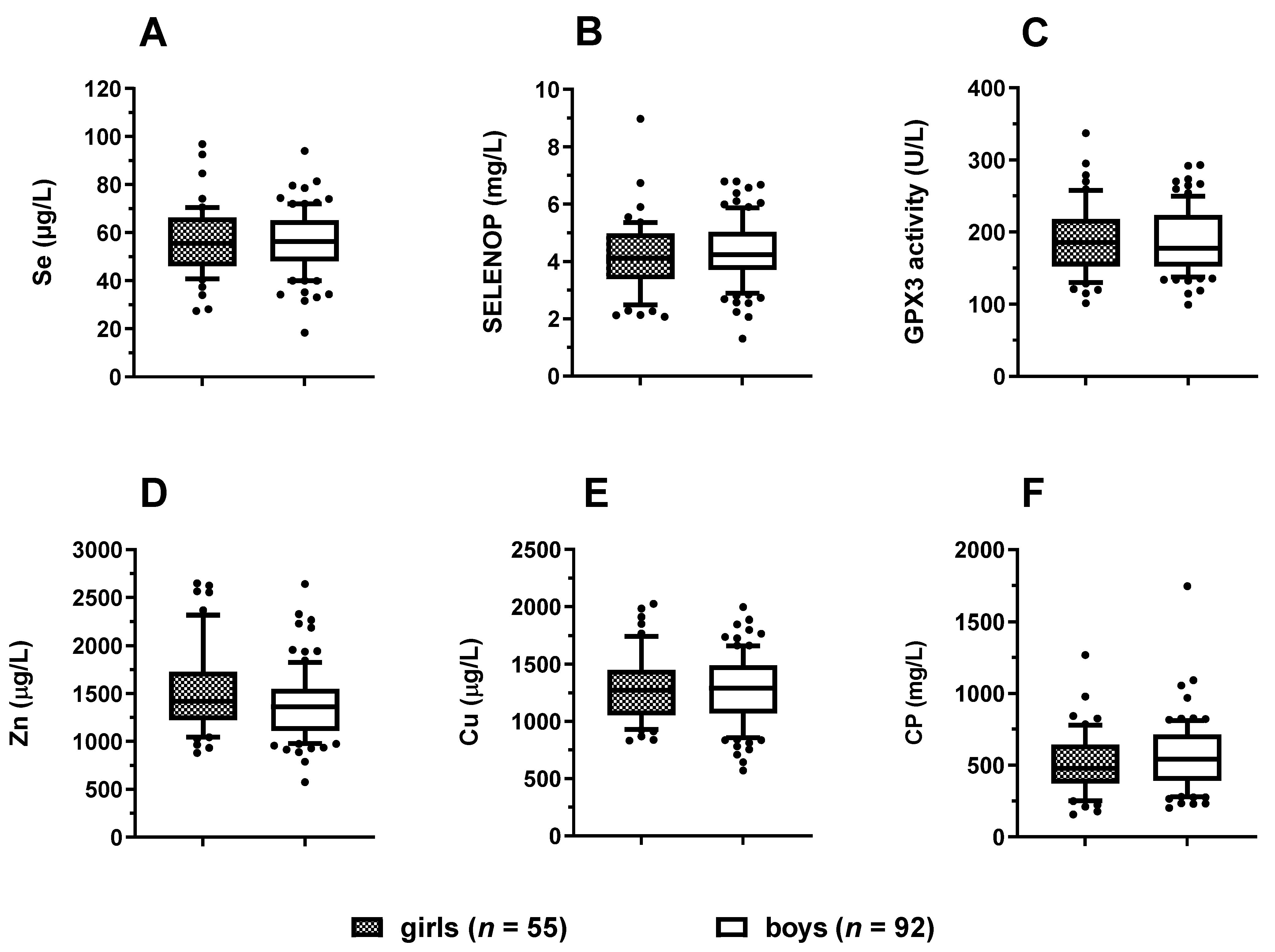
3.2. Trace Element Status in Relation to Age and Sex
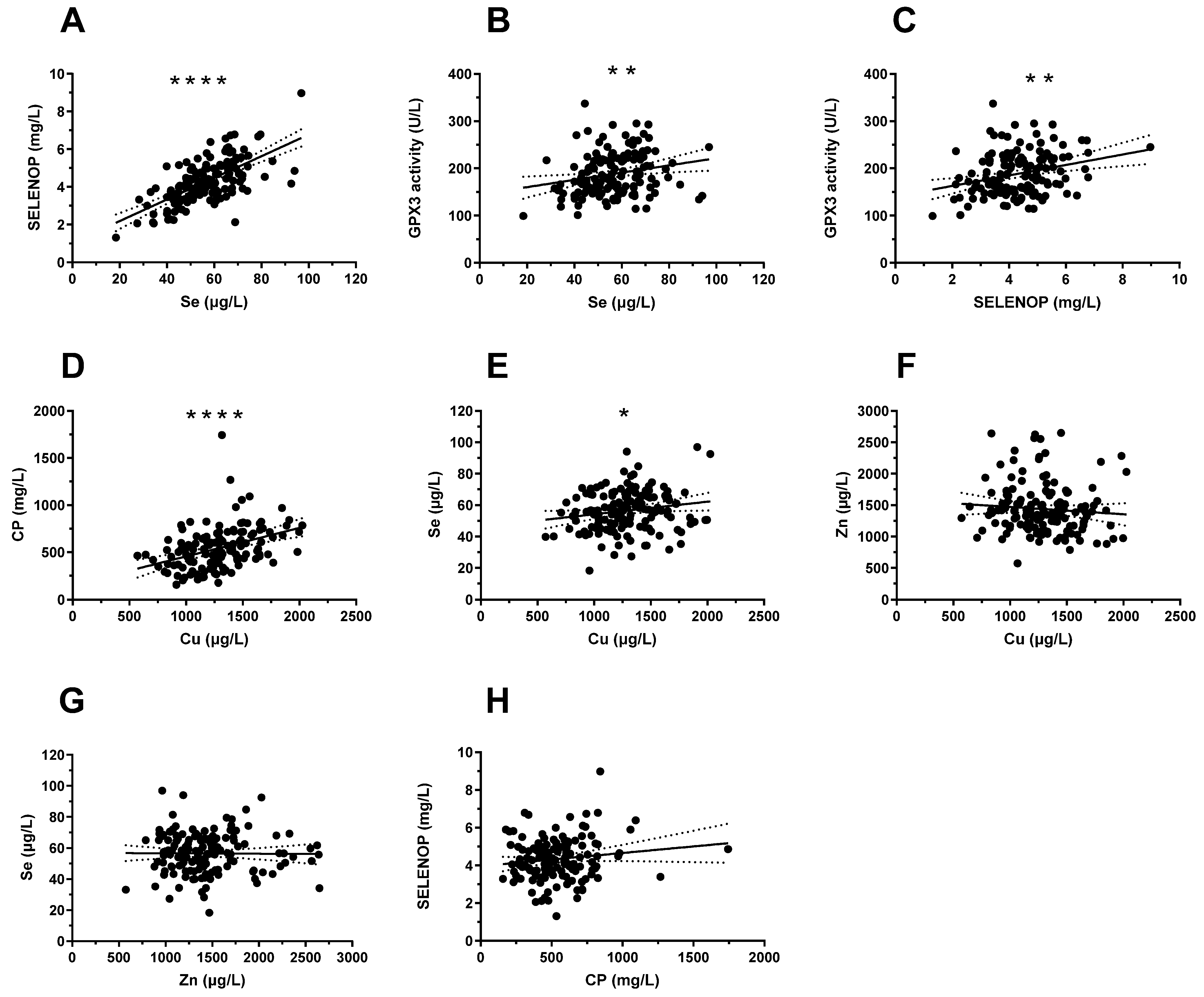
3.3. Correlations between Serum Cu, Zn, and Se Status and Protein Biomarkers
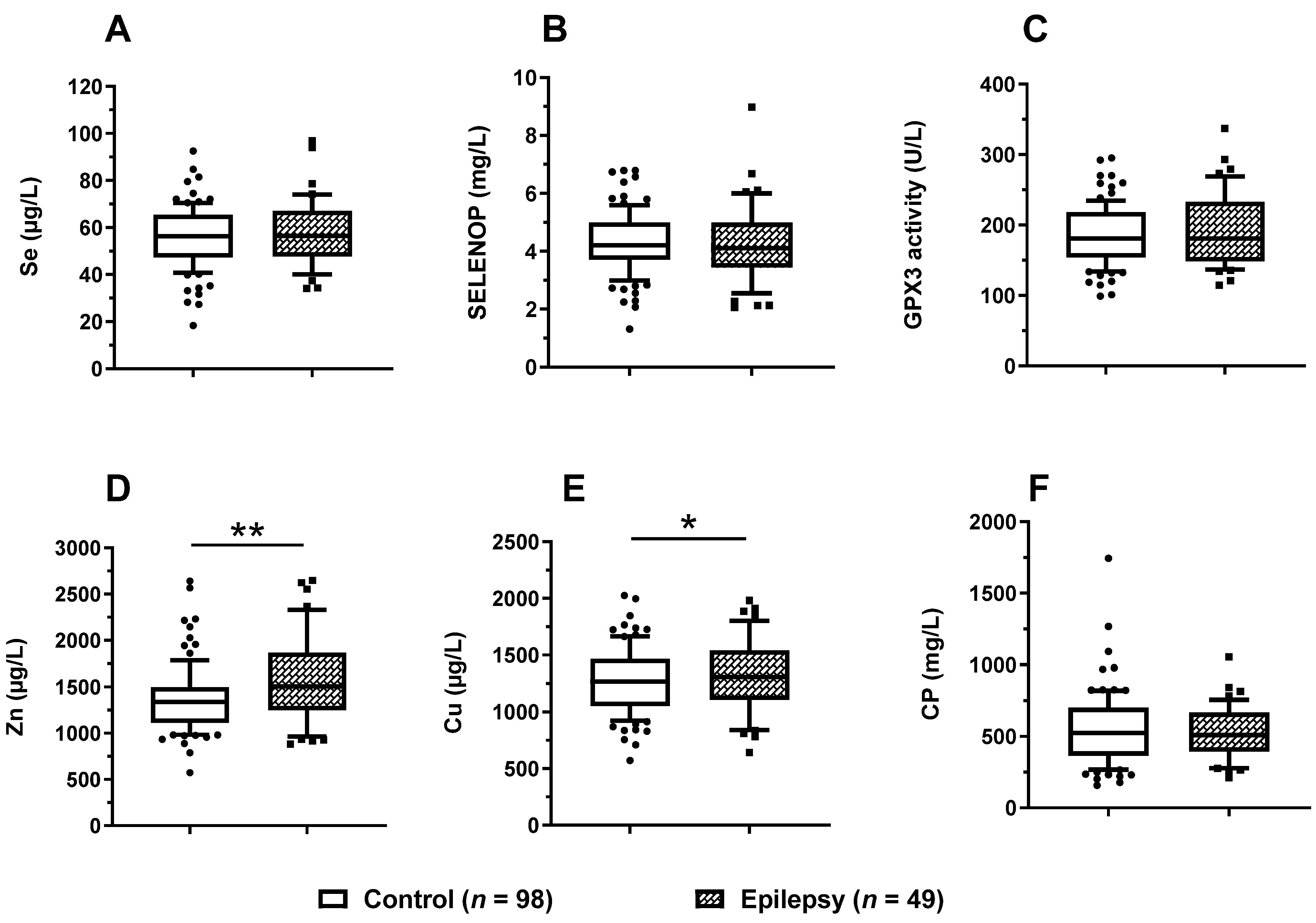
3.4. Trace Element Status in Relation to Epilepsy
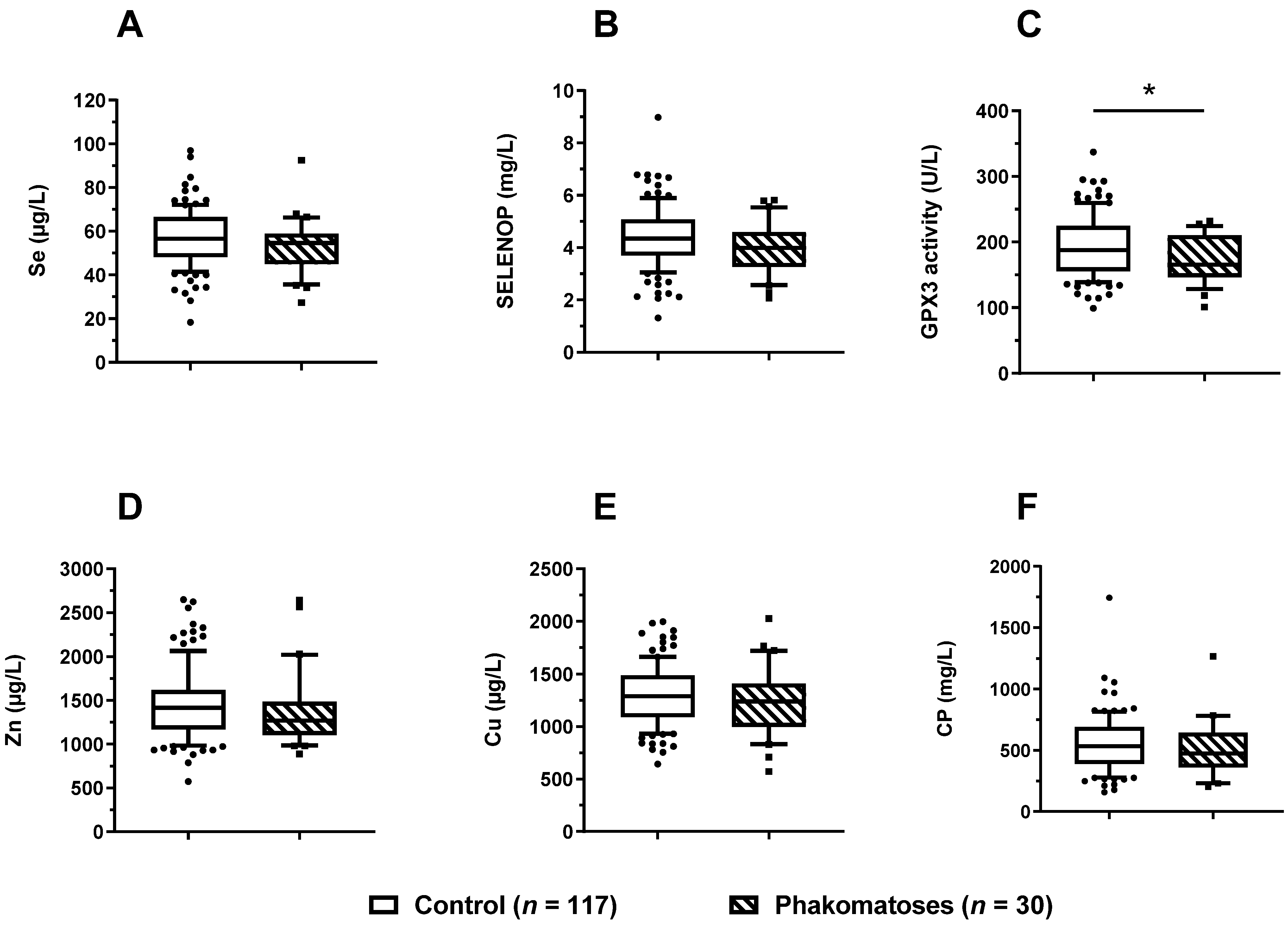
3.5. Trace Element Status in Relation to Phacomatoses
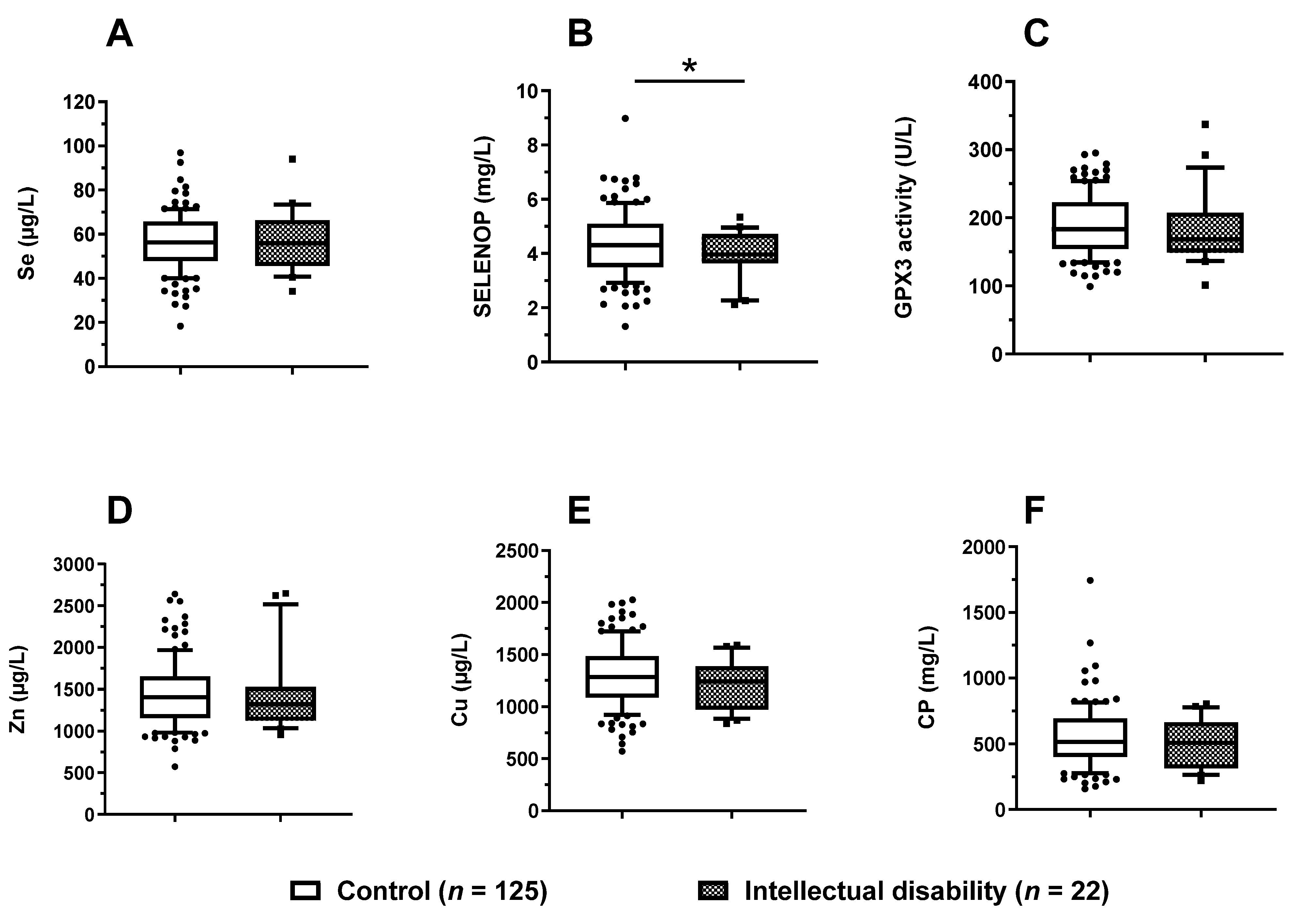
3.6. Trace Element Status in Relation to Mental Development
3.7. Trace Element Status of the Paediatric Patients in Relation to References
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Singh, M. Role of micronutrients for physical growth and mental development. Indian. J. Pediatr. 2004, 71, 59–62. [Google Scholar] [CrossRef] [PubMed]
- Umair, M.; Alfadhel, M. Genetic Disorders Associated with Metal Metabolism. Cells 2019, 8, 1598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zemrani, B.; Bines, J.E. Recent insights into trace element deficiencies: Causes, recognition and correction. Curr. Opin. Gastroen. 2020, 36, 110–117. [Google Scholar] [CrossRef] [PubMed]
- Scott, N.; Delport, D.; Hainsworth, S.; Pearson, R.; Morgan, C.; Huang, S.; Akuoku, J.K.; Piwoz, E.; Shekar, M.; Levin, C.; et al. Ending malnutrition in all its forms requires scaling up proven nutrition interventions and much more: A 129-country analysis. BMC Med. 2020, 18, 356. [Google Scholar] [CrossRef] [PubMed]
- Gassio, R.; Artuch, R.; Vilaseca, M.A.; Fuste, E.; Colome, R.; Campistol, J. Cognitive functions and the antioxidant system in phenylketonuric patients. Neuropsychology 2008, 22, 426–431. [Google Scholar] [CrossRef]
- Bryan, J.; Osendarp, S.; Hughes, D.; Calvaresi, E.; Baghurst, K.; van Klinken, J.W. Nutrients for cognitive development in school-aged children. Nutr. Rev. 2004, 62, 295–306. [Google Scholar] [CrossRef] [PubMed]
- Kasaikina, M.V.; Hatfield, D.L.; Gladyshev, V.N. Understanding selenoprotein function and regulation through the use of rodent models. Biochim. Biophys. Acta 2012, 1823, 1633–1642. [Google Scholar] [CrossRef] [Green Version]
- Schweizer, U.; Fradejas-Villar, N. Why 21? The significance of selenoproteins for human health revealed by inborn errors of metabolism. FASEB J. 2016, 30, 3669–3681. [Google Scholar] [CrossRef] [Green Version]
- Schoenmakers, E.; Chatterjee, K. Human Disorders Affecting the Selenocysteine Incorporation Pathway Cause Systemic Selenoprotein Deficiency. Antioxid. Redox Sign. 2020, 33, 481–497. [Google Scholar] [CrossRef]
- Schweizer, U.; Bohleber, S.; Zhao, W.; Fradejas-Villar, N. The Neurobiology of Selenium: Looking Back and to the Future. Front. Neurosci. 2021, 15, 652099. [Google Scholar] [CrossRef] [PubMed]
- Jones, G.D.; Droz, B.; Greve, P.; Gottschalk, P.; Poffet, D.; McGrath, S.P.; Seneviratne, S.I.; Smith, P.; Winkel, L.H. Selenium deficiency risk predicted to increase under future climate change. Proc. Natl. Acad. Sci. USA 2017, 114, 2848–2853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schiavon, M.; Nardi, S.; dalla Vecchia, F.; Ertani, A. Selenium biofortification in the 21(st)century: Status and challenges for healthy human nutrition. Plant Soil 2020, 453, 245–270. [Google Scholar] [CrossRef] [PubMed]
- Mirlean, N.; Seus-Arrache, E.R.; Vlasova, O. Selenium deficiency in subtropical littoral pampas: Environmental and dietary aspects. Environ. Geochem. Health 2018, 40, 543–556. [Google Scholar] [CrossRef] [PubMed]
- Ligowe, I.S.; Phiri, F.P.; Ander, E.L.; Bailey, E.H.; Chilimba, A.D.C.; Gashu, D.; Joy, E.J.M.; Lark, R.M.; Kabambe, V.; Kalimbira, A.A.; et al. Selenium deficiency risks in sub-Saharan African food systems and their geospatial linkages. Proc. Nutr. Soc. 2020, 79, 457–467. [Google Scholar] [CrossRef] [Green Version]
- Loscalzo, J. Keshan disease, selenium deficiency, and the selenoproteome. N. Engl. J. Med. 2014, 370, 1756–1760. [Google Scholar] [CrossRef]
- Duntas, L.H.; Benvenga, S. Selenium: An element for life. Endocrine 2015, 48, 756–775. [Google Scholar] [CrossRef]
- Thomson, C.D. Selenium and iodine intakes and status in New Zealand and Australia. Br. J. Nutr. 2004, 91, 661–672. [Google Scholar] [CrossRef] [Green Version]
- Schomburg, L. The other view: The trace element selenium as a micronutrient in thyroid disease, diabetes, and beyond. Hormones 2020, 19, 15–24. [Google Scholar] [CrossRef]
- Belhadj, M.; Tani, L.S.K.; Medjati, N.D.; Harek, Y.; Sahi, M.D.; Sun, Q.; Heller, R.; Behar, A.; Charlet, L.; Schomburg, L. Se Status Prediction by Food Intake as Compared to Circulating Biomarkers in a West Algerian Population. Nutrients 2020, 12, 3599. [Google Scholar] [CrossRef]
- Zhou, H.; Wang, T.; Li, Q.; Li, D. Prevention of Keshan Disease by Selenium Supplementation: A Systematic Review and Meta-analysis. Biol. Trace Elem. Res. 2018, 186, 98–105. [Google Scholar] [CrossRef]
- Yu, F.F.; Han, J.; Wang, X.; Fang, H.; Liu, H.; Guo, X. Salt-Rich Selenium for Prevention and Control Children with Kashin-Beck Disease: A Meta-analysis of Community-Based Trial. Biol. Trace Elem. Res. 2016, 170, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Duntas, L.H. Selenium and at-risk pregnancy: Challenges and controversies. Thyroid. Res. 2020, 13, 16. [Google Scholar] [CrossRef] [PubMed]
- Polanska, K.; Hanke, W.; Krol, A.; Gromadzinska, J.; Kuras, R.; Janasik, B.; Wasowicz, W.; Mirabella, F.; Chiarotti, F.; Calamandrei, G. Micronutrients during pregnancy and child psychomotor development: Opposite effects of Zinc and Selenium. Environ. Res. 2017, 158, 583–589. [Google Scholar] [CrossRef] [Green Version]
- Zachara, B.A. Selenium in Complicated Pregnancy. A Review. Adv. Clin. Chem. 2018, 86, 157–178. [Google Scholar] [CrossRef] [PubMed]
- Riese, C.; Michaelis, M.; Mentrup, B.; Gotz, F.; Kohrle, J.; Schweizer, U.; Schomburg, L. Selenium-dependent pre- and posttranscriptional mechanisms are responsible for sexual dimorphic expression of selenoproteins in murine tissues. Endocrinology 2006, 147, 5883–5892. [Google Scholar] [CrossRef] [Green Version]
- Hofstee, P.; Cuffe, J.S.M.; Perkins, A.V. Analysis of Selenoprotein Expression in Response to Dietary Selenium Deficiency During Pregnancy Indicates Tissue Specific Differential Expression in Mothers and Sex Specific Changes in the Fetus and Offspring. Int. J. Mol. Sci. 2020, 21, 2210. [Google Scholar] [CrossRef] [Green Version]
- Ajmone-Cat, M.A.; De Simone, R.; Tartaglione, A.M.; Di Biase, A.; Di Benedetto, R.; D’Archivio, M.; Vari, R.; Ricceri, L.; Aureli, F.; Iacoponi, F.; et al. Critical Role of Maternal Selenium Nutrition in Neurodevelopment: Effects on Offspring Behavior and Neuroinflammatory Profile. Nutrients 2022, 14, 1850. [Google Scholar] [CrossRef]
- Schomburg, L.; Riese, C.; Renko, K.; Schweizer, U. Effect of age on sexually dimorphic selenoprotein expression in mice. Biol. Chem. 2007, 388, 1035–1041. [Google Scholar] [CrossRef]
- Dumitrescu, A.M.; Liao, X.H.; Abdullah, M.S.; Lado-Abeal, J.; Majed, F.A.; Moeller, L.C.; Boran, G.; Schomburg, L.; Weiss, R.E.; Refetoff, S. Mutations in SECISBP2 result in abnormal thyroid hormone metabolism. Nat. Genet. 2005, 37, 1247–1252. [Google Scholar] [CrossRef]
- Azevedo, M.F.; Barra, G.B.; Naves, L.A.; Ribeiro Velasco, L.F.; Godoy Garcia Castro, P.; de Castro, L.C.; Amato, A.A.; Miniard, A.; Driscoll, D.; Schomburg, L.; et al. Selenoprotein-related disease in a young girl caused by nonsense mutations in the SBP2 gene. J. Clin. Endocrinol. Metab. 2010, 95, 4066–4071. [Google Scholar] [CrossRef] [Green Version]
- Di Cosmo, C.; McLellan, N.; Liao, X.H.; Khanna, K.K.; Weiss, R.E.; Papp, L.; Refetoff, S. Clinical and molecular characterization of a novel selenocysteine insertion sequence-binding protein 2 (SBP2) gene mutation (R128X). J. Clin. Endocrinol. Metab. 2009, 94, 4003–4009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schoenmakers, E.; Agostini, M.; Mitchell, C.; Schoenmakers, N.; Papp, L.; Rajanayagam, O.; Padidela, R.; Ceron-Gutierrez, L.; Doffinger, R.; Prevosto, C.; et al. Mutations in the selenocysteine insertion sequence-binding protein 2 gene lead to a multisystem selenoprotein deficiency disorder in humans. J. Clin. Investig. 2010, 120, 4220–4235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seeher, S.; Atassi, T.; Mahdi, Y.; Carlson, B.A.; Braun, D.; Wirth, E.K.; Klein, M.O.; Reix, N.; Miniard, A.C.; Schomburg, L.; et al. Secisbp2 is essential for embryonic development and enhances selenoprotein expression. Antioxid. Redox. Signal 2014, 21, 835–849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agamy, O.; Ben Zeev, B.; Lev, D.; Marcus, B.; Fine, D.; Su, D.; Narkis, G.; Ofir, R.; Hoffmann, C.; Leshinsky-Silver, E.; et al. Mutations disrupting selenocysteine formation cause progressive cerebello-cerebral atrophy. Am. J. Hum. Genet. 2010, 87, 538–544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Puppala, A.K.; French, R.L.; Matthies, D.; Baxa, U.; Subramaniam, S.; Simonovic, M. Structural basis for early-onset neurological disorders caused by mutations in human selenocysteine synthase. Sci. Rep. 2016, 6, 32563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iwama, K.; Sasaki, M.; Hirabayashi, S.; Ohba, C.; Iwabuchi, E.; Miyatake, S.; Nakashima, M.; Miyake, N.; Ito, S.; Saitsu, H.; et al. Milder progressive cerebellar atrophy caused by biallelic SEPSECS mutations. J. Hum. Genet. 2016, 61, 527–531. [Google Scholar] [CrossRef]
- Smith, A.C.; Mears, A.J.; Bunker, R.; Ahmed, A.; MacKenzie, M.; Schwartzentruber, J.A.; Beaulieu, C.L.; Ferretti, E.; Consortium, F.C.; Majewski, J.; et al. Mutations in the enzyme glutathione peroxidase 4 cause Sedaghatian-type spondylometaphyseal dysplasia. J. Med. Genet. 2014, 51, 470–474. [Google Scholar] [CrossRef] [Green Version]
- Wirth, E.K.; Conrad, M.; Winterer, J.; Wozny, C.; Carlson, B.A.; Roth, S.; Schmitz, D.; Bornkamm, G.W.; Coppola, V.; Tessarollo, L.; et al. Neuronal selenoprotein expression is required for interneuron development and prevents seizures and neurodegeneration. FASEB J. 2010, 24, 844–852. [Google Scholar] [CrossRef] [Green Version]
- Seeher, S.; Carlson, B.A.; Miniard, A.C.; Wirth, E.K.; Mahdi, Y.; Hatfield, D.L.; Driscoll, D.M.; Schweizer, U. Impaired selenoprotein expression in brain triggers striatal neuronal loss leading to co-ordination defects in mice. Biochem. J. 2014, 462, 67–75. [Google Scholar] [CrossRef] [Green Version]
- Hill, K.E.; Zhou, J.; McMahan, W.J.; Motley, A.K.; Atkins, J.F.; Gesteland, R.F.; Burk, R.F. Deletion of selenoprotein P alters distribution of selenium in the mouse. J. Biol. Chem. 2003, 278, 13640–13646. [Google Scholar] [CrossRef] [Green Version]
- Schomburg, L.; Schweizer, U.; Holtmann, B.; Flohe, L.; Sendtner, M.; Kohrle, J. Gene disruption discloses role of selenoprotein P in selenium delivery to target tissues. Biochem. J. 2003, 370, 397–402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Renko, K.; Werner, M.; Renner-Muller, I.; Cooper, T.G.; Yeung, C.H.; Hollenbach, B.; Scharpf, M.; Kohrle, J.; Schomburg, L.; Schweizer, U. Hepatic selenoprotein P (SePP) expression restores selenium transport and prevents infertility and motor-incoordination in Sepp-knockout mice. Biochem. J. 2008, 409, 741–749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burk, R.F.; Hill, K.E.; Motley, A.K.; Winfrey, V.P.; Kurokawa, S.; Mitchell, S.L.; Zhang, W. Selenoprotein P and apolipoprotein E receptor-2 interact at the blood-brain barrier and also within the brain to maintain an essential selenium pool that protects against neurodegeneration. FASEB J. 2014, 28, 3579–3588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schomburg, L.; Schweizer, U.; Kohrle, J. Selenium and selenoproteins in mammals: Extraordinary, essential, enigmatic. Cell Mol. Life Sci. 2004, 61, 1988–1995. [Google Scholar] [CrossRef]
- Ramaekers, V.T.; Calomme, M.; Vanden Berghe, D.; Makropoulos, W. Selenium deficiency triggering intractable seizures. Neuropediatrics 1994, 25, 217–223. [Google Scholar] [CrossRef]
- Hughes, D.J.; Fedirko, V.; Jenab, M.; Schomburg, L.; Meplan, C.; Freisling, H.; Bueno-de-Mesquita, H.B.; Hybsier, S.; Becker, N.P.; Czuban, M.; et al. Selenium status is associated with colorectal cancer risk in the European prospective investigation of cancer and nutrition cohort. Int. J. Cancer 2015, 136, 1149–1161. [Google Scholar] [CrossRef]
- Hybsier, S.; Schulz, T.; Wu, Z.; Demuth, I.; Minich, W.B.; Renko, K.; Rijntjes, E.; Kohrle, J.; Strasburger, C.J.; Steinhagen-Thiessen, E.; et al. Sex-specific and inter-individual differences in biomarkers of selenium status identified by a calibrated ELISA for selenoprotein P. Redox Biol. 2017, 11, 403–414. [Google Scholar] [CrossRef]
- Hackler, J.; Wisniewska, M.; Greifenstein-Wiehe, L.; Minich, W.B.; Cremer, M.; Buhrer, C.; Schomburg, L. Copper and selenium status as biomarkers of neonatal infections. J. Trace Elem. Med. Biol. 2020, 58, 126437. [Google Scholar] [CrossRef]
- Gunzler, W.A.; Kremers, H.; Flohe, L. An improved coupled test procedure for glutathione peroxidase (EC 1-11-1-9-) in blood. Z. Klin. Chem. Klin. Biochem. 1974, 12, 444–448. [Google Scholar]
- Heller, R.A.; Sun, Q.; Hackler, J.; Seelig, J.; Seibert, L.; Cherkezov, A.; Minich, W.B.; Seemann, P.; Diegmann, J.; Pilz, M.; et al. Prediction of survival odds in COVID-19 by zinc, age and selenoprotein P as composite biomarker. Redox Biol. 2021, 38, 101764. [Google Scholar] [CrossRef]
- Rukgauer, M.; Klein, J.; Kruse-Jarres, J.D. Reference values for the trace elements copper, manganese, selenium, and zinc in the serum/plasma of children, adolescents, and adults. J. Trace Elem. Med. Biol. 1997, 11, 92–98. [Google Scholar] [CrossRef]
- Rayman, M.P.; Rayman, M.P. The argument for increasing selenium intake. Proc. Nutr. Soc. 2002, 61, 203–215. [Google Scholar] [CrossRef] [PubMed]
- Moghaddam, A.; Heller, R.A.; Sun, Q.; Seelig, J.; Cherkezov, A.; Seibert, L.; Hackler, J.; Seemann, P.; Diegmann, J.; Pilz, M.; et al. Selenium Deficiency Is Associated with Mortality Risk from COVID-19. Nutrients 2020, 12, 2098. [Google Scholar] [CrossRef] [PubMed]
- El-Rashidy, O.F.; Youssef, M.M.; Elgendy, Y.G.; Mohsen, M.A.; Morsy, S.M.; Dawh, S.A.; Saad, K. Selenium and antioxidant levels in children with intractable epilepsy receiving ketogenic diet. Acta Neurol. Belg. 2020, 120, 375–380. [Google Scholar] [CrossRef]
- Sun, Q.; Mehl, S.; Renko, K.; Seemann, P.; Gorlich, C.L.; Hackler, J.; Minich, W.B.; Kahaly, G.J.; Schomburg, L. Natural Autoimmunity to Selenoprotein P Impairs Selenium Transport in Hashimoto’s Thyroiditis. Int. J. Mol. Sci. 2021, 22, 13088. [Google Scholar] [CrossRef]
- Amoros, R.; Murcia, M.; Gonzalez, L.; Rebagliato, M.; Iniguez, C.; Lopez-Espinosa, M.J.; Vioque, J.; Broberg, K.; Ballester, F.; Llop, S. Maternal selenium status and neuropsychological development in Spanish preschool children. Environ. Res. 2018, 166, 215–222. [Google Scholar] [CrossRef] [Green Version]
- Becker, B.; Strowd, R.E., 3rd. Phakomatoses. Dermatol. Clin. 2019, 37, 583–606. [Google Scholar] [CrossRef]
- Korf, B.R.; Bebin, E.M. Neurocutaneous Disorders in Children. Pediatr. Rev. 2017, 38, 119–128. [Google Scholar] [CrossRef] [PubMed]
- Jentarra, G.; Snyder, S.L.; Narayanan, V. Genetic aspects of neurocutaneous disorders. Semin. Pediatr. Neurol. 2006, 13, 43–47. [Google Scholar] [CrossRef]
- Seiderer, J.; Dambacher, J.; Kuhnlein, B.; Pfennig, S.; Konrad, A.; Torok, H.P.; Haller, D.; Goke, B.; Ochsenkuhn, T.; Lohse, P.; et al. The role of the selenoprotein S (SELS) gene -105G>A promoter polymorphism in inflammatory bowel disease and regulation of SELS gene expression in intestinal inflammation. Tissue. Antig. 2007, 70, 238–246. [Google Scholar] [CrossRef]
- Renko, K.; Hofmann, P.J.; Stoedter, M.; Hollenbach, B.; Behrends, T.; Kohrle, J.; Schweizer, U.; Schomburg, L. Down-regulation of the hepatic selenoprotein biosynthesis machinery impairs selenium metabolism during the acute phase response in mice. FASEB J 2009, 23, 1758–1765. [Google Scholar] [CrossRef] [PubMed]
- Korf, B.R. The phakomatoses. Clin. Dermatol. 2005, 23, 78–84. [Google Scholar] [CrossRef] [PubMed]
- Ohh, M. Ubiquitin pathway in VHL cancer syndrome. Neoplasia 2006, 8, 623–629. [Google Scholar] [CrossRef] [Green Version]
- Becker, N.P.; Martitz, J.; Renko, K.; Stoedter, M.; Hybsier, S.; Cramer, T.; Schomburg, L. Hypoxia reduces and redirects selenoprotein biosynthesis. Metallomics 2014, 6, 1079–1086. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhai, R.; Zhang, M.; Liu, J.; Guang, H.; Li, B.; Chen, D.; Zhang, S. Reference Intervals of and Relationships among Essential Trace Elements in Whole Blood of Children Aged 0–14 years. J. Clin. Lab. Anal. 2017, 31, 22043. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, Y.; Yan, C. Gender differences in trace element exposures with cognitive abilities of school-aged children: A cohort study in Wujiang city, China. Environ. Sci. Pollut. Res. Int. 2022, 1–15. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ICD-10 Code | Disease/Syndrome | Number of Children (%) |
|---|---|---|
| G40 | Epilepsy | 49 (33.3) |
| Q85 | Phacomatoses | 30 (20.4) |
| F70–F79 | Intellectual disability | 22 (15.0) |
| diverse | Diverse phenotypes | 46 (31.3) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Görlich, C.L.; Sun, Q.; Roggenkamp, V.; Hackler, J.; Mehl, S.; Minich, W.B.; Kaindl, A.M.; Schomburg, L. Selenium Status in Paediatric Patients with Neurodevelopmental Diseases. Nutrients 2022, 14, 2375. https://doi.org/10.3390/nu14122375
Görlich CL, Sun Q, Roggenkamp V, Hackler J, Mehl S, Minich WB, Kaindl AM, Schomburg L. Selenium Status in Paediatric Patients with Neurodevelopmental Diseases. Nutrients. 2022; 14(12):2375. https://doi.org/10.3390/nu14122375
Chicago/Turabian StyleGörlich, Christian L., Qian Sun, Viola Roggenkamp, Julian Hackler, Sebastian Mehl, Waldemar B. Minich, Angela M. Kaindl, and Lutz Schomburg. 2022. "Selenium Status in Paediatric Patients with Neurodevelopmental Diseases" Nutrients 14, no. 12: 2375. https://doi.org/10.3390/nu14122375