
Epidemiological Dynamics of Foot-and-Mouth Disease in the Horn of Africa: The Role of Virus Diversity and Animal Movement
 , , ,
, , ,
Abstract
:1. Search Strategy and Selection Criteria
2. Introduction
3. The Causes of FMD Virus Diversity
4. Distribution of Foot-and-Mouth Disease
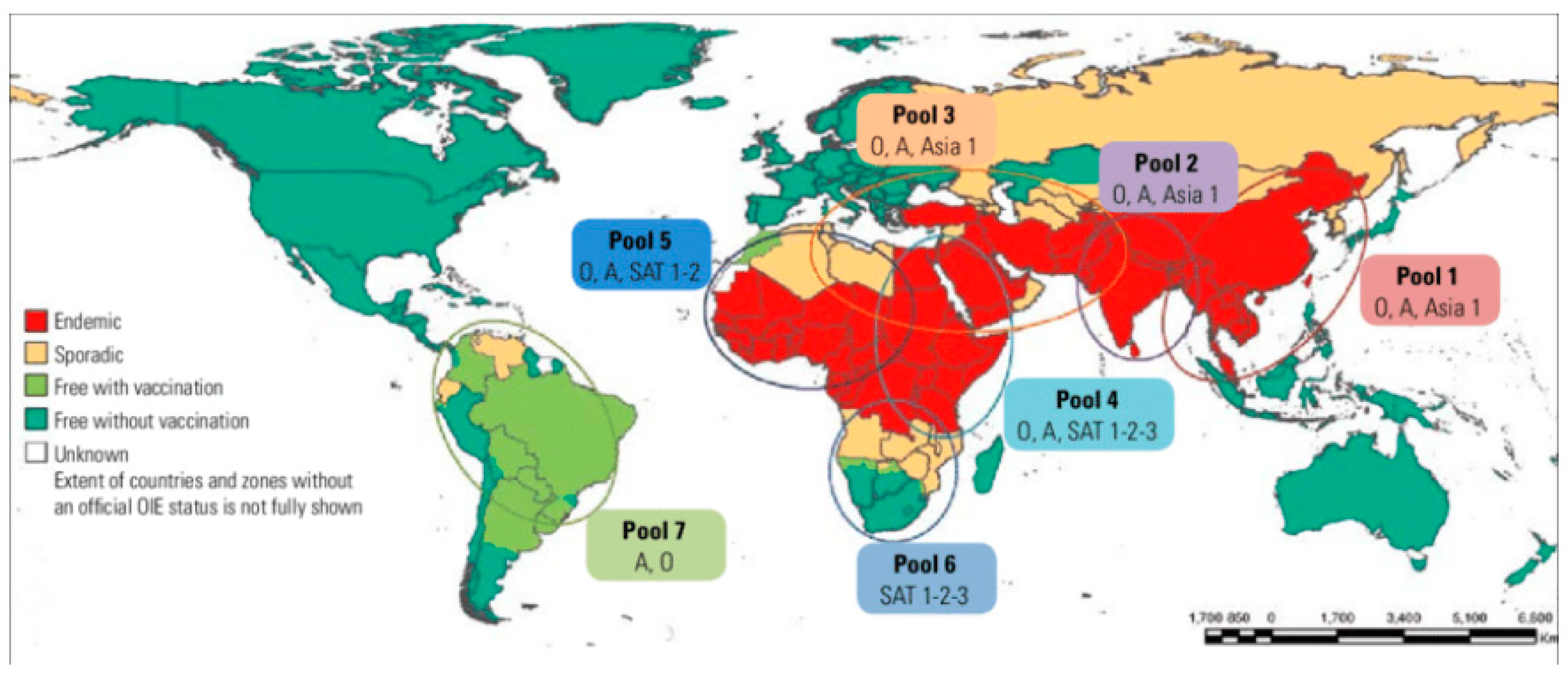
4.1. Global Distribution
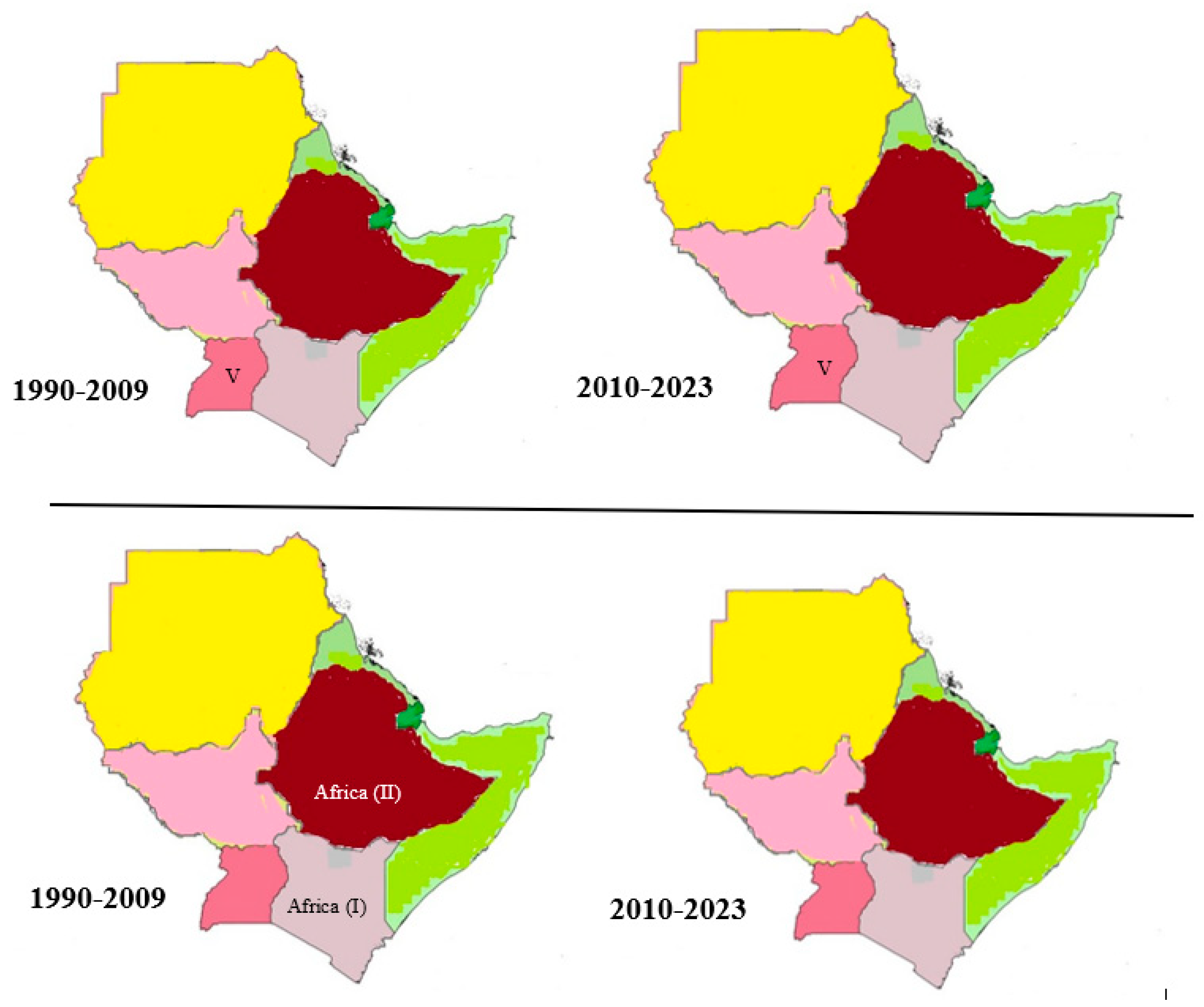
4.2. Distribution and Overall Prevalence of Foot-and-Mouth Disease in the Horn of Africa
4.3. Serotype and Topotypes in the Horn of Africa
4.3.1. Serotype O Topotypes in the Horn of Africa
4.3.2. Serotype A Topotypes in the Horn of Africa
4.3.3. Serotype SAT2 Topotypes in the Horn of Africa
4.3.4. Serotype SAT1 Topotypes in the Horn of Africa
4.3.5. Serotype SAT3 Topotypes in the Horn of Africa
4.3.6. Serotype C Topotypes in the Horn of Africa
5. The Horn of Africa and the Dynamics of FMD
6. Animal Factor Role in the Epidemiology of FMD
6.1. Acute and Persistent Infection
6.2. Animal Movement and Trade
7. Role of Wild Ungulates
8. Vaccination
9. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Ethics Statement
References
- Alary, V.; Bourzat, D. Global Animal Products’ Supply and Demand-Challenges of the Horn of Africa. 2008. Available online: https://agritrop.cirad.fr/548607/1/document_548607.pdf (accessed on 1 January 2022).
- Current Availability and Future Requirements of Livestock Feeds. Available online:https://epashupalan.com/10961/animal-husbandry/current-availability-and-future-requirements-of-livestock-feeds/ (accessed on 1 January 2022).
- The IGAD Regional Peste des Petits Ruminants (PPR) Progressive Control and Eradication Strategy. Available online: https://rr-africa.woah.org/wp-content/uploads/2019/09/progressive-control-and-eradication-strategy-1.pdf (accessed on 2 January 2022).
- Horn of Africa Drought: Regional Humanitarian Overview & Call to Action (Revised 21 September 2022). Available online:https://fscluster.org/news/horn-africa-drought-regional#:~:text=Over%208.9%20million%20livestock%E2%80%94which,latest%20FSNWG%20Drought%20Special%20Report (accessed on 2 January 2022).
- Alexandersen, S.; Mowat, N. Foot-and-mouth disease: Host range and pathogenesis. Curr. Top. Microbiol. Immunol. 2005, 288, 9–42. [Google Scholar] [CrossRef] [PubMed]
- Lloyd-Jones, K.; Mahapatra, M.; Upadhyaya, S.; Paton, D.J.; Babu, A.; Hutchings, G.; Parida, S. Genetic and antigenic characterization of serotype O FMD viruses from East Africa for the selection of suitable vaccine strain. Vaccine 2017, 35, 6842–6849. [Google Scholar] [CrossRef] [PubMed]
- Tekleghiorghis, T.; Moormann, R.J.M.; Weerdmeester, K.; Dekker, A. Foot-and-mouth Disease Transmission in Africa: Implications for Control, a Review. Transbound. Emerg. Dis. 2016, 63, 136–151. [Google Scholar] [CrossRef] [PubMed]
- Ayebazibwe, C.; Mwiine, F.N.; Tjørnehøj, K.; Balinda, S.N.; Muwanika, V.B.; Okurut, A.R.A.; Belsham, G.J.; Normann, P.; Siegismund, H.R.; Alexandersen, S. The role of African buffalos (syncerus caffer) in the maintenance of foot-and-mouth disease in Uganda. BMC Vet. Res. 2010, 6, 54. [Google Scholar] [CrossRef] [Green Version]
- Dhikusooka, M.T.; Tjørnehøj, K.; Ayebazibwe, C.; Namatovu, A.; Ruhweza, S.; Siegismund, H.R.; Wekesa, S.N.; Normann, P.; Belsham, G.J. Foot-and-mouth disease virus serotype SAT 3 in long-horned ankole Calf, Uganda. Emerg. Infect. Dis. 2015, 21, 111–114. [Google Scholar] [CrossRef]
- Paton, D.J.; Di Nardo, A.; Knowles, N.J.; Wadsworth, J.; Pituco, E.M.; Cosivi, O.; Rivera, A.M.; Kassimi, L.B.; Brocchi, E.; de Clercq, K.; et al. The history of foot-and-mouth disease virus serotype C: The first known extinct serotype? Virus Evol. 2021, 7, veab009. [Google Scholar] [CrossRef]
- Foot and Mouth Disease. Available online: https://www.woah.org/en/disease/foot-and-mouth-disease/ (accessed on 5 January 2022).
- Ayelet, G.; Mahapatra, M.; Gelaye, E.; Egziabher, B.G.; Rufeal, T.; Sahle, M.; Ferris, N.P.; Wadsworth, J.; Hutchings, G.H.; Knowles, N.J. Genetic characterization of foot-and-mouth disease viruses, Ethiopia, 1981–2007. Emerg. Infect. Dis. 2009, 15, 1409–1417. [Google Scholar] [CrossRef]
- Rweyemamu, M.; Roeder, P.; MacKay, D.; Sumption, K.; Brownlie, J.; Leforban, Y.; Valarcher, J.F.; Knowles, N.J.; Saraiva, V. Epidemiological patterns of foot-and-mouth disease worldwide. Transbound. Emerg. Dis. 2008, 55, 57–72. [Google Scholar] [CrossRef]
- WRLFMD. Foot-and-Mouth Disease. Quarterly Report January to March 2020. Available online: https://www.wrlfmd.org/sites/world/files/quick_media/WRLMEG-2020-00003B-UGA-GTR-O-SAT2.pdf (accessed on 7 March 2023).
- Megersa, B.; Beyene, B.; Abunna, F.; Regassa, A.; Amenu, K.; Rufael, T. Risk factors for foot and mouth disease seroprevalence in indigenous cattle in Southern Ethiopia: The effect of production system. Trop. Anim. Health Prod. 2009, 41, 891–898. [Google Scholar] [CrossRef]
- Wright, C.F.; Morelli, M.J.; Thébaud, G.; Knowles, N.J.; Herzyk, P.; Paton, D.J.; Haydon, D.T.; King, D.P. Beyond the consensus: Dissecting within-host viral population diversity of foot-and-mouth disease virus by using next-generation genome sequencing. J. Virol. 2011, 85, 2266–2275. [Google Scholar] [CrossRef] [Green Version]
- Ferretti, L.; Pérez-Martín, E.; Zhang, F.; Maree, F.; de Klerk-Lorist, L.M.; van Schalkwykc, L.; Juleff, N.D.; Charleston, B.; Ribeca, P. Pervasive within-host recombination and epistasis as major determinants of the molecular evolution of the foot-and-mouth disease virus capsid. Phil. Trans. R. Soc. B 2020, 374, e1008235. [Google Scholar] [CrossRef] [Green Version]
- Ferretti, L.; Di Nardo, A.; Singer, B.; Lasecka-Dykes, L.; Logan, G.; Wright, C.F.; Pérez-Martín, E.; King, D.P.; Tuthill, T.J.; Ribeca, P. Within-Host Recombination in the Foot-and-Mouth Disease Virus Genome. Viruses 2018, 10, 221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jamal, S.M.; Ferrari, G.; Ahmed, S.; Normann, P.; Belsham, G.J. Molecular characterization of serotype Asia-1 foot-and-mouth disease viruses in Pakistan and Afghanistan; emergence of a new genetic Group and evidence for a novel recombinant virus. Infect. Genet. Evol. 2011, 11, 2049–2062. [Google Scholar] [CrossRef]
- Ahmed, N.H.; Osman, N.A.; Alfouz, W.; Saeed, H.M. Serological detection and genetic characterization of foot-and-mouth disease virus from cattle in northern sudan, 2016–2018. Vet. Anim. Sci. 2021, 13, 100188. [Google Scholar] [CrossRef] [PubMed]
- Domingo, E.; Soria, M.E.; Gallego, I.; de Ávila, A.I.; García-Crespo, C.; Martínez-González, B.; Gómez, J.; Briones, C.; Gregori, J.; Quer, J.; et al. A new implication of quasispecies dynamics: Broad virus diversification in absence of external perturbations. J. Mol. Epidemiol. Evol. Genet. Infect. Dis. 2020, 82, 104278. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Shi, J.; Wang, H.; Rivera-Serrano, E.E.; Yang, D.; Zhou, G.; Sun, C.; Cameron, C.E.; Yu, L. Polymerase fidelity contributes to Foot-and-Mouth Disease Virus pathogenicity and transmissibility in vivo. J. Virol. 2020, 95, e01569-20. [Google Scholar] [CrossRef]
- Domingo, E. Quasispecies. In Encyclopedia of Virology, 2nd ed.; Granoff, A., Webster, R.G., Eds.; Elsevier: Oxford, UK, 1999. [Google Scholar]
- Domingo, E.; Escarmı, C.; Baranowski, E.; Ruiz-Jarabo, C.M.; Carrillo, E.; Núñez, J.I.; Sobrino, F. Evolution of foot-and-mouth disease virus. Virus Res. 2003, 91, 47–63. [Google Scholar] [CrossRef]
- Arzt, J.; Fish, I.H.; Bertram, M.R.; Smoliga, G.R.; Hartwig, E.J.; Pauszek, S.J.; Holinka-Patterson, L.; Segundo, F.C.D.-S.; Sitt, T.; Rieder, E.; et al. Simultaneous and Staggered Foot-and-Mouth Disease Virus Coinfection of Cattle. J. Virol. 2021, 95, e0165021. [Google Scholar] [CrossRef]
- Brito, B.; Pauszek, S.J.; Hartwig, E.J.; Smoliga, G.R.; Vu, L.T.; Dong, P.V.; Stenfeldt, C.; Rodriguez, L.L.; King, D.P.; Knowles, N.J.; et al. A traditional evolutionary history of foot-and-mouth disease viruses in Southeast Asia challenged by analyses of non-structural protein coding sequences. Sci. Rep. 2018, 8, 6472. [Google Scholar] [CrossRef] [Green Version]
- Rweyemamu, M.M.; Astudillo, V.M. Global perspective for foot and mouth disease control. Rev. Sci. Tech. 2002, 21, 765–773. [Google Scholar] [CrossRef]
- Samuel, A.R.; Knowles, N.J. Foot-and-mouth disease type O viruses exhibit genitically and geographically distinct evolutionary lineages (topotypes). J. Gen. Virol. 2001, 82, 609–621. [Google Scholar] [CrossRef] [PubMed]
- Knowles, N.J.; Samuel, A.R. Molecular epidemiology of foot-and-mouth disease virus. Virus Res. 2003, 91, 65–80. [Google Scholar] [CrossRef] [PubMed]
- Paton, D.J.; Sumption, K.J.; Charleston, B. Options for control of foot-and-mouth disease: Knowledge, capability and policy. Philos. Trans. R. Soc. B Biol. Sci. 2009, 364, 2657–2667. [Google Scholar] [CrossRef] [PubMed]
- Knowles, N.J.; Swabey, K.G.; Midgley, R.J.; Davies, P.R.; Wadsworth, J. Global molecular epidemiology of foot-and-mouth disease virus type C. In Proceedings of the 14th Meeting of the European Study Group on the Molecular Biology of Picornavirus, Saariselkä, Inari, Finland, 26 November–1 December 2006. [Google Scholar]
- Roeder, P.L.; Knowles, N.J. Foot-and-mouth disease virus type C situation: The first target for eradication? In Report of the Session of the Research Group of the Standing Technical Committee of EUFMD; Erice: Sicily, Italy, 2008. [Google Scholar]
- Di Nardo, A.; Knowles, N.J.; Paton, D.J. Combining livestock trade patterns with phylogenetics to help understand the spread of foot and mouth disease in sub-Saharan Africa, the Middle East and Southeast Asia. Rev. Sci. Tech. 2011, 30, 63–85. [Google Scholar] [CrossRef]
- Sebhatu, T.T. Foot-and-Mouth Disease Sero-Surveillance in Africa and Vaccine Matching. Available online: https://dspace.library.uu.nl/handle/1874/299019 (accessed on 10 April 2022).
- Rufael, T.; Catley, A.; Bogale, A.; Sahle, M.; Shiferaw, Y. Foot and mouth disease in the Borana pastoral system, southern Ethiopia and implications for livelihoods and international trade. Trop. Anim. Health Prod. 2008, 40, 29–38. [Google Scholar] [CrossRef]
- Dubie, T.; Negash, W. Seroprevalence of bovine foot and mouth disease (FMD) and its associated risk factors in selected districts of Afar region, Ethiopia. Vet. Med. Sci. 2021, 7, 1678–1687. [Google Scholar] [CrossRef]
- Freimanis, G.L.; Di Nardo, A.; Bankowska, K.; King, D.J.; Wadsworth, J.; Knowles, N.J.; King, D.P. Genomics and outbreaks: Foot and mouth disease. Rev. Sci. Tech. 2016, 35, 175–189. [Google Scholar] [CrossRef] [Green Version]
- Abdela, N. Sero-prevalence, risk factors and distribution of foot and mouth disease in Ethiopia. Acta Trop. 2017, 169, 125–132. [Google Scholar] [CrossRef]
- Mesfine, M.; Nigatu, S.; Belayneh, N.; Jemberu, W.T. Sero-Epidemiology of Foot and Mouth Disease in Domestic Ruminants in Amhara Region, Ethiopia. Front. Vet. Sci. 2019, 6, 130. [Google Scholar] [CrossRef] [Green Version]
- Sulayeman, M.; Dawo, F.; Mammo, B.; Gizaw, D.; Shegu, D. Isolation, molecular characterization and sero-prevalence study of foot-and-mouth disease virus circulating in central Ethiopia. BMC Vet. Res. 2018, 14, 110. [Google Scholar] [CrossRef]
- Woldemariyam, F.T.; De Vleeschauwer, A.; Hundessa, N.; Muluneh, A. Risk Factor Assessment, Sero-Prevalence, and Genotyping of the Virus that Causes Foot-and-Mouth Disease on Commercial Farms in Ethiopia from October 2018 to February 2020. Agriculture 2022, 12, 49. [Google Scholar] [CrossRef]
- Kibore, B.; Gitao, C.G.; Sangula, A.; Kitala, P. Foot and mouth disease sero-prevalence in cattle in Kenya. J. Vet. Med. Anim. Health 2014, 5, 262–268. [Google Scholar]
- Nthiwa, D.; Bett, B.; Odongo, D.; Kenya, E.; Wainaina, M.; Grazioli, S.; Foglia, E.; Brocchi, E.; Alonso, S. Seroprevalence of foot-and-mouth disease virus in cattle herds raised in Maasai Mara ecosystem in Kenya. Prev. Vet. Med. 2020, 176, 104929. [Google Scholar] [CrossRef] [PubMed]
- Omondi, G.P.; Gakuya, F.; Arzt, J.; Sangula, A.; Hartwig, E.; Pauszek, S.; Smoliga, G.; Brito, B.; Perez, A.; Obanda, V.; et al. The role of African buffalo in the epidemiology of foot-and-mouth disease in sympatric cattle and buffalo populations in Kenya. Transbound. Emerg. Dis. 2020, 67, 2206–2221. [Google Scholar] [CrossRef]
- Chepkwony, E.C.; Gitao, G.C.; Muchemi, G.M.; Sangula, A.K.; Kairu-Wanyoike, S.W. Epidemiological study on foot-and-mouth disease in small ruminants: Sero-prevalence and risk factor assessment in Kenya. PLoS ONE 2021, 16, e0234286. [Google Scholar] [CrossRef]
- Kibore, B.; Gitao, C.G.; Sangula, A.; Kitala, P. Porcine FMD Sero-prevalence in Kenya and its potential effect. Am. J. Res. Commun. 2014, 2, 1–22. Available online: https://profiles.uonbi.ac.ke/gitao/files/kibore_2.pdf (accessed on 10 April 2022).
- Tekleghiorghis, T.; Weerdmeester, K.; van Hemert-Kluitenberg, F.; Moormann, R.J.M.; Dekker, A. Foot-and-Mouth Disease Seroprevalence in Cattle in Eritrea. Transbound. Emerg. Dis. 2017, 64, 754–763. [Google Scholar] [CrossRef]
- Mwiine, F.N.; Velazquez-Salinas, L.; Ahmed, Z.; Ochwo, S.; Munsey, A.; Kenney, M.; Lutwama, J.J.; Maree, F.F.; Lobel, L.; Perez, A.M.; et al. Serological and phylogenetic characterization of foot and mouth disease viruses from Uganda during cross-sectional surveillance study in cattle between 2014 and 2017. Transbound. Emerg. Dis. 2019, 66, 2011–2024. [Google Scholar] [CrossRef]
- Mwiine, F.N.; Ayebazibwe, C.; Alexandersen, S.; Olaho-Mukani, W.; Ademun, A.R.O.; Tjørnehøj, K. Serotype Specificity of Antibodies against Foot-and-Mouth Disease Virus in Cattle in Selected Districts in Uganda. Available online: https://academicjournals.org/journal/JVMAH/article-full-text-pdf/1776E601433 (accessed on 5 February 2022).
- Ur-Rehman, S.; Arshad, M.; Hussain, I.; Iqbal, Z. Detection and Seroprevalence of Foot and Mouth Disease in Sheep and Goats in Punjab, Pakistan. Transbound. Emerg. Dis. 2014, 61, 25–30. [Google Scholar] [CrossRef]
- Alfouz, W.; Raouf, Y.A.; Ahmed, N.H.; Hamid, A.E.; Osman, N.A. Sero-epidemiology of foot-and-mouth disease in Darfur area, Western Sudan. Vet. Res. Commun. 2021, 45, 343–352. [Google Scholar] [CrossRef]
- Yousif, H.; Almutlab, A.A.; Hassen, A.A.; Al-Majali, A.; Tibbo, M. Role of small ruminants in the epidemiology of foot-and-mouth disease in Sudan. Bull. Anim. Health Prod. Afr. 2017, 65, 145–156. Available online: https://www.ajol.info/index.php/bahpa/article/view/167629 (accessed on 25 April 2022).
- Ochi, E.; Ismail, A.O. A Review on Epidemiology of Foot and Mouth Disease (FMD) in South Sudan. Rep. Opin. 2014, 6, 13–16. Available online: http://www.sciencepub.net/report (accessed on 16 January 2023).
- Tekleghiorghis, T.; Moormann, R.J.M.; Weerdmeester, K.; Dekker, A. Serological evidence of foot-and-mouth disease virus serotype C & SAT-1 infections in Eritrea. Transbound. Emerg. Dis. 2014, 61, e83–e88. [Google Scholar] [CrossRef] [PubMed]
- Tully, D.C.; Fares, M.A. The tale of a modern animal plague: Tracing the evolutionary history and determining the time-scale for foot and mouth disease virus. Virology 2008, 382, 250–256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gizaw, D.; Tesfaye, Y.; Wood, B.A.; Di Nardo, A.; Shegu, D.; Muluneh, A.; Bilata, T.; Belayneh, R.; Fentie, A.; Asgdome, H.; et al. Molecular characterization of foot-and-mouth disease viruses circulating in Ethiopia between 2008 and 2019. Transbound. Emerg. Dis. 2020, 67, 2983–2992. [Google Scholar] [CrossRef]
- Kerfua, S.D.; Shirima, G.; Kusiluka, L.; Ayebazibwe, C.; Martin, E.; Arinaitwe, E.; Cleaveland, S.; Haydon, D.T. Low topotype diversity of recent foot-and-mouth disease vrius serotypes O and A from districts located along the Uganda and Tanzania border. J. Vet. Sci. 2019, 20, e4. [Google Scholar] [CrossRef]
- Balinda, S.N.; Sangula, A.K.; Heller, R.; Muwanika, V.B.; Belsham, G.J.; Masembe, C.; Siegismund, H.R. Diversity and transboundary mobility of serotype O foot-and-mouth disease virus in East Africa: Implications for vaccination policies. Infect. Genet. Evol. J. Mol. Epidemiol. Evol. Genet. Infect. Dis. 2010, 10, 1058–1065. [Google Scholar] [CrossRef]
- WRLFMD. Foot-and-Mouth Disease Report 2013. Available online: https://www.wrlfmd.org/sites/world/files/quick_media/OIE-FAO%20FMD%20Ref%20Lab%20Report%20October-December%202013.pdf (accessed on 5 February 2022).
- Canini, L.; Blaise-Boisseau, S.; Nardo, A.D.; Shaw, A.E.; Romey, A.; Relmy, A.; Bernelin-Cottet, C.; Salomez, A.-L.; Haegeman, A.; Ularamu, H.; et al. Identification of diffusion routes of O/EA-3 topotype of foot-and-mouth disease virus in Africa and Western Asia between 1974 and 2019—A phylogeographic analysis. Transbound. Emerg. Dis. 2022, 69, 5. [Google Scholar] [CrossRef]
- WRLFMD. Foot-and-Mouth Disease Quarterly Report April to June 2018. Available online: https://www.wrlfmd.org/sites/world/files/quick_media/OIE-FAO%20FMD%20Ref%20Lab%20Report%20Apr-Jun%202018.pdf (accessed on 7 March 2023).
- Negusssie, H.; Kyule, M.N.; Yami, M. Outbreak investigations and genetic characterization of foot-and-mouth disease virus in Ethiopia in 2008/2009. Trop. Anim. Health Prod. 2011, 43, 235–243. [Google Scholar] [CrossRef]
- Banda, F.; Sinkala, Y.; Mataa, L.; Lebea, P.; Sikombe, T.; Kangwa, H.L.; Fana, E.M.; Mokopasetso, M.; Wadsworth, J.; Knowles, N.J.; et al. Characterization of Foot-and-Mouth Disease Viruses in Zambia-Implications for the Epidemiology of the Disease in Southern Africa. Viruses 2021, 13, 2195. [Google Scholar] [CrossRef]
- Banda, F.; Shilongo, A.; Hikufe, E.H.; Khaiseb, S.; Kabajani, J.; Shikongo, B.; Set, P.; Kapapero, J.K.; Shoombe, K.K.; Zaire, G.; et al. The first detection of a serotype O foot-and-mouth disease virus in Namibia. Transbound. Emerg. Dis. 2022, 69, e3261–e3267. [Google Scholar] [CrossRef]
- Foot-and-Mouth Disease. Available online: https://www.fao.org/3/cc4292en/cc4292en.pdf (accessed on 20 February 2022).
- Florian Duchatel, M.B. Circulation of Foot-and-Mouth Disease Virus in Africa and identification of the underlying constraints using Phylogeographic methods. bioRxiv 2018. [Google Scholar] [CrossRef] [Green Version]
- Munsey, A.; Norbert, F.; Sylvester, M.; Salinas, L.V.; Ahmed, Z.; Maree, F.; Rodriguez, L.L.; Rieder, E.; Perez, A.; Dellicour, S.; et al. Phylogeographic analysis of foot-and mouth disease virus serotype O dispersal and associated drivers in East Africa. Mol. Ecol. 2021, 30, 3815–3825. [Google Scholar] [CrossRef] [PubMed]
- WRLFMD. Foot-and-Mouth Disease Quarterly Report October to December 2018. Available online: https://www.wrlfmd.org/sites/world/files/quick_media/OIE-FAO%20FMD%20Ref%20Lab%20Report%20Oct-%20Dec%202018.pdf (accessed on 7 March 2023).
- WRLFMD. Foot-and-Mouth Disease Quarterly Report July to September 2018. Available online: https://www.wrlfmd.org/sites/world/files/WRLFMD-2018-00020-SUD-GTR-O-SAT2_001.pdf (accessed on 7 March 2023).
- Lycett, S.; Tanya, V.N.; Hall, M.; King, D.P.; Mazeri, S.; Mioulet, V.; Knowles, N.J.; Wadsworth, J.; Bachanek-Bankowska, K.; Ngu Ngwa, V.; et al. The evolution and phylodynamics of serotype A and SAT2 foot-and-mouth disease viruses in endemic regions of Africa. Sci. Rep. 2019, 9, 5614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bari, F.D.; Parida, S.; Tekleghiorghis, T.; Dekker, A.; Sangula, A.; Reeve, R.; Haydon, D.T.; Paton, D.J.; Mahapatra, M. Genetic and antigenic characterisation of serotype A FMD viruses from East Africa to select new vaccine strains. Vaccine 2014, 32, 5794–5800. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wekesa, S.N.; Sangula, A.K.; Belsham, G.J.; Muwanika, V.B.; Heller, R.; Balinda, S.N.; Masembe, C.; Siegismund, H.R. Infection, Genetics and Evolution Genetic diversity of serotype A foot-and-mouth disease viruses in Kenya from 1964 to 2013; implications for control strategies in eastern Africa. Infect. Genet. Evol. 2014, 21, 408–417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duchatel, F.; Bronsvoort, B.M.; Lycett, S. Phylogeographic Analysis and Identification of Factors Impacting the Diffusion of Foot-and-Mouth Disease Virus in Africa. Front. Ecol. Evol. 2019, 7, 371. [Google Scholar] [CrossRef] [Green Version]
- Xu, W.; Yang, M. Genetic variation and evolution of foot-and-mouth disease virus serotype A in relation to vaccine matching. Vaccine 2021, 39, 1420–1427. [Google Scholar] [CrossRef]
- WRLFMD. Foot-and-Mouth Disease. Quarterly Report January to December 2020. Available online: https://www.fao.org/eufmd/resources/reports/gmr/fr/ (accessed on 7 March 2023).
- Dhikusooka, M.T.; Ayebazibwe, C.; Namatovu, A.; Belsham, G.J.; Siegismund, H.R.; Wekesa, S.N.; Balinda, S.N.; Muwanika, V.B.; Tjørnehøj, K. Unrecognized circulation of SAT 1 foot-and-mouth disease virus in cattle herds around Queen Elizabeth National Park in Uganda. BMC Vet. Res. 2016, 12, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Balinda, S.N.; Tjørnehøj, K.; Muwanika, V.B.; Sangula, A.K.; Mwiine, F.N.; Ayebazibwe, C.; Masembe, H.; Siegismund, R.; Alexandersen, S. Prevalence estimates of antibodies towards foot-and-mouth disease virus in small ruminants in Uganda. Transbound. Emerg. Dis. Dec. 2009, 56, 362–371. [Google Scholar] [CrossRef]
- Sangula, A.K.; Belsham, G.J.; Muwanika, V.B.; Heller, R.; Balinda, S.N.; Masembe, C.; Siegismund, H.R. Evolutionary analysis of foot-and-mouth disease virus serotype SAT 1 isolates from east africa suggests two independent introductions from southern africa. BMC Evol. Biol. 2010, 10, 371. [Google Scholar] [CrossRef] [Green Version]
- Bouslikhane, M. Cross border movements of animals and animal products and their relevance to the epidemiology of animal diseases in Africa. Africa OIE Reg. Comm. 2015, 1–7. Available online: https://www.woah.org/app/uploads/2021/03/2015-afr2-bouslikhane-a.pdf (accessed on 22 January 2022).
- Grace, D.; Little, P. Informal trade in livestock and livestock products. Rev. Sci. Tech. 2020, 39, 183–192. [Google Scholar] [CrossRef] [PubMed]
- Biosecurity. Available online: https://hro.house.texas.gov/focus/biosecure.pdf (accessed on 7 March 2023).
- Munsey, A.; Mwiine, F.N.; Ochwo, S.; Velazquez-Salinas, L.; Ahmed, Z.; Maree, F.; Rodriguez, L.L.; Rieder, E.; Perez, A.; VanderWaal, K. Spatial distribution and risk factors for foot and mouth disease virus in Uganda: Opportunities for strategic surveillance. Prev. Vet. Med. 2019, 171, 104766. [Google Scholar] [CrossRef]
- Hekal, S.H.A.; Al-Gaabary, M.H.; El-Sayed, M.M.; Sobhy, H.M.; Fayed, A.A.A. Seroprevalence of some Infectious transboundry diseases in cattle imported from Sudan to Egypt. J. Adv. Vet. Anim. Res. 2019, 6, 92–99. [Google Scholar] [CrossRef] [PubMed]
- Wubshet, A.K.; Dai, J.; Li, Q.; Zhang, J. Review on outbreak dynamics, the endemic serotypes, and diversified topotypic profiles of foot andmouth disease virus isolates in Ethiopia from 2008 to 2018. Viruses 2019, 11, 1076. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- FAO. Prioritization of Antigen Management with International Surveillance. Available online: https://www.fao.org/3/cb1799en/cb1799en.pdf (accessed on 27 April 2022).
- Robinson, L.; Knight-Jones, T.J.D.; Charleston, B.; Rodriguez, L.L.; Gay, C.G.; Sumption, K.J.; Vosloo, W. Global Foot-and-Mouth Disease Research Update and Gap Analysis: 7—Pathogenesis and Molecular Biology. Transbound. Emerg. Dis. 2016, 63 (Suppl. S1), 63–71. [Google Scholar] [CrossRef]
- Stenfeldt, C.; Arzt, J. The Carrier Conundrum; A Review of Recent Advances and Persistent Gaps Regarding the Carrier State of Foot-and-Mouth Disease Virus. Pathogens 2020, 9, 167. [Google Scholar] [CrossRef] [Green Version]
- Swanepoel, H.; Crafford, J.; Quan, M. A Scoping Review of Viral Diseases in African Ungulates. Vet. Sci. 2021, 8, 17. [Google Scholar] [CrossRef]
- Tadesse, B.; Molla, W.; Mengsitu, A.; Jemberu, W.T. Transmission dynamics of foot and mouth disease in selected outbreak areas of northwest Ethiopia. Epidemiol. Infect. 2019, 147, e189. [Google Scholar] [CrossRef] [Green Version]
- Paton, D.J.; Gubbins, S.; King, D.P. Understanding the transmission of foot-and-mouth disease virus at different scales. Curr. Opin. Virol. 2018, 28, 85–91. [Google Scholar] [CrossRef]
- Bertram, M.R.; Vu, L.T.; Pauszek, S.J.; Brito, B.P.; Hartwig, E.J.; Smoliga, G.R.; Hoang, B.H.; Phuong, N.T.; Stenfeldt, C.; Fish, I.H.; et al. Lack of Transmission of Foot-and-Mouth Disease Virus from Persistently Infected Cattle to Naïve Cattle under Field Conditions in Vietnam. Front. Vet. Sci. 2018, 5, 174. [Google Scholar] [CrossRef] [Green Version]
- Cortey, M.; Ferretti, L.; Pérez-Martín, E.; Zhang, F.; de Klerk-Lorist, L.-M.; Scott, K.; Freimanis, G.; Seago, J.; Ribeca, P.; van Schalkwyk, L.; et al. Persistent Infection of African Buffalo (Syncerus caffer) with Foot-and-Mouth Disease Virus: Limited Viral Evolution and No Evidence of Antibody Neutralization Escape. J. Virol. 2019, 93, e00563-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maree, F.; de Klerk-Lorist, L.-M.; Gubbins, S.; Zhang, F.; Seago, J.; Pérez-Martín, E.; Reid, L.; Scott, K.; van Schalkwyk, L.; Bengis, R.; et al. Differential Persistence of Foot-and-Mouth Disease Virus in African Buffalo Is Related to Virus Virulence. J. Virol. 2016, 90, 5132–5140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arzt, J.; Belsham, G.J.; Lohse, L.; Bøtner, A.; Stenfeldt, C. Transmission of Foot-and-Mouth Disease from Persistently Infected Carrier Cattle to Naive Cattle via Transfer of Oropharyngeal Fluid. mSphere 2018, 3, e00365-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robinson, T.P.; Thornton, P.K.; Francesconi, G.N.; Kruska, R.L.; Chiozza, F.; Notenbaert, A.M.; Cecchi, G.; Herrero, M.T.; Epprecht, M.; Fritz, S.; et al. Global Livestock Production Systems; Food and Agriculture Organization of the United Nations (FAO): Rome, Italy; International Livestock Research Institute (ILRI): Rome, Italy, 2011. [Google Scholar]
- Duchatel, F. Virus Phylogeography at the Wild/Domestic Animal Interface. Ph.D. Thesis, The University of Edinburgh, Edinburgh, Scotland, 2021. Available online: https://era.ed.ac.uk/handle/1842/37962 (accessed on 10 May 2022).
- Aman, E.; Molla, W.; Gebreegizabher, Z.; Jemberu, W.T. Spatial and temporal distribution of foot and mouth disease outbreaks in Amhara region of Ethiopia in the period 1999 to 2016. BMC Vet. Res. 2020, 16, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Chaters, G.L.; Johnson, P.C.D.; Cleaveland, S.; Crispell, J.; De Glanville, W.A.; Doherty, T.; Matthews, L.; Mohr, S.; Nyasebwa, O.M.; Rossi, G.; et al. Analysing livestock network data for infectious disease control: An argument for routine data collection in emerging economies. Philos. Trans. R. Soc. Biol. Sci. 2019, 374, 20180264. [Google Scholar] [CrossRef] [Green Version]
- Cornélis, D.; Melletti, M.; Korte, L.; Ryan, S.J.; Mirabile, M.; Prin, T.; Prins, H.H. African buffalo Syncerus caffer (Sparrman, 1779). In Ecology, Evolution and Behaviour of Wild Cattle: Implications for Conservation; Cambridge University Press: Cambridge, UK, 2014; pp. 326–372. [Google Scholar] [CrossRef]
- Gortázar, C.; Barroso, P.; Nova, R.; Cáceres, G. The role of wildlife in the epidemiology and control of Foot-and-mouth-disease And Similar Transboundary (FAST) animal diseases: A review. Transbound. Emerg. Dis. 2022, 69, 2462–2473. [Google Scholar] [CrossRef] [PubMed]
- Nthiwa, D.; Alonso, S.; Odongo, D.; Kenya, E.; Bett, B. A participatory epidemiological study of major cattle diseases amongst Maasai pastoralists living in wildlife-livestock interfaces in Maasai Mara, Kenya. Trop. Anim. Health Prod. 2019, 51, 1097–1103. [Google Scholar] [CrossRef] [Green Version]
- Jemberu, W.T.; Molla, W.; Dagnew, T.; Rushton, J.; Hogeveen, H. Farmers’ willingness to pay for foot and mouth disease vaccine in different cattle production systems in Amhara region of Ethiopia. PLoS ONE 2020, 15, e0239829. [Google Scholar] [CrossRef]
- Casey-Bryars, M.; Reeve, R.; Bastola, U.; Knowles, N.J.; Auty, H.; Bachanek-Bankowska, K.; Fowler, V.L.; Fyumagwa, R.; Kazwala, R.; Kibona, T.; et al. Waves of endemic foot-and-mouth disease in eastern Africa suggest feasibility of proactive vaccination approaches. Nat. Ecol. Evol. 2018, 2, 1449–1457. Available online: http://www.nature.com/natecolevol (accessed on 12 December 2022). [CrossRef] [Green Version]
- Foot and Mouth Disease (FMD) Vaccine Description. Available online: https://www.nvi.com.et/products/vaccines-against/ruminant-and-equine-diseases/fmd-2/ (accessed on 20 February 2023).
- Tesfaye, Y.; Khan, F.; Gelaye, E. Vaccine matching and antigenic variability of foot-and-mouth disease virus serotypes O and A from 2018 Ethiopian isolates. Int. Microbiol. 2021, 25, 47–59. [Google Scholar] [CrossRef]
- Tesfaye, Y.; Khan, F.; Yami, M.; Wadsworth, J.; Knowles, N.J.; King, D.P.; Gelaye, E. A vaccine-matching assessment of different genetic variants of serotype O foot-and-mouth disease virus isolated in Ethiopia between 2011 and 2014. Arch. Virol. 2020, 165, 1749–1757. [Google Scholar] [CrossRef] [PubMed]
- Kenubih, A. Foot and Mouth Disease Vaccine Development and Challenges in Inducing Long-Lasting Immunity: Trends and Current Perspectives. Vet. Med. Res. Rep. 2021, 12, 205–215. [Google Scholar] [CrossRef] [PubMed]
- Sangula, A.K.; Siegismund, H.R.; Belsham, G.J.; Balinda, S.N.; Masembe, C.; Muwanika, V.B. Low diversity of foot-and-mouth disease serotype C virus in Kenya: Evidence for probable vaccine strain re-introductions in the field. Epidemiol. Infect. 2011, 139, 189–196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jackson, B.; Harvey, Y.; Perez-Martin, E.; Wilsden, G.; Juleff, N.; Charleston, B.; Seago, J. The selection of naturally stable candidate foot-and-mouth disease virus vaccine strains for East Africa. Vaccine 2021, 39, 5015–5024. [Google Scholar] [CrossRef]
- Hammond, J.M.; Maulidi, B.; Henning, N. Targeted FMD Vaccines for Eastern Africa: The AgResults Foot and Mouth Disease Vaccine Challenge Project. Viruses 2021, 13, 1830. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name of Wild Ungulates | Ethiopia | Kenya | Sudan | Eritrea | Djibouti | Uganda | South Sudan | Somalia |
|---|---|---|---|---|---|---|---|---|
| Buffalo, African/Cape | yes | yes | yes | - | - | yes | yes | yes |
| Elephant, African | yes | yes | - | - | - | yes | yes | |
| Bushpig | yes | yes | yes | - | - | yes | yes | yes |
| Grant’s Gazelle | yes | yes | - | - | - | yes | yes | - |
| Gazelle | yes | yes | - | - | - | - | - | yes |
| Kudu | yes | yes | yes | yes | yes | yes | yes | |
| Giraffe | yes | yes | yes | yes | - | yes | yes | yes |
| Oribi | yes | yes | yes | - | - | - | - | - |
| Reedbuck, Bohor | yes | yes | yes | yes | yes | |||
| Roan antelope | yes | yes | yes | - | - | - | - | - |
| Impala | - | yes | - | - | - | yes | - | - |
| warthog | yes | yes | yes | - | - | yes | - | - |
| Günther’s dik-dik, | yes | yes | yes | yes | yes | |||
| Gazella | yes | yes | yes | yes | - | yes | - | |
| Salt’s dik-dik | - | yes | yes | - | yes | - | yes | yes |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Woldemariyam, F.T.; Kariuki, C.K.; Kamau, J.; De Vleeschauwer, A.; De Clercq, K.; Lefebvre, D.J.; Paeshuyse, J. Epidemiological Dynamics of Foot-and-Mouth Disease in the Horn of Africa: The Role of Virus Diversity and Animal Movement. Viruses 2023, 15, 969. https://doi.org/10.3390/v15040969
Woldemariyam FT, Kariuki CK, Kamau J, De Vleeschauwer A, De Clercq K, Lefebvre DJ, Paeshuyse J. Epidemiological Dynamics of Foot-and-Mouth Disease in the Horn of Africa: The Role of Virus Diversity and Animal Movement. Viruses. 2023; 15(4):969. https://doi.org/10.3390/v15040969
Chicago/Turabian StyleWoldemariyam, Fanos Tadesse, Christopher Kinyanjui Kariuki, Joseph Kamau, Annebel De Vleeschauwer, Kris De Clercq, David J. Lefebvre, and Jan Paeshuyse. 2023. "Epidemiological Dynamics of Foot-and-Mouth Disease in the Horn of Africa: The Role of Virus Diversity and Animal Movement" Viruses 15, no. 4: 969. https://doi.org/10.3390/v15040969