

Immune Responses to Influenza D Virus in Calves Previously Infected with Bovine Viral Diarrhea Virus 2
 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Viruses and Cells
2.2. Experimental BVDV and IDV Infection in Calves—Study Design
2.3. BVDV and IDV RT-qPCR
2.4. Hemagglutination Inhibition (HI) and Virus Neutralization (VN) Assays
2.5. Assessment of Cell-Mediated Immune Responses to IDV
3. Results
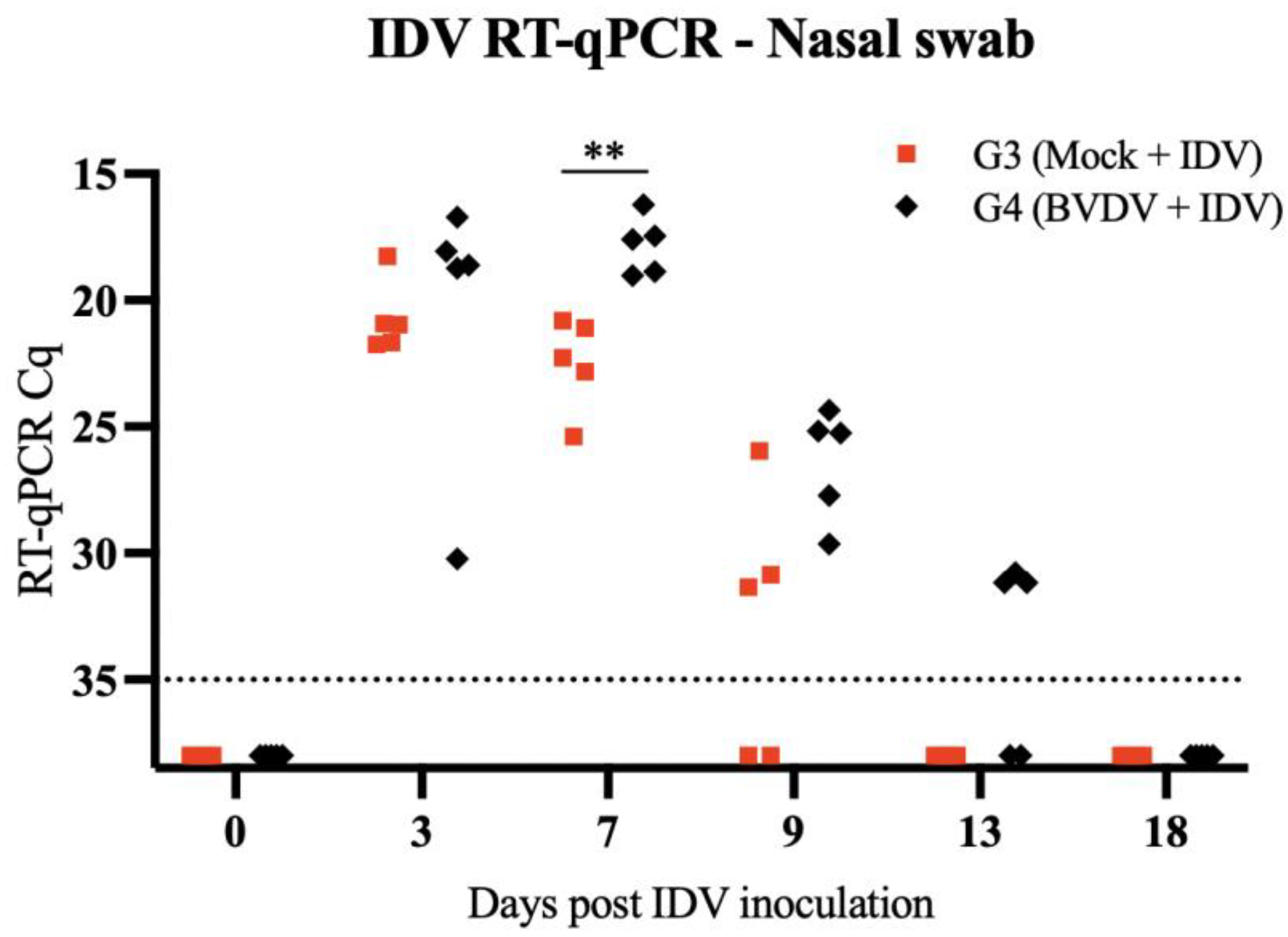
3.1. Clinical Signs and Viral Detection
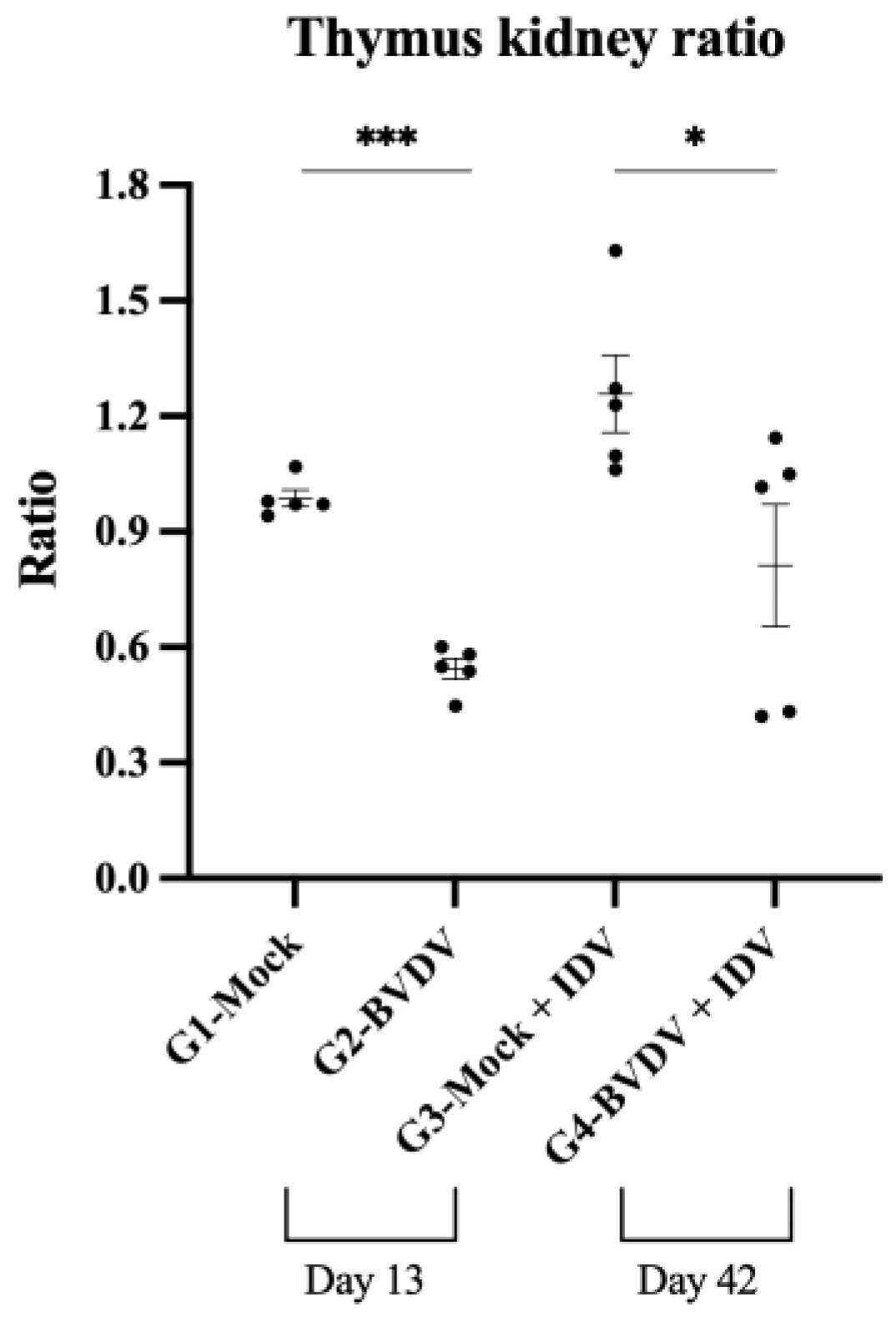
3.2. Thymus Evaluation
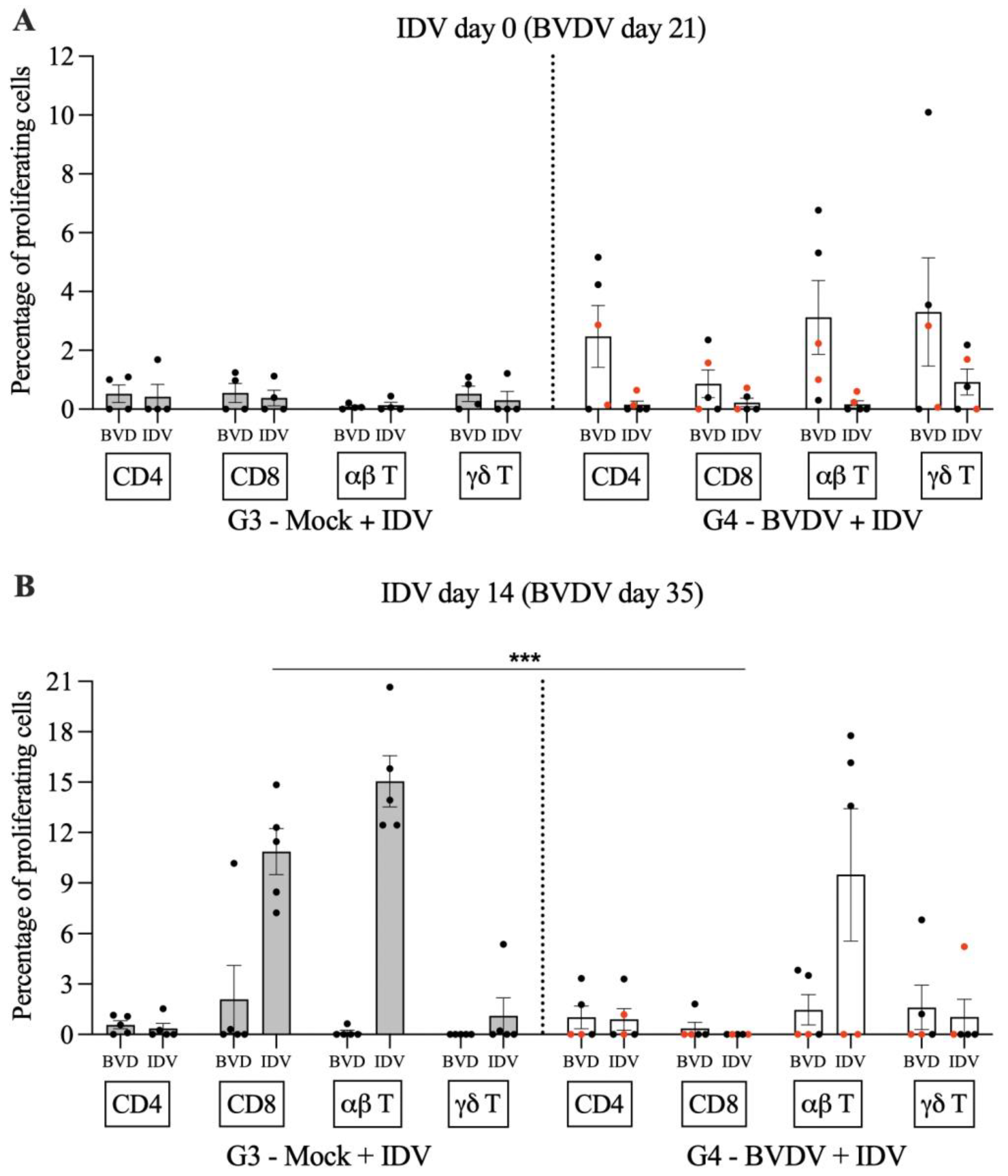
3.3. Cell-Mediated Immune Responses to IDV
3.4. Serological Responses to BVDV and IDV
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- USDA-APHIS. Death Loss in U.S. Cattle and Calves Due to Predator and Nonpredator Causes, 2015; USDA-APHIS: Riverdale, MD, USA, 2017.
- Taylor, J.D.; Fulton, R.W.; Lehenbauer, T.W.; Step, D.L.; Confer, A.W. The epidemiology of bovine respiratory disease: What is the evidence for predisposing factors? Can. Vet. J. 2010, 51, 1095. [Google Scholar] [PubMed]
- Fulton, R.W. Viruses in Bovine Respiratory Disease in North America: Knowledge Advances Using Genomic Testing. Vet. Clin. N. Am. Food Anim. Pract. 2020, 36, 321–332. [Google Scholar] [CrossRef] [PubMed]
- Ridpath, J.F.; Fulton, R.W.; Bauermann, F.V.; Falkenberg, S.M.; Welch, J.; Confer, A.W. Sequential exposure to bovine viral diarrhea virus and bovine coronavirus results in increased respiratory disease lesions: Clinical, immunologic, pathologic, and immunohistochemical findings. J. Vet. Diagn. Investig. Off. Publ. Am. Assoc. Vet. Lab. Diagn. Inc 2020, 32, 513–526. [Google Scholar] [CrossRef] [PubMed]
- Ng, T.F.F.; Kondov, N.O.; Deng, X.; Van Eenennaam, A.; Neibergs, H.L.; Delwart, E. A Metagenomics and Case-Control Study To Identify Viruses Associated with Bovine Respiratory Disease. J. Virol. 2015, 89, 5340–5349. [Google Scholar] [CrossRef] [PubMed]
- Hause, B.M.; Ducatez, M.; Collin, E.A.; Ran, Z.; Liu, R.; Sheng, Z.; Armien, A.; Kaplan, B.; Chakravarty, S.; Hoppe, A.D.; et al. Isolation of a Novel Swine Influenza Virus from Oklahoma in 2011 Which Is Distantly Related to Human Influenza C Viruses. PLoS Pathog. 2013, 9, e1003176. [Google Scholar] [CrossRef] [PubMed]
- Shaw, M.; Palese, P. Fields Virology; Knipe, D., Howley, P., Eds.; Wolters Kluwer Lippincott Williams & Wilkins: Philadelphia, PE, USA, 2013; p. 2456. [Google Scholar]
- Hause, B.M.; Collin, E.A.; Liu, R.; Huang, B.; Sheng, Z.; Lu, W.; Wang, D.; Nelson, E.A.; Li, F. Characterization of a novel influenza virus in cattle and swine: Proposal for a new genus in the Orthomyxoviridae family. MBio 2014, 5, 10–1128. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, L.; Eckard, L.; Epperson, W.B.; Long, L.P.; Smith, D.; Huston, C.; Genova, S.; Webby, R.; Wan, X.F. Influenza D virus infection in Mississippi beef cattle. Virology 2015, 486, 28–34. [Google Scholar] [CrossRef] [PubMed]
- Su, S.; Fu, X.; Li, G.; Kerlin, F.; Veit, M. Novel Influenza D virus: Epidemiology, pathology, evolution and biological characteristics. Virulence 2017, 8, 1580–1591. [Google Scholar] [CrossRef]
- White, S.K.; Ma, W.; McDaniel, C.J.; Gray, G.C.; Lednicky, J.A. Serologic evidence of exposure to influenza D virus among persons with occupational contact with cattle. J. Clin. Virol. 2016, 81, 31–33. [Google Scholar] [CrossRef]
- Ferguson, L.; Olivier, A.K.; Genova, S.; Epperson, W.B.; Smith, D.R.; Schneider, L.; Barton, K.; McCuan, K.; Webby, R.J.; Wan, X.-F.; et al. Pathogenesis of Influenza D Virus in Cattle. J. Virol. 2016, 90, 5636–5642. [Google Scholar] [CrossRef]
- Cassard, H.; Ducatez, M.; Valarcher, J.-F.; Pinard, A.; Näslund, K.; Foret, C.; Hägglund, S.; Salem, E.; Corre, T.; Meyer, G.; et al. Pathogenesis, host innate immune response and aerosol transmission of Influenza D virus in cattle. J. Virol. 2019, 93, 10–1128. [Google Scholar] [CrossRef]
- Hause, B.M.; Huntimer, L.; Falkenberg, S.; Henningson, J.; Lechtenberg, K.; Halbur, T. An inactivated influenza D virus vaccine partially protects cattle from respiratory disease caused by homologous challenge. Vet. Microbiol. 2017, 199, 47–53. [Google Scholar] [CrossRef] [PubMed]
- Bauermann, F.V.; Falkenberg, S.M.; Decaro, N.; Flores, E.F.; Ridpath, J.F. Experimental infection of calves, sheep, goats and pigs with HoBi-like viruses by direct inoculation or exposure to persistently infected calves. Vet. Microbiol. 2015, 181, 289–293. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, B.S.; Falkenberg, S.; Dassanayake, R.; Neill, J.; Velayudhan, B.; Li, F.; Vincent, A.L. Virus strain influenced the interspecies transmission of influenza D virus between calves and pigs. Transbound. Emerg. Dis. 2021, 68, 3396–3404. [Google Scholar] [CrossRef] [PubMed]
- Ridpath, J.F.; Bolin, S.R.; Dubovi, E.J. Segregation of Bovine Viral Diarrhea Virus into Genotypes. Virology 1994, 205, 66–74. [Google Scholar] [CrossRef] [PubMed]
- Houe, H. Economic impact of BVDV infection in dairies. Biologicals 2003, 31, 137–143. [Google Scholar] [CrossRef] [PubMed]
- Larson, R.L. Bovine Viral Diarrhea Virus–Associated Disease in Feedlot Cattle. Vet. Clin. N. Am. Food Anim. Pract. 2015, 31, 367–380. [Google Scholar] [CrossRef] [PubMed]
- Baker, J.C. The clinical manifestations of bovine viral diarrhea infection. Vet. Clin. N. Am. Food Anim. Pract. 1995, 11, 425–445. [Google Scholar] [CrossRef]
- Baker, J.C. Bovine viral diarrhea virus: A review. J. Am. Vet. Med. Assoc. 1987, 190, 1449–1458. [Google Scholar]
- Ridpath, J.F.; Falkenberg, S.M.; Bauermann, F.V.; VanderLey, B.L.; Do, Y.; Flores, E.F.; Rodman, D.M.; Neill, J.D. Comparison of acute infection of calves exposed to a high-virulence or low-virulence bovine viral diarrhea virus or a HoBi-like virus. Am. J. Vet. Res. 2013, 74, 438–442. [Google Scholar] [CrossRef]
- Falkenberg, S.M.; Bauermann, F.V.; Ridpath, J.F. Characterization of thymus-associated lymphoid depletion in bovine calves acutely or persistently infected with bovine viral diarrhea virus 1, bovine viral diarrhea virus 2 or HoBi-like pestivirus. Arch. Virol. 2017, 162, 3473–3480. [Google Scholar] [CrossRef] [PubMed]
- Liebler-Tenorio, E.M.; Ridpath, J.F.; Neill, J.D. Distribution of viral antigen and development of lesions after experimental infection of calves with a BVDV 2 strain of low virulence. J. Vet. Diagn. Investig. 2003, 15, 221–232. [Google Scholar] [CrossRef] [PubMed]
- Liebler-Tenorio, E.M.; Greiser-Wilke, I.; Pohlenz, J.F. Organ and tissue distribution of the antigen of the cytopathogenie bovine virus diarrhea virus in the early and advanced phase of experimental mucosal disease. Arch. Virol. 1997, 142, 1613–1634. [Google Scholar] [CrossRef] [PubMed]
- Liebler-Tenorio, E.M.; Ridpath, J.F.; Neill, J.D. Distribution of viral antigen and tissue lesions in persistent and acute infection with the homologous strain of noncytopathic bovine viral diarrhea virus. J. Vet. Diagn. Investig. 2004, 16, 388–396. [Google Scholar] [CrossRef] [PubMed]
- Frink, S.; Grummer, B.; Pohlenz, J.F.; Liebler-Tenorio, E.M. Changes in distribution and numbers of CD4+ and CD8+ T-lymphocytes in lymphoid tissues and intestinal mucosa in the early phase of experimentally induced early onset mucosal disease in cattle. J. Vet. Med. Ser. B 2002, 49, 476–483. [Google Scholar] [CrossRef] [PubMed]
- Falkenberg, S.M.; Johnson, C.; Bauermann, F.V.; McGill, J.; Palmer, M.V.; Sacco, R.E.; Ridpath, J.F. Changes observed in the thymus and lymph nodes 14 days after exposure to BVDV field strains of enhanced or typical virulence in neonatal calves. Vet. Immunol. Immunopathol. 2014, 160, 70–80. [Google Scholar] [CrossRef]
- Risalde, M.A.; Molina, V.; Sánchez-Cordón, P.J.; Pedrera, M.; Romero-Palomo, F.; Bautista, M.J.; Moreno, A.; Gómez-Villamandos, J.C. Comparison of pathological changes and viral antigen distribution in tissues of calves with and without preexisting bovine viral diarrhea virus infection following challenge with bovine herpesvirus-1. Am. J. Vet. Res. 2013, 74, 598–610. [Google Scholar] [CrossRef]
- Romero-Palomo, F.; Risalde, M.A.; Gómez-Villamandos, J.C. Immunopathologic Changes in the Thymus of Calves Pre-infected with BVDV and Challenged with BHV-1. Transbound. Emerg. Dis. 2017, 64, 574–584. [Google Scholar] [CrossRef]
- Cunningham, C.P.; Kimpton, W.G.; Holder, J.E.; Cahill, R.N.P. Thymic export in aged sheep: A continuous role for the thymus throughout pre- and postnatal life. Eur. J. Immunol. 2001, 31, 802–811. [Google Scholar] [CrossRef]
- Savino, W. The thymus is a common target organ in infectious diseases. PLoS Pathog. 2006, 2, e62. [Google Scholar] [CrossRef]
- Romero-Palomo, F.; Risalde, M.A.; Molina, V.; Lauzi, S.; Bautista, M.J.; Gómez-Villamandos, J.C. Characterization of thymus atrophy in calves with subclinical BVD challenged with BHV-1. Vet. Microbiol. 2015, 177, 32–42. [Google Scholar] [CrossRef] [PubMed]
- Risalde, M.A.; Molina, V.; Sánchez-Cordón, P.J.; Pedrera, M.; Panadero, R.; Romero-Palomo, F.; Gómez-Villamandos, J.C. Response of proinflammatory and anti-inflammatory cytokines in calves with subclinical bovine viral diarrhea challenged with bovine herpesvirus-1. Vet. Immunol. Immunopathol. 2011, 144, 135–143. [Google Scholar] [CrossRef] [PubMed]
- Dudakov, J.A.; Hanash, A.M.; Jenq, R.R.; Young, L.F.; Ghosh, A.; Singer, N.V.; West, M.L.; Smith, O.M.; Holland, A.M.; Tsai, J.J.; et al. Interleukin-22 drives endogenous thymic regeneration in mice. Science 2012, 336, 91–95. [Google Scholar] [CrossRef] [PubMed]
- van den Broek, T.; Delemarre, E.M.; Janssen, W.J.M.; Nievelstein, R.A.J.; Broen, J.C.; Tesselaar, K.; Borghans, J.A.M.; Nieuwenhuis, E.E.S.; Prakken, B.J.; Mokry, M.; et al. Neonatal thymectomy reveals differentiation and plasticity within human naive T cells. J. Clin. Investig. 2016, 126, 1126–1136. [Google Scholar] [CrossRef] [PubMed]
- Granadier, D.; Iovino, L.; Kinsella, S.; Dudakov, J.A. Dynamics of thymus function and T cell receptor repertoire breadth in health and disease. Semin. Immunopathol. 2021, 43, 119–134. [Google Scholar] [CrossRef] [PubMed]
- Ross, E.A.; Coughlan, R.E.; Flores-Langarica, A.; Lax, S.; Nicholson, J.; Desanti, G.E.; Marshall, J.L.; Bobat, S.; Hitchcock, J.; White, A.; et al. Thymic function is maintained during Salmonella-induced atrophy and recovery. J. Immunol. 2012, 189, 4266–4274. [Google Scholar] [CrossRef] [PubMed]
- Anz, D.; Thaler, R.; Stephan, N.; Waibler, Z.; Trauscheid, M.J.; Scholz, C.; Kalinke, U.; Barchet, W.; Endres, S.; Bourquin, C. Activation of melanoma differentiation-associated gene 5 causes rapid involution of the thymus. J. Immunol. 2009, 182, 6044–6050. [Google Scholar] [CrossRef]
- Velardi, E.; Tsai, J.J.; van den Brink, M.R.M. T cell regeneration after immunological injury. Nat. Rev. Immunol. 2021, 21, 277–291. [Google Scholar] [CrossRef]
- Mackall, C.L.; Fleisher, T.A.; Brown, M.R.; Andrich, M.P.; Chen, C.C.; Feuerstein, I.M.; Horowitz, M.E.; Magrath, I.T.; Shad, A.T.; Steinberg, S.M.; et al. Age, Thymopoiesis, and CD4+ T-Lymphocyte Regeneration after Intensive Chemotherapy. N. Engl. J. Med. 1995, 332, 143–149. [Google Scholar] [CrossRef]
- Bauermann, F.V.; Flores, E.F.; Falkenberg, S.M.; Weiblen, R.; Ridpath, J.F. Lack of evidence for the presence of emerging HoBi-like viruses in North American fetal bovine serum lots. J. Vet. Diagn. Investig. 2014, 26, 10–17. [Google Scholar] [CrossRef]
- Kraybill, H.F.; Hiner, R.L.; Farnworth, V.M. The Relation of Organ Weights to Lean Body Mass and Empty Body Weight in Cattle. J. Anim. Sci. 1954, 13, 548–555. [Google Scholar] [CrossRef]
- Thrusfield, M. Veterinary Epidemiologyle. In Veterinary Epidemiology; Thrusfield, M., Christley, R., Eds.; Wiley Blackwell: Singapore, 2018; p. 864. ISBN 978-1-118-28028-7. [Google Scholar]
- Rebhun, W.C.; French, T.W.; Perdrizet, J.A.; Dubovi, E.J.; Dill, S.G.; Karcher, L.F. Thrombocytopenia associated with acute bovine virus diarrhea infection in cattle. J. Vet. Intern. Med. 1989, 3, 42–46. [Google Scholar] [CrossRef] [PubMed]
- Bolin, S.R.; McClurkin, A.W.; Coria, M.F. Effects of bovine viral diarrhea virus on the percentages and absolute numbers of circulating 3 and T lymphocytes in cattle. Am. J. Vet. Res. 1985, 46, 884–886. [Google Scholar] [PubMed]
- Messias, C.V.; Loss-Morais, G.; Carvalho, J.B.d.; González, M.N.; Cunha, D.P.; Vasconcelos, Z.; Arge, L.W.P.; Farias-de-Oliveira, D.A.; Gerber, A.L.; Portari, E.A.; et al. Zika virus targets the human thymic epithelium. Sci. Rep. 2020, 10, 1378. [Google Scholar] [CrossRef] [PubMed]
- Thapa, P.; Farber, D.L. The Role of the Thymus in the Immune Response. Thorac. Surg. Clin. 2019, 29, 123–131. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Ruíz, G.U.; Morales-Sánchez, A.; Bhandoola, A. Transcriptional and epigenetic regulation in thymic epithelial cells. Immunol. Rev. 2022, 305, 43–58. [Google Scholar] [CrossRef] [PubMed]
- Kurd, N.; Robey, E.A. T-cell selection in the thymus: A spatial and temporal perspective. Immunol. Rev. 2016, 271, 114–126. [Google Scholar] [CrossRef]
- Ellis, J.A.; Davis, W.C.; Belden, E.L.; Pratt, D.L. Flow cytofluorimetric analysis of lymphocyte subset alterations in cattle infected with bovine viral diarrhea virus. Vet. Pathol. 1988, 25, 231–236. [Google Scholar] [CrossRef]
- Brodersen, B.W.; Kelling, C.L. Alteration of leukocyte populations in calves concurrently infected with bovine respiratory syncytial virus and bovine viral diarrhea virus. Viral Immunol. 1999, 12, 323–334. [Google Scholar] [CrossRef]
- Gånheim, C.; Johannisson, A.; Ohagen, P.; Persson Waller, K. Changes in peripheral blood leucocyte counts and subpopulations after experimental infection with BVDV and/or Mannheimia haemolytica. J. Vet. Med. B Infect. Dis. Vet. Public Health 2005, 52, 380–385. [Google Scholar] [CrossRef]
- Chase, C.C.L. The impact of BVDV infection on adaptive immunity. Biologicals 2013, 41, 52–60. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number of Animals with Positive BVDV RT-qPCR | Sample Type | D0 | D3 | D7 | D9 | D13 | D18 |
|---|---|---|---|---|---|---|---|
| G2 BVDV | Nasal swab | 0 | 0 | 5 | 4 | 0 | NA * |
| Serum | 0 | 0 | 1 | 5 | 0 | NA | |
| G4 BVDV + IDV | Nasal swab | 0 | 2 | 5 | 5 | 0 | 0 |
| Serum | 0 | 0 | 2 | 3 | 0 | 0 |
| Number of Animals Presenting Fever | D1 | D2 | D3 | D4 | D5 | D6 | D7 | D8 | |
|---|---|---|---|---|---|---|---|---|---|
| Post-BVDV inoculation | G2 BVDV | - | 2 | - | 1 | - | 3 | 4 | 1 |
| G4 BVDV + IDV | - | - | 2 | - | 2 | 2 | 5 | 2 | |
| Post-IDV inoculation | G3 Mock + IDV | 1 | 1 | - | - | - | - | 1 | - |
| G4 BVDV + IDV | 3 | - | - | - | 1 | - | - | - | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vicosa Bauermann, F.; Falkenberg, S.; Rudd, J.M.; Peter, C.M.; Merchioratto, I.; Ritchey, J.W.; Gilliam, J.; Taylor, J.; Ma, H.; Maggioli, M.F. Immune Responses to Influenza D Virus in Calves Previously Infected with Bovine Viral Diarrhea Virus 2. Viruses 2023, 15, 2442. https://doi.org/10.3390/v15122442
Vicosa Bauermann F, Falkenberg S, Rudd JM, Peter CM, Merchioratto I, Ritchey JW, Gilliam J, Taylor J, Ma H, Maggioli MF. Immune Responses to Influenza D Virus in Calves Previously Infected with Bovine Viral Diarrhea Virus 2. Viruses. 2023; 15(12):2442. https://doi.org/10.3390/v15122442
Chicago/Turabian StyleVicosa Bauermann, Fernando, Shollie Falkenberg, Jennifer M. Rudd, Cristina Mendes Peter, Ingryd Merchioratto, Jerry W. Ritchey, John Gilliam, Jared Taylor, Hao Ma, and Mayara Fernanda Maggioli. 2023. "Immune Responses to Influenza D Virus in Calves Previously Infected with Bovine Viral Diarrhea Virus 2" Viruses 15, no. 12: 2442. https://doi.org/10.3390/v15122442