

Antiviral Susceptibilities of Distinct Lineages of Influenza C and D Viruses

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Viruses and Cells
2.2. Antiviral Compounds
2.3. Focus Reduction Assay
2.4. Genetic Analysis
2.5. Statistical Analysis
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Furuta, Y.; Komeno, T.; Nakamura, T. Favipiravir (T-705), a broad spectrum inhibitor of viral RNA polymerase. Proc. Jpn. Acad. Ser. B 2017, 93, 449–463. [Google Scholar] [CrossRef] [Green Version]
- Tisdale, M. Influenza M2 Ion-Channel and Neuraminidase Inhibitors. In Antimicrobial Drug Resistance; Mayers, D.L., Ed.; Humana Press: Totowa, NJ, USA, 2009; Chapter 31; pp. 421–447. [Google Scholar]
- Omoto, S.; Speranzini, V.; Hashimoto, T.; Noshi, T.; Yamaguchi, H.; Kawai, M.; Kawaguchi, K.; Uehara, T.; Shishido, T.; Naito, A.; et al. Characterization of influenza virus variants induced by treatment with the endonuclease inhibitor baloxavir marboxil. Sci. Rep. 2018, 8, 9633. [Google Scholar] [CrossRef]
- Takashita, E. Influenza Polymerase Inhibitors: Mechanisms of Action and Resistance. Cold Spring Harb. Perspect. Med. 2020, 11, a038687. [Google Scholar] [CrossRef] [Green Version]
- Bright, R.A.; Medina, M.-J.; Xu, X.; Perez-Oronoz, G.; Wallis, T.R.; Davis, X.M.; Povinelli, L.; Cox, N.J.; Klimov, A.I. Incidence of adamantane resistance among influenza A (H3N2) viruses isolated worldwide from 1994 to 2005: A cause for concern. Lancet 2005, 366, 1175–1181. [Google Scholar] [CrossRef]
- Imai, M.; Yamashita, M.; Sakai-Tagawa, Y.; Iwatsuki-Horimoto, K.; Kiso, M.; Murakami, J.; Yasuhara, A.; Takada, K.; Ito, M.; Nakajima, N.; et al. Influenza A variants with reduced susceptibility to baloxavir isolated from Japanese patients are fit and transmit through respiratory droplets. Nat. Microbiol. 2019, 5, 27–33. [Google Scholar] [CrossRef]
- Takashita, E.; Ichikawa, M.; Morita, H.; Ogawa, R.; Fujisaki, S.; Shirakura, M.; Miura, H.; Nakamura, K.; Kishida, N.; Kuwahara, T.; et al. Human-to-Human Transmission of Influenza A(H3N2) Virus with Reduced Susceptibility to Baloxavir, Japan, February 2019. Emerg. Infect. Dis. 2019, 25, 2108–2111. [Google Scholar] [CrossRef] [Green Version]
- Takashita, E.; Kawakami, C.; Ogawa, R.; Morita, H.; Fujisaki, S.; Shirakura, M.; Miura, H.; Nakamura, K.; Kishida, N.; Kuwahara, T.; et al. Influenza A(H3N2) virus exhibiting reduced susceptibility to baloxavir due to a polymerase acidic subunit I38T substitution detected from a hospitalised child without prior baloxavir treatment, Japan, January 2019. Eurosurveillance 2019, 24, 1900170. [Google Scholar] [CrossRef] [Green Version]
- Govorkova, E.A.; Takashita, E.; Daniels, R.S.; Fujisaki, S.; Presser, L.D.; Patel, M.C.; Huang, W.; Lackenby, A.; Nguyen, H.T.; Pereyaslov, D.; et al. Global update on the susceptibilities of human influenza viruses to neuraminidase inhibitors and the cap-dependent endonuclease inhibitor baloxavir, 2018–2020. Antivir. Res. 2022, 200, 105281. [Google Scholar] [CrossRef]
- Matsuzaki, Y.; Sugawara, K.; Furuse, Y.; Shimotai, Y.; Hongo, S.; Oshitani, H.; Mizuta, K.; Nishimura, H. Genetic Lineage and Reassortment of Influenza C Viruses Circulating between 1947 and 2014. J. Virol. 2016, 90, 8251–8265. [Google Scholar] [CrossRef] [Green Version]
- Huang, C.; Yu, J.; Hause, B.M.; Park, J.Y.; Sreenivasan, C.; Uprety, T.; Sheng, Z.; Wang, D.; Li, F. Emergence of new phylogenetic lineage of Influenza D virus with broad antigenicity in California, United States. Emerg. Microbes Infect. 2021, 10, 739–742. [Google Scholar] [CrossRef]
- Peng, Q.; Liu, Y.; Peng, R.; Wang, M.; Yang, W.; Song, H.; Chen, Y.; Liu, S.; Han, M.; Zhang, X.; et al. Structural insight into RNA synthesis by influenza D polymerase. Nat. Microbiol. 2019, 4, 1750–1759. [Google Scholar] [CrossRef]
- Furuta, Y.; Takahashi, K.; Fukuda, Y.; Kuno, M.; Kamiyama, T.; Kozaki, K.; Nomura, N.; Egawa, H.; Minami, S.; Watanabe, Y.; et al. In Vitro and In Vivo Activities of Anti-Influenza Virus Compound T-705. Antimicrob. Agents Chemother. 2002, 46, 977–981. [Google Scholar] [CrossRef] [Green Version]
- Mishin, V.P.; Patel, M.C.; Chesnokov, A.; De La Cruz, J.; Nguyen, H.T.; Lollis, L.; Hodges, E.; Jang, Y.; Barnes, J.; Uyeki, T.; et al. Susceptibility of Influenza A, B, C, and D Viruses to Baloxavir. Emerg. Infect. Dis. 2019, 25, 1969–1972. [Google Scholar] [CrossRef] [Green Version]
- Takada, K.; Kawakami, C.; Fan, S.; Chiba, S.; Zhong, G.; Gu, C.; Shimizu, K.; Takasaki, S.; Sakai-Tagawa, Y.; Lopes, T.J.S.; et al. A humanized MDCK cell line for the efficient isolation and propagation of human influenza viruses. Nat. Microbiol. 2019, 4, 1268–1273. [Google Scholar] [CrossRef]
- Takashita, E.; Morita, H.; Ogawa, R.; Nakamura, K.; Fujisaki, S.; Shirakura, M.; Kuwahara, T.; Kishida, N.; Watanabe, S.; Odagiri, T. Susceptibility of Influenza Viruses to the Novel Cap-Dependent Endonuclease Inhibitor Baloxavir Marboxil. Front. Microbiol. 2018, 9, 3026. [Google Scholar] [CrossRef] [Green Version]
- Tilmanis, D.; van Baalen, C.; Oh, D.Y.; Rossignol, J.-F.; Hurt, A.C. The susceptibility of circulating human influenza viruses to tizoxanide, the active metabolite of nitazoxanide. Antivir. Res. 2017, 147, 142–148. [Google Scholar] [CrossRef]
- Takashita, E.; Kinoshita, N.; Yamayoshi, S.; Sakai-Tagawa, Y.; Fujisaki, S.; Ito, M.; Iwatsuki-Horimoto, K.; Chiba, S.; Halfmann, P.; Nagai, H.; et al. Efficacy of Antibodies and Antiviral Drugs against COVID-19 Omicron Variant. N. Engl. J. Med. 2022, 386, 995–998. [Google Scholar] [CrossRef]
- Takashita, E.; Kinoshita, N.; Yamayoshi, S.; Sakai-Tagawa, Y.; Fujisaki, S.; Ito, M.; Iwatsuki-Horimoto, K.; Halfmann, P.; Watanabe, S.; Maeda, K.; et al. Efficacy of Antiviral Agents against the SARS-CoV-2 Omicron Subvariant BA.2. N. Engl. J. Med. 2022, 386, 1475–1477. [Google Scholar] [CrossRef]
- Takashita, E.; Yamayoshi, S.; Fukushi, S.; Suzuki, T.; Maeda, K.; Sakai-Tagawa, Y.; Ito, M.; Uraki, R.; Halfmann, P.; Watanabe, S.; et al. Efficacy of Antiviral Agents against the Omicron Subvariant BA.2.75. N. Engl. J. Med. 2022, 387, 1236–1238. [Google Scholar] [CrossRef]
- Takashita, E.; Yamayoshi, S.; Simon, V.; van Bakel, H.; Sordillo, E.M.; Pekosz, A.; Fukushi, S.; Suzuki, T.; Maeda, K.; Halfmann, P.; et al. Efficacy of Antibodies and Antiviral Drugs against Omicron BA.2.12.1, BA.4, and BA.5 Subvariants. N. Engl. J. Med. 2022, 387, 468–470. [Google Scholar] [CrossRef]
- Takashita, E.; Yamayoshi, S.; Halfmann, P.; Wilson, N.; Ries, H.; Richardson, A.; Bobholz, M.; Vuyk, W.; Maddox, R.; Baker, D.A.; et al. In Vitro Efficacy of Antiviral Agents against Omicron Subvariant BA.4.6. N. Engl. J. Med. 2022, 387, 2094–2097. [Google Scholar] [CrossRef]
- Hause, B.M.; Ducatez, M.; Collin, E.A.; Ran, Z.; Liu, R.; Sheng, Z.; Armien, A.; Kaplan, B.; Chakravarty, S.; Hoppe, A.D.; et al. Isolation of a Novel Swine Influenza Virus from Oklahoma in 2011 Which Is Distantly Related to Human Influenza C Viruses. PLoS Pathog. 2013, 9, e1003176. [Google Scholar] [CrossRef] [Green Version]
- Murakami, S.; Sato, R.; Ishida, H.; Katayama, M.; Takenaka-Uema, A.; Horimoto, T. Influenza D Virus of New Phylogenetic Lineage, Japan. Emerg. Infect. Dis. 2020, 26, 168–171. [Google Scholar] [CrossRef] [Green Version]
- Takashita, E.; Kawakami, C.; Morita, H.; Ogawa, R.; Fujisaki, S.; Shirakura, M.; Miura, H.; Nakamura, K.; Kishida, N.; Kuwahara, T.; et al. Detection of influenza A(H3N2) viruses exhibiting reduced susceptibility to the novel cap-dependent endonuclease inhibitor baloxavir in Japan, December 2018. Eurosurveillance 2019, 24, 1800698. [Google Scholar] [CrossRef] [Green Version]
- Takashita, E.; Abe, T.; Morita, H.; Nagata, S.; Fujisaki, S.; Miura, H.; Shirakura, M.; Kishida, N.; Nakamura, K.; Kuwahara, T.; et al. Influenza A(H1N1)pdm09 virus exhibiting reduced susceptibility to baloxavir due to a PA E23K substitution detected from a child without baloxavir treatment. Antivir. Res. 2020, 180, 104828. [Google Scholar] [CrossRef]
- Sugawara, K.; Nishimura, H.; Hongo, S.; Kitame, F.; Nakamura, K. Antigenic Characterization of the Nucleoprotein and Matrix Protein of Influenza C Virus with Monoclonal Antibodies. J. Gen. Virol. 1991, 72, 103–109. [Google Scholar] [CrossRef]
- Ishida, H.; Murakami, S.; Kamiki, H.; Matsugo, H.; Takenaka-Uema, A.; Horimoto, T. Establishment of a Reverse Genetics System for Influenza D Virus. J. Virol. 2020, 94, e01767-19. [Google Scholar] [CrossRef]
- Zhang, Y.; Aevermann, B.D.; Anderson, T.K.; Burke, D.F.; Dauphin, G.; Gu, Z.; He, S.; Kumar, S.; Larsen, C.N.; Lee, A.J.; et al. Influenza Research Database: An integrated bioinformatics resource for influenza virus research. Nucleic Acids Res. 2016, 45, D466–D474. [Google Scholar] [CrossRef] [Green Version]
- Noshi, T.; Kitano, M.; Taniguchi, K.; Yamamoto, A.; Omoto, S.; Baba, K.; Hashimoto, T.; Ishida, K.; Kushima, Y.; Hattori, K.; et al. In vitro characterization of baloxavir acid, a first-in-class cap-dependent endonuclease inhibitor of the influenza virus polymerase PA subunit. Antivir. Res. 2018, 160, 109–117. [Google Scholar] [CrossRef]
- Uehara, T.; Hayden, F.G.; Kawaguchi, K.; Omoto, S.; Hurt, A.C.; De Jong, M.D.; Hirotsu, N.; Sugaya, N.; Lee, N.; Baba, K.; et al. Treatment-Emergent Influenza Variant Viruses With Reduced Baloxavir Susceptibility: Impact on Clinical and Virologic Outcomes in Uncomplicated Influenza. J. Infect. Dis. 2019, 221, 346–355. [Google Scholar] [CrossRef]
- Hirotsu, N.; Sakaguchi, H.; Sato, C.; Ishibashi, T.; Baba, K.; Omoto, S.; Shishido, T.; Tsuchiya, K.; Hayden, F.G.; Uehara, T.; et al. Baloxavir Marboxil in Japanese Pediatric Patients With Influenza: Safety and Clinical and Virologic Outcomes. Clin. Infect. Dis. 2019, 71, 971–981. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayden, F.G.; Sugaya, N.; Hirotsu, N.; Lee, N.; De Jong, M.D.; Hurt, A.C.; Ishida, T.; Sekino, H.; Yamada, K.; Portsmouth, S.; et al. Baloxavir Marboxil for Uncomplicated Influenza in Adults and Adolescents. N. Engl. J. Med. 2018, 379, 913–923. [Google Scholar] [CrossRef] [PubMed]
- Goldhill, D.H.; te Velthuis, A.J.W.; Fletcher, R.A.; Langat, P.; Zambon, M.; Lackenby, A.; Barclay, W.S. The mechanism of resistance to favipiravir in influenza. Proc. Natl. Acad. Sci. USA 2018, 115, 11613–11618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asha, K.; Kumar, B. Emerging Influenza D Virus Threat: What We Know so Far! J. Clin. Med. 2019, 8, 192. [Google Scholar] [CrossRef] [Green Version]
- Taylor, R.M. Studies on Survival of Influenza Virus Between Epidemics and Antigenic Variants of the Virus. Am. J. Public Health Nations Health 1949, 39, 171–178. [Google Scholar] [CrossRef]
- Sreenivasan, C.C.; Sheng, Z.; Wang, D.; Li, F. Host Range, Biology, and Species Specificity of Seven-Segmented Influenza Viruses—A Comparative Review on Influenza C and D. Pathogens 2021, 10, 1583. [Google Scholar] [CrossRef]
- Matsuzaki, Y.; Katsushima, N.; Nagai, Y.; Shoji, M.; Itagaki, T.; Sakamoto, M.; Kitaoka, S.; Mizuta, K.; Nishimura, H. Clinical Features of Influenza C Virus Infection in Children. J. Infect. Dis. 2006, 193, 1229–1235. [Google Scholar] [CrossRef]
- Odagiri, T.; Matsuzaki, Y.; Okamoto, M.; Suzuki, A.; Saito, M.; Tamaki, R.; Lupisan, S.P.; Sombrero, L.T.; Hongo, S.; Oshitani, H. Isolation and Characterization of Influenza C Viruses in the Philippines and Japan. J. Clin. Microbiol. 2015, 53, 847–858. [Google Scholar] [CrossRef] [Green Version]
- Shimizu, Y.; Abiko, C.; Ikeda, T.; Mizuta, K.; Matsuzaki, Y. Influenza C Virus and Human Metapneumovirus Infections in Hospitalized Children With Lower Respiratory Tract Illness. Pediatr. Infect. Dis. J. 2015, 34, 1273–1275. [Google Scholar] [CrossRef]
- Gouarin, S.; Vabret, A.; Dina, J.; Petitjean, J.; Brouard, J.; Cuvillon-Nimal, D.; Freymuth, F. Study of influenza C virus infection in France. J. Med. Virol. 2008, 80, 1441–1446. [Google Scholar] [CrossRef]
- Thielen, B.K.; Friedlander, H.; Bistodeau, S.; Shu, B.; Lynch, B.; Martin, K.; Bye, E.; Como-Sabetti, K.; Boxrud, D.; Strain, A.K.; et al. Detection of Influenza C Viruses Among Outpatients and Patients Hospitalized for Severe Acute Respiratory Infection, Minnesota, 2013–2016. Clin. Infect. Dis. 2017, 66, 1092–1098. [Google Scholar] [CrossRef] [PubMed]
- Ducatez, M.F.; Pelletier, C.; Meyer, G. Influenza D Virus in Cattle, France, 2011–2014. Emerg. Infect. Dis. 2015, 21, 368–371. [Google Scholar] [CrossRef] [PubMed]
- Chiapponi, C.; Faccini, S.; De Mattia, A.; Baioni, L.; Barbieri, I.; Rosignoli, C.; Nigrelli, A.; Foni, E. Detection of Influenza D Virus among Swine and Cattle, Italy. Emerg. Infect. Dis. 2016, 22, 352–354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mitra, N.; Cernicchiaro, N.; Torres, S.; Li, F.; Hause, B.M. Metagenomic characterization of the virome associated with bovine respiratory disease in feedlot cattle identified novel viruses and suggests an etiologic role for influenza D virus. J. Gen. Virol. 2016, 97, 1771–1784. [Google Scholar] [CrossRef] [PubMed]
- Horimoto, T.; Hiono, T.; Mekata, H.; Odagiri, T.; Lei, Z.; Kobayashi, T.; Norimine, J.; Inoshima, Y.; Hikono, H.; Murakami, K.; et al. Nationwide Distribution of Bovine Influenza D Virus Infection in Japan. PLoS ONE 2016, 11, e0163828. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhai, S.L.; Zhang, H.; Chen, S.N.; Zhou, X.; Lin, T.; Liu, R.; Lv, D.H.; Wen, X.H.; Wei, W.K.; Wang, D.; et al. Influenza D Virus in Animal Species in Guangdong Province, Southern China. Emerg. Infect. Dis. 2017, 23, 1392–1396. [Google Scholar] [CrossRef] [Green Version]
- Flynn, O.; Gallagher, C.; Mooney, J.; Irvine, C.; Ducatez, M.; Hause, B.; McGrath, G.; Ryan, E. Influenza D Virus in Cattle, Ireland. Emerg. Infect. Dis. 2018, 24, 389–391. [Google Scholar] [CrossRef] [Green Version]
- Song, H.; Qi, J.; Khedri, Z.; Diaz, S.; Yu, H.; Chen, X.; Varki, A.; Shi, Y.; Gao, G.F. Correction: An Open Receptor-Binding Cavity of Hemagglutinin-Esterase-Fusion Glycoprotein from Newly-Identified Influenza D Virus: Basis for Its Broad Cell Tropism. PLoS Pathog. 2016, 12, e1005505. [Google Scholar] [CrossRef] [Green Version]
- Holwerda, M.; Kelly, J.; Laloli, L.; Stürmer, I.; Portmann, J.; Stalder, H.; Dijkman, R. Determining the Replication Kinetics and Cellular Tropism of Influenza D Virus on Primary Well-Differentiated Human Airway Epithelial Cells. Viruses 2019, 11, 377. [Google Scholar] [CrossRef] [Green Version]
- Choi, J.Y.; Zemke, J.; Philo, S.E.; Bailey, E.S.; Yondon, M.; Gray, G.C. Aerosol Sampling in a Hospital Emergency Room Setting: A Complementary Surveillance Method for the Detection of Respiratory Viruses. Front. Public Health 2018, 6, 174. [Google Scholar] [CrossRef]
- Bailey, E.S.; Choi, J.Y.; Zemke, J.; Yondon, M.; Gray, G.C. Molecular surveillance of respiratory viruses with bioaerosol sampling in an airport. Trop. Dis. Travel Med. Vaccines 2018, 4, 11. [Google Scholar] [CrossRef]
- Borkenhagen, L.K.; Mallinson, K.A.; Tsao, R.W.; Ha, S.-J.; Lim, W.-H.; Toh, T.-H.; Anderson, B.D.; Fieldhouse, J.; Philo, S.E.; Chong, K.-S.; et al. Surveillance for respiratory and diarrheal pathogens at the human-pig interface in Sarawak, Malaysia. PLoS ONE 2018, 13, e0201295. [Google Scholar] [CrossRef]
- White, S.K.; Ma, W.; McDaniel, C.J.; Gray, G.C.; Lednicky, J.A. Serologic evidence of exposure to influenza D virus among persons with occupational contact with cattle. J. Clin. Virol. 2016, 81, 31–33. [Google Scholar] [CrossRef]
- Yesilbag, K.; Toker, E.B.; Ates, O. Recent strains of influenza D virus create a new genetic cluster for European strains. Microb. Pathog. 2022, 172, 105769. [Google Scholar] [CrossRef]


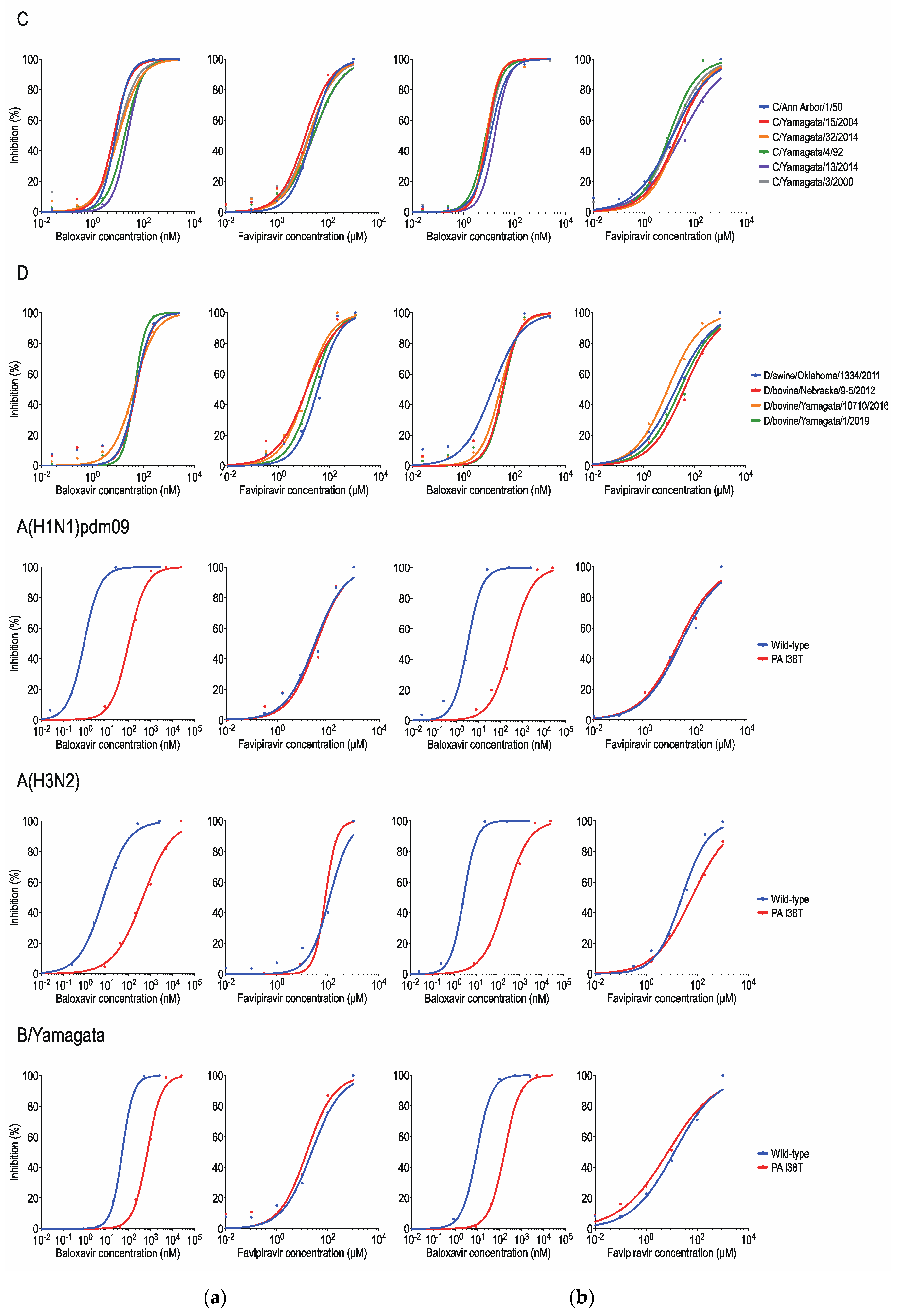{kind=link}
| Type | Subtype/Lineage | Virus | Mean IC50 ± SD * | |||
|---|---|---|---|---|---|---|
| MDCK Cells | ST Cells | |||||
| Baloxavir (nM) | Favipiravir (μM) | Baloxavir (nM) | Favipiravir (μM) | |||
| C | C/Taylor | C/Ann Arbor/1/50 | 7.73 ± 2.67 | 22.37 ± 3.58 | 11.29 ± 0.58 | 20.07 ± 10.00 |
| C/Yamagata | C/Yamagata/15/2004 | 6.20 ± 0.92 | 13.20 ± 1.15 | 8.87 ± 1.90 | 21.45 ± 3.46 | |
| C/Sao Paulo | C/Yamagata/32/2014 | 8.49 ± 3.75 | 19.44 ± 3.45 | 11.45 ± 1.26 | 16.16 ± 5.18 | |
| C/Aichi | C/Yamagata/4/92 | 19.74 ± 3.39 | 24.72 ± 1.38 | 8.19 ± 1.87 | 16.86 ± 5.63 | |
| C/Kanagawa | C/Yamagata/13/2014 | 13.28 ± 3.89 | 17.54 ± 2.41 | 9.35 ± 2.41 | 58.34 ± 14.33 | |
| C/Mississippi | C/Yamagata/3/2000 | 24.83 ± 9.48 | 28.62 ± 4.85 | 17.48 ± 6.00 | 22.22 ± 6.47 | |
| D | D/OK | D/swine/Oklahoma/1334/2011 | 49.83 ± 5.02 | 35.83 ± 12.83 | 13.91 ± 1.48 | 20.81 ± 1.27 |
| D/660 | D/bovine/Nebraska/9-5/2012 | 51.02 ± 12.11 | 13.68 ± 3.55 | 34.76 ± 1.94 | 46.86 ± 8.63 | |
| D/Yama2016 | D/bovine/Yamagata/10710/2016 | 44.51 ± 9.12 | 12.48 ± 2.71 | 27.40 ± 3.22 | 10.21 ± 1.47 | |
| D/Yama2019 | D/bovine/Yamagata/1/2019 | 46.36 ± 6.47 | 21.05 ± 1.49 | 36.75 ± 5.86 | 33.51 ± 4.52 | |
| A | A(H1N1)pdm09 | A/Kanagawa/ZC1931/2019 (wild type) | 0.88 ± 0.12 | 31.07 ± 7.86 | 3.09 ± 0.39 | 57.96 ± 8.90 |
| A/Kanagawa/IC1890/2019 (PA I38T) † | 98.19 ± 10.33 | 35.07 ± 6.34 | 329.50 ± 48.38 | 40.65 ± 15.85 | ||
| A(H3N2) | A/Yokohama/136/2018 (wild type) | 6.70 ± 0.65 | 136.20 ± 41.54 | 2.65 ± 0.20 | 30.38 ± 7.83 | |
| A/Yokohama/133/2018 (PA I38T) † | 568.03 ± 251.22 | 73.87 ± 9.86 | 240.80 ± 71.37 | 59.65 ± 7.27 | ||
| B | B/Yamagata | RG-B/Yamanashi/166/1998 (wild type) | 46.79 ± 2.93 | 23.29 ± 1.10 | 9.09 ± 2.88 | 19.44 ± 2.89 |
| RG-B/Yamanashi/166/1998 (PA I38T) † | 680.97 ± 21.18 | 15.56 ± 5.54 | 169.30 ± 32.24 | 34.00 ± 5.56 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Takashita, E.; Murakami, S.; Matsuzaki, Y.; Fujisaki, S.; Morita, H.; Nagata, S.; Katayama, M.; Mizuta, K.; Nishimura, H.; Watanabe, S.; et al. Antiviral Susceptibilities of Distinct Lineages of Influenza C and D Viruses. Viruses 2023, 15, 244. https://doi.org/10.3390/v15010244
Takashita E, Murakami S, Matsuzaki Y, Fujisaki S, Morita H, Nagata S, Katayama M, Mizuta K, Nishimura H, Watanabe S, et al. Antiviral Susceptibilities of Distinct Lineages of Influenza C and D Viruses. Viruses. 2023; 15(1):244. https://doi.org/10.3390/v15010244
Chicago/Turabian StyleTakashita, Emi, Shin Murakami, Yoko Matsuzaki, Seiichiro Fujisaki, Hiroko Morita, Shiho Nagata, Misa Katayama, Katsumi Mizuta, Hidekazu Nishimura, Shinji Watanabe, and et al. 2023. "Antiviral Susceptibilities of Distinct Lineages of Influenza C and D Viruses" Viruses 15, no. 1: 244. https://doi.org/10.3390/v15010244