
Detection and Molecular Characterization of the SARS-CoV-2 Delta Variant and the Specific Immune Response in Companion Animals in Switzerland
 , , , , , , , , ,
, , , , , , , , ,  , , and
, , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design and Recruitment of COVID-19-Affected Households
2.2. Swab and Blood Sample Collection
2.3. SARS-CoV-2 Nucleic Acid Extraction, Analysis, and Confirmation
2.4. Next-Generation Sequencing (NGS) and Phylogenetic Tree Construction
2.5. Serological SARS-CoV-2 Testing and Variant Analysis
3. Results
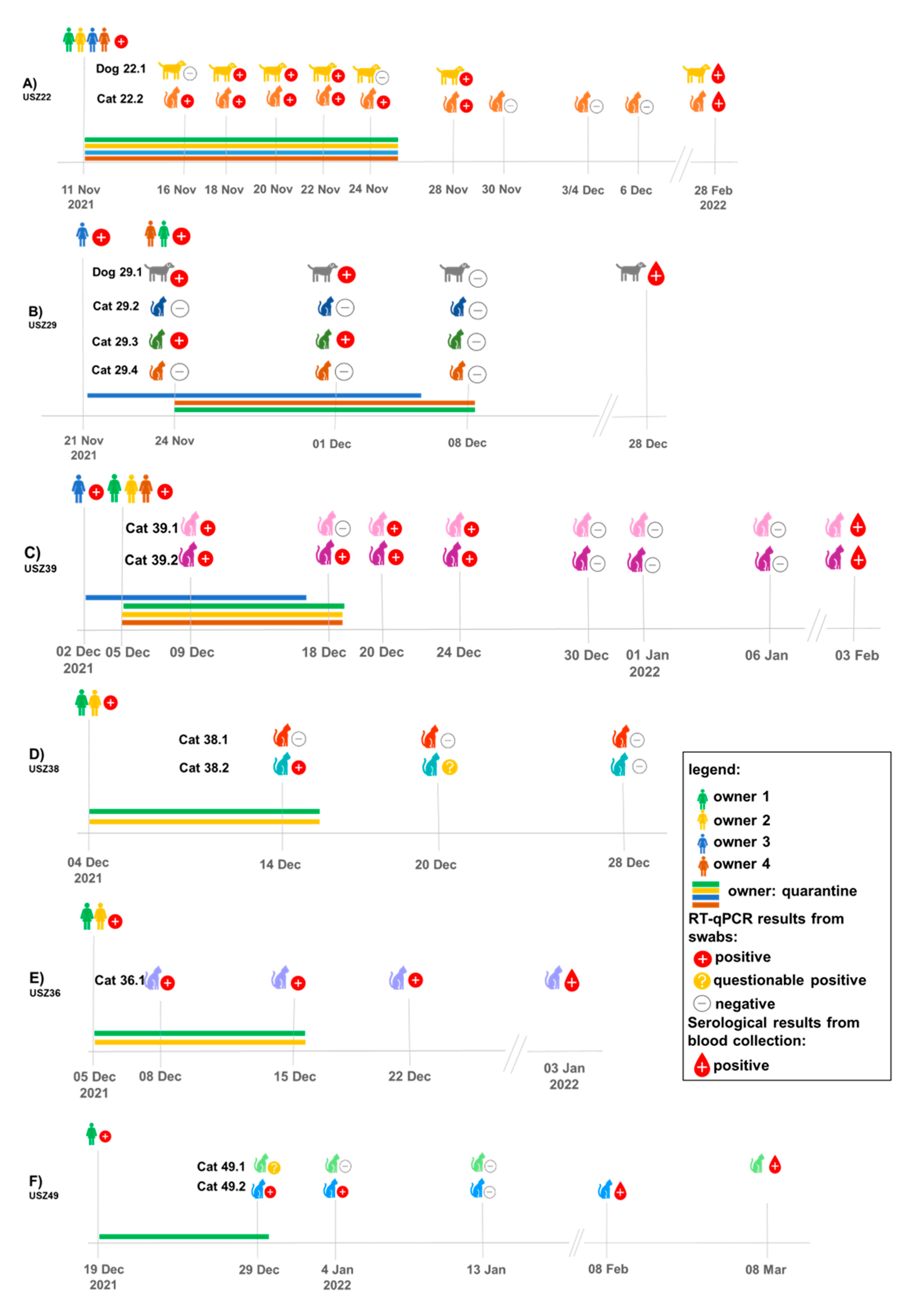
3.1. Overview of COVID-19-Affected Households Included in This Study, Characterization of Cats and Dogs and Available Material, and Test Results
3.2. Timeline of Sample Collection and Molecular and Serological Testing for SARS-CoV-2 in the Different Households
3.3. Detection of SARS-CoV-2 Viral RNA and RNA Loads in Different Samples from Companion Animals and the Environment (Fur, Bedding)
3.4. Confirmation of SARS-CoV-2 Infection in Cats and Dogs by Serology
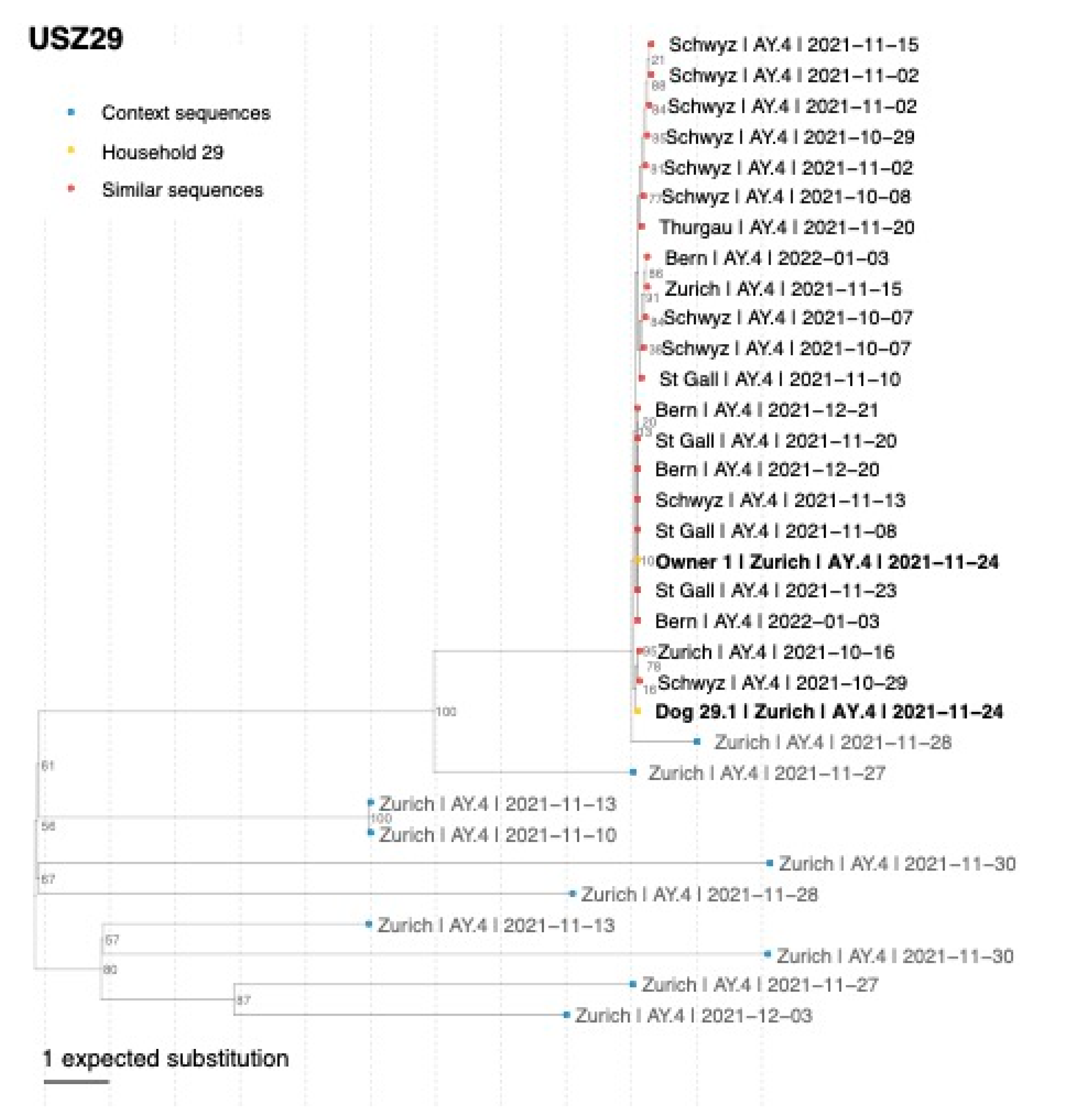
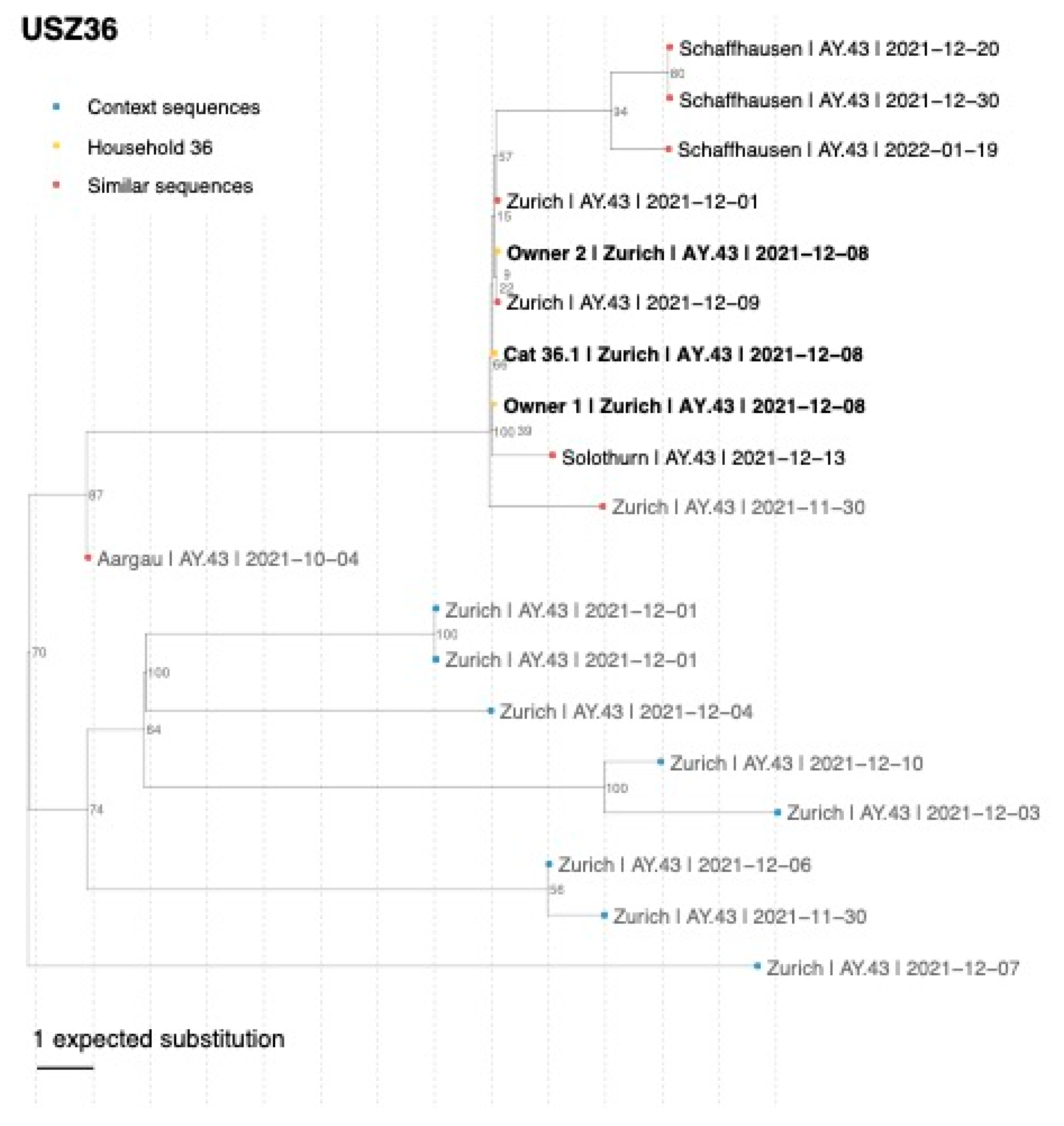
3.5. Confirmation of SARS-CoV-2 Delta Variant Infection and Determination of the Pangolin Lineage in Cats, Dogs, and Owners by NGS
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SNP | Aa Mutation | Samples | % Human Sequences GISAID | % Animal Sequences GISAID | % Companion Animal Sequences GISAID | |
|---|---|---|---|---|---|---|
| 1 | A10323G | ORF1a:K3353R | 22.2, 22.1 | 1.41 (1.4, 1.42) | 0.93 (0.61, 1.42) | 2.62 (1.28, 5.31) |
| 2 | A929G | ORF1a:I222V | 36.1 | 0.06 (0.06, 0.06) | 0.13 (0.05, 0.39) | 0.75 (0.21, 2.69) |
| 3 | C15952A | ORF1b:L829I | 39.1, 39.2 | 2.36 (2.35, 2.36) | 0.44 (0.24, 0.81) | 0.75 (0.21, 2.69) |
| 4 | C20844T | - | 36.1 | 0.17 (0.17, 0.17) | 0.88 (0.57, 1.36) | 1.12 (0.38, 3.25) |
| 5 | C22792T | - | 39.1, 39.2 | 5.66 (5.65, 5.68) | 0.97 (0.64, 1.47) | 1.12 (0.38, 3.25) |
| 6 | C6026T | ORF1a:P1921S | 29.1 | 0.15 (0.15, 0.15) | 0.13 (0.05, 0.39) | 1.12 (0.38, 3.25) |
| 7 | C9286T | - | 39.1, 39.2 | 0.2 (0.2, 0.2) | 0.97 (0.64, 1.47) | 1.87 (0.8, 4.31) |
| 8 | G2272A | - | 29.1 | 0.03 (0.03, 0.04) | 0.13 (0.05, 0.39) | 0.37 (0.02, 2.09) |
| 9 | G25244T | S:V1228L | 29.1 | 0.12 (0.12, 0.12) | 0.44 (0.24, 0.81) | 1.12 (0.38, 3.25) |
| 10 | G487A | - | 36.1 | 0.04 (0.04, 0.04) | 0.13 (0.05, 0.39) | 0.75 (0.21, 2.69) |
| 11 | T27534C | - | 38.2 | 0.03 (0.02, 0.03) | 0.35 (0.18, 0.7) | 1.5 (0.58, 3.79) |
| 12 | T29867A | - | 36.1, 38.2, 39.1 | 0.12 (0.12, 0.13) | 0.18 (0.07, 0.45) | 1.12 (0.38, 3.25) |
References
- World Health Organization. WHO Coronavirus (COVID-19) Dashboard. Available online: https://covid19.who.int/ (accessed on 17 November 2022).
- Zhu, Z.; Lian, X.; Su, X.; Wu, W.; Marraro, G.A.; Zeng, Y. From SARS and MERS to COVID-19: A brief summary and comparison of severe acute respiratory infections caused by three highly pathogenic human coronaviruses. Respir. Res. 2020, 21, 224. [Google Scholar] [CrossRef]
- Zhou, P.; Yang, X.L.; Wang, X.G.; Hu, B.; Zhang, L.; Zhang, W.; Si, H.R.; Zhu, Y.; Li, B.; Huang, C.L.; et al. A pneumonia outbreak associated with a new coronavirus of probable bat origin. Nature 2020, 588, E6. [Google Scholar] [CrossRef]
- Lam, T.T.; Jia, N.; Zhang, Y.W.; Shum, M.H.; Jiang, J.F.; Zhu, H.C.; Tong, Y.G.; Shi, Y.X.; Ni, X.B.; Liao, Y.S.; et al. Identifying SARS-CoV-2-related coronaviruses in Malayan pangolins. Nature 2020, 583, 282–285. [Google Scholar] [CrossRef] [Green Version]
- Segales, J.; Puig, M.; Rodon, J.; Avila-Nieto, C.; Carrillo, J.; Cantero, G.; Terron, M.T.; Cruz, S.; Parera, M.; Noguera-Julian, M.; et al. Detection of SARS-CoV-2 in a cat owned by a COVID-19-affected patient in Spain. Proc. Natl. Acad. Sci. USA 2020, 117, 24790–24793. [Google Scholar] [CrossRef]
- Klaus, J.; Meli, M.L.; Willi, B.; Nadeau, S.; Beisel, C.; Stadler, T.; Egberink, H.; Zhao, S.; Lutz, H.; Riond, B.; et al. Detection and Genome Sequencing of SARS-CoV-2 in a Domestic Cat with Respiratory Signs in Switzerland. Viruses 2021, 13, 496. [Google Scholar] [CrossRef] [PubMed]
- Neira, V.; Brito, B.; Aguero, B.; Berrios, F.; Valdes, V.; Gutierrez, A.; Ariyama, N.; Espinoza, P.; Retamal, P.; Holmes, E.C.; et al. A household case evidences shorter shedding of SARS-CoV-2 in naturally infected cats compared to their human owners. Emerg. Microbes Infect. 2021, 10, 376–383. [Google Scholar] [CrossRef] [PubMed]
- Hosie, M.J.; Epifano, I.; Herder, V.; Orton, R.J.; Stevenson, A.; Johnson, N.; MacDonald, E.; Dunbar, D.; McDonald, M.; Howie, F.; et al. Detection of SARS-CoV-2 in respiratory samples from cats in the UK associated with human-to-cat transmission. Vet. Rec. 2021, 188, e247. [Google Scholar] [CrossRef] [PubMed]
- Garigliany, M.; Van Laere, A.S.; Clercx, C.; Giet, D.; Escriou, N.; Huon, C.; van der Werf, S.; Eloit, M.; Desmecht, D. SARS-CoV-2 Natural Transmission from Human to Cat, Belgium, March 2020. Emerg. Infect. Dis. 2020, 26, 3069–3071. [Google Scholar] [CrossRef]
- Newman, A.; Smith, D.; Ghai, R.R.; Wallace, R.M.; Torchetti, M.K.; Loiacono, C.; Murrell, L.S.; Carpenter, A.; Moroff, S.; Rooney, J.A.; et al. First Reported Cases of SARS-CoV-2 Infection in Companion Animals—New York, March-April 2020. MMWR Morb. Mortal. Wkly. Rep. 2020, 69, 710–713. [Google Scholar] [CrossRef]
- Fuentealba, N.A.; More, G.; Bravi, M.E.; Unzaga, J.M.; De Felice, L.; Salina, M.; Viegas, M.; Nabaes Jodar, M.S.; Valinotto, L.E.; Rivero, F.D.; et al. First detection and molecular analysis of SARS-CoV-2 from a naturally infected cat from Argentina. Vet. Microbiol. 2021, 260, 109179. [Google Scholar] [CrossRef]
- Ruiz-Arrondo, I.; Portillo, A.; Palomar, A.M.; Santibanez, S.; Santibanez, P.; Cervera, C.; Oteo, J.A. Detection of SARS-CoV-2 in pets living with COVID-19 owners diagnosed during the COVID-19 lockdown in Spain: A case of an asymptomatic cat with SARS-CoV-2 in Europe. Transbound. Emerg. Dis. 2020, 68, 973–976. [Google Scholar] [CrossRef]
- Van Aart, A.E.; Velkers, F.C.; Fischer, E.A.J.; Broens, E.M.; Egberink, H.; Zhao, S.; Engelsma, M.; Hakze-van der Honing, R.W.; Harders, F.; de Rooij, M.M.T.; et al. SARS-CoV-2 infection in cats and dogs in infected mink farms. Transbound. Emerg. Dis. 2021, 69, 3001–3007. [Google Scholar] [CrossRef] [PubMed]
- Chandler, J.C.; Bevins, S.N.; Ellis, J.W.; Linder, T.J.; Tell, R.M.; Jenkins-Moore, M.; Root, J.J.; Lenoch, J.B.; Robbe-Austerman, S.; Deliberto, T.J.; et al. SARS-CoV-2 exposure in wild white-tailed deer (Odocoileus virginianus). Proc. Natl. Acad. Sci. USA 2021, 118, e2114828118. [Google Scholar] [CrossRef]
- Kuchipudi, S.V.; Surendran-Nair, M.; Ruden, R.M.; Yon, M.; Nissly, R.H.; Vandegrift, K.J.; Nelli, R.K.; Li, L.; Jayarao, B.M.; Maranas, C.D.; et al. Multiple spillovers from humans and onward transmission of SARS-CoV-2 in white-tailed deer. Proc. Natl. Acad. Sci. USA 2022, 119, e2121644119. [Google Scholar] [CrossRef]
- World Health Organization. WHO Director-General’s Opening Remarks at the Media Briefing on COVID-19—11 March 2020. Available online: https://www.who.int/director-general/speeches/detail/who-director-general-s-opening-remarks-at-the-media-briefing-on-covid-19 (accessed on 11 March 2020).
- World Health Organization. Tracking SARS-CoV-2 Variants. Available online: https://www.who.int/activities/tracking-SARS-CoV-2-variants (accessed on 21 September 2021).
- World Organisation for Animal Health Situation Report #19 on SARS-CoV-2 in Animals. Available online: https://www.woah.org/app/uploads/2022/12/sars-cov-2-situation-report-19.pdf (accessed on 30 November 2022).
- Sit, T.H.C.; Brackman, C.J.; Ip, S.M.; Tam, K.W.S.; Law, P.Y.T.; To, E.M.W.; Yu, V.Y.T.; Sims, L.D.; Tsang, D.N.C.; Chu, D.K.W.; et al. Infection of dogs with SARS-CoV-2. Nature 2020, 586, 776–778. [Google Scholar] [CrossRef] [PubMed]
- Klaus, J.; Zini, E.; Hartmann, K.; Egberink, H.; Kipar, A.; Bergmann, M.; Palizzotto, C.; Zhao, S.; Rossi, F.; Franco, V.; et al. SARS-CoV-2 Infection in Dogs and Cats from Southern Germany and Northern Italy during the First Wave of the COVID-19 Pandemic. Viruses 2021, 13, 1453. [Google Scholar] [CrossRef]
- Fritz, M.; Rosolen, B.; Krafft, E.; Becquart, P.; Elguero, E.; Vratskikh, O.; Denolly, S.; Boson, B.; Vanhomwegen, J.; Gouilh, M.A.; et al. High prevalence of SARS-CoV-2 antibodies in pets from COVID-19+ households. One Health 2021, 11, 100192. [Google Scholar] [CrossRef] [PubMed]
- Goryoka, G.W.; Cossaboom, C.M.; Gharpure, R.; Dawson, P.; Tansey, C.; Rossow, J.; Mrotz, V.; Rooney, J.; Torchetti, M.; Loiacono, C.M.; et al. One Health Investigation of SARS-CoV-2 Infection and Seropositivity among Pets in Households with Confirmed Human COVID-19 Cases—Utah and Wisconsin, 2020. Viruses 2021, 13, 1813. [Google Scholar] [CrossRef]
- Hamer, S.A.; Pauvolid-Correa, A.; Zecca, I.B.; Davila, E.; Auckland, L.D.; Roundy, C.M.; Tang, W.; Torchetti, M.; Killian, M.L.; Jenkins-Moore, M.; et al. Natural SARS-CoV-2 infections, including virus isolation, among serially tested cats and dogs in households with confirmed human COVID-19 cases in Texas, USA. BioRxiv, 2020; preprint. [Google Scholar]
- Barroso-Arevalo, S.; Rivera, B.; Dominguez, L.; Sanchez-Vizcaino, J.M. First Detection of SARS-CoV-2 B.1.1.7 Variant of Concern in an Asymptomatic Dog in Spain. Viruses 2021, 13, 1379. [Google Scholar] [CrossRef]
- Barroso-Arevalo, S.; Sanchez-Morales, L.; Perez-Sancho, M.; Dominguez, L.; Sanchez-Vizcaino, J.M. First Detection of SARS-CoV-2 B.1.617.2 (Delta) Variant of Concern in a Symptomatic Cat in Spain. Front. Vet. Sci. 2022, 9, 841430. [Google Scholar] [CrossRef]
- Barroso, R.; Vieira-Pires, A.; Antunes, A.; Fidalgo-Carvalho, I. Susceptibility of Pets to SARS-CoV-2 Infection: Lessons from a Seroepidemiologic Survey of Cats and Dogs in Portugal. Microorganisms 2022, 10, 345. [Google Scholar] [CrossRef] [PubMed]
- Pomorska-Mol, M.; Turlewicz-Podbielska, H.; Gogulski, M.; Ruszkowski, J.J.; Kubiak, M.; Kuriga, A.; Barket, P.; Postrzech, M. A cross-sectional retrospective study of SARS-CoV-2 seroprevalence in domestic cats, dogs and rabbits in Poland. BMC Vet. Res. 2021, 17, 322. [Google Scholar] [CrossRef] [PubMed]
- Mohebali, M.; Hassanpour, G.; Zainali, M.; Gouya, M.M.; Khayatzadeh, S.; Parsaei, M.; Sarafraz, N.; Hassanzadeh, M.; Azarm, A.; Salehi-Vaziri, M.; et al. SARS-CoV-2 in domestic cats (Felis catus) in the northwest of Iran: Evidence for SARS-CoV-2 circulating between human and cats. Virus Res. 2022, 310, 198673. [Google Scholar] [CrossRef] [PubMed]
- Zoccola, R.; Beltramo, C.; Magris, G.; Peletto, S.; Acutis, P.; Bozzetta, E.; Radovic, S.; Zappulla, F.; Porzio, A.M.; Gennero, M.S.; et al. First detection of an Italian human-to-cat outbreak of SARS-CoV-2 Alpha variant—lineage B.1.1.7. One Health 2021, 13, 100295. [Google Scholar] [CrossRef]
- Pecora, A.; Malacari, D.A.; Mozgovoj, M.V.; Diaz, M.L.A.; Peralta, A.V.; Cacciabue, M.; Puebla, A.F.; Carusso, C.; Mundo, S.L.; Gonzalez Lopez Ledesma, M.M.; et al. Anthropogenic Infection of Domestic Cats With SARS-CoV-2 Alpha Variant B.1.1.7 Lineage in Buenos Aires. Front. Vet. Sci. 2022, 9, 790058. [Google Scholar] [CrossRef]
- Jairak, W.; Charoenkul, K.; Chamsai, E.; Udom, K.; Chaiyawong, S.; Bunpapong, N.; Boonyapisitsopa, S.; Tantilertcharoen, R.; Techakriengkrai, N.; Surachetpong, S.; et al. First cases of SARS-CoV-2 infection in dogs and cats in Thailand. Transbound. Emerg. Dis. 2021, 69, e979–e991. [Google Scholar] [CrossRef]
- Miro, G.; Regidor-Cerrillo, J.; Checa, R.; Diezma-Diaz, C.; Montoya, A.; Garcia-Cantalejo, J.; Botias, P.; Arroyo, J.; Ortega-Mora, L.M. SARS-CoV-2 Infection in One Cat and Three Dogs Living in COVID-19-Positive Households in Madrid, Spain. Front. Vet. Sci. 2021, 8, 779341. [Google Scholar] [CrossRef]
- Hamer, S.A.; Ghai, R.R.; Zecca, I.B.; Auckland, L.D.; Roundy, C.M.; Davila, E.; Busselman, R.E.; Tang, W.; Pauvolid-Correa, A.; Killian, M.L.; et al. SARS-CoV-2 B.1.1.7 variant of concern detected in a pet dog and cat after exposure to a person with COVID-19, USA. Transbound. Emerg. Dis. 2022, 69, 1656–1658. [Google Scholar] [CrossRef]
- Keller, M.; Hagag, I.T.; Balzer, J.; Beyer, K.; Kersebohm, J.C.; Sadeghi, B.; Wernike, K.; Hoper, D.; Wylezich, C.; Beer, M.; et al. Detection of SARS-CoV-2 variant B.1.1.7 in a cat in Germany. Res. Vet. Sci. 2021, 140, 229–232. [Google Scholar] [CrossRef]
- Ferasin, L.; Fritz, M.; Ferasin, H.; Becquart, P.; Corbet, S.; Ar Gouilh, M.; Legros, V.; Leroy, E.M. Infection with SARS-CoV-2 variant B.1.1.7 detected in a group of dogs and cats with suspected myocarditis. Vet. Rec. 2021, 189, e944. [Google Scholar] [CrossRef]
- Fernandez-Bastit, L.; Rodon, J.; Pradenas, E.; Marfil, S.; Trinite, B.; Parera, M.; Roca, N.; Pou, A.; Cantero, G.; Lorca-Oro, C.; et al. First Detection of SARS-CoV-2 Delta (B.1.617.2) Variant of Concern in a Dog with Clinical Signs in Spain. Viruses 2021, 13, 2526. [Google Scholar] [CrossRef] [PubMed]
- Wendling, N.M.; Carpenter, A.; Liew, A.; Ghai, R.R.; Gallardo-Romero, N.; Stoddard, R.A.; Tao, Y.; Zhang, J.; Retchless, A.C.; Ahmad, A.; et al. Transmission of SARS-CoV-2 Delta variant (B.1.617.2) from a fully vaccinated human to a canine in Georgia, July 2021. Zoonoses Public Health 2022, 69, 587–592. [Google Scholar] [CrossRef]
- Kang, K.; Chen, Q.; Gao, Y.; Yu, K.J. Detection of SARS-CoV-2 B.1.617.2 (Delta) variant in three cats owned by a confirmed COVID-19 patient in Harbin, China. Vet. Med. Sci. 2021, 8, 945. [Google Scholar] [CrossRef] [PubMed]
- Doerksen, T.; Lu, A.; Noll, L.; Almes, K.; Bai, J.; Upchurch, D.; Palinski, R. Near-Complete Genome of SARS-CoV-2 Delta (AY.3) Variant Identified in a Dog in Kansas, USA. Viruses 2021, 13, 2104. [Google Scholar] [CrossRef] [PubMed]
- Lenz, O.C.; Marques, A.D.; Kelly, B.J.; Rodino, K.G.; Cole, S.D.; Perera, R.A.P.M.; Weiss, S.R.; Bushman, F.D.; Lennon, E.M. SARS-CoV-2 Delta Variant (AY.3) in the Feces of a Domestic Cat. Viruses 2022, 14, 421. [Google Scholar] [CrossRef]
- Klaus, J.; Palizzotto, C.; Zini, E.; Meli, M.L.; Leo, C.; Egberink, H.; Zhao, S.; Hofmann-Lehmann, R. SARS-CoV-2 Infection and Antibody Response in a Symptomatic Cat from Italy with Intestinal B-Cell Lymphoma. Viruses 2021, 13, 527. [Google Scholar] [CrossRef] [PubMed]
- Studer, N.; Lutz, H.; Saegerman, C.; Gonczi, E.; Meli, M.L.; Boo, G.; Hartmann, K.; Hosie, M.J.; Moestl, K.; Tasker, S.; et al. Pan-European Study on the Prevalence of the Feline Leukaemia Virus Infection—Reported by the European Advisory Board on Cat Diseases (ABCD Europe). Viruses 2019, 11, 993. [Google Scholar] [CrossRef] [Green Version]
- Berger, A.; Willi, B.; Meli, M.L.; Boretti, F.S.; Hartnack, S.; Dreyfus, A.; Lutz, H.; Hofmann-Lehmann, R. Feline calicivirus and other respiratory pathogens in cats with Feline calicivirus-related symptoms and in clinically healthy cats in Switzerland. BMC Vet. Res. 2015, 11, 282. [Google Scholar] [CrossRef] [Green Version]
- Gomes-Keller, M.A.; Tandon, R.; Gonczi, E.; Meli, M.L.; Hofmann-Lehmann, R.; Lutz, H. Shedding of feline leukemia virus RNA in saliva is a consistent feature in viremic cats. Vet. Microbiol. 2006, 112, 11–21. [Google Scholar] [CrossRef]
- Nadeau, S.A.; Vaughan, T.G.; Beckmann, C.; Topolsky, I.; Chen, C.; Hodcroft, E.; Schär, T.; Nissen, I.; Santacroce, N.; Burcklen, E.; et al. Swiss public health measures associated with reduced SARS-CoV-2 transmission using genome data. Sci. Transl. Med. 2022, 14, eabn7979. [Google Scholar] [CrossRef] [PubMed]
- Posada-Cespedes, S.; Seifert, D.; Topolsky, I.; Jablonski, K.P.; Metzner, K.J.; Beerenwinkel, N. V-pipe: A computational pipeline for assessing viral genetic diversity from high-throughput data. Bioinformatics 2021, 37, 1673–1680. [Google Scholar] [CrossRef] [PubMed]
- Shu, Y.; McCauley, J. GISAID: Global initiative on sharing all influenza data—From vision to reality. Euro Surveill. 2017, 22, 30494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Toole, A.; Scher, E.; Underwood, A.; Jackson, B.; Hill, V.; McCrone, J.T.; Colquhoun, R.; Ruis, C.; Abu-Dahab, K.; Taylor, B.; et al. Assignment of epidemiological lineages in an emerging pandemic using the pangolin tool. Virus Evol. 2021, 7, veab064. [Google Scholar] [CrossRef]
- Hadfield, J.; Megill, C.; Bell, S.M.; Huddleston, J.; Potter, B.; Callender, C.; Sagulenko, P.; Bedford, T.; Neher, R.A. Nextstrain: Real-time tracking of pathogen evolution. Bioinformatics 2018, 34, 4121–4123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minh, B.Q.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; von Haeseler, A.; Lanfear, R. IQ-TREE 2: New Models and Efficient Methods for Phylogenetic Inference in the Genomic Era. Mol. Biol. Evol. 2020, 37, 1530–1534. [Google Scholar] [CrossRef] [Green Version]
- Hoang, D.T.; Chernomor, O.; von Haeseler, A.; Minh, B.Q.; Vinh, L.S. UFBoot2: Improving the Ultrafast Bootstrap Approximation. Mol. Biol. Evol. 2018, 35, 518–522. [Google Scholar] [CrossRef]
- Sagulenko, P.; Puller, V.; Neher, R.A. TreeTime: Maximum-likelihood phylodynamic analysis. Virus Evol. 2018, 4, vex042. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Nadeau, S.; Yared, M.; Voinov, P.; Xie, N.; Roemer, C.; Stadler, T. CoV-Spectrum: Analysis of Globally Shared SARS-CoV-2 Data to Identify and Characterize New Variants. Bioinformatics 2021, 38, 1735–1737. [Google Scholar] [CrossRef]
- Chaoran Chen, C.R. Tanja Stadler, LAPIS is a fast web API for massive open virus sequencing databases. arXiv 2022, arXiv:2206.01210. [Google Scholar]
- Spiri, A.M.; Meli, M.L.; Riond, B.; Herbert, I.; Hosie, M.J.; Hofmann-Lehmann, R. Environmental Contamination and Hygienic Measures After Feline Calicivirus Field Strain Infections of Cats in a Research Facility. Viruses 2019, 11, 958. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spiri, A.M.; Rodriguez-Campos, S.; Matos, J.M.; Glaus, T.M.; Riond, B.; Reusch, C.E.; Hofmann-Lehmann, R.; Willi, B. Clinical, serological and echocardiographic examination of healthy field dogs before and after vaccination with a commercial tetravalent leptospirosis vaccine. BMC Vet. Res. 2017, 13, 138. [Google Scholar] [CrossRef]
- Davis, C.; Logan, N.; Tyson, G.; Orton, R.; Harvey, W.T.; Perkins, J.S.; Mollett, G.; Blacow, R.M.; Consortium, C.-G.U.; Peacock, T.P.; et al. Reduced neutralisation of the Delta (B.1.617.2) SARS-CoV-2 variant of concern following vaccination. PLoS Pathog. 2021, 17, e1010022. [Google Scholar] [CrossRef] [PubMed]
- Tyson, G.B.; Jones, S.; Logan, N.; McDonald, M.; Murcia, P.R.; Willett, B.J.; Weir, W.; Hosie, M.J. Rising SARS-CoV-2 Seroprevalence and Patterns of Cross-Variant Antibody Neutralization in UK Domestic Cats. BioRxiv 2022. [Google Scholar] [CrossRef]
- Global Lineage Reports. Available online: https://cov-lineages.org/index.html (accessed on 21 September 2022).
- Pagani, G.; Lai, A.; Bergna, A.; Rizzo, A.; Stranieri, A.; Giordano, A.; Paltrinieri, S.; Lelli, D.; Decaro, N.; Rusconi, S.; et al. Human-to-Cat SARS-CoV-2 Transmission: Case Report and Full-Genome Sequencing from an Infected Pet and Its Owner in Northern Italy. Pathogens 2021, 10, 252. [Google Scholar] [CrossRef] [PubMed]
- Sailleau, C.; Dumarest, M.; Vanhomwegen, J.; Delaplace, M.; Caro, V.; Kwasiborski, A.; Hourdel, V.; Chevaillier, P.; Barbarino, A.; Comtet, L.; et al. First detection and genome sequencing of SARS-CoV-2 in an infected cat in France. Transbound. Emerg. Dis. 2020, 67, 2324–2328. [Google Scholar] [CrossRef]
- Bienzle, D.; Rousseau, J.; Marom, D.; MacNicol, J.; Jacobson, L.; Sparling, S.; Prystajecky, N.; Fraser, E.; Weese, J.S. Risk Factors for SARS-CoV-2 Infection and Illness in Cats and Dogs. Emerg. Infect. Dis. 2022, 28, 1154–1162. [Google Scholar] [CrossRef]
- Amicone, M.; Borges, V.; Alves, M.J.; Isidro, J.; Zé-Zé, L.; Duarte, S.; Vieira, L.; Guiomar, R.; Gomes, J.P.; Gordo, I. Mutation rate of SARS-CoV-2 and emergence of mutators during experimental evolution. Evol. Med. Public Health 2022, 10, 142–155. [Google Scholar] [CrossRef]
- Meredith, L.W.; Hamilton, W.L.; Warne, B.; Houldcroft, C.J.; Hosmillo, M.; Jahun, A.S.; Curran, M.D.; Parmar, S.; Caller, L.G.; Caddy, S.L.; et al. Rapid implementation of SARS-CoV-2 sequencing to investigate cases of health-care associated COVID-19: A prospective genomic surveillance study. Lancet Infect. Dis. 2020, 20, 1263–1271. [Google Scholar] [CrossRef]
- Lumley, S.F.; Constantinides, B.; Sanderson, N.; Rodger, G.; Street, T.L.; Swann, J.; Chau, K.K.; O’Donnell, D.; Warren, F.; Hoosdally, S.; et al. Epidemiological data and genome sequencing reveals that nosocomial transmission of SARS-CoV-2 is underestimated and mostly mediated by a small number of highly infectious individuals. J. Infect. 2021, 83, 473–482. [Google Scholar] [CrossRef]
- Rausch, J.W.; Capoferri, A.A.; Katusiime, M.G.; Patro, S.C.; Kearney, M.F. Low genetic diversity may be an Achilles heel of SARS-CoV-2. Proc. Natl. Acad. Sci. USA 2020, 117, 24614–24616. [Google Scholar] [CrossRef]
- Li, Y.; Si, H.R.; Zhu, Y.; Xie, N.; Li, B.; Zhang, X.P.; Han, J.F.; Bao, H.H.; Yang, Y.; Zhao, K.; et al. Characteristics of SARS-CoV-2 transmission in a medium-sized city with traditional communities during the early COVID-19 epidemic in China. Virol. Sin. 2022, 37, 187–197. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.I.; Kim, S.G.; Kim, S.M.; Kim, E.H.; Park, S.J.; Yu, K.M.; Chang, J.H.; Kim, E.J.; Lee, S.; Casel, M.A.B.; et al. Infection and Rapid Transmission of SARS-CoV-2 in Ferrets. Cell Host Microbe 2020, 27, 704–709 e2. [Google Scholar] [CrossRef]
- Richard, M.; Kok, A.; de Meulder, D.; Bestebroer, T.M.; Lamers, M.M.; Okba, N.M.A.; Fentener van Vlissingen, M.; Rockx, B.; Haagmans, B.L.; Koopmans, M.P.G.; et al. SARS-CoV-2 is transmitted via contact and via the air between ferrets. Nat. Commun. 2020, 11, 3496. [Google Scholar] [CrossRef]
- Shi, J.; Wen, Z.; Zhong, G.; Yang, H.; Wang, C.; Huang, B.; Liu, R.; He, X.; Shuai, L.; Sun, Z.; et al. Susceptibility of ferrets, cats, dogs, and other domesticated animals to SARS-coronavirus 2. Science 2020, 368, 1016–1020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bosco-Lauth, A.M.; Hartwig, A.E.; Porter, S.M.; Gordy, P.W.; Nehring, M.; Byas, A.D.; VandeWoude, S.; Ragan, I.K.; Maison, R.M.; Bowen, R.A. Experimental infection of domestic dogs and cats with SARS-CoV-2: Pathogenesis, transmission, and response to reexposure in cats. Proc. Natl. Acad. Sci. USA 2020, 117, 26382–26388. [Google Scholar] [CrossRef]
- Gaudreault, N.N.; Trujillo, J.D.; Carossino, M.; Meekins, D.A.; Morozov, I.; Madden, D.W.; Indran, S.V.; Bold, D.; Balaraman, V.; Kwon, T.; et al. SARS-CoV-2 infection, disease and transmission in domestic cats. Emerg. Microbes Infect. 2020, 9, 2322–2332. [Google Scholar] [CrossRef] [PubMed]
- Halfmann, P.J.; Hatta, M.; Chiba, S.; Maemura, T.; Fan, S.; Takeda, M.; Kinoshita, N.; Hattori, S.I.; Sakai-Tagawa, Y.; Iwatsuki-Horimoto, K.; et al. Transmission of SARS-CoV-2 in Domestic Cats. N. Engl. J. Med. 2020, 383, 592–594. [Google Scholar] [CrossRef]
- Sia, S.F.; Yan, L.M.; Chin, A.W.H.; Fung, K.; Choy, K.T.; Wong, A.Y.L.; Kaewpreedee, P.; Perera, R.; Poon, L.L.M.; Nicholls, J.M.; et al. Pathogenesis and transmission of SARS-CoV-2 in golden hamsters. Nature 2020, 583, 834–838. [Google Scholar] [CrossRef]
- Schlottau, K.; Rissmann, M.; Graaf, A.; Schön, J.; Sehl, J.; Wylezich, C.; Höper, D.; Mettenleiter, T.C.; Balkema-Buschmann, A.; Harder, T.; et al. SARS-CoV-2 in fruit bats, ferrets, pigs, and chickens: An experimental transmission study. Lancet Microbe 2020, 1, e218–e225. [Google Scholar] [CrossRef]
- Bashor, L.; Gagne, R.B.; Bosco-Lauth, A.M.; Bowen, R.A.; Stenglein, M.; VandeWoude, S. SARS-CoV-2 evolution in animals suggests mechanisms for rapid variant selection. Proc. Natl. Acad. Sci. USA 2021, 118, e2105253118. [Google Scholar] [CrossRef] [PubMed]
- Braun, K.M.; Moreno, G.K.; Halfmann, P.J.; Hodcroft, E.B.; Baker, D.A.; Boehm, E.C.; Weiler, A.M.; Haj, A.K.; Hatta, M.; Chiba, S.; et al. Transmission of SARS-CoV-2 in domestic cats imposes a narrow bottleneck. PLoS Pathog. 2021, 17, e1009373. [Google Scholar] [CrossRef]
- Al Bayat, S.; Mundodan, J.; Hasnain, S.; Sallam, M.; Khogali, H.; Ali, D.; Alateeg, S.; Osama, M.; Elberdiny, A.; Al-Romaihi, H.; et al. Can the cycle threshold (Ct) value of RT-PCR test for SARS CoV2 predict infectivity among close contacts? J. Infect. Public Health 2021, 14, 1201–1205. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.; Zhao, S.; He, D.; Yang, L.; Wang, M.H.; Li, Y.; Mei, S.; Zou, X. Positive RT-PCR tests among discharged COVID-19 patients in Shenzhen, China. Infect. Control Hosp. Epidemiol 2020, 41, 1110–1112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- AlJishi, J.M.; Al-Tawfiq, J.A. Intermittent viral shedding in respiratory samples of patients with SARS-CoV-2: Observational analysis with infection control implications. J. Hosp. Infect. 2021, 107, 98–100. [Google Scholar] [CrossRef] [PubMed]
- Yuan, J.; Kou, S.; Liang, Y.; Zeng, J.; Pan, Y.; Liu, L. PCR Assays Turned Positive in 25 Discharged COVID-19 Patients. Clin. Infect. Dis. 2020, 71, 2230–2232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martins, M.; Boggiatto, P.M.; Buckley, A.; Cassmann, E.D.; Falkenberg, S.; Caserta, L.C.; Fernandes, M.H.V.; Kanipe, C.; Lager, K.; Palmer, M.V.; et al. From Deer-to-Deer: SARS-CoV-2 is efficiently transmitted and presents broad tissue tropism and replication sites in white-tailed deer. PLoS Pathog. 2022, 18, e1010197. [Google Scholar] [CrossRef] [PubMed]
- Lee, P.H.; Tay, W.C.; Sutjipto, S.; Fong, S.W.; Ong, S.W.X.; Wei, W.E.; Chan, Y.H.; Ling, L.M.; Young, B.E.; Toh, M.P.H.; et al. Associations of viral ribonucleic acid (RNA) shedding patterns with clinical illness and immune responses in Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2) infection. Clin. Transl Immunol. 2020, 9, e1160. [Google Scholar] [CrossRef]
- Wong, J.; Koh, W.C.; Momin, R.N.; Alikhan, M.F.; Fadillah, N.; Naing, L. Probable causes and risk factors for positive SARS-CoV-2 test in recovered patients: Evidence from Brunei Darussalam. J. Med. Virol. 2020, 92, 2847–2851. [Google Scholar] [CrossRef]
- Zhou, J.; Otter, J.A.; Price, J.R.; Cimpeanu, C.; Meno Garcia, D.; Kinross, J.; Boshier, P.R.; Mason, S.; Bolt, F.; Holmes, A.H.; et al. Investigating Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2) Surface and Air Contamination in an Acute Healthcare Setting During the Peak of the Coronavirus Disease 2019 (COVID-19) Pandemic in London. Clin. Infect. Dis. 2021, 73, e1870–e1877. [Google Scholar] [CrossRef]
- Hamer, S.A.; Pauvolid-Correa, A.; Zecca, I.B.; Davila, E.; Auckland, L.D.; Roundy, C.M.; Tang, W.; Torchetti, M.K.; Killian, M.L.; Jenkins-Moore, M.; et al. SARS-CoV-2 Infections and Viral Isolations among Serially Tested Cats and Dogs in Households with Infected Owners in Texas, USA. Viruses 2021, 13, 938. [Google Scholar] [CrossRef] [PubMed]
- Rak, A.; Donina, S.; Zabrodskaya, Y.; Rudenko, L.; Isakova-Sivak, I. Cross-Reactivity of SARS-CoV-2 Nucleocapsid-Binding Antibodies and Its Implication for COVID-19 Serology Tests. Viruses 2022, 14, 2041. [Google Scholar] [CrossRef] [PubMed]
- Tan, C.W.; Chia, W.N.; Qin, X.; Liu, P.; Chen, M.I.; Tiu, C.; Hu, Z.; Chen, V.C.; Young, B.E.; Sia, W.R.; et al. A SARS-CoV-2 surrogate virus neutralization test based on antibody-mediated blockage of ACE2-spike protein-protein interaction. Nat. Biotechnol. 2020, 38, 1073–1078. [Google Scholar] [CrossRef] [PubMed]




| Household | Owners | Dogs | Cats | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| ID | Date Entering Study | Canton | People in Household | RT-qPCR Positive 1 | ID | RT-qPCR Positive | Sero-Positive | ID | RT-qPCR Positive | Seropositive |
| USZ22 | 11 November 2021 | ZH | 2 adults, 3 children | 2 adults, 2 children | Dog 22.1 | Yes | Yes 2 | Cat 22.2 | Yes | Yes |
| USZ29 | 21 November 2021 | ZH | 2 adults, 2 children | 1 adult, 2 children | Dog 29.1 | Yes | Yes | Cat 29.2 Cat 29.3 Cat 29.4 | No Yes No | N/A N/A N/A |
| USZ39 | 2 December 2021 | ZH | 2 adults, 2 children | 2 adults, 2 children | Cat 39.1 Cat 39.2 | Yes Yes | Yes Yes | |||
| USZ38 | 4 December 2021 | ZH | 1 adult, 2 children | 2 children | Cat 38.1 Cat 38.2 | No Yes | N/A N/A | |||
| USZ36 | 5 December 2021 | ZH | 2 adults, 2 children | 2 adults | Cat 36.1 | Yes | Yes | |||
| USZ174 | 11 December 2021 | ZH | 2 adults | 2 adults | Cat 174.1 3 Cat 174.2 3 | N/A N/A | Yes Yes | |||
| USZ49 | 19 December 2021 | ZH | 1 adult | 1 adult | Cat 49.1 Cat 49.2 | Yes 4 Yes | Yes 5 Yes | |||
| USZ149 | 21 December 2021 | SG | ≥1 6,7,8 | At least 1 | Dog 149.1 3 | N/A | Yes | |||
| USZ211 | 9 February 2022 | SG | ≥1 6,7 | At least 1 | Cat 211.1 3 | N/A | Yes | |||
| Total | 2/2 | 3/3 | 8/11 | 9/9 | ||||||
| Cat ID | SARS-CoV-2 Positivity | Age (Years) | Sex | Breed | Outdoor Access | Pre-Existing Condition | Clinical Signs | Clinical Signs Observed 1 |
|---|---|---|---|---|---|---|---|---|
| Cat 22.2 | RNA, Abs | unknown | fn | Oriental Shorthair | No | No | ||
| Cat 29.3 | RNA | 5 | f | Europ. Shorthair | Yes | No | ||
| Cat 39.1 Cat 39.2 | RNA, Abs RNA, Abs | 8 13 | mn fn | Unknown Unknown | Yes Yes | Arthrosis | Yes No | Respiratory (nasal discharge) |
| Cat 38.1 Cat 38.2 | No RNA | 1.5 1.5 | mn mn | Bengal Bengal | Giardia infection Giardia infection | Yes Yes | Respiratory and gastrointestinal (ocular discharge, vomiting) Respiratory and gastrointestinal (ocular discharge, vomiting); lethargy, apathy, reduced appetite, bacterial cystitis 2 | |
| Cat 36.1 | RNA | 18 | fn | Mixed breed | Hyper-thyreosis | No | ||
| Cat 49.1 Cat 49.2 | RNA, Abs RNA, Abs | 11 17 | mn mn | Somali Maine Coon | No No | No No | ||
| Cat 174.1 Cat 174.2 | Abs Abs | 9 9 | mn mn | Europ. Shorthair Europ. Shorthair | Yes Yes | No No | ||
| Cat 211.1 | Abs | 10 | f | Europ. Shorthair | No | Yes | Gastrointestinal 2 (vomiting, absent defecation) |
| Dog ID | SARS-CoV-2 Positivity | Age (Years) | Sex | Breed | Pre-Existing Condition | Clinical Signs | Clinical Signs Observed 1 |
|---|---|---|---|---|---|---|---|
| Dog 22.1 | RNA, Abs | unknown | mn | Australian Labradoodle | No | ||
| Dog 29.1 | RNA, Abs | 1 | f | Labrador–Bernese Mountain Mix | Yes | Lethargy/apathy | |
| Dog 149.1 | Abs | 9 | f | Rhodesian Ridgeback | Mammary tumor, pyometra | Yes | Respiratory (sneezing) 2 |
| Animal ID 1 | Date of Blood Collection for Serology | Days after First Positive RT-qPCR of the Animal | RBD Elisa | Percentage of Positive Control | sVNT | Percentage Inhibition |
|---|---|---|---|---|---|---|
| Dog 22.1 | 28 February 2022 | 102 | Negative | Negative | ||
| Cat 22.2 | 28 February 2022 | 100 | Positive | 71 | Positive | 88 |
| Dog 29.1 | 28 December 2021 | 34 | Positive | 33 | Positive | 62 |
| Cat 39.1 | 3 February 2022 | 56 | Positive | 151 | Positive | 100 |
| Cat 39.2 | 3 February 2022 | 56 | Positive | 125 | Positive | 95 |
| Cat 36.1 | 3 January 2022 | 26 | Positive | 130 | Positive | 94 |
| Cat 49.1 | 8 March 2022 | 69 | Positive | 102 | Positive | 100 |
| Cat 49.2 | 8 February 2022 | 41 | Positive | 96 | Negative 2 | 69 |
| Dog 149.1 | 21 December 2022 | N/A 3 | Positive | 55 | Positive | 58 |
| Cat 174.1 | 21 December 2022 | N/A 3 | Positive | 125 | Positive | 100 |
| Cat 174.2 | 21 December 2022 | N/A 3 | Positive | 135 | Positive | 100 |
| Cat 211.1 | 9 February 2022 | N/A 3 | Positive | 147 | Positive | 101 |
| Animal ID | Pseudotype-Based Neutralization Assay (Neutralization Titers 1) | Identified Variant | |||
|---|---|---|---|---|---|
| Alpha | Beta | Delta | Omicron | ||
| Dog 22.1 | <50 | <50 | 58 | <17 | Delta |
| Cat 22.2 | <50 | <50 | 115 | 20 | Delta |
| Dog 29.1 | <50 | 55 | 94 | <17 | Delta |
| Cat 39.1 | 68 | 71 | 166 | 27 | Delta |
| Cat 39.2 | 143 | 101 | 224 | 25 | Delta |
| Cat 36.1 | 49 | <50 | 463 | 22 | Delta |
| Cat 49.1 | 91 | 72 | 229 | <50 | Delta |
| Cat 49.2 | <50 | <50 | 74 | <50 | Delta |
| Dog 149.1 | <50 | <50 | 261 | <17 | Delta |
| Cat 174.1 | 225 | 276 | 550 | 74 | Delta |
| Cat 174.2 | 411 | 707 | 1890 | 379 | Delta |
| Cat 211.1 | 240 | 254 | 341 | 52 | Delta |
| Household ID | Sample from | Date of Sampling for Sequencing | Material of Sequenced Sample | Breadth of Sequencing Coverage | Depth of Sequencing Coverage | SARS-CoV-2 Variant | Pangolin Lineage |
|---|---|---|---|---|---|---|---|
| USZ22 | Owner 1 Owner 2 Owner 3 Owner 4 Dog 22.1 Cat 22.2 | 16 November 2021 16 November 2021 16 November 2021 16 November 2021 18 November 2021 16 November 2021 | Nasal swab Oral swab Nasal swab Nasal swab Oral swab Oral swab | 99.8% 99.8% 99.8% 99.8% 99.8% 99.0% | 10,078.325 7357.6606 5991.9077 8399.461 7603.86 8748.53 | Delta Delta Delta Delta Delta Delta | AY.129 AY.129 AY.129 AY.129 AY.129 AY.129 |
| USZ29 | Owner 1 Dog 29.1 Cat 29.3 | 24 November 2021 24 November 2021 24 November 2021 | Nasal swab Nasal swab Oral swab | 99.8% 99.4% 71.3% | 7085.115 8431.732 16,659.176 | Delta Delta - | AY.4 AY.4 - |
| USZ39 | Owner 1 Owner 2 Owner 3 Owner 4 Cat 39.1 Cat 39.2 | 9 December 2021 9 December 2021 9 December 2021 9 December 2021 9 December 2021 9 December 2021 | Nasal swab Nasal swab Oral swab Oral swab Oral swab Oral swab | 99.7% 99.8% 99.7% 99.9% 99.7% 98.8% | 21,950.77 20,430.922 303.0392 14,888.675 19,432.705 22,502.582 | Delta Delta Delta Delta Delta Delta | B.1.617.2 B.1.617.2 B.1.617.2 B.1.617.2 B.1.617.2 B.1.617.2 |
| USZ38 | Owner 1 Owner 2 Cat 38.2 | 14 December 2021 14 December 2021 14 December 2021 | Nasal swab Nasal swab Nasal swab | 98.8% 90.3% 98.7% | 15,422.453 13,471.375 16,428.799 | Delta - Delta | AY.4 - AY.4 |
| USZ36 | Owner 1 Owner 2 Cat 36.1 | 8 December 2021 8 December 2021 8 December 2021 | Nasal swab Nasal swab Oral swab | 99.8% 99.8% 99.1% | 18,054.1 17,920.172 18,757.414 | Delta Delta Delta | AY.43 AY.43 AY.43 |
| Household ID | Sample from | Minimum Distance (SNPs) to Sequence from Animal in the Household | Minimum Distance (SNPs) to Sequence from Owner in the Household | Most Similar Sequence from Owner in the Household | Minimum Distance (SNPs) to Sequence from Animal in Another Household | Most Similar Sequence from Animal in Another Household | Minimum Distance (SNPs) to Another Swiss Sequence |
|---|---|---|---|---|---|---|---|
| USZ22 | Dog 22.1 Cat 22.2 | 1 1 | 0 1 | Owners 1, 2, 4 Owners 1, 2, 4 | 13 14 | Cat 38.2 Cat 38.2 | 0 1 |
| USZ29 | Dog 29.1 | - | 0 | Owner 1 | 15 | Dog 22.1 | 0 |
| USZ39 | Cat 39.1 Cat 39.2 | 2 2 | 0 1 | Owner 1 Owners 3, 4 | 18 17 | Dog 22.1 and cat 22.2 Dog 22.1 and cat 22.2 | 1 1 |
| USZ38 | Cat 38.2 | - | 3 | Owner 1 | 13 | Dog 22.1 | 2 |
| USZ36 | Cat 36.1 | - | 0 | Owners 1, 2 | 16 | Dog 22.1 and cat 22.2 | 0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kuhlmeier, E.; Chan, T.; Agüí, C.V.; Willi, B.; Wolfensberger, A.; Beisel, C.; Topolsky, I.; Beerenwinkel, N.; Stadler, T.; Swiss SARS-CoV-2 Sequencing Consortium; et al. Detection and Molecular Characterization of the SARS-CoV-2 Delta Variant and the Specific Immune Response in Companion Animals in Switzerland. Viruses 2023, 15, 245. https://doi.org/10.3390/v15010245
Kuhlmeier E, Chan T, Agüí CV, Willi B, Wolfensberger A, Beisel C, Topolsky I, Beerenwinkel N, Stadler T, Swiss SARS-CoV-2 Sequencing Consortium, et al. Detection and Molecular Characterization of the SARS-CoV-2 Delta Variant and the Specific Immune Response in Companion Animals in Switzerland. Viruses. 2023; 15(1):245. https://doi.org/10.3390/v15010245
Chicago/Turabian StyleKuhlmeier, Evelyn, Tatjana Chan, Cecilia Valenzuela Agüí, Barbara Willi, Aline Wolfensberger, Christian Beisel, Ivan Topolsky, Niko Beerenwinkel, Tanja Stadler, Swiss SARS-CoV-2 Sequencing Consortium, and et al. 2023. "Detection and Molecular Characterization of the SARS-CoV-2 Delta Variant and the Specific Immune Response in Companion Animals in Switzerland" Viruses 15, no. 1: 245. https://doi.org/10.3390/v15010245