

Back to Where It Was First Described: Vectors of Sylvatic Yellow Fever Transmission in the 2017 Outbreak in Espírito Santo, Brazil
 , , , , , ,
, , , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Entomological Surveys
2.2. Taxonomic Characterization
2.3. YFV Detection
2.4. Statistical Analysis
3. Results
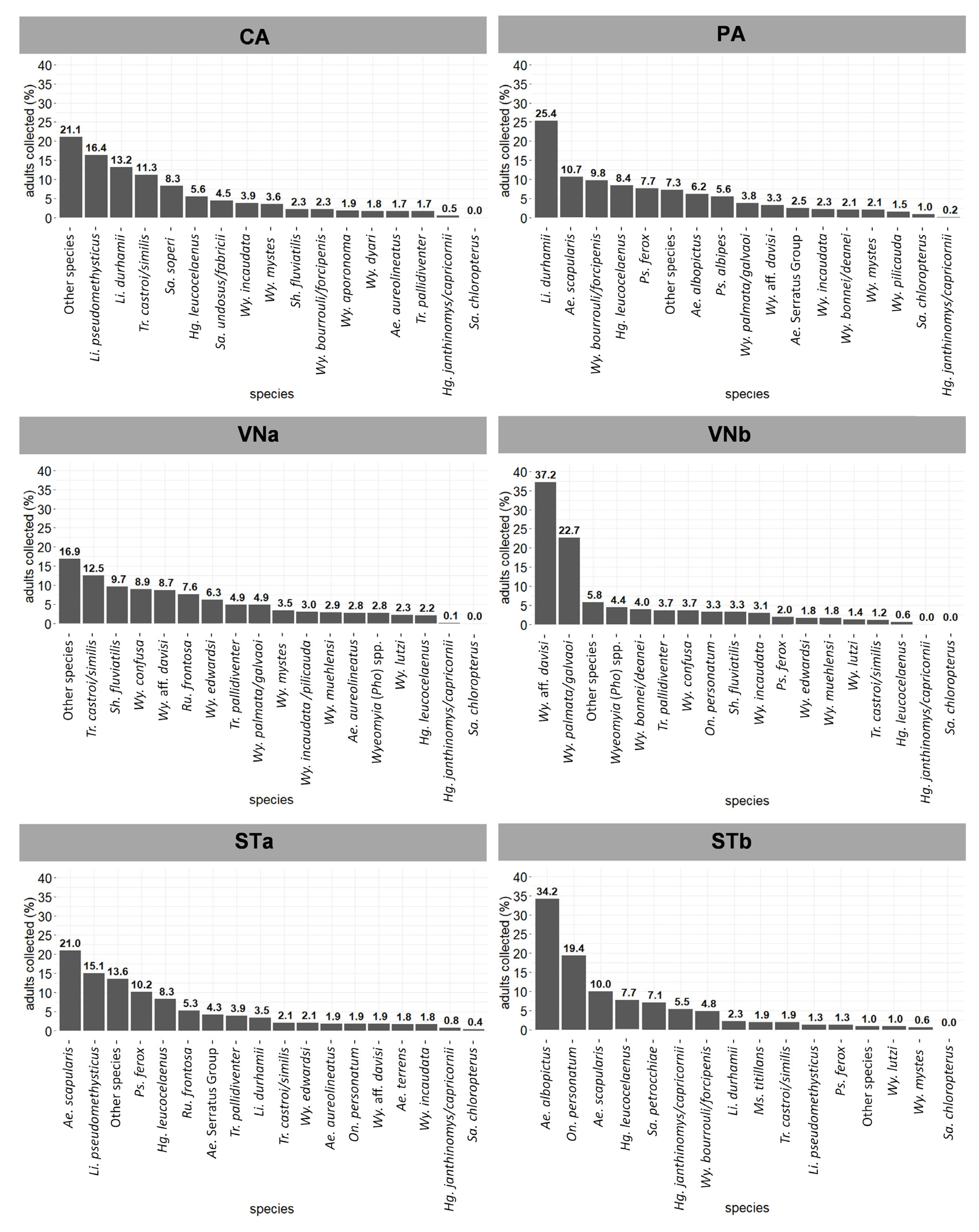
3.1. Entomological Survey Results
3.2. Detection of Natural Infection by YFV
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Monath, T.P.; Vasconcelos, P.F.C. Yellow Fever. J. Clin. Virol. 2015, 64, 160–173. [Google Scholar] [CrossRef] [PubMed]
- Possas, C.; Lourenço-de-Oliveira, R.; Tauil, P.L.; Pinheiro, F.D.P.; Pissinatti, A.; Cunha, R.V.; da Freire, M.; Martins, R.M.; Homma, A. Yellow Fever Outbreak in Brazil: The Puzzle of Rapid Viral Spread and Challenges for Immunisation. Mem. Inst. Oswaldo Cruz 2018, 113, e180278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franco, O. História da Febre Amarela no Brasil; Ministério da Sáude, Departamento Nacional de Endemias Rurais: Rio de Janeiro, Brazil, 1969; Volume 1. [Google Scholar]
- Abreu, F.V.S.; Ferreira-De-Brito, A.; de Souza Azevedo, A.; Linhares, J.H.R.; de Oliveira Santos, V.; Miranda, E.H.; Neves, M.S.A.S.; Yousfi, L.; Ribeiro, I.P.; dos Santos, A.A.C.; et al. Survey on Non-Human Primates and Mosquitoes Does Not Provide Evidences of Spillover/Spillback between the Urban and Sylvatic Cycles of Yellow Fever and Zika Viruses Following Severe Outbreaks in Southeast Brazil. Viruses 2020, 12, 364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vasconcelos, P.F.C.; Rosa, A.P.A.T.; Rodrigues, S.G.; Rosa, E.S.T.; Monteiro, H.A.O.; Cruz, A.C.R.; Barros, V.L.R.S.; Souza, M.R.; Rosa, J.F.S.T. Yellow Fever in Pará State, Amazon Region of Brazil, 1998–1999: Entomologic and Epidemiologic Findings. Emerg. Infect. Dis. 2001, 7, 565–569. [Google Scholar] [PubMed]
- Cardoso, J.D.C.; de Almeida, M.A.B.; dos Santos, E.; da Fonseca, D.F.; Sallum, M.A.M.; Noll, C.A.; Monteiro, H.A.d.O.; Cruz, A.C.R.; Carvalho, V.L.; Pinto, E.v.; et al. Yellow Fever Virus in Haemagogus Leucocelaenus and Aedes Serratus Mosquitoes, Southern Brazil, 2008. Emerg. Infect. Dis. 2010, 16, 1918–1924. [Google Scholar] [CrossRef]
- de Abreu, F.V.S.; Ribeiro, I.P.; Ferreira-de-Brito, A.; dos Santos, A.A.C.; de Miranda, R.M.; Bonelly, I.D.S.; Neves, M.S.A.S.; Bersot, M.I.; dos Santos, T.P.; Gomes, M.Q.; et al. Haemagogus Leucocelaenus and Haemagogus Janthinomys Are the Primary Vectors in the Major Yellow Fever Outbreak in Brazil, 2016–2018. Emerg. Microbes. Infect. 2019, 8, 218–231. [Google Scholar] [CrossRef] [Green Version]
- Soper, F.L.; Penna, H.; Cardoso, E.; Serafim, J.; Frobisher, M.; Pinheiro, J. Yellow Fever without Aedes Aegypti. Study of a Rural Epidemic in the Valle Do Chanaan, Espirito Santo, Brazil, 1932. Am. J. Epidemiol. 1933, 18, 555–587. [Google Scholar] [CrossRef]
- Fernandes, N.C.C.d.A.; Cunha, M.S.; Guerra, J.M.; Réssio, R.A.; Cirqueira, C.D.S.; D’Andretta Iglezias, S.; de Carvalho, J.; Araujo, E.L.L.; Catão-Dias, J.L.; Díaz-Delgado, J. Outbreak of Yellow Fever among Nonhuman Primates, Espirito Santo, Brazil, 2017. Emerg. Infect. Dis. 2017, 23, 2038–2041. [Google Scholar] [CrossRef] [Green Version]
- Cunha, M.S.; da Costa, A.C.; de Azevedo Fernandes, N.C.C.; Guerra, J.M.; dos Santos, F.C.P.; Nogueira, J.S.; D’Agostino, L.G.; Komninakis, S.V.; Witkin, S.S.; Ressio, R.A.; et al. Epizootics Due to Yellow Fever Virus in São Paulo State, Brazil: Viral Dissemination to New Areas (2016–2017). Sci. Rep. 2019, 9, 5474. [Google Scholar] [CrossRef] [Green Version]
- Delatorre, E.; Santos De Abreu, F.V.; Ribeiro, I.P.; Gómez, M.M.; Cunha Dos Santos, A.A.; Ferreira-De-Brito, A.; Alberto Santos Neves, M.S.; Bonelly, I.; de Miranda, R.M.; Furtado, N.D.; et al. Distinct YFV Lineages Co-Circulated in the Central-Western and Southeastern Brazilian Regions from 2015 to 2018. Front. Microbiol. 2019, 10, 1079. [Google Scholar] [CrossRef]
- Ministério da Saúde. Informe 43—Febre Amarela/2017; Ministério da Saúde, Secretaria de Vigilância em Saúde, Centro de Operações de Emergências em Saúde sobre Febre Amarela: Cuiabá, Brazil, 2017. [Google Scholar]
- Ministério da Saúde. Informe 21—Monitoramento Do Período Sazonal Da Febre Amarela Brasil-2017/2018; Monitoramento Da Situação Epidemiológica Da Febre Amarela No Brasil: Cuiabá, Brasil, 2018. [Google Scholar]
- De Rezende, I.M.; Sacchetto, L.; Munhoz de Mello, É.; Alves, P.A.; Iani, F.C.d.M.; Adelino, T.É.R.; Duarte, M.M.; Cury, A.L.F.; Bernardes, A.F.L.; Santos, T.A.; et al. Persistence of Yellow Fever Virus Outside the Amazon Basin, Causing Epidemics in Southeast Brazil, from 2016 to 2018. PLoS Negl. Trop. Dis. 2018, 12, e0006538. [Google Scholar] [CrossRef] [Green Version]
- Siqueira, P.C.; Maciel, E.L.N.; Catão, R.D.C.; Brioschi, A.P.; da Silva, T.C.C.; do Prado, T.N. Completude Das Fichas de Notificação de Febre Amarela No Estado Do Espírito Santo, 2017. Epidemiol. Serv. Saude 2020, 29, e2019402. [Google Scholar] [CrossRef]
- Siqueira, P.C.; Catão, R.C.; Gava, C.; Maciel, E.L.N.; Prado, T.N. Difusão Espaço-Temporal Da Febre Amarela Silvestre Em Um Estado Do Sudeste Do Brasil, 2017. Cad. Saúde Pública 2021, 37, 1–13. [Google Scholar] [CrossRef]
- INCAPER—Instituto Capixaba de Pesquisa Assistência Técnica e Extensão Rural. Programa de Assistência Técnica e Extensão Rural, PROATER 2020-2023; INCAPER: Vitoria, Brazil, 2020. [Google Scholar]
- Moussallem, T.M.; Gava, C.; Ardisson, K.S.; Marques, C.S.; Graceli, G.C.; Valadares Koski, A.d.P.; Almada, G.L.; da Silva, A.R.; de Jesus, F.A.A.; Rodrigues, G.A.P.; et al. Yellow Fever Outbreak in a Rural-Urban Mixed Community of Espírito Santo, Brazil: Epidemiological Aspects. Rev. Panam. Salud Pública 2019, 43, 1. [Google Scholar] [CrossRef] [Green Version]
- Consoli, R.A.G.B.; Lourenço-de-Oliveira, R. Principais Mosquitos de Importância Sanitária No Brasil, 1st ed.; Editora Fiocruz: Rio de Janeiro, Brazil, 1994; ISBN 85-85676-03-5. [Google Scholar]
- Couto-Lima, D.; Andreazzi, C.S.; Leite, P.J.; Bersot, M.I.L.; Alencar, J.; Lourenço-De-oliveira, R. Seasonal Population Dynamics of the Primary Yellow Fever Vector Haemagogus Leucocelaenus (Dyar & Shannon) (Diptera: Culicidae) Is Mainly Influenced by Temperature in the Atlantic Forest, Southeast Brazil. Mem. Inst. Oswaldo Cruz 2020, 115, 1–13. [Google Scholar] [CrossRef]
- Service, M.W. Mosquito Ecology; Springer: Dordrecht, The Netherlands, 1993. [Google Scholar]
- Fay, R.W.; Eliason, D.A. A Preferred Oviposition Site as a Surveillance Method for Aedes Aegypti. Mosq. News 1966, 26, 531–535. [Google Scholar]
- Lane, J. Neotropical Culicidae; Editora Universidade de São Paulo: São Paulo, Brazil, 1953; Volume 2. [Google Scholar]
- Correa, R.R.; Ramalho, G.R. Revisão de Phoniomyia Theobald, 1903 (Diptera, Culicidae, Sabethini). Folia Clin. Biol. 1956, 25, 1–176. [Google Scholar]
- Forattini, O.P. Culicidologia Médica Identificação, Biologia, Epidemiologia; Editora Universidade de São Paulo: São Paulo, Brazil, 2002; Volume 2. [Google Scholar]
- Sallum, M.A.M.; Obando, R.G.; Carrejo, N.; Wilkerson, R.C. Identification Keys to the Anopheles Mosquitoes of South America (Diptera: Culicidae). IV. Adult Females. Parasites Vectors 2020, 13, 584. [Google Scholar] [CrossRef]
- R Core Team R: A Language and Environment for Statistical. Available online: https://www.R-project.org/ (accessed on 24 May 2020).
- Shannon, C.E.; Weaver, W. A Mathematical Theory of Communication; University of Illinois Press: Urbana, IL, USA, 1949; ISBN 13-978-0252725487. [Google Scholar]
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis, 1st ed.; Springer: New York, NY, USA, 2009. [Google Scholar]
- QGIS Team Software QGIS 3.12.2. Available online: https://www.qgis.org/pt_BR/site/ (accessed on 24 May 2020).
- Belkin, J.N.; Heinemann, S.J.; Page, W.A. Mosquito Studies (Diptera: Culicidae) XXI. The Culicidae of Jamaica. Am. Entomol. Inst. Contrib. 1970, 6, 1–458. [Google Scholar]
- Pinheiro, G.G.; Rocha, M.N.; de Oliveira, M.A.; Moreira, L.A.; Filho, J.D.A. Detection of Yellow Fever Virus in Sylvatic Mosquitoes during Disease Outbreaks of 2017–2018 in Minas Gerais State, Brazil. Insects 2019, 10, 136. [Google Scholar] [CrossRef] [Green Version]
- Câmara, D.C.P.; Pinel, C.d.S.; Rocha, G.P.; Codeço, C.T.; Honório, N.A. Diversity of Mosquito (Diptera: Culicidae) Vectors in a Heterogeneous Landscape Endemic for Arboviruses. Acta Trop. 2020, 212, 105715. [Google Scholar] [CrossRef] [PubMed]
- Cunha, M.S.; Tubaki, R.M.; de Menezes, R.M.T.; Pereira, M.; Caleiro, G.S.; Coelho, E.; Saad, L.d.C.; Fernandes, N.C.C.d.A.; Guerra, J.M.; Nogueira, J.S.; et al. Possible Non-Sylvatic Transmission of Yellow Fever between Non-Human Primates in São Paulo City, Brazil, 2017–2018. Sci. Rep. 2020, 10, 15751. [Google Scholar] [CrossRef] [PubMed]
- Das Virgens, T.M.; Rezende, H.R.; Pinto, I.S.; Falqueto, A. Fauna of Mosquitoes (Diptera: Culicidae) in Goytacazes National Forest and Surrounding Area, State of Espírito Santo, Southeastern Brazil. Biota Neotrop. 2018, 18, 1–6. [Google Scholar] [CrossRef]
- Cunha, M.S.; Faria, N.R.; Caleiro, G.S.; Candido, D.S.; Hill, S.C.; Claro, I.M.; da Costa, A.C.; Nogueira, J.S.; Maeda, A.Y.; da Silva, F.G.; et al. Genomic Evidence of Yellow Fever Virus in Aedes Scapularis, Southeastern Brazil, 2016. Acta Trop. 2020, 205, 105390. [Google Scholar] [CrossRef] [PubMed]
- Shannon, R.; Whitman, L.; Franca, M. Yellow Fever Virus in Jungle Mosquitoes. Science 1938, 88, 110–111. [Google Scholar] [CrossRef]
- Wilk-da-Silva, R.; Mucci, L.F.; Ceretti-Junior, W.; Duarte, A.M.R.d.; Marrelli, M.T.; Medeiros-Sousa, A.R. Influence of Landscape Composition and Configuration on the Richness and Abundance of Potential Sylvatic Yellow Fever Vectors in a Remnant of Atlantic Forest in the City of São Paulo, Brazil. Acta Trop. 2020, 204, 105385. [Google Scholar] [CrossRef]
- De Abreu, F.V.S.; de Andreazzi, C.S.; Neves, M.S.A.S.; Meneguete, P.S.; Ribeiro, M.S.; Dias, C.M.G.; de Albuquerque Motta, M.; Barcellos, C.; Romão, A.R.; Magalhães, M.d.A.F.M.; et al. Ecological and Environmental Factors Affecting Transmission of Sylvatic Yellow Fever in the 2017–2019 Outbreak in the Atlantic Forest, Brazil. Parasites Vectors 2022, 15, 23. [Google Scholar] [CrossRef]
- CMN. Coleção de Mosquitos Neotropicais. Available online: http://cmn.fiocruz.br/catalogue (accessed on 27 October 2022).
- Carpenter, S.J.; Galindo, P. Trapido Harold Forest Mosquito Studies in an Endemic YF Area in Panama. Mosq. News 1952, 12, 156–164. [Google Scholar]
- Alencar, J.; de Mello, C.F.; Gil-Santana, H.R.; Guimarães, A.É.; de Almeida, S.A.S.; Gleiser, R.M. Vertical Oviposition Activity of Mosquitoes in the Atlantic Forest of Brazil with Emphasis on the Sylvan Vector, Haemagogus Leucocelaenus (Diptera: Culicidae). J. Vector Ecol. 2016, 41, 18–26. [Google Scholar] [CrossRef] [Green Version]
- Alencar, J.; Morone, F.; de Mello, C.F.; Dégallier, N.; Lucio, P.S.; da Serra-Freire, N.M.; Guimarães, A.É. Flight Height Preference for Oviposition of Mosquito (Diptera: Culicidae) Vectors of Sylvatic Yellow Fever Virus Near the Hydroelectric Reservoir of Simplício, Minas Gerais, Brazil. J. Med. Entomol. 2013, 50, 791–795. [Google Scholar] [CrossRef] [Green Version]
- Hendy, A.; Hernandez-Acosta, E.; Valério, D.; Mendonça, C.; Costa, E.R.; Júnior, J.T.A.; Assunção, F.P.; Scarpassa, V.M.; Gordo, M.; Fé, N.F.; et al. The Vertical Stratification of Potential Bridge Vectors of Mosquito-Borne Viruses in a Central Amazonian Forest Bordering Manaus, Brazil. Sci. Rep. 2020, 10, 18254. [Google Scholar] [CrossRef]
- Rawlinsl, S.C.; Hull’, B.; Chadee, D.D.; Martinez’, R.; Lemaitre, A.; James’, F.; Webb, L. Sylvatic Yellow Fever Activity in Trinidad, 1988 & 1989. Trans. R. Soc. Trop. Med. Hyg. 1990, 84, 142–143. [Google Scholar]
- Dégallier, N.; Travassos da Rosa, A.P.A.; Vasconcelos, P.F.C.; Travassos da Rosa, E.S.; Rodrigues, S.G.; Sá Filho, G.C.; Travassos da Rosa, J.F.S. New Entomological and Virological Data on the Vectors of Sylvatic Yellow Fever in Brazil. Cienc. Cult. 1992, 44, 136–142. [Google Scholar]
- Vasconcelos, P.F.C.; Rodrigues, S.G.; Degallier, N.; Moraes, M.A.P.; Travassos Da Rosa, J.F.S.; Travassos Da Rosa, E.S.; Mondet, B.; Barros, V.L.R.S.; Travassos Da Rosa, A.P.A. An Epidemic of Sylvatic Yellow Fever in the Southeast Region of Maranhao State, Brazil, 1993–1994: Epidemiologic and Entomologic Findings. Am. J. Trop. Med. Hyg. 1997, 57, 132–137. [Google Scholar] [CrossRef] [Green Version]
- Vasconcelos, P.F.C.; Sperb, F.; Monteiro, H.A.O.; Tortes, M.A.N.; Sousa, M.R.S.; Vasconcelos, H.B.; Mardini, L.B.L.F.; Rodrigues, S.G. Isolations of Yellow Fever Virus from Haemagogus Leucocelaenus in Rio Grande Do Sul State, Brazil. Trans. R. Soc. Trop. Med. Hyg. 2003, 97, 60–62. [Google Scholar] [CrossRef]
- Hervé, J.; Dégallier, N.; da Rosa, A. Instituto Evandro Chagas: 50 Anos de Contribuição Às Ciências Biológicas e à Medicina Tropical; Fund, Servo Saúde Pública: Belém, Brazil, 1986; Volume 1. [Google Scholar]
- Moraes, M.A.P.; Travassos da Rosa, A.P.A.; Pinheiro, F.P. An Epidemic of Yellow Fever in Central Brazil, 1972–1973. Am. J. Trop. Med. Hyg. 1981, 30, 204–211. [Google Scholar] [CrossRef]
- De Abreu, F.V.S.; Delatorre, E.; dos Santos, A.A.C.; Ferreira-De-Brito, A.; de Castro, M.G.; Ribeiro, I.P.; Furtado, N.D.; Vargas, W.P.; Ribeiro, M.S.; Meneguete, P.; et al. Combination of Surveillance Tools Reveals That Yellow Fever Virus Can Remain in the Same Atlantic Forest Area at Least for Three Transmission Seasons. Mem. Inst. Oswaldo Cruz 2019, 114, e190076. [Google Scholar] [CrossRef]
- Taylor, R.M.; da Cunha, J.F. An Epidemiological Study of Jungle Yellow Fever in an Endemic Area in Brazil. Part I. Epidemiology of Human Infections. Am. J. Trop. Med. 1946, 26 (Suppl. S1), 1–21. [Google Scholar]
- Giovanetti, M.; de Mendonça, M.C.L.; Fonseca, V.; Mares-Guia, M.A.; Fabri, A.; Xavier, J.; de Jesus, J.G.; Gräf, T.; dos Santos Rodrigues, C.D.; dos Santos, C.C.; et al. Yellow Fever Virus Reemergence and Spread in Southeast Brazil, 2016–2019. J. Virol. 2019, 94, e01623-19. [Google Scholar] [CrossRef]
- Fioravanti, C. O Alarme Dos Macacos; FAPESP: São Paulo, Brazil, 2018. [Google Scholar]
- Cunha, M.d.P.; Duarte-Neto, A.N.; Pour, S.Z.; Ortiz-Baez, A.S.; Černý, J.; Pereira, B.B.d.S.; Braconi, C.T.; Ho, Y.L.; Perondi, B.; Sztajnbok, J.; et al. Origin of the São Paulo Yellow Fever Epidemic of 2017–2018 Revealed through Molecular Epidemiological Analysis of Fatal Cases. Sci. Rep. 2019, 9, 20418. [Google Scholar] [CrossRef] [Green Version]
- Pan American Helth Organization; World Health Organization. Epidemiological Alert Yellow Fever; WHO: Washington, DC, USA, 2022. [Google Scholar]
- De Almeida, M.A.B.; dos Santos, E.; Cardoso, J.d.C.; da Silva, L.G.; Rabelo, R.M.; Bicca-Marques, J.C. Predicting Yellow Fever Through Species Distribution Modeling of Virus, Vector, and Monkeys. EcoHealth 2019, 16, 95–108. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxon | Adults | Immature | Total | Ab (%) 1 | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| CA | PA | STa | STb | VNa | VNb | CA | VNA | |||
| Subfamily Anophelinae | 2 | 4 | 8 | 14 | 0.1 | |||||
| Tribe Anophelini | 2 | 4 | 8 | 14 | 0.1 | |||||
| Anopheles (Kerteszia) cruzii Dyar & Knab, 1908 | 3 | 4 | 7 | 0.07 | ||||||
| Anopheles (Kerteszia) laneanus? (Corrêa & Cerqueira, 1944) | 2 | 2 | 0.02 | |||||||
| Anopheles (Kerteszia) sp. | 2 | 2 | 0.02 | |||||||
| Anopheles (Nyssorhynchus) triannulatus (Neiva & Pinto, 1922) | 1 | 1 | 0.01 | |||||||
| Anopheles (Stethomyia) nimbus (Theobald, 1902)/thomasi Shannon, 1933/acanthotorynus Komp 1937 | 2 | 2 | 0.02 | |||||||
| Subfamily Culicinae | 2005 | 3170 | 886 | 310 | 2677 | 1081 | 258 | 257 | 10,644 | 99.9 |
| Tribe Aedini | 181 | 749 | 439 | 184 | 181 | 37 | 1771 | 16.6 | ||
| Aedes (Howardina) aureolineatus Berlin, 1969 | 35 | 17 | 1 | 29 | 82 | 0.77 | ||||
| Aedes (Howardina) fulvithorax (Lutz, 1904) | 2 | 2 | 4 | 0.04 | ||||||
| Aedes (Ochlerotatus) scapularis (Rondani, 1848) | 2 | 183 | 187 | 31 | 3 | 406 | 3.81 | |||
| Aedes (Ochlerotatus) serratus (Theobald, 1901)/hastatus Dyar, 1922/oligopistus Dyar, 1918/eucephalaeus Dyar, 1918 | 2 | 52 | 38 | 1 | 2 | 1 | 96 | 0.90 | ||
| Aedes (Ochlerotatus) taeniorhynchus (Wiedemann, 1821) | 1 | 1 | 0.01 | |||||||
| Aedes (Protomacleaya) terrens (Walker, 1856) | 3 | 3 | 16 | 43 | 3 | 68 | 0.64 | |||
| Aedes (Stegomyia) aegypti (Linnaeus,1762) | 1 | 1 | 0.01 | |||||||
| Aedes (Stegomyia) albopictus (Skuse, 1894) | 7 | 95 | 4 | 106 | 1 | 213 | 2.00 | |||
| Aedes sp. | 1 | 1 | 2 | 0.02 | ||||||
| Haemagogus (Haemagogus) janthinomys Dyar, 1921/Hg. capricornii Lutz, 1904 | 10 | 21 | 7 | 17 | 15 | 70 | 0.66 | |||
| Haemagogus (Conopostegus) leucocelaenus (Dyar & Shannon, 1924) | 112 | 171 | 74 | 24 | 86 | 7 | 474 | 4.45 | ||
| Psorophora (Janthinosoma) albigenu (Peryassú, 1908) | 1 | 1 | 0.01 | |||||||
| Psorophora (Janthinosoma) albipes (Theobald, 1907) | 74 | 74 | 0.69 | |||||||
| Psorophora (Janthinosoma) albipes (Theobald, 1907)/albigenu (Peryassú, 1908)/varipes (Coquillett, 1904) | 3 | 3 | 0.03 | |||||||
| Psorophora (Janthinosoma) ferox (von Humboldt, 1819) | 8 | 141 | 91 | 4 | 1 | 22 | 267 | 2.51 | ||
| Psorophora (Janthinosoma) lanei Shannon and Cerqueira, 1943 | 1 | 1 | 2 | 4 | 0.04 | |||||
| Psorophora (Janthinosoma) sp. | 4 | 1 | 5 | 0.05 | ||||||
| Tribe Culicini | 8 | 26 | 20 | 22 | 12 | 72 | 101 | 261 | 2.4 | |
| Culex (Carrollia) sp. | 72 | 101 | 173 | 1.62 | ||||||
| Culex (Culex) Declarator Group | 2 | 2 | 0.02 | |||||||
| Culex (Culex) declarator Dyar and Knab, 1906 | 1 | 1 | 0.01 | |||||||
| Culex (Culex) nigripalpus Theobald, 1901 | 3 | 6 | 9 | 2 | 20 | 0.19 | ||||
| Culex (Culex) sp. | 2 | 6 | 7 | 2 | 17 | 0.16 | ||||
| Culex (Melanoconion) sp. | 9 | 7 | 16 | 0.15 | ||||||
| Culex (Microculex) neglectus Lutz, 1904 | 1 | 1 | 2 | 0.02 | ||||||
| Culex (Microculex) imitator Theobald, 1903 | 1 | 1 | 0.01 | |||||||
| Culex (Microculex) sp. | 2 | 2 | 0.02 | |||||||
| Culex sp. | 3 | 11 | 9 | 3 | 1 | 27 | 0.25 | |||
| Tribe Mansoninii | 1 | 6 | 1 | 6 | 14 | 0.1 | ||||
| Coquillettidia (Rhynchotaenia) albicosta (Peryassú, 1908) | 1 | 1 | 0.01 | |||||||
| Coquillettidia (Rhynchotaenia) chrysonotum (Peryassú, 1922) | 1 | 1 | 0.01 | |||||||
| Mansonia (Mansonia) titillans (Walker, 1848) | 5 | 6 | 11 | 0.10 | ||||||
| Mansonia (Mansonia) sp. | 1 | 1 | 0.01 | |||||||
| Tribe Sabethini | 1815 | 2389 | 426 | 120 | 2474 | 1032 | 186 | 156 | 8598 | 80.7 |
| Limatus durhamii Theobald, 1901 | 264 | 646 | 31 | 7 | 14 | 49 | 1011 | 9.49 | ||
| Limatus flavisetosus Oliveira Castro, 1935 | 18 | 1 | 1 | 5 | 15 | 8 | 48 | 0.45 | ||
| Limatus pseudomethysticus (Bonne-Wepster & Bonne, 1920) | 329 | 134 | 4 | 28 | 495 | 4.64 | ||||
| Limatus flavisetosus? Oliveira Castro, 1935 | 1 | 1 | 0.01 | |||||||
| Limatus sp. | 1 | 1 | 0.01 | |||||||
| Onirion personatum (Lutz, 1904) | 34 | 93 | 17 | 60 | 31 | 36 | 7 | 278 | 2.61 | |
| Runchomyia (Runchomyia) cerqueirai (Stone, 1944) | 25 | 1 | 3 | 29 | 0.27 | |||||
| Runchomyia (Runchomyia) frontosa (Theobald, 1903) | 26 | 7 | 47 | 93 | 1 | 174 | 1.63 | |||
| Runchomyia (Runchomyia) humboldti (Lane & Cerqueira, 1942) | 28 | 5 | 3 | 36 | 0.34 | |||||
| Runchomyia (Runchomyia) reversa (Lane & Cerqueira, 1942) | 1 | 1 | 0.01 | |||||||
| Runchomyia (Runchomyia) theobaldi | 2 | 2 | 0.02 | |||||||
| Runchomyia (Runchomyia) sp. | 1 | 1 | 2 | 0.02 | ||||||
| Sabethes (Davismyia) petrocchiae (Shannon & Del Ponte, 1928) | 10 | 22 | 4 | 36 | 0.34 | |||||
| Sabethes (Peytonulus) aurescens (Lutz, 1905) | 21 | 6 | 27 | 0.25 | ||||||
| Sabethes (Peytonulus) fabricii Lane & Cerqueira, 1942 | 19 | 1 | 20 | 0.19 | ||||||
| Sabethes (Peytonulus) undosus (Coquillett, 1906)/fabricii Lane & Cerqueira 1942 | 90 | 3 | 1 | 6 | 100 | 0.94 | ||||
| Sabethes (Peytonulus) hadrognathus Harbach, 1995 | 1 | 1 | 0.01 | |||||||
| Sabethes (Peytonulus) identicus Dyar & Knab, 1907 | 19 | 1 | 3 | 5 | 28 | 0.26 | ||||
| Sabethes (Peytonulus) aff. Ignotus | 7 | 1 | 8 | 0.08 | ||||||
| Sabethes (Peytonulus) soperi Lane & Cerqueira, 1942 | 166 | 1 | 6 | 173 | 1.62 | |||||
| Sabethes (Peytonulus) whitmani Lane and Cerqueira, 1942 | 3 | 3 | 0.03 | |||||||
| Sabethes (Peytonulus) sp. | 1 | 1 | 0.01 | |||||||
| Sabethes (Sabethes) albiprivus Theobald, 1903 | 3 | 7 | 11 | 30 | 51 | 0.48 | ||||
| Sabethes (Sabethes) batesi Lane & Cerqueira, 1942 | 1 | 1 | 2 | 0.02 | ||||||
| Sabethes (Sabethes) forattinii Cerqueira, 1961 | 10 | 10 | 0.09 | |||||||
| Sabethes (Sabethes) purpureus (Theobald, 1907) | 3 | 32 | 3 | 38 | 0.36 | |||||
| Sabethes (Sabethinus) intermedius (Lutz, 1904) | 2 | 1 | 3 | 0.03 | ||||||
| Sabethes (Sabethinus) melanonymphe Dyar, 1924 | 3 | 1 | 4 | 0.04 | ||||||
| Sabethes (Sabethinus) xhyphydes Harbach, 1994 | 1 | 1 | 1 | 3 | 0.03 | |||||
| Sabethes (Sabethoides) chloropterus (von Humboldt, 1819) | 33 | 4 | 37 | 0.35 | ||||||
| Sabethes sp. | 18 | 18 | 0.17 | |||||||
| Shannoniana (Shannoniana) fluviatilis (Theobald, 1903) | 46 | 11 | 114 | 36 | 7 | 120 | 334 | 3.13 | ||
| Trichoprosopon castroi Lane & Cerqueira, 1942/similis Lane & Cerqueira, 1942 | 226 | 19 | 6 | 123 | 13 | 1 | 388 | 3.64 | ||
| Trichoprosopon compressum Lutz, 1905 | 6 | 1 | 2 | 5 | 14 | 0.13 | ||||
| Trichoprosopon digitatum Rondani, 1848 | 6 | 64 | 27 | 97 | 0.91 | |||||
| Trichoprosopon pallidiventer (Lutz,1905) | 35 | 1 | 35 | 52 | 40 | 12 | 175 | 1.64 | ||
| Trichoprosopon soaresi Lane & Cerqueira, 1942 | 6 | 2 | 8 | 0.08 | ||||||
| Trichoprosopon sp. | 1 | 1 | 2 | 0.02 | ||||||
| Wyeomyia (Cruzmyia) dyari Lane & Cerqueira, 1942 | 36 | 4 | 40 | 0.38 | ||||||
| Wyeomyia (Miamyia) codiocampa Dyar & Knab, 1907 | 1 | 1 | 5 | 7 | 0.07 | |||||
| Wyeomyia (Miamyia) limai | 2 | 1 | 3 | 0.03 | ||||||
| Wyeomyia (Miamyia) lutzi (Costa Lima, 1930) | 8 | 1 | 12 | 3 | 48 | 15 | 87 | 0.82 | ||
| Wyeomyia (Miamyia) oblita (Lutz, 1904) | 15 | 15 | 0.14 | |||||||
| Wyeomyia (Miamyia) sabethea Lane & cerqueira, 1942 | 10 | 11 | 5 | 1 | 2 | 4 | 33 | 0.31 | ||
| Wyeomyia (Phoniomyia) antunesi (Lane & Guimarães, 1937) | 6 | 6 | 0.06 | |||||||
| Wyeomyia (Phoniomyia) bonnei (Lane and Cerqueira, 1942)/deanei (Lourenço-de-Oliveira, 1983) | 6 | 75 | 10 | 43 | 134 | 1.26 | ||||
| Wyeomyia (Phoniomyia) aff. davisi | 33 | 212 | 17 | 391 | 402 | 1055 | 9.90 | |||
| Wyeomyia (Phoniomyia) edwardsi (Lane & Cerqueira, 1942) | 8 | 64 | 19 | 306 | 19 | 416 | 3.90 | |||
| Wyeomyia (Phoniomyia) incaudata (Root, 1928) | 78 | 117 | 16 | 301 | 33 | 545 | 5.11 | |||
| Wyeomyia (Phoniomyia) pilicauda Root, 1928 | 20 | 131 | 10 | 10 | 171 | 1.60 | ||||
| Wyeomyia (Phoniomyia) incaudata (Root, 1928)/pilicauda Root, 1928 | 154 | 1 | 155 | 1.45 | ||||||
| Wyeomyia (Phoniomyia) muehlensi Petrocchi, 1927 | 2 | 39 | 24 | 19 | 84 | 0.79 | ||||
| Wyeomyia (Phoniomyia) palmata (Lane & Cerqueira, 1942) | 53 | 53 | 0.50 | |||||||
| Wyeomyia (Phoniomyia) palmata (Lane & Cerqueira, 1942)/galvaoi (Correa & Ramalho, 1956) | 20 | 454 | 12 | 408 | 245 | 1139 | 10.69 | |||
| Wyeomyia (Phoniomyia) theobaldi (Lane & Cerqueira, 1942) | 2 | 3 | 9 | 4 | 18 | 0.17 | ||||
| Wyeomyia (Phoniomyia) sp. | 8 | 34 | 8 | 52 | 48 | 150 | 1.41 | |||
| Wyeomyia (Prosopolepis) confusa (Lutz, 1905) | 1 | 1 | 108 | 40 | 150 | 1.41 | ||||
| Wyeomyia (Spilonympha) bourrouli (Lutz, 1905)/forcipenis Lourenço-de-Oliveira & Silva, 1985 | 46 | 212 | 7 | 15 | 8 | 1 | 289 | 2.71 | ||
| Wyeomyia (Spilonympha) mystes Dyar, 1924 | 73 | 135 | 3 | 2 | 84 | 297 | 2.79 | |||
| Wyeomyia (Triamyia) aporonoma Dyar and Knab 1906 | 38 | 38 | 0.36 | |||||||
| Wyeomyia (Triamyia) aporonoma Dyar and Knab 1906/staminifera Lourenço-de-Oliveira, Motta & Castro,1992 | 12 | 12 | 0.11 | |||||||
| Wyeomyia shannoni Lane & Cerqueira, 1942 | 8 | 8 | 0.08 | |||||||
| Wyeomyia (Wyeomyia) sp. | 1 | 1 | 0.01 | |||||||
| Wyeomyia sp. | 7 | 18 | 1 | 6 | 1 | 33 | 0.31 | |||
| Total | 2007 | 3170 | 890 | 310 | 2685 | 1081 | 258 | 257 | 10,658 | - |
| 10,143 | 515 | |||||||||
| N. Taxa 2 | 57 | 44 | 42 | 17 | 47 | 29 | 11 | 5 | ||
| Adults Sampling 1 | Adults Sampling 2 | |||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CA | PA | STa | STb | VNa | VNb | Total | PA | VNa | Total | |||||||||||
| Sampling Effort 1 (h) | 60 | 42 | 50.5 | 13 | 52 | 33 | 250.5 | 97 | 66 | 163 | ||||||||||
| Species | N 2 | BF 3 | N | BF | N | BF | N | BF | N | BF | N | BF | N | BF | N | BF | N | BF | N | BF |
| Hg. leucocelaenus | 112 | 1.87 | 104 | 2.48 | 74 | 1.47 | 24 | 1.85 | 18 | 0.35 | 7 | 0.21 | 339 | 1.35 | 67 | 0.69 | 68 | 1.03 | 135 | 0.83 |
| Hg. janthinomys/capricornii | 10 | 0.17 | 2 | 0.05 | 7 | 0.14 | 17 | 1.31 | 1 | 0.02 | 37 | 0.15 | 19 | 0.20 | 14 | 0.21 | 33 | 0.20 | ||
| Sa. chloropterus | 12 | 0.29 | 4 | 0.08 | 16 | 0.06 | 21 | 0.22 | 0.00 | 21 | 0.13 | |||||||||
| Ae. aureolineatus | 35 | 0.58 | 17 | 0.34 | 1 | 0.08 | 23 | 0.44 | 76 | 0.30 | 6 | 0.09 | 6 | 0.04 | ||||||
| Sa. identicus | 19 | 0.32 | 2 | 0.04 | 5 | 0.15 | 26 | 0.10 | 1 | 0.01 | 1 | 0.02 | 2 | 0.01 | ||||||
| Sa. soperi | 166 | 2.77 | 1 | 0.02 | 167 | 0.67 | 0 | 0.00 | ||||||||||||
| Sh. fluviatilis | 46 | 0.77 | 11 | 0.22 | 80 | 1.54 | 36 | 1.09 | 173 | 0.69 | 34 | 0.52 | 34 | 0.21 | ||||||
| TOTAL | 388 | 6.47 | 118 | 2.81 | 114 | 2.26 | 42 | 3.23 | 124 | 2.38 | 48 | 1.45 | 834 | 3.33 | 108 | 1.11 | 123 | 1.86 | 231 | 1.42 |
| Adults Sampling 1 | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Local | CA | PA | STa | STb | VNa | VNb | ||||||
| Species | MIR 1 | MLE 2 | MIR | MLE | MIR | MLE | MIR | MLE | MIR | MLE | MIR | MLE |
| Ae. aureolineatus | 0.00 | 0.00 | 71.43 | 117.75 | 0.00 | 0.00 | 0.00 | 0.00 | ||||
| Hg. janthinomys/capricornii | 100.00 | 129.45 | 500.00 | 500.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | ||
| Hg. leucocelaenus | 36.04 | 38.96 | 0.00 | 0.00 | 13.51 | 13.89 | 83.33 | 102.19 | 166.67 | 338.45 | 142.86 | 276.98 |
| Sa. chloropterus | 83.33 | 91.44 | 0.00 | 0.00 | ||||||||
| Sa. identicus | 52.63 | 59.25 | 0.00 | 0.00 | 0.00 | 0.00 | ||||||
| Sa. soperi | 12.35 | 12.66 | 0.00 | 0.00 | ||||||||
| Sh. fluviatilis | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 27.78 | 32.02 | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stanzani, L.M.d.A.; Motta, M.d.A.; Erbisti, R.S.; Abreu, F.V.S.d.; Nascimento-Pereira, A.C.; Ferreira-de-Brito, A.; Neves, M.S.A.S.; Pereira, G.R.; Pereira, G.R.; Santos, C.B.d.; et al. Back to Where It Was First Described: Vectors of Sylvatic Yellow Fever Transmission in the 2017 Outbreak in Espírito Santo, Brazil. Viruses 2022, 14, 2805. https://doi.org/10.3390/v14122805
Stanzani LMdA, Motta MdA, Erbisti RS, Abreu FVSd, Nascimento-Pereira AC, Ferreira-de-Brito A, Neves MSAS, Pereira GR, Pereira GR, Santos CBd, et al. Back to Where It Was First Described: Vectors of Sylvatic Yellow Fever Transmission in the 2017 Outbreak in Espírito Santo, Brazil. Viruses. 2022; 14(12):2805. https://doi.org/10.3390/v14122805
Chicago/Turabian StyleStanzani, Luciana Matos de Abreu, Monique de Albuquerque Motta, Rafael Santos Erbisti, Filipe Vieira Santos de Abreu, Agostinho Cardoso Nascimento-Pereira, Anielly Ferreira-de-Brito, Maycon Sebastião Alberto Santos Neves, Gláucio Rocha Pereira, Glauber Rocha Pereira, Claudiney Biral dos Santos, and et al. 2022. "Back to Where It Was First Described: Vectors of Sylvatic Yellow Fever Transmission in the 2017 Outbreak in Espírito Santo, Brazil" Viruses 14, no. 12: 2805. https://doi.org/10.3390/v14122805