
Restoring Subtropical Forests: Alleviating P Limitation and Introducing C Limitation Using Evergreen Broad-Leaved Tree Species

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area Description
2.2. Experimental Design and Soil Sampling
2.3. Soil Physicochemical Analyses
2.4. Soil Enzyme Activities
2.5. Calculation and Statistical Analysis
3. Results
3.1. Soil Physicochemical Properties
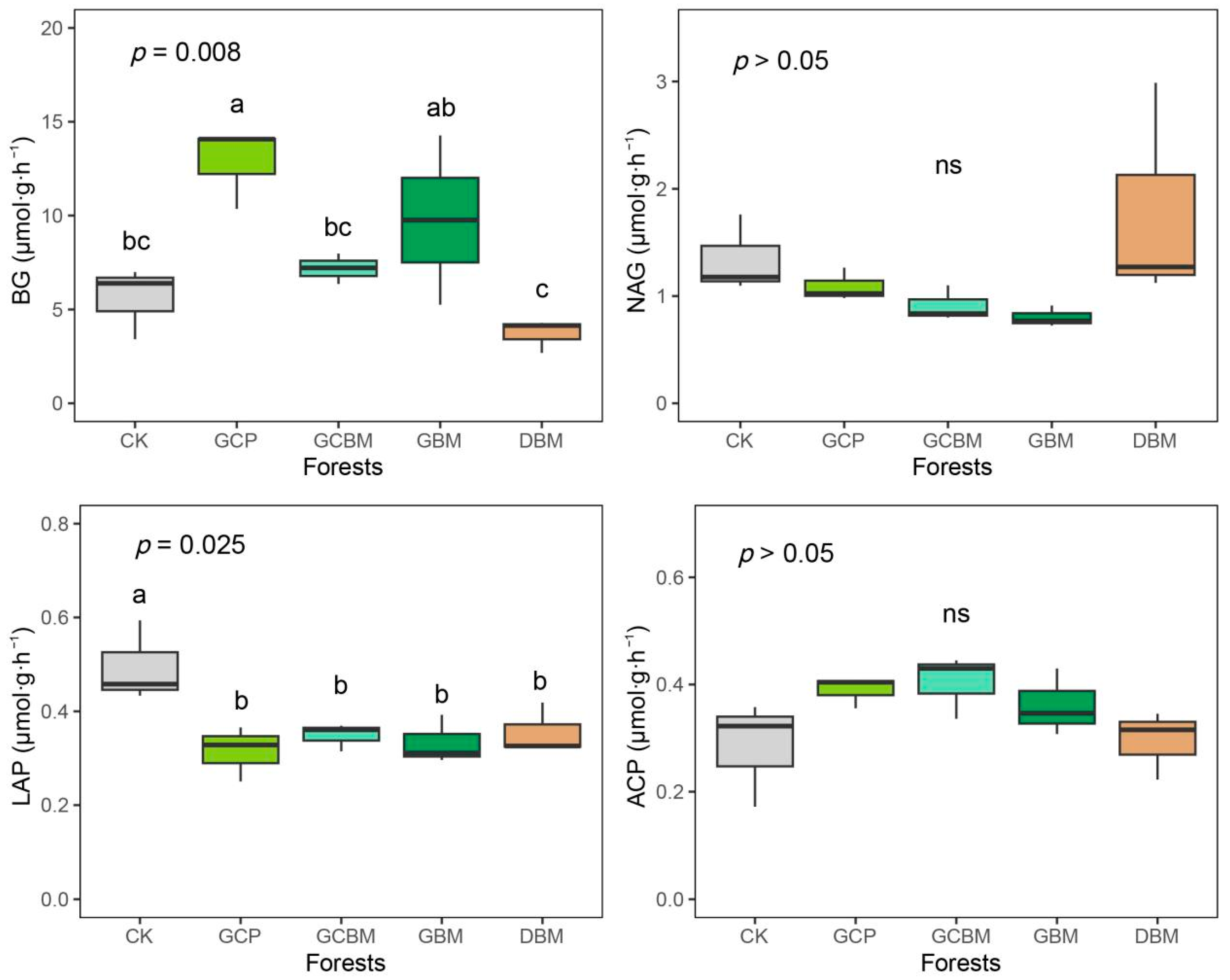
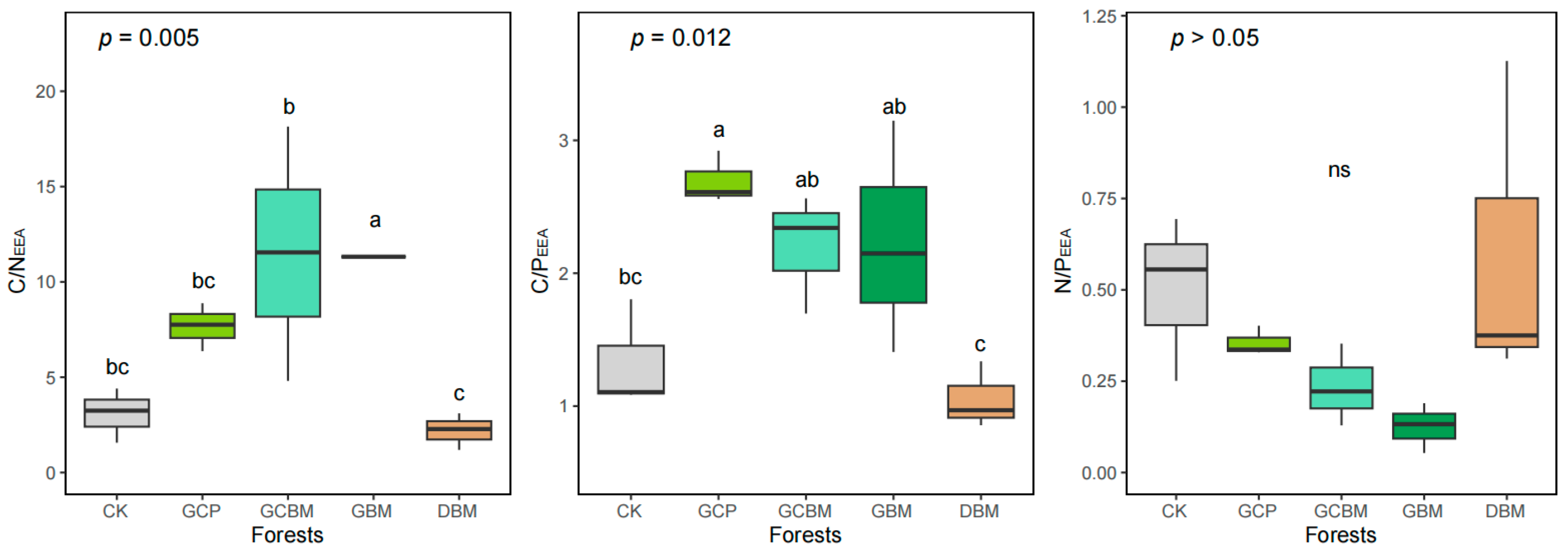
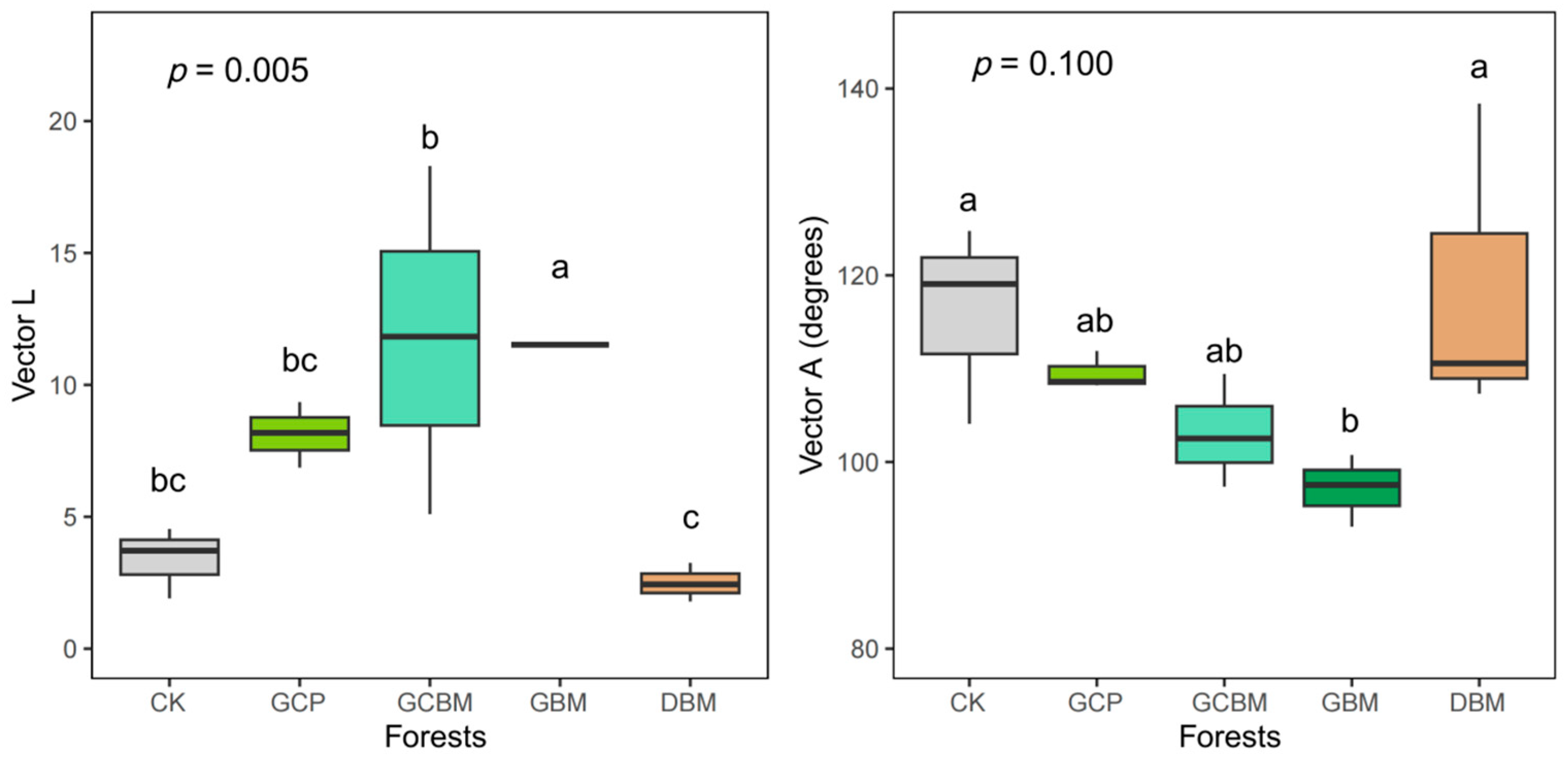
3.2. Soil Enzyme Activities and Their Stoichiometry
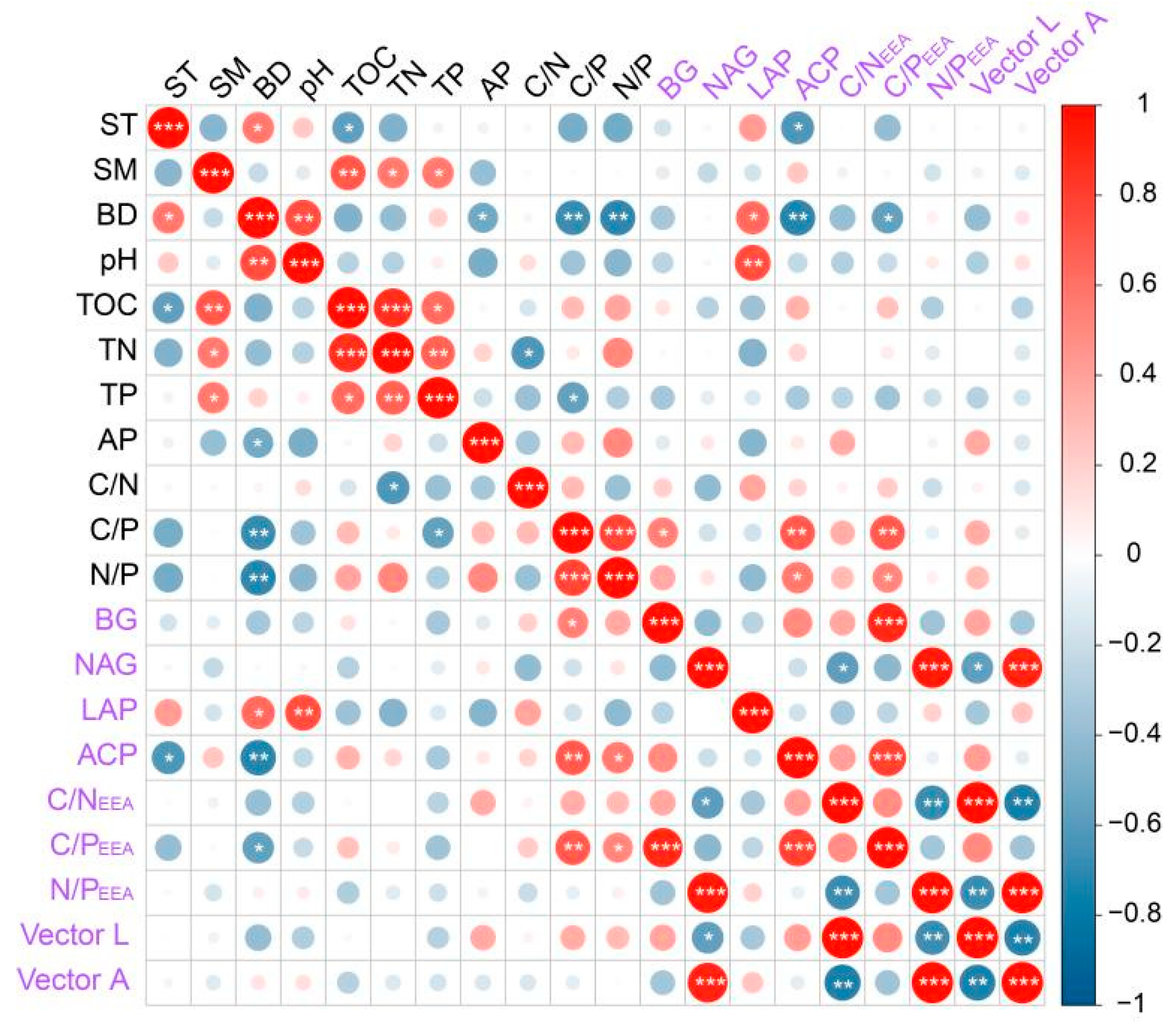
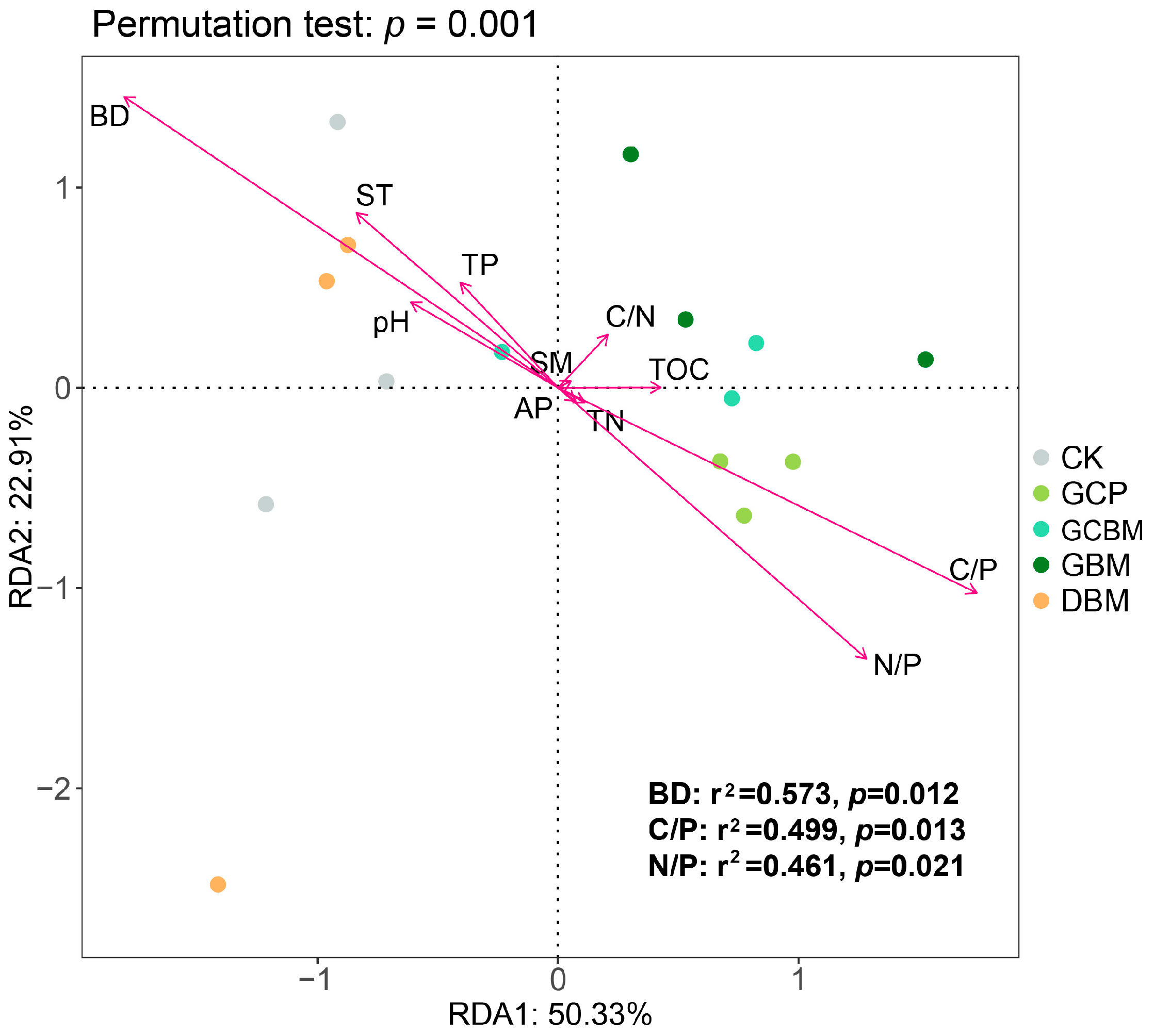
3.3. Relations between Soil Physicochemical Properties and Soil Enzymes
4. Discussion
4.1. Soil Enzyme Activities
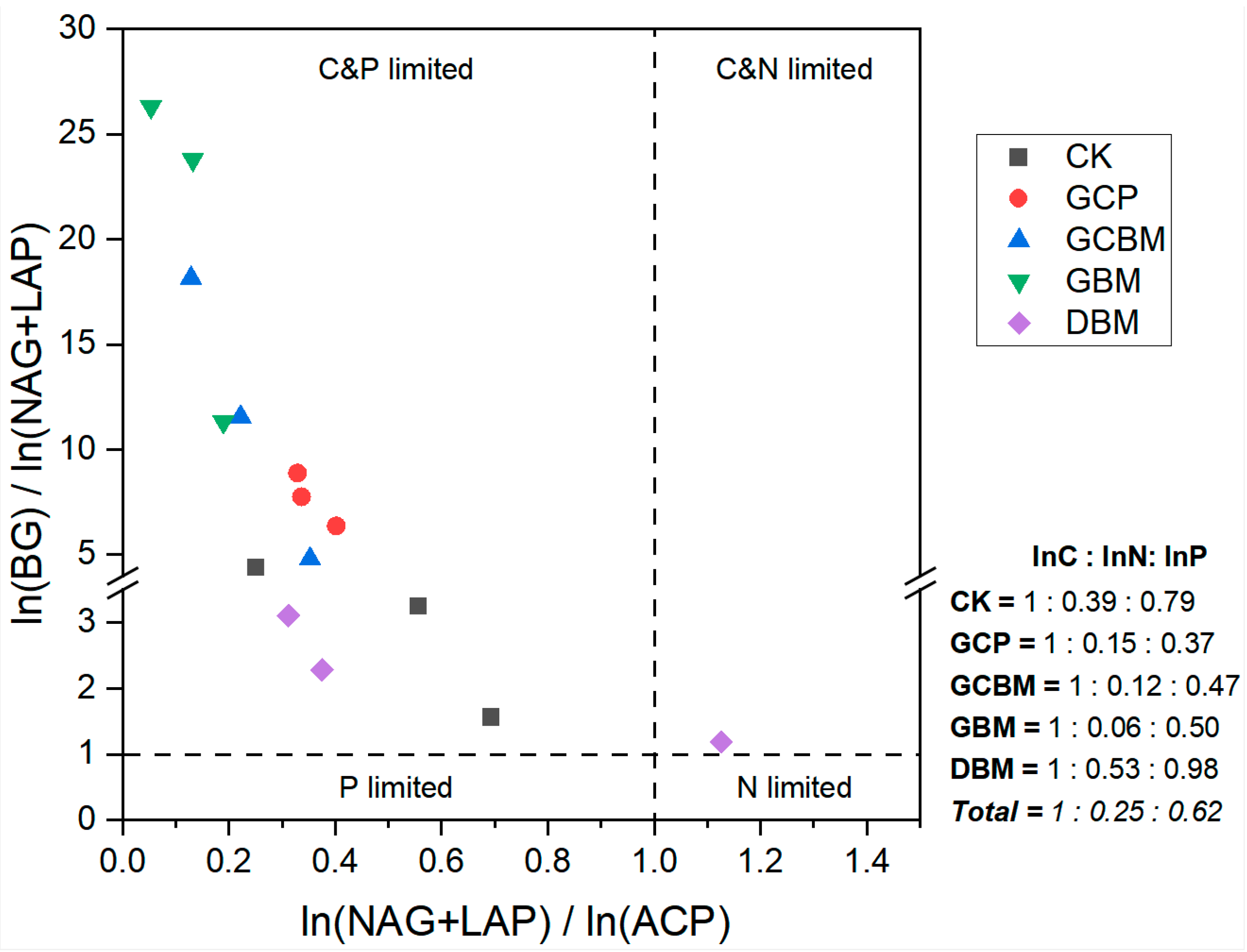
4.2. Nutrient Limitation Status of Soil Microbes
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lozano, Y.M.; Hortal, S.; Armas, C.; Pugnaire, F.I. Interactions among soil, plants, and microorganisms drive secondary succession in a dry environment. Soil Biol. Biochem. 2014, 78, 298–306. [Google Scholar] [CrossRef]
- Li, X.; Liu, Y.; Wu, G.; Lie, Z.; Sheng, H.; Aguila, L.C.R.; Khan, M.S.; Liu, X.; Zhou, S.; Wu, T.; et al. Mixed plantations do not necessarily provide higher ecosystem multifunctionality than monoculture plantations. Sci. Total Environ. 2024, 914, 170156. [Google Scholar] [CrossRef]
- Xu, H.; Yue, C.; Zhang, Y.; Liu, D.; Piao, S. Forestation at the right time with the right species can generate persistent carbon benefits in China. Proc. Natl. Acad. Sci. USA 2023, 120, e2304988120. [Google Scholar] [CrossRef]
- Bahram, M.; Hildebrand, F.; Forslund, S.K.; Anderson, J.L.; Soudzilovskaia, N.A.; Bodegom, P.M.; Bengtsson-Palme, J.; Anslan, S.; Coelho, L.P.; Harend, H.; et al. Structure and function of the global topsoil microbiome. Nature 2018, 560, 233–237. [Google Scholar] [CrossRef]
- Berg, G.; Smalla, K. Plant species and soil type cooperatively shape the structure and function of microbial communities in the rhizosphere. FEMS Microbiol. Ecol. 2009, 68, 1–13. [Google Scholar] [CrossRef]
- Zuccarini, P.; Sardans, J.; Asensio, L.; Peñuelas, J. Altered activities of extracellular soil enzymes by the interacting global environmental changes. Glob. Chang. Biol. 2023, 29, 2067–2091. [Google Scholar] [CrossRef]
- Henry, H.A.L. Soil extracellular enzyme dynamics in a changing climate. Soil Biol. Biochem. 2012, 47, 53–59. [Google Scholar] [CrossRef]
- Sinsabaugh, R.L.; Belnap, J.; Findlay, S.G.; Shah, J.J.F.; Hill, B.H.; Kuehn, K.A.; Kuske, C.R.; Litvak, M.E.; Martinez, N.G.; Moorhead, D.L.; et al. Extracellular enzyme kinetics scale with resource availability. Biogeochemistry 2014, 121, 287–304. [Google Scholar] [CrossRef]
- Sinsabaugh, R.L.; Hill, B.H.; Follstad Shah, J.J. Ecoenzymatic stoichiometry of microbial organic nutrient acquisition in soil and sediment. Nature 2009, 462, 795–798. [Google Scholar] [CrossRef]
- Sinsabaugh, R.L.; Shah, J.J.F. Ecoenzymatic Stoichiometry and Ecological Theory. Annu. Rev. Ecol. Evol. Syst. 2012, 43, 313–343. [Google Scholar] [CrossRef]
- Sinsabaugh, R.L.; Lauber, C.L.; Weintraub, M.N.; Ahmed, B.; Zeglin, L.H. Stoichiometry of soil enzyme activity at global scale. Ecol. Lett. 2008, 11, 1252–1264. [Google Scholar] [CrossRef]
- Burns, R.G.; DeForest, J.L.; Marxsen, J.; Sinsabaugh, R.L.; Stromberger, M.E.; Wallenstein, M.D.; Weintraub, M.N.; Zoppini, A. Soil enzymes in a changing environment: Current knowledge and future directions. Soil Biol. Biochem. 2013, 58, 216–234. [Google Scholar] [CrossRef]
- Sinsabaugh, R.L.; Moorhead, D.L. Resource allocation to extracellular enzyme production: A model for nitrogen and phosphorus control of litter decomposition. Soil Biol. Biochem. 1994, 26, 1305–1311. [Google Scholar] [CrossRef]
- Zhou, L.; Liu, S.; Shen, H.; Zhao, M.; Xu, L.; Xing, A.; Fang, J. Soil extracellular enzyme activity and stoichiometry in China’s forests. Funct. Ecol. 2020, 34, 1461–1471. [Google Scholar] [CrossRef]
- Yuan, X.; Niu, D.; Gherardi, L.A.; Liu, Y.; Wang, Y.; Elser, J.J.; Fu, H. Linkages of stoichiometric imbalances to soil microbial respiration with increasing nitrogen addition: Evidence from a long-term grassland experiment. Soil Biol. Biochem. 2019, 138, 107580. [Google Scholar] [CrossRef]
- Jian, Z.; Ni, Y.; Zeng, L.; Lei, L.; Xu, J.; Xiao, W.; Li, M.-H. Latitudinal patterns of soil extracellular enzyme activities and their controlling factors in Pinus massoniana plantations in subtropical China. For. Ecol. Manag. 2021, 495, 119358. [Google Scholar] [CrossRef]
- Zuo, Y.; Li, J.; Zeng, H.; Wang, W. Vertical pattern and its driving factors in soil extracellular enzyme activity and stoichiometry along mountain grassland belts. Biogeochemistry 2018, 141, 23–39. [Google Scholar] [CrossRef]
- Zheng, H.; Liu, Y.; Zhang, J.; Chen, Y.; Yang, L.; Li, H.; Wang, L. Factors influencing soil enzyme activity in China’s forest ecosystems. Plant Ecol. 2018, 219, 31–44. [Google Scholar] [CrossRef]
- Guo, Z.; Zhang, X.; Green, S.M.; Dungait, J.A.J.; Wen, X.; Quine, T.A. Soil enzyme activity and stoichiometry along a gradient of vegetation restoration at the Karst Critical Zone Observatory in Southwest China. Land Degrad. Dev. 2019, 30, 1916–1927. [Google Scholar] [CrossRef]
- Nwe, T.Z.; Maaroufi, N.I.; Allan, E.; Soliveres, S.; Kempel, A. Plant attributes interact with fungal pathogens and nitrogen addition to drive soil enzymatic activities and their temporal variation. Funct. Ecol. 2023, 37, 564–575. [Google Scholar] [CrossRef]
- Wu, L.; Zou, B.; Wang, S.; Zhou, L.; Zheng, Y.; Huang, Z.; He, J.-Z. Effects of multispecies restoration on soil extracellular enzyme activity stoichiometry in Pinus massoniana plantations of subtropical China. Soil Biol. Biochem. 2023, 178, 108967. [Google Scholar] [CrossRef]
- Xu, Z.W.; Yu, G.R.; Zhang, X.Y.; He, N.P.; Wang, Q.F.; Wang, S.Z.; Wang, R.L.; Zhao, N.; Jia, Y.L.; Wang, C.Y. Soil enzyme activity and stoichiometry in forest ecosystems along the North-South Transect in eastern China (NSTEC). Soil Biol. Biochem. 2017, 104, 152–163. [Google Scholar] [CrossRef]
- Huang, Y.-X.; Wu, Z.-J.; Zong, Y.-Y.; Li, W.-Q.; Chen, F.-S.; Wang, G.G.; Li, J.-J.; Fang, X.-M. Mixing with coniferous tree species alleviates rhizosphere soil phosphorus limitation of broad-leaved trees in subtropical plantations. Soil Biol. Biochem. 2022, 175, 108853. [Google Scholar] [CrossRef]
- Fanin, N.; Fromin, N.; Buatois, B.; Hättenschwiler, S. An experimental test of the hypothesis of non-homeostatic consumer stoichiometry in a plant litter–microbe system. Ecol. Lett. 2013, 16, 764–772. [Google Scholar] [CrossRef]
- Xiao, L.; Liu, G.; Li, P.; Li, Q.; Xue, S. Ecoenzymatic stoichiometry and microbial nutrient limitation during secondary succession of natural grassland on the Loess Plateau, China. Soil Tillage Res. 2020, 200, 104605. [Google Scholar] [CrossRef]
- Liu, G.; Wang, H.; Yan, G.; Wang, M.; Jiang, S.; Wang, X.; Xue, J.; Xu, M.; Xing, Y.; Wang, Q. Soil enzyme activities and microbial nutrient limitation during the secondary succession of boreal forests. Catena 2023, 230, 107268. [Google Scholar] [CrossRef]
- Peng, Z.; Wu, Y.; Guo, L.; Yang, L.; Wang, B.; Wang, X.; Liu, W.; Su, Y.; Wu, J.; Liu, L. Foliar nutrient resorption stoichiometry and microbial phosphatase catalytic efficiency together alleviate the relative phosphorus limitation in forest ecosystems. New Phytol. 2023, 238, 1033–1044. [Google Scholar] [CrossRef]
- Du, E.Z.; Terrer, C.; Pellegrini, A.F.A.; Ahlström, A.; van Lissa, C.J.; Zhao, X.; Xia, N.; Wu, X.H.; Jackson, R.B. Global patterns of terrestrial nitrogen and phosphorus limitation. Nat. Geosci. 2020, 13, 221–226. [Google Scholar] [CrossRef]
- Cai, L.; Yang, Y.; Chong, Y.; Xiong, J.; Wu, J.; Ai, X.; Guo, Q.; Yuan, Y.; Li, Z. Higher soil aggregate stability in subtropical coniferous plantations than natural forests due to microbial and aggregate factors. Forests 2022, 13, 2110. [Google Scholar] [CrossRef]
- Saiya-Cork, K.R.; Sinsabaugh, R.L.; Zak, D.R. The effects of long term nitrogen deposition on extracellular enzyme activity in an Acer saccharum forest soil. Soil Biol. Biochem. 2002, 34, 1309–1315. [Google Scholar] [CrossRef]
- Moorhead, D.L.; Sinsabaugh, R.L.; Hill, B.H.; Weintraub, M.N. Vector analysis of ecoenzyme activities reveal constraints on coupled C, N and P dynamics. Soil Biol. Biochem. 2016, 93, 1–7. [Google Scholar] [CrossRef]
- Liu, R.; Zhang, Y.; Hu, X.-F.; Wan, S.; Wang, H.; Liang, C.; Chen, F.-S. Litter manipulation effects on microbial communities and enzymatic activities vary with soil depth in a subtropical Chinese fir plantation. For. Ecol. Manag. 2021, 480, 118641. [Google Scholar] [CrossRef]
- Šnajdr, J.; Valášková, V.; Merhautová, V.; Herinková, J.; Cajthaml, T.; Baldrian, P. Spatial variability of enzyme activities and microbial biomass in the upper layers of Quercus petraea forest soil. Soil Biol. Biochem. 2008, 40, 2068–2075. [Google Scholar] [CrossRef]
- Pérez-Suárez, M.; Arredondo-Moreno, J.T.; Huber-Sannwald, E.; Vargas-Hernández, J.J. Production and quality of senesced and green litterfall in a pine–oak forest in central-northwest Mexico. For. Ecol. Manag. 2009, 258, 1307–1315. [Google Scholar] [CrossRef]
- Liang, C.; Schimel, J.P.; Jastrow, J.D. The importance of anabolism in microbial control over soil carbon storage. Nat. Microbiol. 2017, 2, 17105. [Google Scholar] [CrossRef]
- Cao, X.W.; Shi, Z.M.; Chen, J.; Liu, S.; Zhang, M.M.; Chen, M.; Xu, G.X.; Wu, J.M.; Xing, H.S.; Li, F.F. Extracellular enzyme characteristics and microbial metabolic limitation in soil of subalpine forest ecosystems on the eastern Qinghai-Tibetan Plateau. Plant Soil 2022, 479, 337–353. [Google Scholar] [CrossRef]
- Yan, B.; Duan, M.; Wang, R.; Li, J.; Wei, F.; Chen, J.; Wang, J.; Wu, Y.; Wang, G. Planted forests intensified soil microbial metabolic nitrogen and phosphorus limitation on the Loess Plateau, China. Catena 2022, 211, 105982. [Google Scholar] [CrossRef]
- Zhou, M.J.; Wang, J.L.; Hu, H.B.; Chen, J.Y.; Zhu, Z.Y.; Heng, Y.C.; Feng, Y.Y. Simulated Nitric Acid Rain Aggravated the C and P Limits of Forest Soil Microorganisms. Forests 2023, 14, 1044. [Google Scholar] [CrossRef]
- Lehmann, J.; Kleber, M. The contentious nature of soil organic matter. Nature 2015, 528, 60–68. [Google Scholar] [CrossRef]
- Cotrufo, M.F.; Ranalli, M.G.; Haddix, M.L.; Six, J.; Lugato, E. Soil carbon storage informed by particulate and mineral-associated organic matter. Nat. Geosci. 2019, 12, 989–994. [Google Scholar] [CrossRef]
- Xiang, H.M.; Luo, X.Z.; Zhang, L.L.; Hou, E.Q.; Li, J.; Zhu, Q.D.; Wen, D.Z. Forest succession accelerates soil carbon accumulation by increasing recalcitrant carbon stock in subtropical forest topsoils. Catena 2022, 212, 106030. [Google Scholar] [CrossRef]
- Vitousek, P.M.; Farrington, H. Nutrient limitation and soil development: Experimental test of a biogeochemical theory. Biogeochemistry 1997, 37, 63–75. [Google Scholar] [CrossRef]
- Chen, Y.; Li, S.; Zeng, L.; An, B.; Xiao, T.; Mao, R.; Zhang, Y. Effects of Mycorrhizal and Extraradical Hyphae of Subtropical Native Tree Species on Soil Enzyme Activities and Their Stoichiometric Ratios. Forests 2023, 14, 2112. [Google Scholar] [CrossRef]
- DeForest, J.L.; Moorhead, D.L. Effects of elevated pH and phosphorus fertilizer on soil C, N and P enzyme stoichiometry in an acidic mixed mesophytic deciduous forest. Soil Biol. Biochem. 2020, 150, 107996. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Forest | Forest Type | Longitude (E) | Latitude (N) | Altitude (m) | Slope (°) | Aspect | Stand Age | Canopy Density |
|---|---|---|---|---|---|---|---|---|
| 1 | CK | 114.4917274 | 30.5067658 | 113.5 | 15 | South | 20 | 0.60 |
| 2 | CK | 114.4916772 | 30.5067728 | 110.1 | 14 | South | 20 | 0.65 |
| 3 | CK | 114.4914149 | 30.5067738 | 117.9 | 12 | South | 20 | 0.80 |
| 4 | GCP | 114.4905969 | 30.5068831 | 121.5 | 29 | South | 20 | 0.55 |
| 5 | GCP | 114.4905451 | 30.5068259 | 115.5 | 27 | South | 20 | 0.50 |
| 6 | GCP | 114.4906427 | 30.5067447 | 122.6 | 36 | South | 20 | 0.60 |
| 7 | GCBM | 114.4891612 | 30.5065437 | 123.7 | 12 | South | 15 | 0.70 |
| 8 | GCBM | 114.489269 | 30.5067259 | 125.4 | 16 | South | 15 | 0.75 |
| 9 | GCBM | 114.4891927 | 30.506788 | 127.4 | 15 | South | 15 | 0.66 |
| 10 | GBM | 114.4965209 | 30.5068437 | 107 | 21 | South | 25 | 0.78 |
| 11 | GBM | 114.4965414 | 30.5066902 | 107.8 | 26 | South | 25 | 0.75 |
| 12 | GBM | 114.4962807 | 30.506741 | 95.8 | 23 | South | 25 | 0.85 |
| 13 | DBM | 114.4990471 | 30.5071404 | 86.3 | 15 | Southeast | 23 | 0.70 |
| 14 | DBM | 114.4993576 | 30.507338 | 65.1 | 16 | Southeast | 23 | 0.75 |
| 15 | DBM | 114.4992954 | 30.5073709 | 72.6 | 16 | Southeast | 23 | 0.68 |
| Enzyme | Abbreviation | EC | Function |
|---|---|---|---|
| β-1,4-glucosidase | BG | 3.2.1.21 | Cellulose degradation: hydrolyzes glucose from cellobiose |
| β-1,4-N-acetylglucosaminidase | NAG | 3.2.1.14 | Chitin and peptidoglycan degradation: hydrolyzes glucosamine from chitobiose |
| Leucine aminopeptidase | LAP | 3.4.11.1 | Proteolysis: hydrolyzes leucine and other hydrophobic amino acids from the N terminus of polypeptides |
| Acid phosphatase | ACP | 3.1.3.1 | Hydrolyzes phosphate from phosphosaccarides and phospholipids |
| Soil Properties | CK | GCP | GCBM | GBM | DBM | p-Value |
|---|---|---|---|---|---|---|
| ST (°C) | 25.61 ± 1.05 a | 23.42 ± 0.29 b | 23.15 ± 0.17 b | 25.44 ± 0.27 a | 24.69 ± 0.08 ab | 0.018 |
| SM (%) | 9.17 ± 1.56 | 10.64 ± 2.39 | 12.73 ± 1.95 | 8.24 ± 0.56 | 10.95 ± 2.19 | >0.05 |
| BD (g/cm3) | 1.20 ± 0.12 a | 0.71 ± 0.06 b | 0.73 ± 0.06 b | 0.75 ± 0.02 b | 0.81 ± 0.09 b | 0.005 |
| pH | 5.79 ± 0.13 a | 4.23 ± 0.10 c | 5.02 ± 0.17 b | 4.28 ± 0.15 c | 4.27 ± 0.02 c | <0.001 |
| TOC (g/kg) | 31.27 ± 0.89 c | 36.85 ± 0.96 ab | 39.28 ± 1.96 a | 32.48 ± 1.05 bc | 37.20 ± 2.15 ab | 0.017 |
| TN (g/kg) | 2.16 ± 0.14 | 2.54 ± 0.12 | 2.76 ± 0.27 | 2.24 ± 0.10 | 2.75 ± 0.06 | >0.05 |
| TP (g/kg) | 0.70 ± 0.05 | 0.69 ± 0.02 | 0.75 ± 0.07 | 0.64 ± 0.03 | 0.79 ± 0.07 | >0.05 |
| AP (mg/kg) | 5.83 ± 0.86 | 8.00 ± 1.84 | 8.70 ± 0.55 | 9.81 ± 0.95 | 9.40 ± 1.17 | >0.05 |
| C/N | 14.62 ± 1.02 | 14.54 ± 0.34 | 14.39 ± 0.76 | 14.54 ± 0.21 | 13.52 ± 0.49 | >0.05 |
| C/P | 45.09 ± 3.66 | 53.53 ± 1.09 | 53.03 ± 2.41 | 50.88 ± 3.63 | 47.51 ± 1.48 | >0.05 |
| N/P | 3.08 ± 0.04 | 3.69 ± 0.15 | 3.71 ± 0.25 | 3.50 ± 0.28 | 3.53 ± 0.24 | >0.05 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, Y.; Xia, W.; Fan, Y.; Chong, Y.; Xiong, J.; Yu, W. Restoring Subtropical Forests: Alleviating P Limitation and Introducing C Limitation Using Evergreen Broad-Leaved Tree Species. Forests 2024, 15, 568. https://doi.org/10.3390/f15030568
Yang Y, Xia W, Fan Y, Chong Y, Xiong J, Yu W. Restoring Subtropical Forests: Alleviating P Limitation and Introducing C Limitation Using Evergreen Broad-Leaved Tree Species. Forests. 2024; 15(3):568. https://doi.org/10.3390/f15030568
Chicago/Turabian StyleYang, Yujing, Wei Xia, Yixuan Fan, Yujie Chong, Jiatai Xiong, and Wenjing Yu. 2024. "Restoring Subtropical Forests: Alleviating P Limitation and Introducing C Limitation Using Evergreen Broad-Leaved Tree Species" Forests 15, no. 3: 568. https://doi.org/10.3390/f15030568